
Abstract
A panzootic caused by highly pathogenic avian influenza (HPAI) A(H5N1) virus, clade 2.3.4.4b, has affected many animal species around the world since 2021. In March 2024, genotype B3.13 of this virus was identified in dairy cattle in the United States, following a spillover event from wild birds. Mammary gland lesions were a key finding in infected cows, with infectious virus detected in their milk. Raw milk is sold legally in retail establishments in multiple US states, including California. In November 2024, HPAI A(H5N1) virus, clade 2.3.4.4b, genotype B3.13, was detected in raw milk sold commercially in California and then in bulk milk tanks. The affected product later was recalled. We describe an 8-mo-old cat with a history of severe illness after consuming this raw milk before it was recalled. The cat was euthanized and submitted for postmortem examination and diagnostic workup. Autopsy and histopathology revealed icterus, nasal discharge, hydrothorax, gliosis, and necrotizing pneumonia, hepatitis, and salpingitis, among other lesions. Immunohistochemistry for influenza A virus revealed intralesional immunolabeling in many organs. Molecular detection was positive for HPAI A(H5N1) virus, clade 2.3.4.4b, genotype B3.13. To our knowledge, HPAI A(H5N1) virus has not been reported previously in cats after consuming raw milk purchased from a retail establishment, nor has salpingitis been associated with HPAI A(H5N1) virus infection in a mammal. Hepatic damage and icterus were prominent findings in our case rather than primary involvement of the CNS.
Influenza A virus (
The molecular composition of the hemagglutinin cleavage site motif (
Since 2021, HPIAV subtype Eurasian H5N1, clade 2.3.4.4b [HPAI A(H5N1) virus] has caused an unprecedented panzootic, impacting wild and domestic bird populations around the globe.22,27 HPAI A(H5N1) virus has spilled over to multiple terrestrial and marine mammalian species,1,19,21,27,29,39 including humans.13,15,26 In marine mammals, such as cetaceans, lesions caused by HPAI A(H5N1) virus infection, mainly targeting the CNS, have been reported.37,48
One well-known route of natural infection with HPIAV in felids is by direct contact or ingestion of infected birds, 24 and these species appear to be particularly susceptible to infection with HPAI A(H5N1) virus. 36 Severe disease and death have been reported in infected domestic cats, bobcats, Bengal tigers, and cougars, among others.20,21,42 Conclusive evidence of mammal-to-mammal transmission is still elusive, but has been suggested to have occurred in mink as a result of a mutation of the virus.1,25
In March 2024, HPAI A(H5N1) virus, clade 2.3.4.4b, genotype B3.13 was first detected in commercial dairy cattle in Texas. Since then, the virus has spread to dairy farms across the country. As of 2025 May 13, >1,053 dairy herds in 17 states had been affected, with most cases occurring in California. 16 Clinical signs in cattle infected with HPAI A(H5N1) virus are nonspecific and include an abrupt drop in milk production, with reduced rumination and feed consumption.9,12 The virus targets the mammary gland and can be detected in milk. 9 On 2025 Jan 31, the USDA reported a second strain of HPAI A(H5N1) virus called D1.1 (H5N1 clade 2.3.4.4b, genotype D1.1) in dairy cattle. The strain produced an outbreak on a dairy farm in Nevada, 50 with new mutations in the clade that could potentially result in a change of tissue tropism and greater pathogenicity and higher mortality.18,33
Domestic cats on infected dairy farms have also been affected.9,36 In most cases, these cats had been fed colostrum or raw milk from infected cows, and developed lethargy, weakness, inappetence, oculonasal discharge, and neurologic signs before death.9,36 The primary lesions found in these cases included encephalitis, myocarditis, chorioretinitis, sialadenitis, and interstitial pneumonia, with virus detected in several tissues by PCR and immunohistochemistry (IHC). 36
We describe a case of HPAI A(H5N1) virus infection in a domestic cat that became infected after consuming raw milk sold for human consumption in California. The cat was from a suburban Los Angeles household with an indoor-only group of 7 cats from different litters. The raw milk was offered as a treat in late November. The raw milk was from a retail establishment and was recalled after detection of HPAI A(H5N1) virus in milk bulk tanks. 10 The owner of the cat did not work with cattle, poultry, or wildlife. We describe the gross, microscopic, and ultrastructural lesions and the viral antigen distribution in this cat.
