
Abstract
An atypical urease-negative mutant of Actinobacillus pleuropneumoniae serovar 2 was isolated in Japan. Nucleotide sequence analysis of the urease gene cluster revealed that the insertion of a short DNA sequence into the cbiM gene was responsible for the urease-negative activity of the mutant. Veterinary diagnostic laboratories should be watchful for the presence of aberrant urease-negative A. pleuropneumoniae isolates.
Keywords
Actinobacillus pleuropneumoniae is the etiologic agent of porcine pleuropneumonia, a severe respiratory disease that causes serious economic losses in the pig-rearing industry. 8 Sixteen serovars have been recognized, based mainly on the antigenic diversity of their capsular polysaccharides.3,12
The urease-positive activity of A. pleuropneumoniae is a key characteristic for differentiating these organisms from other V-factor–dependent Pasteurellaceae species isolated from pigs. 4 One exception is that a urease-negative variant of A. pleuropneumoniae serovar 1 was isolated from the lungs of dead pigs in the United States. 4 However, the genetic background of the urease-negative activity of the isolate was not characterized. Later, it was revealed that the urease gene cluster involved in urease activity of A. pleuropneumoniae consisted of 3, 4, and 5 genes encoding the structural subunit proteins (UreABC), accessory proteins (UreEFGD), 5 and nickel transport proteins (CbiKLMQO), 7 respectively.
Urease activity is considered to be a virulence factor of A. pleuropneumoniae. The activity is not required for development of acute pleuropneumonia 13 but is required for chronic infection. 2 Urease activity may also contribute to the ability of A. pleuropneumoniae to establish infection in the porcine respiratory tract. 6 We report herein our isolation of a urease-negative mutant of A. pleuropneumoniae serovar 2 in Japan and our genetic characterization of the urease gene cluster, which we performed to elucidate the molecular basis of the urease-negative activity.
On a farrow-to-finish operation farm, six 90-d-old pigs showed clinical signs, such as coughing and dyspnea, and 3 of the pigs died in September 2013. A. pleuropneumoniae–like organisms were isolated from lung lesions of the 3 dead pigs with pleuropneumonia. Lung specimens were inoculated on Nagasaki Haemophilus medium agar (Kyokuto Pharmaceutical Industrial, Tokyo Japan; a medium that contains 0.02% β-nicotinamide dinucleotide [NAD] plus 5% defibrinated horse blood) and incubated at 37°C in the presence of 5% CO2. One bacterial colony from each of the 3 pigs was chosen for further analysis; the 3 isolates were tentatively identified as A. pleuropneumoniae by the presence of hemolysis and the requirement for NAD for in vitro growth, although they showed aberrant urease-negative activity.
The biochemical characteristics of the 3 isolates were further examined using a biochemical identification kit (ID Test HN20-Rapid, Nissui Pharmaceutical, Tokyo, Japan). The 7-digit biochemical profile number generated by the ID test was 7007373. These biochemical properties as well as the biological characteristics, such as hemolytic activity and absence of growth on MacConkey agar plates, were consistent with those of A. pleuropneumoniae, except that the 3 isolates were urease negative. All 3 isolates were serotyped as serovar 2 by slide agglutination tests as described previously. 11
These 3 isolates were named UN16, UN21, and UN26, respectively, and the nucleotide sequences of the 16S ribosomal (r)DNA of each isolate were determined. The primers 5’-ATTGAACGCTGGCGGCAGGCTTAACACATG-3’ and 5’-AAGGCCCGGGAACGTATTCACCGCAACATT-3’ were used for PCR amplification of the 16S rDNA gene. The PCR assay, sequencing, and sequence analysis were essentially the same as described previously. 9 The nucleotide sequence of the 16S rDNA of the 3 strains was identical to the 16S rDNA sequence of A. pleuropneumoniae serovar 2 (GenBank accession AF033058). Nucleotide sequences of the 3 urease-negative isolates were deposited in the DNA Data Bank of Japan (DDBJ)/European Molecular Biology Laboratory (EMBL)/GenBank databases as accessions LC227658, LC227659, and LC227660, respectively.
