
Abstract
Background
Lung cancer is a foremost global health challenge and remains the leading cause of cancer-related mortality worldwide. Despite advancements in early diagnosis and treatment, most cases are detected at advanced stages, which severely compromises survival rates. Current chemotherapeutic agents are often limited by toxicity and resistance, highlighting the urgent need for safer and more effective preventive strategies.
Purpose
The present study evaluated the chemopreventive potential of ajmalicine, an indole alkaloid, against benzo(a)pyrene (B(a)P)-induced lung carcinogenesis in rats.
Materials and Methods
Lung carcinogenesis was induced by oral administration of B(a)P for 4 consecutive weeks. Ajmalicine (20 mg/kg) was administered orally 1 week before and throughout the 16-week experimental period. Post-treatment, antioxidant status, xenobiotic detoxification enzymes, inflammatory cytokines, and caspase-3/9 activities were measured using standard commercial kits. Histopathological examination of lung tissues was performed to confirm the protective role of ajmalicine.
Results
Ajmalicine treatment significantly enhanced antioxidant markers while reducing xenobiotic detoxification enzyme activity in B(a)P-exposed rats. Furthermore, ajmalicine attenuated pro-inflammatory cytokines and increased caspase-3/9 activity, suggesting the induction of apoptosis. Histopathological findings corroborated its protective role by demonstrating improved lung tissue architecture.
Conclusion
These findings reveal, for the first time, that ajmalicine exhibits strong chemopreventive effects against B(a)P-induced lung carcinogenesis by modulating oxidative stress, inflammation, and apoptosis. Ajmalicine may represent a promising natural candidate for preventing carcinogen-induced lung cancer.
Introduction
Lung cancer is a major health concern worldwide, characterized by unregulated cell division in the lungs, and is the predominant cause of cancer deaths in the world. The prognosis of lung cancer is poor, even with the advancement in diagnostic and therapeutic procedures, and the 5-year survival rate is dismal in most countries. The mortality rate associated with lung cancer can be attributed to late detection, its devastating effects, and the emergence of resistance to the standard treatment (Bray et al., 2024). Lung cancer is successfully treated through early detection, screening, and prevention to reduce the disease mortality. The drawbacks of the existing diagnostic modalities, including invasiveness and cost of pathological diagnosis, emphasize the necessity of less invasive and cheaper diagnostic tools. Understanding of the pathophysiology of tumor development and the biomarkers specific to lung cancer allows the development of effective therapy (Vicidomini, 2023). Lung cancer is a complex process that is triggered by the genetic and epigenetic changes of lung cells. These alterations can interfere with significant cellular pathways that are implicated in cell growth, differentiation, and apoptosis. Individuals who are exposed to environmental carcinogens, including tobacco smoke, asbestos, and radon, also have a high risk of lung cancer (Ravanbakhsh et al., 2023). These carcinogens can damage DNA, resulting in oncogene mutation and tumor suppressor genes. Specifically, benzo(a)pyrene (B(a)P) is a polycyclic aromatic hydrocarbon and a potent carcinogen of the lung, which induces DNA adducts that result in mutations and genomic instability (Stading et al., 2021).
The discovery of genes that cause cancer development has significantly enhanced the molecular knowledge of lung cancer. Molecular-targeted therapy has done a lot to enhance the treatment of lung cancer; however, the resistance that eventually develops reduces the effectiveness of the therapy. New drugs that target drug-resistant mutations have been developed, and they help patients who are intolerant of first-line targeted drugs. Nevertheless, these medications are not fully tested (Li et al., 2022). The tumor microenvironment is a crucial factor in lung cancer progression and metastasis, which affects the response to treatment and resistance. Tumor cell–tumor microenvironment interfaces may stimulate tumor growth and angiogenesis. The immunotherapies have transformed the treatment of lung cancer by enhancing the immune reaction of the host against cancerous cells. However, immunotherapy only works in a small portion of patients, and predictive biomarkers are required to determine the patients who will respond most (Zhou et al., 2024).