All 7 cats became sick 8–14 d after drinking the raw milk, and 5 died or were euthanized because of their illness. The first cat to become affected had tested positive for feline panleukopenia virus (
Our case was an 8-mo-old, intact female Siamese-mix cat. At the time of presentation of the cat, the cat owner assumed that it was ill with FPV because of the test result on another cat in the group. However, after our case and a second cat in the home tested negative for FPV, the cat owner reported to the veterinary practice that all the cats had been fed raw milk 11 d before it was recalled because of HPAI A(H5N1) virus contamination. The veterinary practice reported this to the local public health agency, triggering a public health investigation and PCR testing for HPAI A(H5N1) virus in multiple cats in the home (manuscript in preparation). This was the only cat in the cluster that was submitted for autopsy.
Initial clinical signs in this cat and clinicopathologic alterations included fever (39.7°C, rising to 40.9°C the next day), dehydration, elevated liver enzyme activities and bilirubin values (ALT 267 U/L [RI: 6–60 U/L]) rising to 752 U/L the next day; bilirubin (65 µmol/L [RI: 0–8.6 µmol/L], 3.8 mg/dL [RI: 0–0.5 mg/dL]) rising to 84 µmol/L (4.9 mg/dL) the next day, thrombocytopenia (40–56 × 109/L [RI: 200–500 × 109/L], 40–56,000/µL [RI: 200-500 K/µL]), bilirubinuria (3+), and bilirubin crystalluria (1–5/hpf). The cat appeared eupneic, and no significant