
Abstract
Background
Diabetes mellitus (DM) is a metabolic disorder that has exhibited a significant rise in global incidence in recent years.
Purpose
Our study aims to investigate the wound-protective effect of fargesin against streptozotocin (STZ)-induced wounds in rats and to explore the underlying mechanism.
Materials and Methods
Intraperitoneal administration of STZ was used for the induction of DM in rats, and rats were randomized into five groups as follows: Gp I: normal control, Gp II: diabetic control group untreated wound, Gp III: diabetes and fargesin (5 mg/kg) treated wound, Gp IV: diabetes and fargesin (10 mg/kg) treated wound, and Gp V: diabetes and fargesin (20 mg/kg), respectively. Blood glucose level, insulin level, lipid parameters, collagen, hexosamine, macrophage migration inhibitory factor (MIF), nitric oxide (NO), epidermal growth factor (EGF), C-C motif chemokine ligand 2 (CCL-2), C-C motif chemokine ligand 3 (CCL-3), C-X-C motif chemokine ligand 2 (CXCL2), antioxidant parameters, inflammatory cytokines, inflammatory parameters, and mRNA expression were scrutinized.
Results
Fargesin treatment altered the levels of MIF, collagen, hexosamine, NO, EGF, C-C motif chemokine ligand (CCL2, CCL3), and CXCL2. Fargesin treatment decreased the glucose level and improved the insulin level. Fargesin treatment altered the level of lipid parameters, antioxidant, and inflammatory cytokines, namely, inflammatory parameters such as heme oxygenase-1 (HO-1) and nuclear factor erythroid 2-related factor 2 (Nrf2). Fargesin treatment altered the mRNA expression of cytokines and inflammatory parameters.
Conclusion
In conclusion, oral administration of fargesin ameliorates wound healing via alteration of inflammation and oxidative stress through the regulation of nuclear factor kappa B (NF-κB)/vascular endothelial growth factor (VEGF) and HO-1/Nrf2 signaling pathway.
Introduction
Diabetes mellitus (DM) is a complex group of chronic metabolic disorders characterized by impaired insulin secretion, hyperglycemia, and/or altered insulin action. It is strongly associated with an increased risk of various disease-related complications. The hallmark of DM is persistent hyperglycemia, which, over time, can lead to the accumulation of toxic metabolites in tissues (Verma et al., 2021; Yadav et al., 2024). Chronic hyperglycemia has long been associated with impaired wound healing, a major concern for diabetic patients. The prolonged exposure to elevated blood glucose levels leads to significant physiological impairments, among which endothelial dysfunction is particularly notable. This dysfunction plays a crucial role in the pathogenesis of various diabetic complications (Gautam et al., 2023). This dysfunction results in the downregulation of pro-angiogenic factors, thereby hindering the formation of new blood vessels necessary for tissue repair and regeneration. The combined effect of impaired angiogenesis and increased oxidative stress significantly compromises the wound healing capacity, rendering diabetic patients particularly susceptible to chronic, non-healing wounds and subsequent complications (Hyun et al., 2018; Tie et al., 2013).
Chronic hyperglycemia has been traditionally linked with impaired wound healing, posing a significant concern for diabetic patients. The prolonged exposure to elevated blood glucose levels in diabetes leads to a range of physiological impairments, among which endothelial dysfunction is particularly critical. It serves as a major factor in triggering and worsening numerous diabetes-related complications (Hyun et al., 2018; Singh et al., 2022).
During diabetes, these cells become dysfunctional, resulting in impaired regulation of blood flow, increased inflammation, and reduced ability to form new blood vessels. This endothelial dysfunction leads to the downregulation of pro-angiogenic factors, thereby hindering the formation of new blood vessels necessary for tissue repair and regeneration (Chen et al., 2021; Liu et al., 2023). The formation of new blood vessels, known as angiogenesis, is essential for effective wound healing as it provides sufficient blood flow to the repairing tissue. In diabetic patients, impairment of this process significantly delays wound healing and enhances the risk of developing chronic, non-healing wounds (Abdou et al., 2020; Hyun et al., 2018; Jin et al., 2018; Tie et al., 2013)
Wound repair is a complex and dynamic biological process comprising an orchestrated sequence of physiological and cellular events initiated to restore disrupted skin tissue integrity. This turnover is controlled by a complex interplay of cytokines and growth factors, which are important for triggering and modulating immune reactions and cellular compensatory processes. Immunity is a critical component of wound healing, as it prevents infection and promotes tissue regeneration. However, in conditions such as diabetes, impaired wound healing can disrupt this process, leading to a cascade of detrimental effects on the healing process (Ansell et al., 2018; Huang et al., 2019). One of the primary consequences of impaired wound healing is the overproduction of matrix metalloproteinases (MMPs). These enzymes play an essential role in normal wound healing by breaking down extracellular matrix components, which is a necessary step in normal wound healing. However, excessive MMP activity leads to disproportionate extracellular matrix (ECM) degradation, which further disrupts degradation and synthesis, contributing to the expansion of chronic, non-healing wounds, which are a common complication in diabetic patients (Anand et al., 2022; Ansell et al., 2018; Cheng et al., 2018; Senturk et al., 2016).