The animal models, especially those triggered by B(a)P, have played a critical role in the pathophysiology of lung cancer and in the assessment of new treatment approaches. The lung cancer model caused by B(a)P is a good model to investigate the preclinical research because they are similar to the molecular and histological features of human lung cancer (Yang et al., 2024). The models enable the researchers to determine the impact of carcinogens on the lung tissue, early biomarkers of lung cancer development, and the potential ability of the chemopreventive candidates (Sehgal et al., 2012). Newer forms of treatment are being found to counter the inefficiencies of the existing forms of treatment and to increase the survival of lung cancer patients. The necessity for safer and more effective alternative treatment options of lung cancer proves the inadequacy of the current treatment methods and the increasing importance of the necessity to focus on the complex processes that take place and lead to the onset of lung cancer (Vostrikova et al., 2020). Ajmalicine is a bioactive monoterpenoid indole alkaloid compound of natural origin that is isolated in plants like Rauvolfia serpentina and Catharanthus roseus, and it has been mentioned to show anti-hypertensive activity (Misra et al., 2006). Previous research already stated that ajmalicine possesses neuroprotective (Liu et al., 2020) and antioxidant (Jaleel et al., 2007) effects. In addition, Sun et al. (2024) have indicated that ajmalicine exhibited anti-tumor effects through pyroptosis induction in hepatoma cells.
Previous studies have demonstrated the chemopreventive potential of several plant-derived alkaloids and flavonoids against chemically induced cancers, largely through mechanisms involving antioxidant enhancement, modulation of inflammatory signaling, and regulation of apoptosis. However, investigations into the protective role of ajmalicine, an indole alkaloid primarily known for its anti-hypertensive and neuroprotective effects, in the context of carcinogen-induced lung cancer are extremely limited. To the best of our knowledge, no prior study has systematically evaluated ajmalicine’s ability to counteract B(a)P-induced lung carcinogenesis, a well-established model that closely mimics environmental and tobacco-related carcinogen exposure in humans. The novelty of this research lies in demonstrating, for the first time, that ajmalicine not only enhances antioxidant defense systems but also suppresses xenobiotic enzyme activation, attenuates pro-inflammatory cytokine release, and promotes caspase-mediated apoptosis in B(a)P-exposed rats. Furthermore, the integration of biochemical, molecular, and histopathological evidence provides a comprehensive understanding of ajmalicine’s chemopreventive mechanism. These unique findings broaden the pharmacological relevance of ajmalicine beyond its known therapeutic roles and suggest its potential as a natural candidate for lung cancer chemoprevention. However, there are no reports to claim the anti-cancer properties of ajmalicine against lung cancer. The objective of this study was to investigate the chemopreventive potential of ajmalicine against B(a)P-induced lung carcinogenesis in rats by assessing its effects on oxidative stress, xenobiotic detoxification enzymes, inflammatory cytokines, apoptotic markers, and histopathological alterations. Consequently, the present work evaluated the anti-cancer properties of ajmalicine against B(a)P-induced lung cancer in rats.
Materials and Methods
Chemicals and Reagents
B(a)P, ajmalicine, and other analytical-grade reagents were procured from Sigma–Aldrich (St. Louis, MO, USA) unless otherwise specified. Commercial enzyme-linked immunosorbent assay (ELISA) kits for antioxidant enzymes—superoxide dismutase (SOD), catalase (CAT), and glutathione (GSH)—and the lipid peroxidation marker malondialdehyde (MDA) (Abcam, USA), inflammatory cytokines—tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), interleukin-6 (IL-6), and nuclear factor kappa B p65 (NF-κB p65) (MyBioSource, USA)—and apoptotic markers (caspase-3 and caspase-9) were purchased from Elabscience, USA. All solvents and chemicals used were of analytical-grade.
Experimental Rats
Male Wistar rats (n = 24; 10–12 weeks old) were obtained from the institutional animal facility. The animals were randomly assigned to four experimental groups (n = 6 per group) using a computer-generated random number table, with each rat given a unique identification number to ensure unbiased distribution of body weight and age across groups. Rats were housed under hygienic conditions, maintained on a 12 h light/dark cycle at 21°C–24°C with 50%–60% relative humidity, and provided with a standard pellet diet and water ad libitum. All animals were acclimatized to the laboratory environment for 7 days prior to the start of the experiment. This study was approved by the Ethics Committee of Hebei Provincial Hospital of Traditional Chinese Medicine (approval number: HBZY2022-KY-086-01).
Experimental Groups
After acclimatization, the rats were randomly divided into four experimental groups:
Group I (control): Received corn oil only.
Group II (B(a)P): Administered B(a)P (50 mg/kg body weight) twice weekly for 4 weeks to induce lung cancer.
Group III (ajmalicine + B(a)P): Pre-treated with ajmalicine (20 mg/kg body weight, orally) one day prior to B(a)P exposure and continued daily for 16 weeks.
Group IV (ajmalicine alone): Received ajmalicine (20 mg/kg body weight, orally) daily for 16 weeks without B(a)P exposure.
