
Abstract
In April–June 2017, a cluster of 4 third-trimester abortions caused by Coxiella burnetii was diagnosed on a dairy farm with 356 Holstein cows in Uruguay. We investigated 1) the seroprevalence of C. burnetii in cows delivering normal calves in this herd, 2) whether non-aborting cows gave birth to seropositive and/or PCR-positive newborn calves in the year of the abortion cluster, and 3) whether seronegative newborn calves seroconverted after ingesting colostrum replacer. In March–September 2017, serum samples were collected from 133 calving cows and their calves immediately after birth and before colostrum ingestion. All 266 sera were analyzed for anti–C. burnetii IgG by ELISA. Additionally, 179 blood samples from some of these animals (82 cows, 97 calves) were PCR-tested for C. burnetii DNA. Thirty of the 133 (22.5%) cows delivering normal calves were seropositive at parturition; 4 of 82 (4.9%) were PCR-positive. All calves tested at birth by ELISA and PCR were negative. Our results suggest that congenital infection is not an important route of C. burnetii transmission to live-born calves in this herd. We observed that 76 of 80 (95%) seronegative newborn calves in 2021 became seropositive for anti–C. burnetii IgG at 24-h-old after drinking an imported commercial colostrum replacer, which could confound seroepidemiologic surveys in herds using colostrum replacers.
Keywords
Coxiella burnetii, a gram-negative bacterium, is the causative agent of the globally important zoonosis Q fever.
15
Despite the existence of diverse reservoirs, domestic ruminants are by far the main source of C. burnetii infection for humans.2,15 Clinically, C. burnetii infection leads to the abortion, premature delivery, stillbirth, and weak offspring (
Although the airborne route is the main mode of transmission for C. burnetii in animals and humans, little is known about other transmission pathways in cattle. 2 The existence of transplacental transmission of C. burnetii is still debated. In murine models, the fetoplacental junction resisted C. burnetii transmission, with vertical infection typically occurring via aerosol inhalation during parturition. 3 Intrauterine infection of calves with C. burnetii was suggested in 1952, although postnatal exposure through inhalation of maternal secretions during or shortly after parturition could not be ruled out, given the experimental design used in that study. 22 The role of calves as carriers of this bacterium remains largely unknown.
C. burnetii invades the placenta and can cause placental inflammation and necrosis, leading to abortion. 11 C. burnetii can also colonize trophoblasts and use them as a protective replicative niche without necessarily interfering with pregnancy.4,11 In ruminants, which have a synepitheliochorial placenta with 6 layers separating maternal and fetal blood, C. burnetii may remain confined to the placenta, or eventually reach the fetus following amniotic-oral or amniotic-respiratory routes, leading to bacterial detection in the gastrointestinal tract and lungs.1,5 Fetal infection may also occur hematogenously through the umbilical blood vessels, leading to systemic bacteremia. 1 Perinatal infection also may eventually occur via the inhalation of contaminated aerosols generated during or immediately after calving. Distinguishing between in utero and perinatal C. burnetii infection is challenging, especially under field conditions. In a case-control study, 9 4 of 56 blood samples from stillborn calves were PCR positive, and 1 of 56 samples was positive for anti–C. burnetii phase II antibodies; neither DNA nor antibodies against C. burnetii were detected in 30 calves born alive.
For effective passive transfer of immunoglobulins, colostrum replacers are an alternative for the first feeding of neonatal calves when maternal colostrum is unavailable. Commercial products are powders that are usually free of pathogens and other infectious agents and are reconstituted to fluid form before use.
In 2019, we reported a cluster of 4 bovine abortions caused by C. burnetii that occurred during the third trimester of pregnancy (diagnosed in Apr–Jun 2017 in a dairy farm in Uruguay). 14 We conducted serologic and molecular evaluations of dam-calf pairs at calving from this dairy herd, 14 which also involved C. burnetii exposure among farm workers and laboratory technicians. 18
We subsequently evaluated cows and calves from the same herd of 356 lactating and dry Holstein cows on this farm endemically infected with C. burnetii. Sampling procedures were approved by the Instituto Nacional de Investigación Agropecuaria, Ethics Committee on the Use of Animals (protocols 2017.1, 2019.5, 2019.9). This herd had seasonal calving, was free of Brucella abortus, and was in a region free of Rhipicephalus microplus ticks. No vaccination against C. burnetii had been implemented. We investigated: 1) the seroprevalence of C. burnetii in cows delivering normal calves in this herd, 2) whether non-aborting cows gave birth to seropositive and/or PCR-positive newborn calves in the year of the abortion cluster, and 3) whether seronegative newborn calves seroconverted after ingesting colostrum replacer.
To assess vertical transmission and exposure, in March–September 2017, blood samples were collected from 133 cows at calving selected by convenience sampling, as well as from their 133 calves before colostrum intake. Sera were harvested for the detection of anti–C. burnetii IgG by ELISA. Concurrently, whole blood from 82 of the cows and 97 of the calves was tested with a PCR assay for C. burnetii detection.