Fargesin, a natural neolignan, has received extensive attention in scientific research for its multi-pharmacological activities (Lee et al., 2021; Lu et al., 2021). Besides the traditional applications in East Asian medicine, further therapeutic activities have been unearthed through ongoing scientific examination. Fargesin displays multiple biological activities, such as anti-inflammatory, neuroprotective, and anti-oxidative effects, suggesting that fargesin could be a candidate drug for inflammatory disease therapy and neurodegenerative disease therapy (Lu et al., 2021; Pham et al., 2017; Sha et al., 2015; Wang et al., 2020). The structurally related fargesin has also been shown to have potential as a cardiovasculotonic agent, with studies showing involvement in the regulation of blood pressure and endothelial function. Fargesin may naturally have bioactivity due to its homolytic structure and to the multi-target action in the cell. This flexibility drew the attention of researchers to investigate its potential also in cancer therapy, for which observational studies have shown, in the beginning, anti-proliferative and proapoptotic effects in some cancer cell lines (Fu et al., 2019; Lu et al., 2021; Park et al., 2021). Even more intriguing is the suggested relevance to neurology due to the ability of the compound to pass the blood–brain barrier. This effect should greatly promote the fargesin in further drug discovery and development, and in fact, there are some investigations worldwide to explore the whole efficiency and the detailed mechanism of fargesin in the treatment (Lee et al., 2021; Park et al., 2021; Pham et al., 2017; Sha et al., 2015). The current study investigates the wound healing protective effect of fargesin against streptozotocin (STZ)-induced wounds in rats.
Materials and Methods
Experimental Animals
Preparation of the Toxicant and the Test Drug
Briefly, STZ was dissolved in citrate buffer (pH 4.5) and administered intraperitoneally at a dose of 60 mg/kg to overnight-fasted rats for the induction of DM (Gautam et al., 2023). The experimental rats were left for 7 days, and rats having the highest glucose levels (more than 300 mg/dL) were considered diabetic. The test drug (fargesin) was given to the rats in the form of an oral suspension using an oral gauge. The suspension was prepared using 1% carboxymethylcellulose (CMC) as the suspending agent. The suspension was freshly prepared before being given to the rats.
Experimental Design
The rats were randomly selected into five groups and each group contained six rats.
Gp I: normal control,
Gp II: diabetic control group wound untreated,
Gp III: diabetes and fargesin (5 mg/kg) treated wound,
Gp IV: diabetes and fargesin (10 mg/kg) treated wound, and
Gp V: diabetes and fargesin (20 mg/kg), respectively.
The dose of the tested drug (fargesin) was selected on the basis of the previous report method (Lu et al., 2021).
Wounding Operation
On day 14, the rats were anesthetized by intraperitoneal injection of ketamine (60 mg/kg body weight). The back hair was shaved with an electric hair clipper, and the WSG was marked with a skin marker. A full-thickness wound was then generated at the dorsal side by excision of the epidermal and dermal layers using a sterile scalpel.
Blood Glucose and Insulin Levels
Blood samples were drawn from the rat tail vein with acid-citrate dextrose-filled capillary tubes. The serum obtained was immediately centrifuged for 10 min at 10,000 rpm. Blood glucose levels were determined using a glucose assay kit (BioVision) following the manufacturer’s instructions. Serum insulin was measured with the ultrasensitive rat insulin ELISA kit (Morinaga Co., Ltd., Tokyo, Japan), according to the manufacturer’s instructions.
Lipid Parameters
Lipid parameters, such as total cholesterol (TC), triglyceride (TG), and high-density lipoprotein (HDL), were estimated using the available kit (Spam Diagnostic, India). The levels of low-density lipoprotein (LDL) and very low-density lipoprotein (VLDL) were estimated using the previously reported formula.
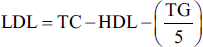
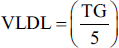
Collagen, Hexosamine, and Macrophage Migration Inhibitory Factor
The levels of collagen, hexosamine, and macrophage migration inhibitory factor (MIF) were estimated using previously reported methods with minor modifications (Abdou et al., 2020; Elson & Morgan, 1933; Woessner, 1961).