At the end of the treatment period, all rats were anesthetized and humanely euthanized. Blood and lung tissue samples were collected for further biochemical and histopathological analyses.
Analysis of Antioxidant Markers
The levels of antioxidant markers were quantified in lung tissue homogenates. The lungs were homogenized, centrifuged for 10 min at 12,000 rpm, and the resultant supernatant was retrieved and preserved at −20°C. The SOD, CAT, GSH, glutathione peroxidase (GPx), glutathione-S-transferase (GST), vitamin E (vit E), and vitamin C (vit C) were analyzed using kits. The tests were performed in triplicate according to the manufacturer’s specifications (Abcam, USA) (Munusamy et al., 2025).
Analysis of Biochemical Markers
The concentrations of cytochrome (cyt) P450, cyt B5, NADPH cyt C reductase, and caspase-3/9 in the experimental rats were analyzed utilizing commercial kits. The tests were performed in triplicate in accordance with the manufacturer’s specifications (MyBioSource, USA) (Sarathbabu et al., 2019).
Analysis of Inflammatory Markers
The TNF-α, IL-1β, and IL-6 concentrations in the serum of control and experimental rats were evaluated using the test kits and following the manufacturer’s specifications (Elabscience, USA).
Histopathology Analysis
The collected lungs were promptly immersed in formalin (10%), fixed in paraffin, and sectioned into 5–6 µm slices. Hematoxylin and eosin (H&E) staining was conducted to examine alterations in the lung tissue architecture under a microscope.
Statistical Analysis
Statistical analysis was performed using GraphPad Prism 9.0 (GraphPad Software, San Diego, CA, USA). Data are expressed as mean ± standard deviation (SD). Differences among multiple groups were assessed using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test for multiple comparisons. A p value of < .05 and < .01 was considered statistically significant.
Results
Effect of Ajmalicine on the Antioxidant Markers in Benzo(a)pyrene-induced Rats
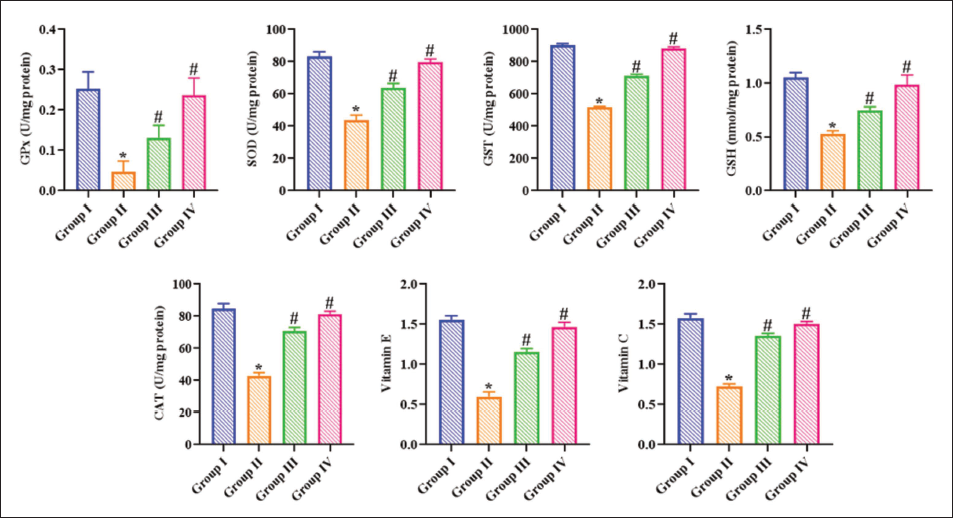
The present study observed marked diminution in the SOD, GPx, CAT, GSH, GST, vit E, and vit C levels in the lungs of B(a)P-treated rats relative to other groups. Whereas the ajmalicine treatment at a dose of 20 mg/kg remarkably increased those antioxidant concentrations in the lung tissues of B(a)P-induced rats, as seen in Figure 1. The administration of the same concentration of ajmalicine alone did not show a major effect on the levels of these antioxidants in the rats. These data suggest that providing ajmalicine to B(a)P rats may aid in detoxifying the carcinogen via its antioxidant properties.