abnormalities were noted in thoracic or abdominal radiographs. The cat was hospitalized and given fluids IV, treatment for potential toxoplasmosis (clindamycin, enrofloxacin), and additional supportive care, but declined rapidly over the following day, becoming visibly icteric. Late the next day, the cat developed respiratory arrest, with bradycardia and a ventricular escape rhythm. It regurgitated a large volume of fluid, which was partly aspirated, and then developed full cardiopulmonary arrest. The cat arrested and was resuscitated 2 times before being euthanized.
The carcass was submitted to the California Animal Health and Food Safety Laboratory System (CAHFS; San Bernardino, CA, USA) in December 2024 for postmortem examination and diagnostic workup. A full autopsy was performed ~36 h after death. The carcass was moderately fresh, and in good nutritional condition, with moderate amounts of fat reserves and well-fleshed. The conjunctivae, oral mucous membranes, and fat were diffusely yellow. Bilateral serous nasal discharge was observed. The thoracic cavity contained ~80 mL of serosanguineous fluid. The tracheal mucosa was diffusely red, and the lungs failed to collapse when the thoracic cavity was opened. The lung parenchyma had dark-red foci, and all lobes appeared congested and edematous (
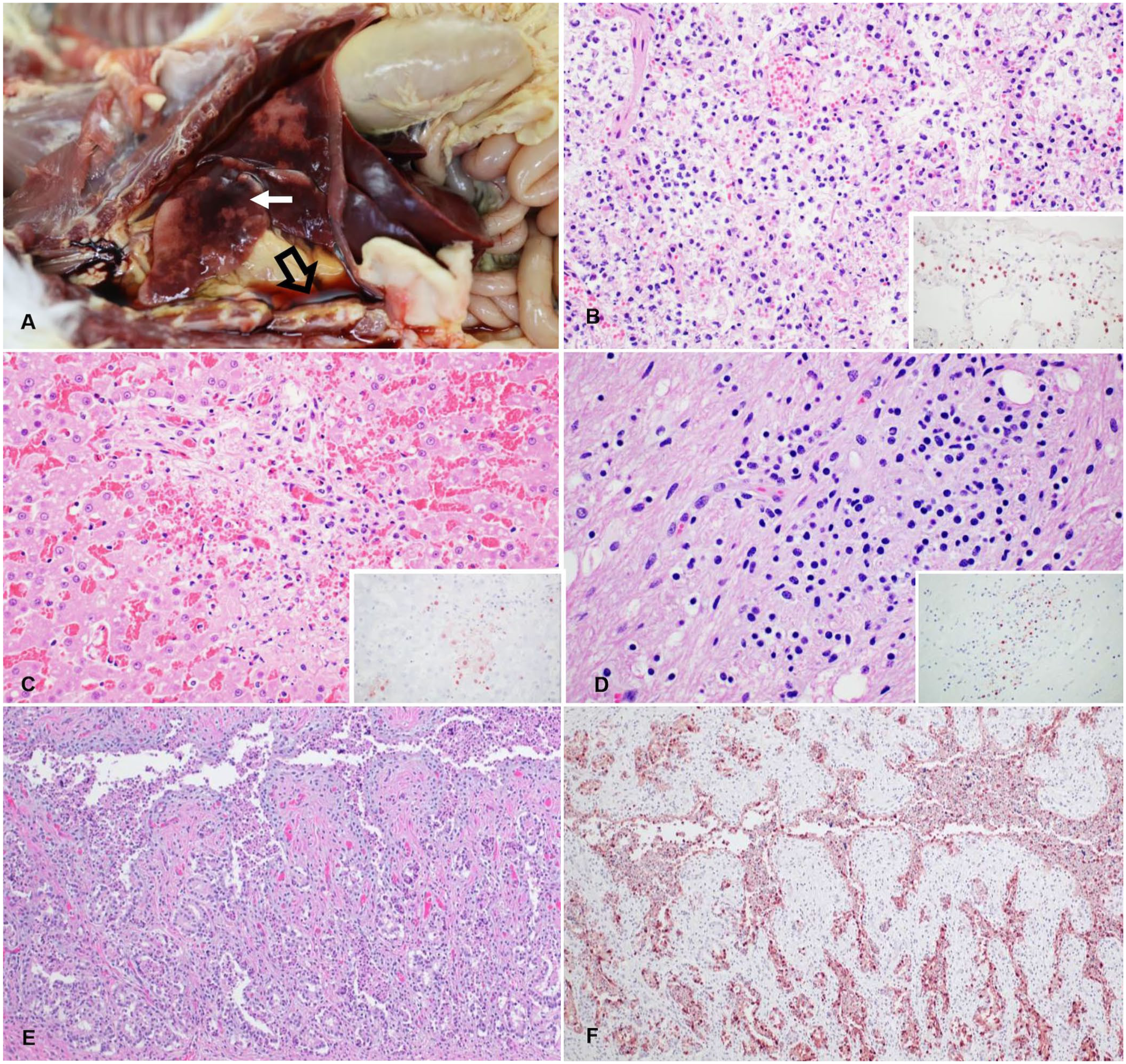
Gross and microscopic changes in a cat infected with highly pathogenic avian influenza A(H5N1) virus after consuming recalled raw milk.