Epidermal Growth Factor, Nitric Oxide, C-C Motif Chemokine Ligand 2, C-C Motif Chemokine Ligand 3, C-X-C Motif Chemokine Ligand 2, Heme Oxygenase-1, and Nuclear Factor Erythroid 2-related Factor 2
The levels of epidermal growth factor (EGF), nitric oxide (NO), C-C motif chemokine ligand 2 (CCL2), C-C motif chemokine ligand 3 (CCL3), C-X-C motif chemokine ligand 2 (CXCL2), heme oxygenase-1 (HO-1), and Nrf2 were estimated using the commercial ELISA kit following the manufacturer’s instructions (R&D System, France).
Antioxidant Parameters
The levels of antioxidant parameters, including malondialdehyde (MDA), glutathione peroxidase (GPx), glutathione (GSH), catalase (CAT), superoxide dismutase (SOD), and total protein, were determined using previously reported methods with minor modifications (Abdou et al., 2020; Liu et al., 2023; Xie et al., 2022).
Inflammatory Cytokines and Parameters
The levels of inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), interleukin-1β (IL-1β), interleukin-17 (IL-17), interleukin-18 (IL-18), and interleukin-10 (IL-10), and inflammatory parameters, such as cyclooxygenase-2 (COX-2), prostaglandin E2 (PGE2), vascular endothelial growth factor (VEGF), transforming growth factor beta 1 (TGF-β1), and nuclear factor kappa B (NF-κB), were estimated using the ELISA kits (FANKEW, Shanghai, China).
Real-time Reverse Transcription Polymerase Chain Reaction
RNA was isolated from skin samples using TRIzol reagent (CWBio, Jiangsu, China) according to the manufacturer’s protocol. The extracted RNA was then reverse-transcribed into complementary DNA (cDNA) using a commercial reverse transcription kit (Yeasen, Shanghai, China). Quantitative polymerase chain reaction (qPCR) was conducted using the SYBR Green Real-Time PCR Master Mix (Yeasen, Shanghai, China) on a Roche LightCycler 480 II system (Germany). The PCR protocol included initial denaturation at 95°C for 5 min, followed by 40 amplification cycles consisting of denaturation at 95°C for 15 s, annealing at 58°C for 20 s, and extension at 72°C for 20 s. The relative expression of target genes was analyzed using the 2−∆∆Ct method, with β-actin used as the housekeeping gene for normalization.
Statistical Analysis
The data were analyzed using GraphPad Prism 8.0. Values are expressed as mean ± standard error of the mean (SEM). An unpaired Student’s t-test was used to compare two groups. For group comparison, one-way analysis of variance (ANOVA) test and Tukey’s post hoc test for multiple comparisons were performed. The level of statistical significance was set at p < .05.
Results
Glucose, Insulin, Collagen, Hexosamine, and Macrophage Migration Inhibitory Factor Levels
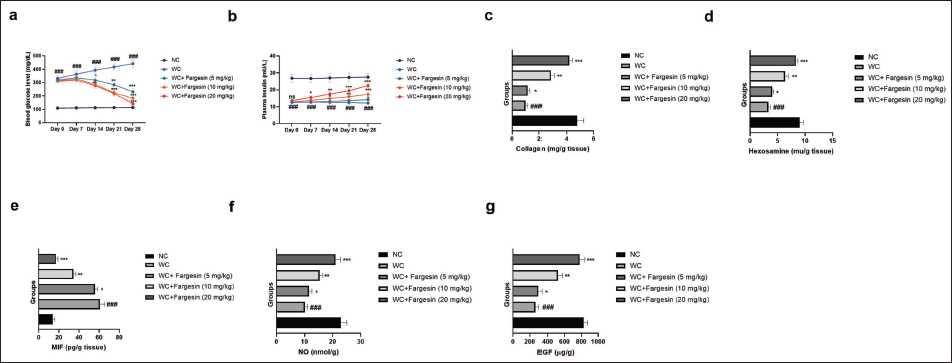
In this study, glucose and insulin levels were measured at regular time intervals. The normal control group did not exhibit any significant changes in glucose levels throughout the study. In contrast, the STZ-induced wound control (WC) group showed a significant (p < .001) increase in blood glucose levels, which remained elevated at all measured time points (days 0, 7, 14, 21, and 28). Treatment with fargesin significantly (p < .001) reduced glucose levels compared to the WC group (Figure 1a). STZ-induced group rats exhibited reduced insulin levels, collagen levels (Figure 1c), hexosamine levels (Figure 1d), and boosted levels of MIF (Figure 1e). Fargesin treatment significantly (p < .001) altered the levels of insulin, collagen, hexosamine, and MIF.