Effect of Ajmalicine on the Xenobiotic Detoxification Enzymes in Benzo(a)pyrene-induced Rats
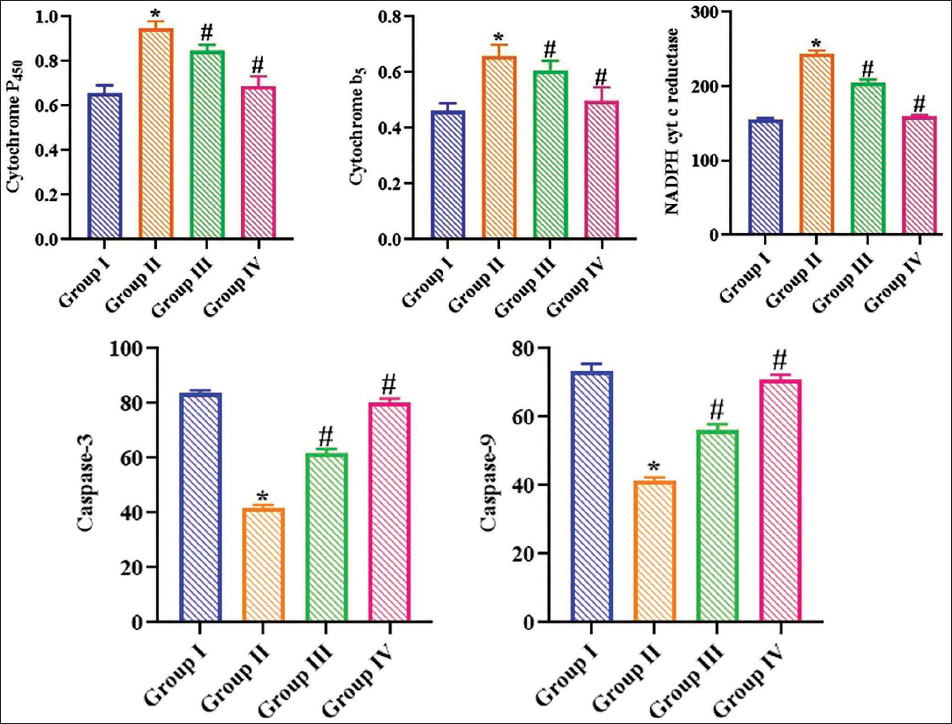
Figure 2 depicts the impact of ajmalicine on the concentrations of xenobiotic detoxification enzymes in the experimental rats. A significant increase in cyt P450, cyt B5, and NADPH cyt C reductase levels was seen in B(a)P-exposed rats. Captivatingly, the treatment of 20 mg/kg of ajmalicine markedly decreased the concentrations of cyt P450, cyt B5, and NADPH cyt C reductase in the B(a)P-treated rats. Furthermore, there are no significant alterations in these marker levels in the control and ajmalicine-alone administered rats.
Effect of Ajmalicine on the Inflammatory Markers in Benzo(a)pyrene-induced Rats
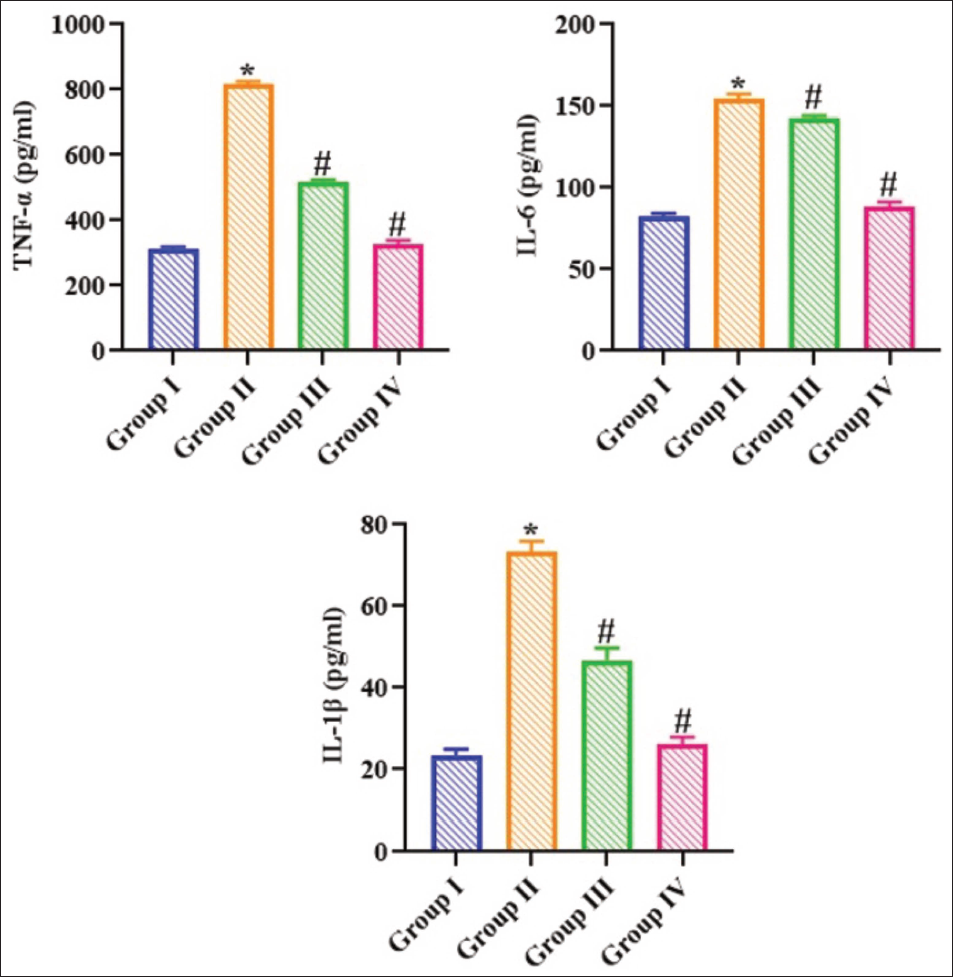
The B(a)P alone-treated rats had profound elevation in the inflammatory cytokines TNF-α, IL-1β, and IL-6, in comparison to the control group. The levels of these inflammatory cytokines were markedly diminished in B(a)P-exposed rats after the administration of 20 mg/kg of ajmalicine in Figure 3. The results indicated that ajmalicine exhibited considerable anti-inflammatory effects on the B(a)P-exposed rats.