Samples of brain, kidney, lung, liver, lymph nodes, spleen, trachea, larynx, tonsil, thyroid gland, thymus, ovary, oviduct, stomach, urinary bladder, small and large intestine, esophagus, adrenal gland, heart, and eyeballs were collected and fixed in 10% neutral-buffered formalin for ~24 h. All the tissues were routinely processed, and 4-µm-thick, H&E-stained sections were produced. Briefly, the lesions of severe interstitial pneumonia were multifocal necrosis of alveolar septa; capillary thrombosis; capillaries expanded by lymphocytes and macrophages; and polymerized fibrin, edema, macrophages, neutrophils, and debris within the alveolar lumina (
Location of histologic lesions and immunohistochemistry (IHC) immunoreactivity for influenza A virus (IAV) nucleoprotein in a cat infected with highly pathogenic influenza A(H5N1) virus, results of RT-qPCR for IAV, H5, and H5GsGd subtypes, and H5 2.3.4.4b clade expressed as Ct values in an infected cat after raw milk consumption.
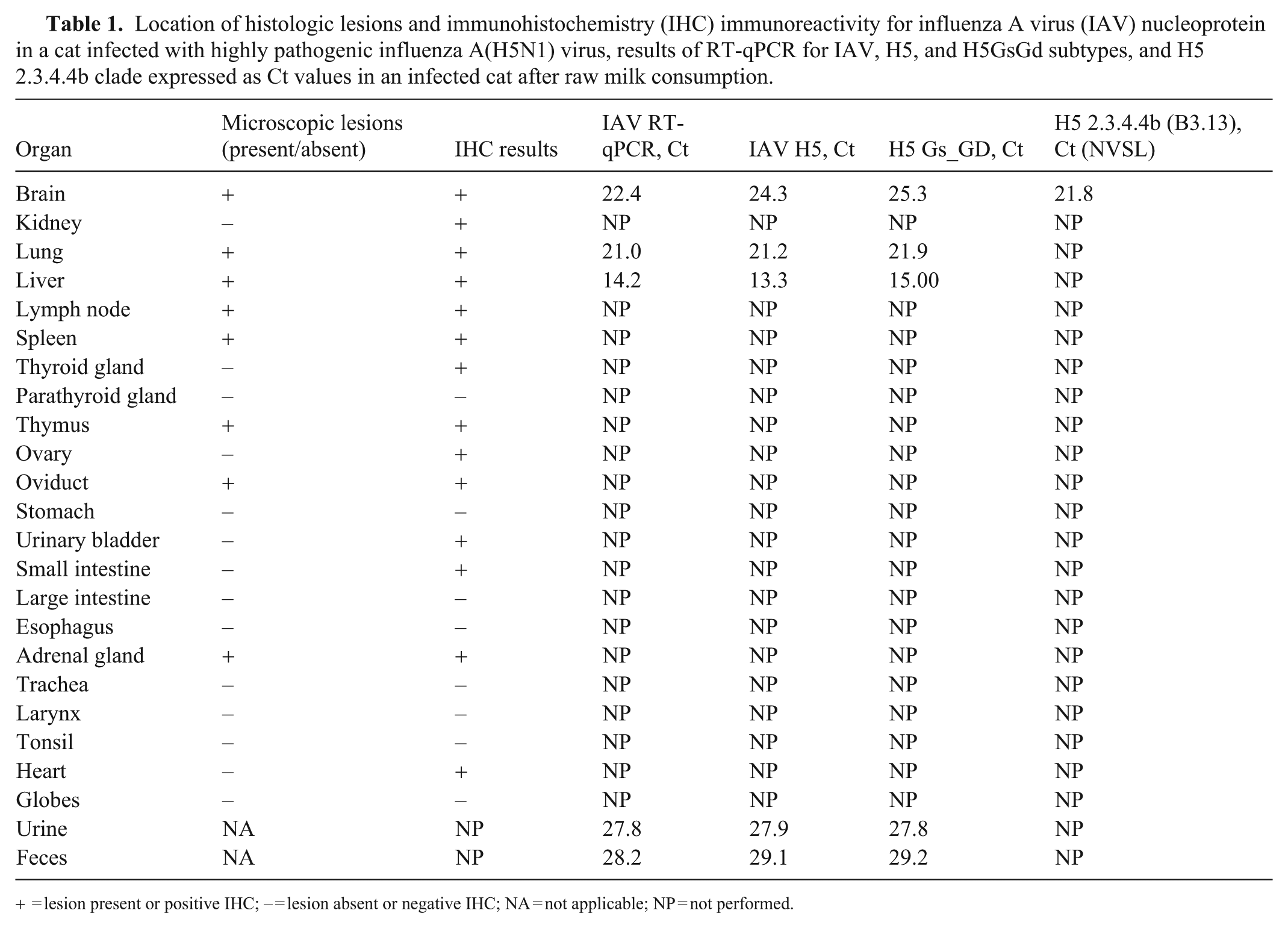
= lesion present or positive IHC; – = lesion absent or negative IHC; NA = not applicable; NP = not performed.