Effect of Fargesin on the Blood Glucose, Plasma Insulin, Collagen, Hexosamine, Macrophage Migration Inhibitory Factor (MIF), Nitric Oxide (NO), and Epidermal Growth Factor (EGF) Level Against Streptozotocin (STZ)-induced Diabetic Rats. (a) Blood Glucose Level, (b) Plasma Insulin Level. (c) Collagen, (d) Hexosamine, (e) MIF, (f) NO, and (g) EGF. Data are Expressed as Mean ± Standard Error of the Mean (SEM). Statistical Significance was Determined by One-way Analysis of Variance (ANOVA) Followed by Tukey’s Post Hoc Test. p < .05, p < .01, p < .001 Versus Control Group; ###p < .001 Versus Model Group.
Nitric Oxide and Epidermal Growth Factor Levels
The levels of NO (Figure 1f) and EGF (Figure 1g) were reduced in the STZ-induced WC group rats, and fargesin treatment significantly (p < .001) improved these levels.
Lipid Profile
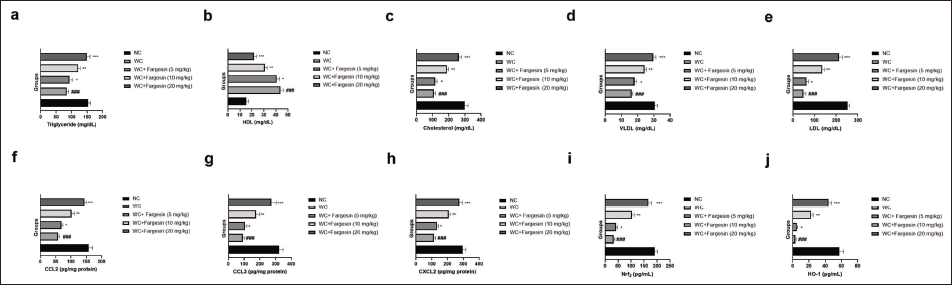
STZ-induced WC group rats exhibited altered lipid profile parameters such as triglyceride (Figure 2a), HDL (Figure 2b), cholesterol (Figure 2c), VLDL (Figure 2d), LDL (Figure 2e), CCL2 (Figure 2f), CCL3 (Figure 2g), CXCL2 (Figure 2h), nuclear factor erythroid 2-related factor 2 (Nrf2) (Figure 2i), and HO-1 (Figure 2j). Fargesin treatment significantly (p < .001) modulated the levels.
Effect of Fargesin on the Lipid, C-C Motif Chemokine Ligand 2 (CCL2), C-C Motif Chemokine Ligand 3 (CCL3), C-X-C motif chemokine ligand 2 (CXCL2), Nuclear Factor Erythroid 2-related Factor 2 (Nrf2), and Heme Oxygenase-1 (HO-1) Level Against Streptozotocin (STZ)-induced Diabetic Rats. (a) Triglyceride, (b) High-density Lipoprotein (HDL), (c) Cholesterol, (d) Very Low-density Lipoprotein (VLDL), (e) Low-density Lipoprotein (LDL), (f) CCL2, (g) CCL3, (h) CXCL2, (i) Nrf2, and (j) HO-1. Data are Expressed as Mean ± Standard Error of the Mean (SEM). Statistical Significance was Determined by One-way Analysis of Variance (ANOVA) Followed by Tukey’s Post Hoc Test. p < .05, p < .01, p < .001 Versus Control Group; ###p < .001 Versus Model group.
Oxidative Stress Parameters
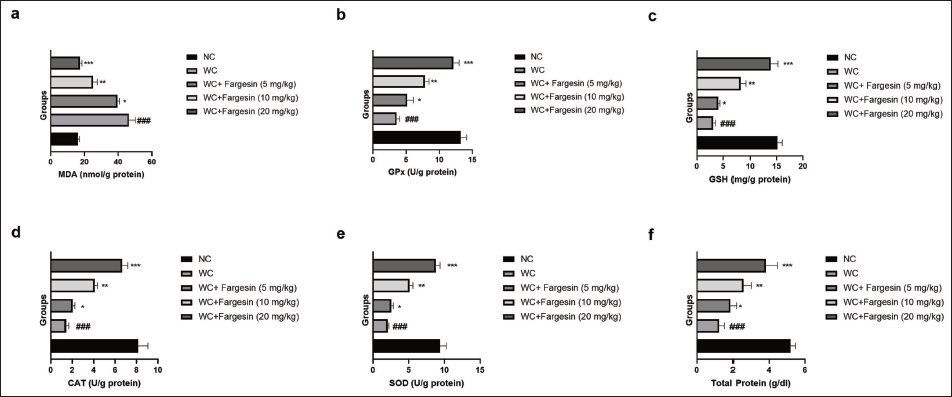
STZ-induced WC group rats showed altered levels of MDA (Figure 3a), GPx (Figure 3b), GSH (Figure 3c), CAT (Figure 3d), SOD (Figure 3e), and total protein (Figure 3f), and fargesin treatment significantly (p < .001) restored the levels of these oxidative stress parameters.