Effect of Ajmalicine on the Caspase Activities in Benzo(a)pyrene-induced Rats
The activities of caspase-3/9 in experimental rats were measured, and the results are presented in Figure 2. The present findings showed that the activities of caspase-3/9 were considerably decreased in B(a)P-treated rats compared to controls. Conversely, the administration of ajmalicine (20 mg/kg) enhanced the activities of caspase-3/9 in the B(a)P-induced rats.
Effect of Ajmalicine on the Lung histology of Benzo(a)pyrene-induced Rats
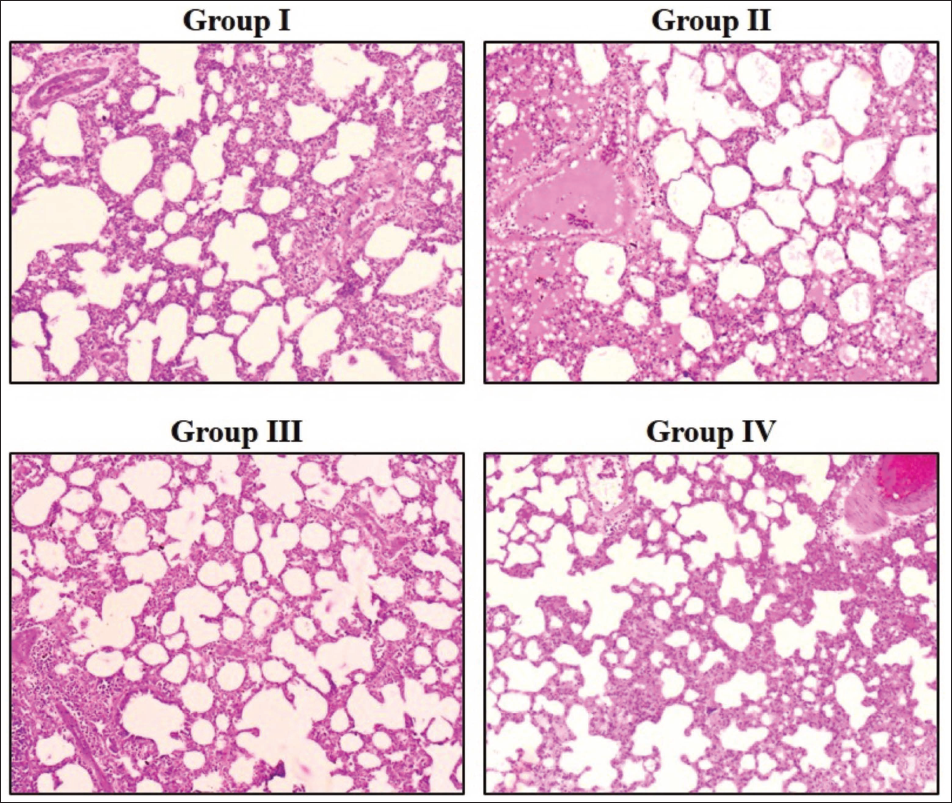
The histological examination of the lung tissues of the experimental rats was performed, and the results are indicated in Figure 4. The B(a)P rats showed deformed alveolar structure, hyperchromatic nuclei, proliferative lesions, focal bronchial changes, and alveolar epithelial hyperplasia of the lung tissues as compared to the control. Surprisingly, proliferative lesions and hyperplasia in the lungs of B(a)P rats were considerably decreased by the treatment with 20 mg/kg of ajmalicine.