We used a commercial indirect ELISA kit (PrioCHECK ruminant Q fever Ab plate kit; ThermoFisher), manufactured with a mix of phase I and II antigens from a C. burnetii sheep strain isolated in France, to detect anti–C. burnetii total IgG antibodies in serum, following the manufacturer’s instructions. Optical densities (ODs) were assessed with a microplate ELISA reader (Multiskan FC; ThermoFisher) at 450 nm wavelength. Positive and negative controls provided with the kit were included in each ELISA plate. The antibody levels were estimated as the sample:positive (S:P) ratio calculated using the formula S:P = (ODSample – ODNC)/(ODPC – ODNC), where ODSample is the OD of the well containing the sample, and ODNC and ODPC are the mean ODs of 2 replicate negative and positive control wells, respectively. The samples were classified as negative for S:P ≤40% or positive for S:P >40% and further subclassified on an ordinal scale as + = S:P >40–100%, ++ = >100–200%, +++ = >200–300%, and ++++ = >300%.
Total DNA was extracted from the 179 whole blood samples (97 calves, 82 cows) collected in 2017 (MagMAX pathogen RNA/DNA kit; ThermoFisher). DNA quality and quantity were assessed (NanoDrop ND-1000 spectrophotometer; ThermoFisher). C. burnetii detection targeted the multicopy IS1111 sequence using the β-actin gene as an internal control ( Table 1).
Details of the real-time PCR procedure for DNA amplification of the Coxiella burnetii multicopy IS1111 gene and the bovine β-actin gene.
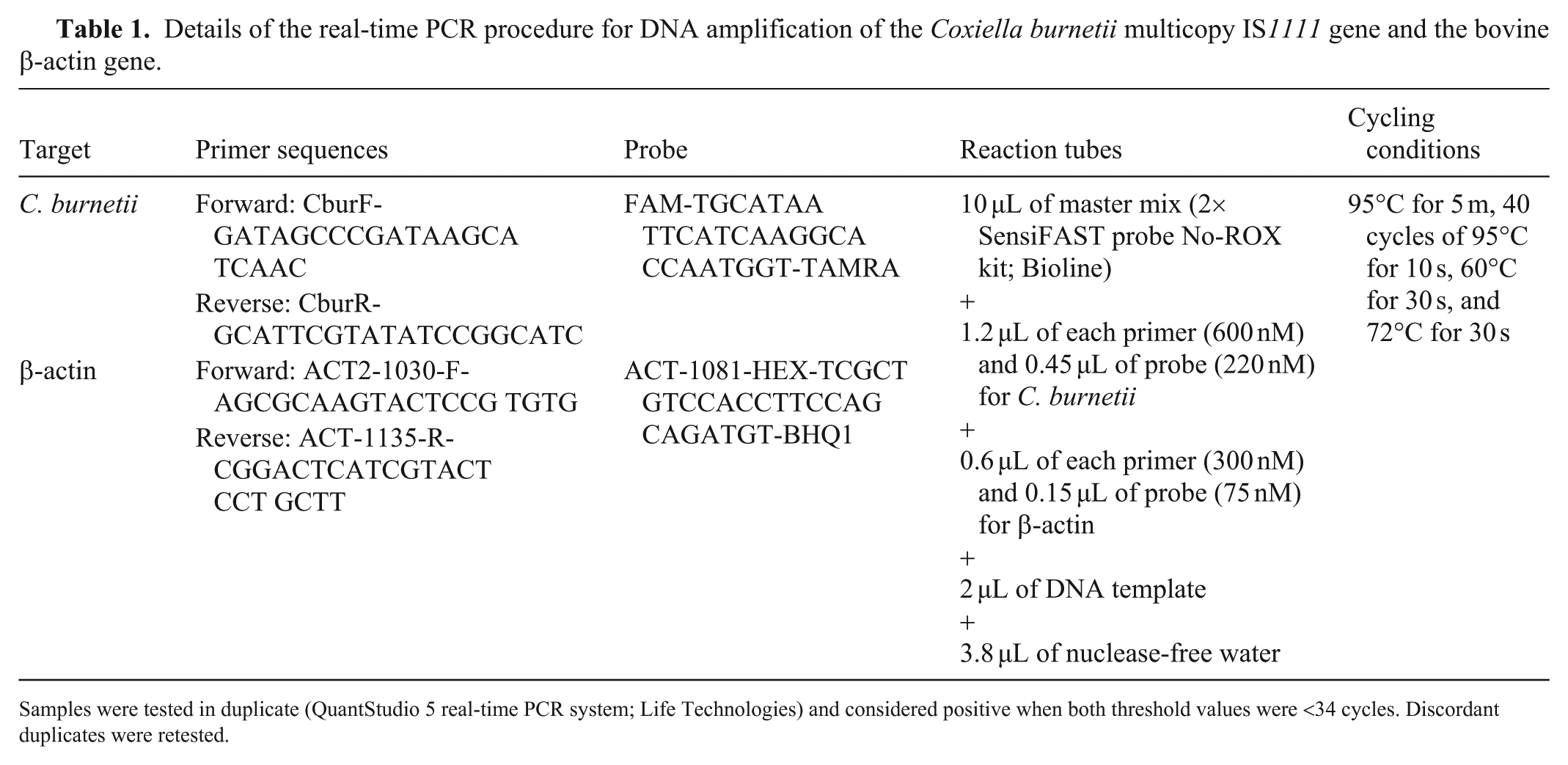
Samples were tested in duplicate (QuantStudio 5 real-time PCR system; Life Technologies) and considered positive when both threshold values were <34 cycles. Discordant duplicates were retested.