Effect of Fargesin on the Level of Oxidative Stress Parameters Against Streptozotocin (STZ)-induced Diabetic Rats. (a) Malondialdehyde (MDA), (b) Glutathione Peroxidase (GPx), (c) Glutathione (GSH), (d) Catalase (CAT), (e) Superoxide Dismutase (SOD), and (f) Total Protein. Data are Expressed as Mean ± Standard Error of the Mean (SEM). Statistical Significance was Determined by One-way Analysis of Variance (ANOVA) Followed by Tukey’s Post Hoc Test. p < .05, p < .01, p < .001 Versus Control group; ###p < .001 Versus Model group.
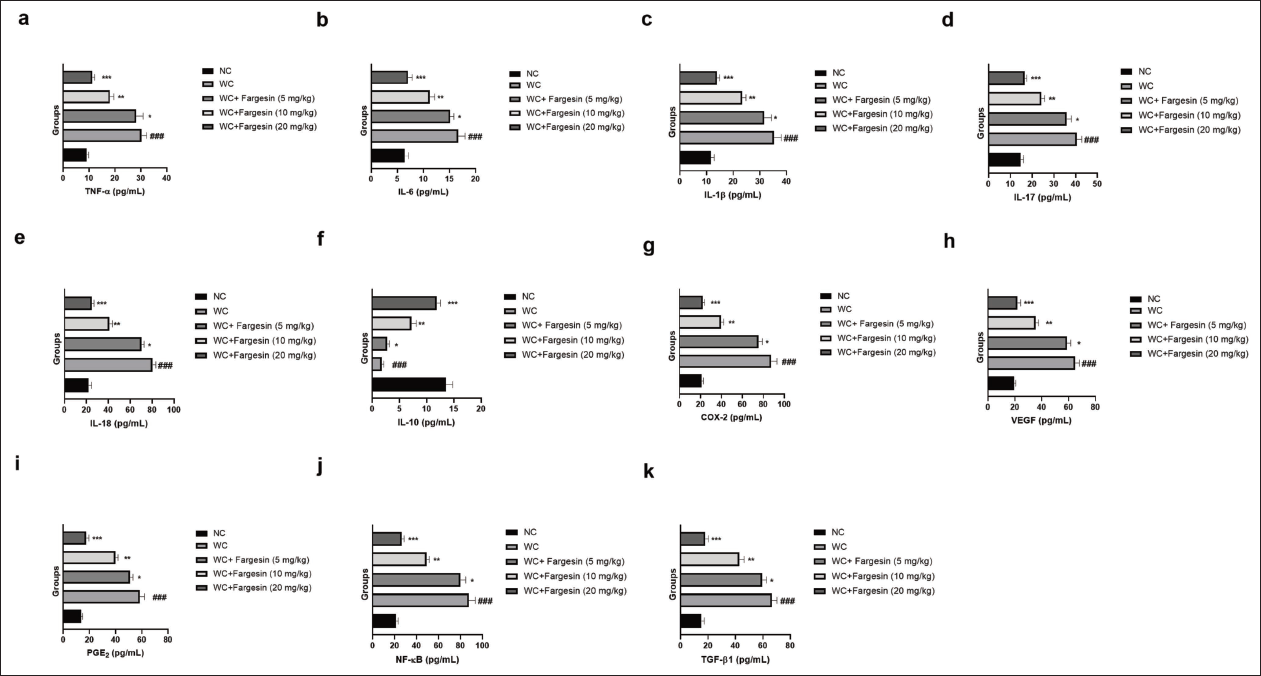
Cytokines and Inflammatory Parameters
STZ-induced WC group rats exhibited altered levels of TNF-α (Figure 4a), IL-6 (Figure 4b), IL-1β (Figure 4c), IL-17 (Figure 4d), IL-18 (Figure 4e), IL-10 (Figure 4f), COX-2 (Figure 4g), VEGF (Figure 4h), PGE2 (Figure 4i), NF-κB (Figure 4j), and TGF-β1 (Figure 4k), and fargesin treatment significantly (p < .001) modulated the levels of inflammatory cytokines and inflammatory parameters.