Discussion
Lung cancer is one of the challenging health problems in the world, which is characterized as the leading cause of cancer-associated deaths globally. Although the early diagnosis mechanisms have been enhanced, a high percentage of lung cancer patients are diagnosed at the later stages, a factor that contributes to low overall survival rates (Rina et al., 2024). Lung cancer is a very insidious illness, the diagnosis of which is usually done at a later stage, which complicates the treatment process even more and decreases the survival rates. The absence of particular symptoms during the initial stages of the disease is a factor that leads to late detection of the disease, and as a result, the disease progresses and spreads to other parts of the body before it is treated. The late diagnosis is a critical prognosis factor, and the 5-year survival rate is much lower than in the situation with early-stage disease (Bade & Dela Cruz, 2020). To complicate the situation further, lung cancer is a heterogeneous disease, and it possesses a number of histological subtypes and molecular profiles, which should be chosen to select the treatment and predict sufficient response. In order to escalate the outcomes, screening is required in addition to early treatment that should be followed by proper staging. The molecular phenotype of lung cancer has been established clearly, and it is a complicated interdependence of genetic mutations, epigenetic variations, and changes of signaling pathways that cause tumorigenesis and evolution of the disease (de Groot et al., 2018). These molecular changes not only participate in the onset of the disease but are also subject to therapeutic targeting. The absence of a long-term effect of the current methods of treatment impairs the necessity to develop new methods of treatment that can effectively deal with the mechanisms of resistance and enhance patient survival (Boolell et al., 2015).
The oxidative stress, which is characterized by the disproportion between the increases in reactive oxygen species (ROS) and the antioxidant systems, has recently been determined as one of the primary causes of cancer development. The ROS generated during regular metabolism serves two functions in cell signaling and homeostasis (Shi et al., 2021). Though moderate levels of ROS play an essential role in the management of different signaling pathways, high levels can inhibit the cellular antioxidant mechanisms, leading to oxidative stress and then cell damage. These oxidative damages could be either DNA mutation, lipid peroxidation, or protein alterations, which are all part of causing tumorigenesis. This ability of ROS to cause carcinogenic effects is associated with the tendency to trigger genotoxicity and perturb essential cellular processes by oxidative stress (Bukowska & Duchnowicz, 2022). Oxidative stress can cause damage to proteins, lipids, DNA, and other cell compounds. A variety of environmental agents, including tobacco smoking, environmental carcinogens, chronic inflammation, and lifestyle-related etiologies, which include fat intake in the diet, are responsible for oxidative DNA damage, which results in tumor development. The cellular cycle is controlled in the interaction process of ROS with organic molecules (Gorrini et al., 2013).
Cancer cells possess very high intensity of oxidative stress than normal cells, such that these cells are reliant on the antioxidant systems to survive. ROS and oxidative stress have to be in balance to maintain the survival of cancer cells. The enhancements of metabolism, mitochondrial malfunctioning, and oncogene activation have been linked to this high oxidative stress and activate the formation of ROS. Tumor aggressiveness can also be influenced by high production of ROS by the cancer cells, which leads to cell proliferation, angiogenesis, and metastasis (Zhang et al., 2013). Pathways under hypoxia-inducible factors and endoplasmic reticulum stress responses are involved in the powerful cell adaptation to hypoxia, including its physiological and pathological types. Therapeutic methods have many obstacles and challenges that remain to be addressed. Another promising strategy toward the treatment of tumors is the redox status of tumor cells themselves, to specifically kill cancer cells without destroying normal cells. Modulation of antioxidant systems and oxidative stress is a potential approach to prevent or treat cancer. Greater interest is given to the oxidative pathway in its therapeutic application to enhance antioxidant defenses against cancer cells (Prasad et al., 2017). The intricate nature of the relationships between oxidative stress, the antioxidant defense, and the generation of cancers highlights the importance of redox behavior in cellular health and cancer prevention. Further studies should be conducted to understand the entire scope of the role of oxidative stress and antioxidants in cancer pathophysiology and to develop effective methods for preventing and treating cancer based on redox balance modulation (Aboelella et al., 2021). This study proved that the antioxidants of the lungs of B(a)P rats decreased significantly. But these antioxidants were increased in B(a)P rats due to the administration of ajmalicine, which evidences that ajmalicine may aid in the detoxification of the carcinogen through its antioxidant effect.