Liver lesions included periportal and centrilobular areas of hepatocellular necrosis with rare lymphocytes, macrophages, and fibrin disrupting the hepatic cords. Some hepatocytes adjacent to the areas of necrosis had vacuolar degeneration (
Necrotizing salpingitis was associated with immunolabeling of the necrotic glandular epithelial cells (
IHC for IAV (mouse monoclonal antibody H16-L10-4R5 [ATCC, https://www.atcc.org]; 1:100, Discovery PSS diluent, Discovery anti-mouse HQ, Discovery anti-HQ HRP) was performed on sections of brain, kidney, lung, liver, lymph node, spleen, thyroid gland, parathyroid gland, thymus, ovary, oviduct, stomach, urinary bladder, small intestine, large intestine, esophagus, adrenal gland, trachea, larynx, tonsil, heart, and eyes, as described. 9 IAV antigen was detected by IHC in multiple tissues (Table 1). Briefly, immunolabeling was noted in the nucleus and cytoplasm of some neurons, in foci of gliosis in the cortex of the brain, in unidentified necrotic cells in the alveolar spaces and alveolar septa of the lungs, in necrotic lymphocytes in lymph nodes, in mononuclear cells in the spleen, in necrotic epithelial cells in oviduct glands, in necrotic hepatocytes, and in necrotic cortical cells in the adrenal gland.
Immunolabeling was also observed—with no associated significant microscopic abnormalities—in some follicular cells in the thyroid glands, endothelial cells of interstitial vessels and mesangial cells in the renal glomeruli, cardiomyocytes, epithelial cells in the thymus, superficial germinal epithelium of the ovary, mesothelial cells of the urinary bladder, and, cytoplasmic immunolabeling, in a few mononuclear cells in the lamina propria of the intestine. We did not observe microscopic ocular lesions or immunolabeling associated with HPAI A(H5N1) virus in our case, although such lesions have been reported in infected cats. 36
Transmission electron microscopy (TEM) was performed on fresh and formalin-fixed tissue (Table 1). 4 Samples from lung, liver, brain, urine, and feces were extracted, cleaned, concentrated, and negatively contrasted using phosphotungstic acid. For ultrastructure of virus assembly and replication in tissue, 1–2 mm3 cubes of trachea, lung, liver, kidney, adrenal gland, and brain were post-fixed in 0.1 M cacodylate-buffered 2.5% glutaraldehyde and 1% osmium tetroxide (Electron Microscopy Sciences). Tissue fragments were embedded in resin in an automatic tissue processor, and 60–90-nm ultrathin sections were stained with toluidine blue, examined to identify the regions of interest, transferred to a copper grid, and examined with a transmission electron microscope (1400; JEOL). All tissues had mild-to-moderate postmortem autolysis and preservation artifacts.
In TEM of the lung, acute, severe, alveolar damage was noted, with necrosis of pneumocytes and denudation of the basement membrane; fibrin replaced pneumocytes and was deposited over the basement membrane; alveolar spaces were filled with pneumocyte debris, erythrocytes, and fibrin (Fig. 2). Both direct and embedding-TEM results were negative, with no viral particles or viral assembly found. TEM has limitations or low sensitivity when postmortem artifacts (as in this case) or extensive tissue processing 8 are present, increasing the risk that target elements might be misidentified as viral factories. In addition, viral particles in tissues can have some degree of similarity with normal intracellular structures. 7

Ultrastructure of lung lesions in a cat infected with highly pathogenic avian influenza A(H5N1) virus.