Effect of Fargesin on the Level of Inflammatory Cytokines and Inflammatory Parameters Against Streptozotocin (STZ)-induced Diabetic Rats. (a) Tumor Necrosis Factor-Alpha (TNF-α), (b) Interleukin-6 (IL-6), (c) Interleukin-1β (IL-1β), (d) Interleukin-17 (IL-17), (e) Interleukin-18 (IL-18), (f) Interleukin-10 (IL-10), (g) Cyclooxygenase-2 (COX-2), (h) Vascular Endothelial Growth Factor (VEGF), (i) Prostaglandin E2 (PGE2), (j) Nuclear Factor Kappa B (NF-κB) and (k) Transforming Growth Factor Beta 1 (TGF-β1). Data are Expressed as Mean ± Standard Error of the Mean (SEM). Statistical Significance was Determined by One-way Analysis of Variance (ANOVA) Followed by Tukey’s Post Hoc Test. p < .05, p < .01, p < .001 Versus Control Group; ###p < .001 Versus Model Group.
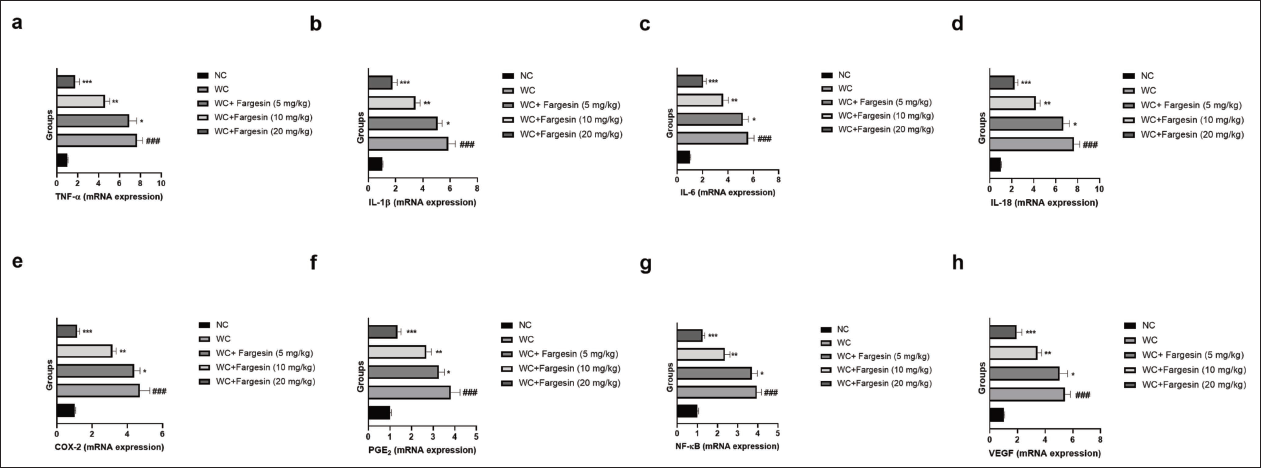
mRNA Expression
STZ-induced WC group rats exhibited the modulated mRNA expression levels of TNF-α (Figure 5a), IL-1β (Figure 5b), IL-6 (Figure 5c), IL-18 (Figure 5d), COX-2 (Figure 5e), PGE2 (Figure 5f), NF-κB (Figure 5g), and VEGF (Figure 5h), and fargesin treatment significantly (p < .001) altered the mRNA expression levels.
Effect of Fargesin on the mRNA Expression of Inflammatory Cytokines and Inflammatory Parameters Against Streptozotocin (STZ)-induced Diabetic Rats. (a) Tumor Necrosis Factor-Alpha (TNF-α), (b) Interleukin-1β (IL-1β), (c) Interleukin-6 (IL-6), (d) Interleukin-18 (IL-18), (e) Cyclooxygenase-2 (COX-2), (f) Prostaglandin E2 (PGE2), (g) Nuclear Factor Kappa B (NF-κB), and (h) Vascular Endothelial Growth Factor (VEGF). Data are Expressed as Mean ± Standard Error of the Mean (SEM). Statistical Significance was Determined by One-way Analysis of Variance (ANOVA) Followed by Tukey’s Post Hoc Test. p < .05, p < .01, p < .001 Versus Control Group; ###p < .001 Versus Model Group.
Discussion
Among its many complications, chronic wounds are particularly troublesome and prevalent. These non-healing wounds frequently lead to lower extremity amputations and are a major contributor to the increased morbidity and mortality associated with diabetes (Shofler et al., 2021; Villines, 2019). The delayed wound healing in DM is associated with a multifactorial pathogenesis, including impaired glucose metabolism and the development of both microvascular and macrovascular diseases. The STZ-induced DM model is widely used in research to study diabetes-related complications such as nephropathy, retinopathy, and foot ulcers, due to its similar pathogenesis to human DM (Baltzis et al., 2014; Bazrafshan et al., 2014; Chen et al., 2021).