The three most important enzymes that are involved in the onset of lung cancer because of the exposure to B(a)P are Cyt P450, cyt b5, and NADPH cyt c reductase, which are highly involved in a complex process of metabolic activation and detoxification. Cyt P450 enzymes or CYPs are terminal oxidases that are found in electron transfer chains (Fuscoe et al., 2020). Such enzymes are strategically placed in the endoplasmic reticulum and the mitochondria, which are very important in the following ways: Drug metabolism, steroid, fatty acids, and prostaglandins formation. Cyt P450 enzymes are important in drug metabolism since they determine the drug activities, including their side effects, and can result in the formation of ions, oxygen-free radicals, or other active molecules (Zhao et al., 2021b). B(a)P is a ubiquitous environmental pollutant that is produced during incomplete combustion reactions; it is metabolically activated by cyt P450 enzymes, resulting in the production of reactive metabolites. This carcinogenic form of benzopyrene is then able to react with DNA, creating DNA adducts, which trigger mutations and genomic instability, which are the first steps towards the development of lung cancer (Shimada et al., 1999). CYP3A4, a major drug-metabolizing enzyme, is highly variable in its expression and activity across individuals and within individuals, with many environmental, genetic, and physiological factors playing a role, leading to unpredictable drug response and toxicity. Phase I drug-metabolizing enzymes, such as cyt P450s, catalyze oxidation, reduction, and hydrolysis reactions, which introduce or unmask small functional groups to the substrates, and moderately enhance their water solubility (Baum et al., 2001).
An electron transfer protein, cyt b5, is important in modulating the activity of some cyt P450 isoforms and affects the overall metabolic fate of benzopyrene. It also interacts with certain P450 enzymes, which help in transferring electrons and enhance the capacity of the catalytic cycle, thereby affecting the production of both detoxified and activated metabolites of benzopyrene (Geller et al., 1999). Cyt b5 enhances the catalytic properties of CYP3A4 and other P450 enzymes by transferring the second electron that is necessary to complete the monooxygenase reaction cycle. Cyt b5, as an allosteric effector, can activate the catalytic activity of some CYP isoforms through enhanced conformational changes, leading to more binding and unloading of substrates and products. Moreover, the balance between a bioactivation process and a detoxification process can be described by the combination of cytochrome b5 and P450 enzymes (Reed et al., 2018). The NADPH cyt c reductase is the central electron transferring component of cyt P450, whose feedback requires the reductases for the catalytic activity. This flavoprotein allows the movements of the electrons of NADPH towards cyt P450 and allows the monooxygenase reaction to occur. The reductase domain of NADPH cyt P450 reductase is linked to the heme-containing cyt P450s domain to transfer the electrons, and this is significant in the catalytic activity of P450 enzymes (Pandey & Flück, 2013). NADPH cyt c reductase is a key site in the cascade of electron transfer and can be a recipient of electrons donated by NADPH and subsequently transfers these to a broad range of redox partners, such as cyt P450s and heme oxygenase. The redox status of the cells may also regulate the activity of NADPH cyt c reductase and further change the total activity of cyt P450 enzymes and the metabolism of benzopyrene (Lin et al., 2012). Due to modifications in the activity of CYP enzymes, susceptibility of a person to benzopyrene-induced lung cancer can be significantly altered. In this study, we found that the concentration of the cyt P450, the cyt B5, and the NADPH cyt C reductase was significantly higher in the B(a)P rats. The concentrations of these enzymes were, however, reduced in the B(a)P-treated rats by the treatment of ajmalicine.
Inflammation is a complex biological reaction to harmful stimuli, which is now reported as a mandatory player in the development of cancers, including lung cancer. Although acute inflammation is a protective process that eliminates pathogens and triggers a tissue repair mechanism, chronic inflammation, however, may paradoxically promote tumorigenesis (Zhao et al., 2021a). The complex relationship between inflammation and cancer encompasses numerous factors, including immune cells, inflammatory mediators, and cellular signaling pathways. Tumors are frequently detected in the inflammatory tissues, which indicates the great importance of local inflammatory mediators in the carcinogenesis process (Hibino et al., 2021). Inflammation plays a key role in changing the tumor microenvironment to stimulate cell proliferation, cell survival, angiogenesis, and metastasis. DNA damage is an important connection between inflammation and the onset of tumors and may be caused by inflammatory conditions. In addition, the presence of inflammation influences the efficiency of anti-cancer therapy, which is a factor that facilitates treatment resistance (Kawanishi et al., 2017). The benzopyrene exposure triggers a chain of cellular and molecular processes that enhance lung inflammation. It is established that the tumor and inflammatory cells’ interactions result in an inflammatory tumor microenvironment. Such a milieu is marked by the release of various inflammatory markers, which in combination contribute to the onset of benzopyrene-induced lung cancer (Malik et al., 2018). Inflammation affects epithelial-mesenchymal transition and tumor stem cells, which are essential to cancer metastasis and treatment resistance. Angiogenesis is also triggered by inflammation, which makes metastasis more probable by breaking down the extracellular matrix that must be used to form new blood vessels (Shi et al., 2017).