In order to elucidate the molecular basis of the absence of urease activity by the mutant, the urease gene cluster was amplified from strain UN16 and a urease-positive serovar 2 strain CB87 that was isolated from the lungs of a dead pig raised on a different farm. PCR primers for the amplification of the urease gene cluster (5’-TACGGCTCGGCTAATAACCGGCGGAGAAAC-3’ and 5’-TTGGTCGTTAAGCGAAGCACGAATTAAGTT-3’) were designed from previously published sequences.5,7 The conditions for PCR, so-called stepdown PCR with KOD-FX Neo (Toyobo, Osaka, Japan), were as follows: 94°C for 2 min (1 cycle); 98°C for 10 s and 74°C for 10 min (5 cycles); 98°C for 10 s and 72°C for 10 min (5 cycles); 98°C for 10 s and 70°C for 10 min (5 cycles); 98°C for 10 s and 68°C for 10 min (20 cycles); and 68°C for 10 min. Purification of the amplified DNA and determination of the complete nucleotide sequence using a primer walking strategy were carried out as described above.
The genetic map and comparison of the nucleotide sequences of the urease gene cluster of the urease-positive and -negative strains are shown in Figure 1. The nucleotide sequence analysis of the urease-negative A. pleuropneumoniae strain UN16 revealed that a cause of the urease-negative activity of the strain might be an insertion of 7 nucleotides (AATTACC) into position 392 of the cbiM sequence, which yields a tandem direct repeat of this sequence leading to a frameshift mutation, resulting in truncation of the CbiM protein. Urease activity of strain UN16 was recovered in the presence of NiCl2 at a concentration of 20 μM. This suggests that an alternative system, such as the magnesium uptake system, 7 with a lower affinity for nickel, mediates uptake at higher concentrations. Urease activity of UN16 in the presence of elevated nickel is consistent with the mutation of the cbiM gene being the cause of the urease-negative phenotype of this strain when tested under normal conditions. The nucleotide sequences of the urease gene cluster of strains CB87 and UN16 have been deposited in the DDBJ/EMBL/GenBank databases as accessions LC227661 and LC227662, respectively.