The skin wound healing process is divided into three stages:
Inflammatory stage: This initial phase involves the secretion of high levels of inflammatory mediators and the activation of the immune system in response to injury. This stage helps clear the wound of debris and pathogens.
Proliferation stage: In this intermediate phase, fibroblasts proliferate, leading to the deposition of collagen fibers. This stage is crucial for tissue formation, as collagen provides structural support to the healing wound.
Angiogenesis and remodeling stage: The final phase involves angiogenesis, where new blood vessels form to supply the healing tissue. The ulcer tissue undergoes remodeling, which strengthens and matures the newly formed tissue, restoring skin integrity (Fang et al., 2023; Tan et al., 2018). This coordinated process is essential for effective wound healing. If any of these stages are disrupted, it can lead to delayed or impaired healing.
Reactive oxygen species (ROS) are key contributors to cellular damage and oxidative stress, which in turn can trigger the activation of multiple signaling pathways, notably the NF-κB pathway. In chronic inflammatory conditions such as diabetes, sustained ROS generation promotes the prolonged activation of NF-κB (Chen et al., 2021). The interplay between ROS and NF-κB not only exacerbates inflammation but also perpetuates the cycle of oxidative stress and tissue damage, making the management of inflammation a critical aspect of diabetic wound healing (Dasari et al., 2021; Salazar et al., 2016; Wu et al., 2022). Targeting ROS production or NF-κB activation represents a potential therapeutic strategy for managing chronic inflammation and improving healing outcomes in diabetic patients.
The presence of VEGF at the wound site is essential for delivering oxygen and nutrients to the regenerating tissue. However, the timing and regulation of VEGF expression are critical to ensuring effective and orderly wound repair (Johnson and Wilgus, 2014). During the early stages of wound healing, there is a significant upregulation of VEGF. This growth factor is primarily produced by keratinocytes, macrophages, and fibroblasts in response to hypoxia and inflammatory cytokines (Johnson and Wilgus, 2014; Keswani et al., 2013; Wilgus et al., 2008). This can facilitate the initial inflammatory response and tissue regeneration. While VEGF-induced angiogenesis is beneficial for healing, an excessively high level of VEGF can delay wound contraction. This is because the newly formed blood vessels can interfere with the normal contractile function of myofibroblasts, which are responsible for pulling the wound edges together. The delayed contraction may lead to prolonged healing times and potentially contribute to the development of chronic wounds, particularly in diabetic patients. As wound healing progresses, the levels of VEGF typically decrease, especially after the epithelialization phase (the formation of new epithelial tissue over the wound) (Graiani et al., 2004; Johnson and Wilgus, 2014; Keswani et al., 2013; Wilgus et al., 2008). At this stage, the need for new blood vessels diminishes, and a balance is struck between the formation of new tissue and the resolution of inflammation. Proper regulation of VEGF expression is essential for the transition from the proliferative phase to the remodeling phase of wound healing. High levels of VEGF in the later stages may disrupt this balance, leading to excessive vascularization and impaired wound closure. VEGF plays a dual role in wound healing, promoting angiogenesis and tissue repair early on but requiring careful regulation to avoid delayed wound contraction and chronic inflammation (Johnson and Wilgus, 2014; Wilgus et al., 2008).
The cellular defense to oxidative stress, especially the diabetic wound healing process, plays an important role in the Nrf2/HO-1 signaling pathway. In diabetes, sustained hyperglycemia leads to increased oxidation in the body, resulting in elevated ROS and oxidative damage that inhibit normal wound healing events, including inflammation resolution, cellular proliferation, and angiogenesis. Nrf2 induction increases the expression of an antioxidant protein, HO-1, which protects against oxidative stress. By decreasing ROS and modulating inflammation, apoptosis, fibrosis, cell proliferation, and angiogenesis, HO-1 is a major factor in enhancing wound healing in the diabetic setting (Accipe et al., 2023; Long et al., 2016).