Inflammation has been referred to as a mandatory factor that determines the manner in which tumors evolve and react to therapy. Metabolic abnormalities, mechanical forces, and a hypoxic environment that make up the tumor microenvironment have been linked to the development of therapy resistance alongside biochemical signaling (Greten & Grivennikov, 2019). The cytokines released by tumor cells attract tumor-infiltrating inflammatory cells to the tumor microenvironment. These inflammatory cells are initially an important first line of defense against the tumor through the production of ROS, the release of pro-inflammatory markers, and the promotion of cytotoxicity. The inflammatory cells may, however, adopt a different phenotype to become tumor-supportive cells as the tumor progresses. The latter is induced by continuous exposure to inflammatory cues and contact with tumor cells (Hou et al., 2021). These inflammatory cells that support tumors then secrete growth factors, pro-angiogenic factors, and immunosuppressive molecules that promote tumor growth, angiogenesis, and immune evasion. Inflammation-mediated phenotypic switch of inflammatory cells is critical to the creation of a tumor-supportive microenvironment. This leads to the development of tumors and resistance to treatment. Pro-inflammatory cytokines have multi-faceted roles in the case of benzopyrene-induced lung cancer (Cohen et al., 2015). The results in this work showed a severe increase in the inflammatory cytokines, including TNF-α, IL-1β, and IL-6. Remarkably, the levels of these cytokines were reduced in B(a)P rats after treating them with ajmalicine. The results indicate that ajmalicine showed an anti-inflammatory effect on the B(a)P rats.
Conclusion
In conclusion, our findings indicate that ajmalicine has chemopreventive effects on lung cancer induced by B(a)P in rats. The chemopreventive effect of ajmalicine in B(a)P-exposed lung cancer is linked to its antioxidant properties and its ability to detoxify the carcinogen through CYP enzymes. In addition, the inflammatory cytokines were decreased, and caspase-3/9 was increased by the ajmalicine in the B(a)P rats. It was therefore clear that ajmalicine possessed chemopreventive effects on the progression of lung cancer induced by carcinogens. However, additional confirmatory studies are strongly advised in the future to get a clear picture of the chemopreventive effect of ajmalicine.
However, while these findings provide strong preclinical evidence, further investigations are required to translate these effects into clinical applications. This research expands the pharmacological profile of ajmalicine and lays the groundwork for its future development as a potential chemopreventive or adjunct therapeutic agent against lung cancer.
Abbreviations
ANOVA: Analysis of variance; B(a)P: Benzo(a)pyrene; CAT: Catalase; CYP: Cytochrome P450; ELISA: Enzyme-linked immunosorbent assay; GSH: Reduced glutathione; GST: Glutathione-S-transferase; H&E: Hematoxylin and eosin; IL-1β: Interleukin-1 beta; IL-6: Interleukin-6; MDA: Malondialdehyde; NF-κB: Nuclear factor kappa-light-chain-enhancer of activated B cells; ROS: Reactive oxygen species; SOD: Superoxide dismutase; TNF-α: Tumor necrosis factor alpha.
Footnotes
Acknowledgments
The authors sincerely acknowledge the support of the Respiratory Department, Hebei Provincial Hospital of Traditional Chinese Medicine, Shijiazhuang, Hebei Province, China, for providing the necessary facilities and technical assistance to carry out this research work. The authors are also grateful to all laboratory staff and colleagues for their valuable help during the experimental procedures and analysis.
Data Availability
Data will be made available on request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This study is approved by the Ethics Committee of Hebei Provincial Hospital of Traditional Chinese Medicine (approval number: HBZY2022-KY-086-01).
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the research project titled “Study on the synergistic and attenuated effects of Lìfèi Jíedu Yìsǔi Decoction on chemotherapy for advanced lung cancer and its mechanism” (grant number: 2021088).
Informed Consent
NA.