In our case, HPAI A(H5N1) virus infection was confirmed with severe lesions in several organs. Similar necrotizing lesions have been described in cats infected with HPAI A(H5N1) virus, 42 and in cats infected experimentally and naturally with HPIAV.30,41 Several cats in the cluster of 7, including this cat, were icteric, which has not been reported previously in cats with influenza. Random hepatocellular necrosis was reported in 3 cats infected with HPIAV, but with no evidence of icterus. 30 Given the severity of the liver lesions and elevated liver enzyme activities in our case, associated with intralesional IAV antigen detected by IHC, it is likely that liver failure occurred because of HPAI A(H5N1) virus infection. This virus should be considered a potential cause of acute liver failure in cats.
Based on the IHC results, we suggest that the HPAI A(H5N1) virus replicated and assembled in neurons and glial cells; mesangial cells of the glomeruli; mononuclear cells in the lung, intestines, lymph node, and spleen; parafollicular cells of the thyroid; epithelial cells of the thymus; superficial germinal epithelium of the ovary; mesothelial cells of the urinary bladder; cortical cells of the adrenal glands, oviduct gland epithelium, hepatocytes, and cardiomyocytes.
Retroviral and FPV infections in cats are associated with severe immunodeficiency that may predispose to HPIAV infection.30,31 Sections of intestine were processed for FPV IHC using a mouse monoclonal antibody (MCA2064, 1:500 dilution for 60 min; Bio-Rad), as described previously. 21 No immunolabeling for FPV was noted in the intestine nor detected by a SNAP antigen test at the veterinary clinic.
Reported clinical signs of HPAI A(H5N1) virus infection in domestic cats are variable. Many infected cats have neurologic signs, including trismus, epileptic seizures, depression, inappetence, and limb stiffness.20,36 Several reports of HPAI A(H5N1) virus infections in domestic cats describe mainly mononuclear and necrotizing encephalitis. 36 In our case, no neurologic clinical signs were observed, despite mild gliosis being observed in the white matter of the brain and IAV being detected in the brain by IHC, which confirms virus replication in this organ. This absence of significant neurologic clinical signs and lesions is in contrast with previous reports that describe mononuclear encephalitis as a frequent lesion of influenza in cats. 36 The clinical course in our case involved rapid decline; with more time, neurologic signs might have developed. The main target organ in our case was the liver rather than the CNS.
The salpingitis observed in our case suggests that the virus may be able to cause reproductive harm (which may have been more evident had this cat been pregnant), with abortion as a clinical sign. Future investigations are required to evaluate the prevalence of the virus in the reproductive tract in mammals, and its potential sexual transmission. Although such transmission has not been reported in animals or humans, high-risk infection via artificial insemination has been suggested in turkeys. 11 We encourage evaluation of the reproductive tissues in suspected cases of HPAI A(H5N1) virus infection and recommend considering this virus in the differential diagnosis when the etiologic agent has not been elucidated completely.
Necrotizing interstitial pneumonia is frequent in cases of influenza in felids and plays an important role in pathogenesis.9,30,42 The lesions in the lungs of our case were most likely associated with the viral infection rather the aspiration of the fluid reported by the clinician, given that we did not observe foreign intra-alveolar material in H&E sections. In addition, chorioretinitis, sialadenitis, and myocarditis have been described in this viral infection. 36 Multifocally, the cardiomyocytes in our cat had strong immunolabeling for IAV. However, we noted no lesions in H&E sections, suggesting that microscopic lesions may not have had sufficient time to develop.
PCR Ct values provided semiquantitative information on viral loads in different samples using a common cut-off value (usually ~30).