HO-1 has emerged as a potential therapeutic target for improving diabetic wound healing. Strategies that enhance Nrf2 activation or directly increase HO-1 expression may help to accelerate wound healing, reduce chronic inflammation, and improve overall outcomes in diabetic patients (Bhatti et al., 2022). These genes play vital roles in protecting cells from oxidative damage and promoting tissue regeneration (Bhatti et al., 2022). The Nrf2/antioxidant response element (ARE) pathway is crucial for counteracting this oxidative stress by upregulating antioxidant enzymes. Antioxidant enzymes are critically involved in the neutralization of reactive oxidants and protection of cells and tissues from oxidative injury, and wound healing. The Nrf2/ARE signaling pathway is also involved in resolving inflammation, which is crucial for the switch from the inflammatory to the proliferative phase of healing. Nrf2 contributes to tissue repair through suppression of pro-inflammatory cytokines and activation of anti-inflammatory mediators, allowing tissue healing to be effective (Aleksunes & Manautou, 2007; Mittal et al., 2014). The Nrf2/ARE pathway supports the proliferation and survival of cells involved in wound healing, including fibroblasts and keratinocytes. By protecting these cells from oxidative stress and apoptosis, Nrf2 ensures that sufficient cell populations are available to regenerate tissue and close the wound. Superoxide dismutase 1 (SOD-1) is an essential antioxidant enzyme that converts superoxide radicals (a type of ROS) into less harmful molecules like hydrogen peroxide. This enzyme is crucial for reducing oxidative stress during wound healing. Deficiencies in SOD-1 have been shown to delay wound healing, as elevated levels of superoxide radicals can lead to cellular damage, inflammation, and impaired tissue repair (Süntar et al., 2021). The Nrf2/ARE pathway, through the regulation of key antioxidant enzymes like SOD-1 and HO-1, plays an important role in facilitating wound repair. Deficiencies in these enzymes result in delayed wound healing due to increased oxidative stress, inflammation, and impaired cellular functions. Therefore, targeting the Nrf2/ARE pathway to enhance the expression of SOD-1, HO-1, and other cytoprotective genes holds significant therapeutic potential for improving wound healing (Liu et al., 2023).
One of the limitations of our study is the absence of direct mechanistic proof of Nrf2 and NF-κB as targets modulated by fargesin. While the results suggest a possible regulatory role based on functional analysis, validation test at the protein level, such as Western blot analysis or gene expression profiling, was not conducted to verify the Nrf2 activation and the NF-κB inhibition. Further investigation of downstream pathways and the molecular mechanism of fargesin is needed using molecular techniques such as immunoblotting, qPCR, or reporter assays.
Conclusion
Fargesin treatment demonstrated significant therapeutic potential by markedly reducing glucose levels and enhancing insulin levels. It also significantly improved lipid profiles, oxidative stress parameters, and various biomarkers involved in collagen synthesis, hexosamine, and macrophage migration (MIF, CCL2, CCL3, CXCL2). Additionally, fargesin effectively modulated inflammatory responses by reducing levels of pro-inflammatory cytokines such as TNF-α, IL-6, IL-1β, IL-17, IL-18, and the anti-inflammatory cytokine IL-10. Furthermore, it inhibited key inflammatory mediators such as COX-2, VEGF, PGE2, NF-κB, and TGF-β1. The suppression of mRNA expression of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6, IL-18) and other inflammatory markers (COX-2, PGE2, NF-κB, VEGF) suggests that fargesin exerts potent anti-inflammatory and antioxidant effects, indicating its potential as a therapeutic agent for managing metabolic and inflammatory disorders.
Footnotes
Abbreviations
ANOVA: Analysis of variance; ARE: Antioxidant response element; CAT: Catalase; CCL2: C-C motif chemokine ligand 2; CCL3: C-C motif chemokine ligand 3; CMC: Carboxymethylcellulose; COX-2: Cyclooxygenase-2; CXCL2: C-X-C motif chemokine ligand 2; DM: Diabetes mellitus; ECM: Extracellular matrix; EGF: Epidermal growth factor; GPx: Glutathione peroxidase; GSH: Glutathione; HDL: High-density lipoprotein; HO-1: Heme oxygenase-1; IL-1β: Interleukin-1β; IL-6: Interleukin-6; IL-10: Interleukin-10; IL-17: Interleukin-17; IL-18: Interleukin-18; LDL: Low-density lipoprotein; MDA: Malondialdehyde; MIF: Macrophage migration inhibitory factor; MMPs: Matrix metalloproteinases; NF-κB: Nuclear factor kappa B; NO: Nitric oxide; Nrf2: Nuclear factor erythroid 2-related factor 2; PGE2: Prostaglandin E2; qPCR: Quantitative polymerase chain reaction; SD: Sprague-Dawley; SEM: Standard error of the mean; SOD: Superoxide dismutase; SOD-1: Superoxide dismutase 1; STZ: Streptozotocin; TC: Total cholesterol; TG: Triglycerides; TGF-β1: Transforming growth factor beta 1; TNF-α: Tumor necrosis factor-alpha; VEGF: Vascular endothelial growth factor; VLDL: Very low-density lipoprotein; WC: Wounded control.
Acknowledgment
The authors are very thankful to the Fourth People’s Hospital of Longgang District for its support in the research.
Declaration of Conflict of Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The whole study was approved from the departmental animal house. The current study was approved from the The Fourth People’s Hospital of Longgang District (Ethical approval no. 20241104).
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.