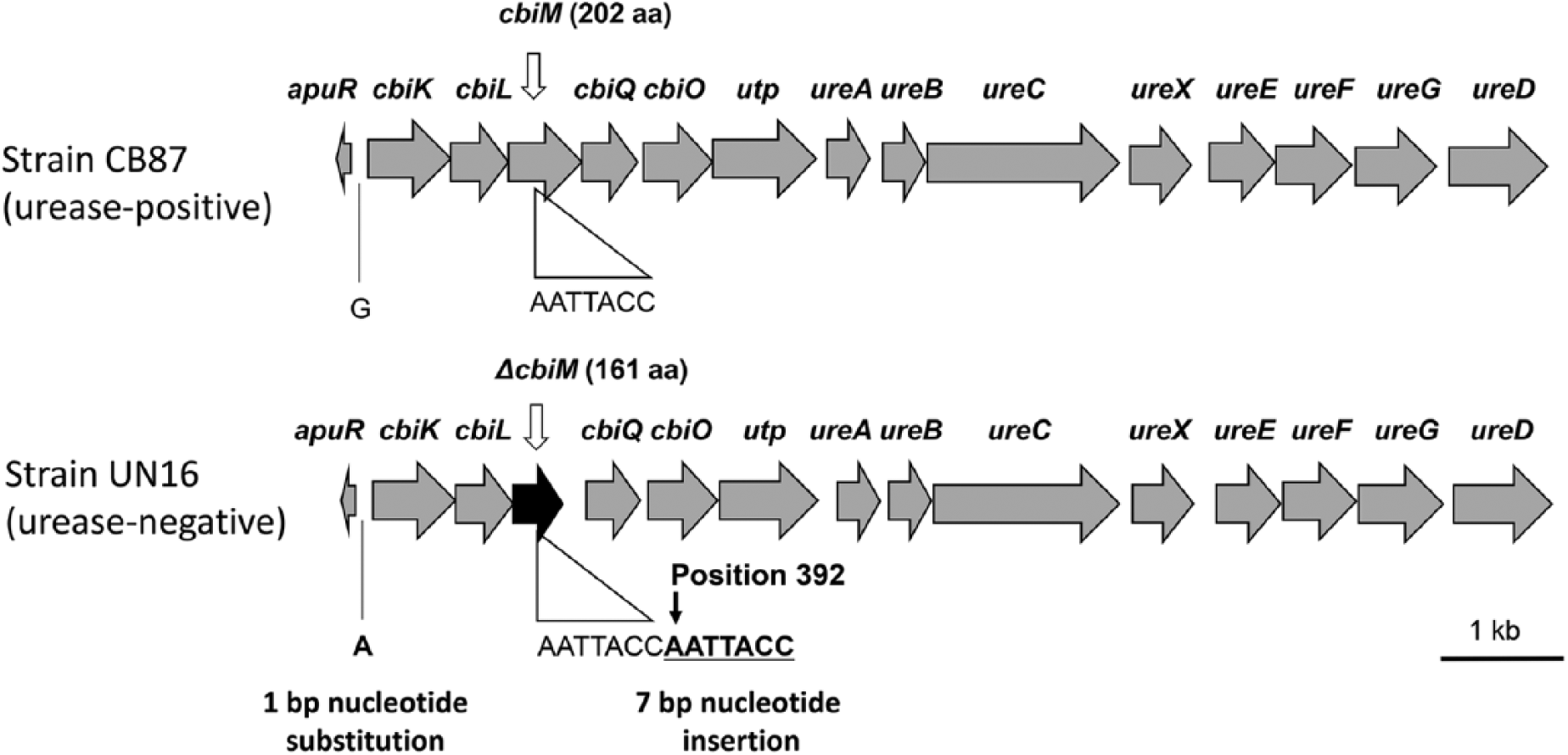
Comparison of the nucleotide sequences of the urease gene cluster of urease-positive and -negative strains. Genes of the urease-positive strain CB87 are indicated by gray arrows. Genes of the urease-negative strain UN16, which has a nucleotide sequence identical to that of strain CB87, are also shown by gray arrows. cbiM genes of strain UN16, which are truncated by an insertion of DNA (ΔcbiM), are shown by black arrows. The number plus amino acid (aa) in parenthesis after cbiM and ΔcbiM genes indicates the length of the aa sequence, which differed between strains CB87 and UN16.
To date, a number of A. pleuropneumoniae mutants that do not express one of the virulence factors, such as Apx toxins,1,14 a biofilm, 10 a capsule, 9 have been isolated. For these mutants, insertion of a mobile genetic element, such as ISApl1 (a 1,070-bp insertion element of the IS30 family)9,10,14 or an uncharacterized mobile genetic element (1,201 bp), 1 was seen to disrupt a gene encoding the respective phenotype. However, our study revealed that tandem duplication of a short DNA sequence might be involved in the urease-negative activity of an A. pleuropneumoniae mutant.
Footnotes
Acknowledgements
We thank Drs. T Shimada (Chiba Prefecture Agricultural Mutual Aid Associations), M Ooi (Toyoura Veterinary Clinic, Japan), Y Okamura (Okamura Animal Clinic, Japan), and T Fujiwara (Fujiwara Animal Hospital, Japan) for providing the lung specimens. We also thank Drs. J Sekiya (Japan Livestock Industry Association), S Miyajima (Japan Livestock Industry Association), A Ogura (Ministry of Agriculture, Forestry and Fisheries), and K Ishikawa (Ministry of Agriculture, Forestry and Fisheries) for arranging the collection of the bacterial isolates.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This study was funded in part by the Ministry of Agriculture, Forestry and Fisheries, Japan.