6
Samples of feces, urine, and swabs of brain, lung, trachea, and liver were collected for IAV reverse-transcription quantitative real-time PCR (RT-qPCR). PCR testing for IAV was performed using the protocol provided by the National Animal Health Laboratory Network (
The lesions and the Ct values obtained in the organs of this cat were correlated. The Ct values of the H5 2.3.4.4b in our case were >21 in the brain, which is similar to previous reports. 36 This result was observed despite the mild lesions that were associated with the positive IHC. In other tissues, the Ct values of IAV RT-qPCR were 14–28, the IAV H5 Ct values were 13–19, and the H5 Gs_GD Ct values were 14–29. The liver was the organ with the lowest Ct value (13) in all the PCR assays performed (IAV PCR test, IAV H5, H5 Gs_GD). The liver also was one of the most affected organs histologically, with strong immunolabeling on IHC in the necrotic areas. The lung had a Ct of 21 and had immunolabeling on IHC in the affected areas. Additionally, the virus was detected with a Ct value >27 in urine and 28 in feces (Table 1).
HPIAV has been reported in rectal swabs from some cats, 41 but not from others. 36 In our case, PCR testing confirmed that the virus was in the feces and small intestine (where some round cells also had positive immunolabeling for IAV by IHC). However, we did not observe necrosis or associated lesions. This finding highlights the importance of considering possible environmental contamination and transmission to other animal species that are in close contact with this type of waste.
Viable virus in milk from infected cows raised concerns about the risk of transmission of the virus via milk. Cow’s milk containing HPAI A(H5N1) virus has been shown to produce illness in experimental mice. 28 However, strong evidence suggests that pasteurization efficiently inactivates HPAI A(H5N1) virus, making dairy products derived from treated milk safe for conusmption.2,45 However, in the United States, raw milk and its derived products are legal to sell to the general public in multiple states, including California. In December 2024 in California, several batches of raw milk sold commercially were recalled after HPAI A(H5N1) virus was detected first in raw milk sold in retail establishments, and later in bulk milk tanks. 10 IAV can remain infectious from 5 d up to >5 wk in raw milk kept at 4°C.28,53 Consumption of raw milk can, therefore, carry the risk of transmission of HPAI A(H5N1) virus. 14
The cat in our case was positive for HPAI A(H5N1) after consuming raw milk shortly before it was recalled because it was found to be infected with HPAI A(H5N1) virus. The milk was not available for testing. The consumption of raw milk presents a health risk not only to cats, but also to people because it can harbor multiple pathogenic microorganisms, including IAV. 49
Several reports have described fatal HPAI A(H5N1) virus infection in cats after they consumed raw pet food products containing poultry, such as duck, turkey, or chicken, that contained the virus. 3 Reports also describe indoor cats infected with HPIAV that lived in households of dairy workers. Those cats had not consumed any unpasteurized milk or milk products and were infected presumably by close contact with the workers or fomites (such as contaminated clothing).38,40 Carnivores, particularly if exposed to HPIAV-infected material or fomites or aerosols, could become infected. 23 Susceptibility to illness could be associated with factors such as malnutrition, immunosuppression, and the D701N amino acid mutation that enhances the replicative adaptation or spillover of the virus in mammalian cells.23,47
In addition, the tissue tropism of IAV is associated with the ability of the virus HA to bind to host sialic acid (SA) receptors such as α-2,3Gal and α-2,6Gal. 18 Although these receptors are present in multiple organs in cats, 51 a complete full-body mapping, including the reproductive organs, has not been reported. Such mapping is crucial to understand the mechanisms of disease of this virus in different organs and will be essential for evaluating the tissue tropism and the routes of infection in felids.
Footnotes
Acknowledgements
We thank Juliann Beingesser and Jose Constante (CAHFS San Bernardino, UCDavis) for their excellent technical assistance.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.