To evaluate the passive transfer of anti–C. burnetii IgG from imported colostrum replacer in neonatal calves, serum samples were collected from 80 live-born calves from the same herd in March–June 2021, immediately after birth and before colostrum replacer ingestion. All these calves were artificially fed a commercial colostrum replacer (Calf’s Choice Total, Saskatoon Colostrum Company, Canada) following the manufacturer’s recommendation, within 2 h of birth; serum samples were collected again when 24-h-old. All 160 sera were processed by ELISA for anti–C. burnetii IgG detection, as described above.
Among the 133 cows that calved in March–September 2017, 30 tested positive for C. burnetii antibodies; 4 of 82 were PCR-positive (
ELISA and PCR results for Coxiella burnetii in 133 cows at calving and their newborn calves before colostrum ingestion.
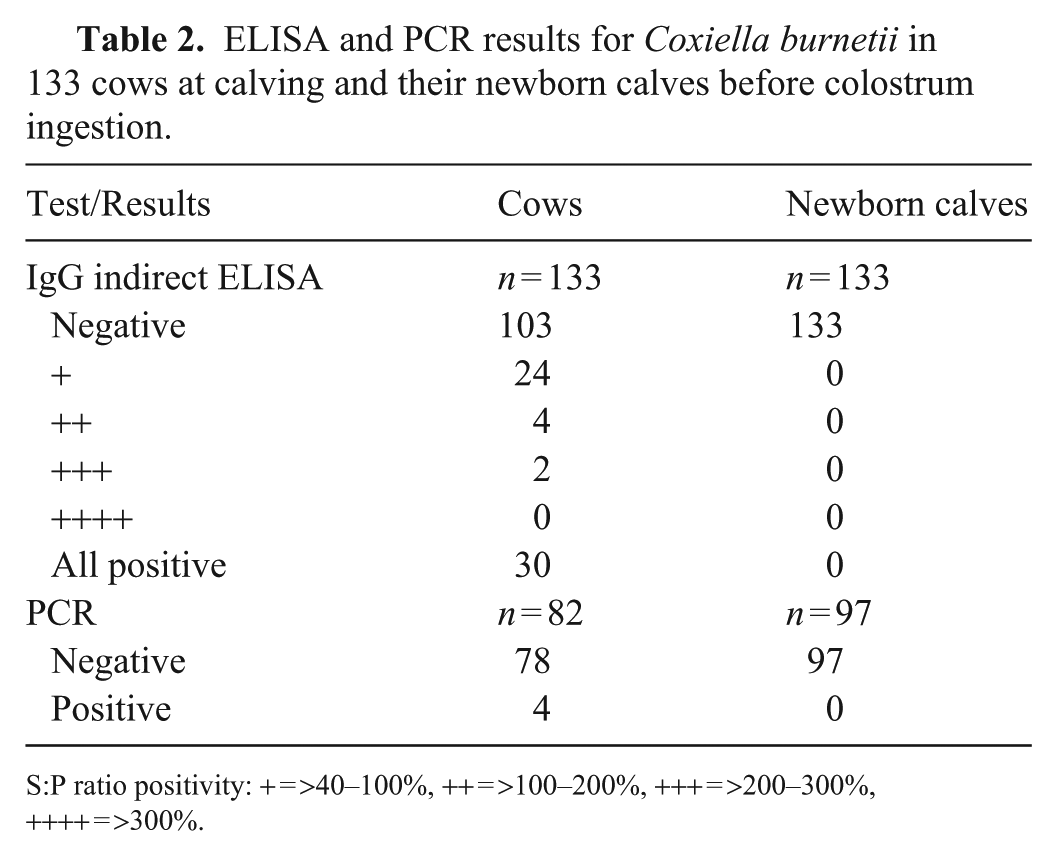
S:P ratio positivity: + = >40–100%, ++ = >100–200%, +++ = >200–300%, ++++ = >300%.
After ingestion of commercial colostrum replacer, 76 of 80 (95%) newborn calves sampled in 2021 that were seronegative at birth had detectable specific anti–C. burnetii IgG in serum 24 h later (
ELISA results for anti–Coxiella burnetii IgG in 80 newborn calves immediately after birth before colostrum intake (pre-colostrum), and the same calves when 24-h-old after ingestion of an imported colostrum replacer (post-colostrum).
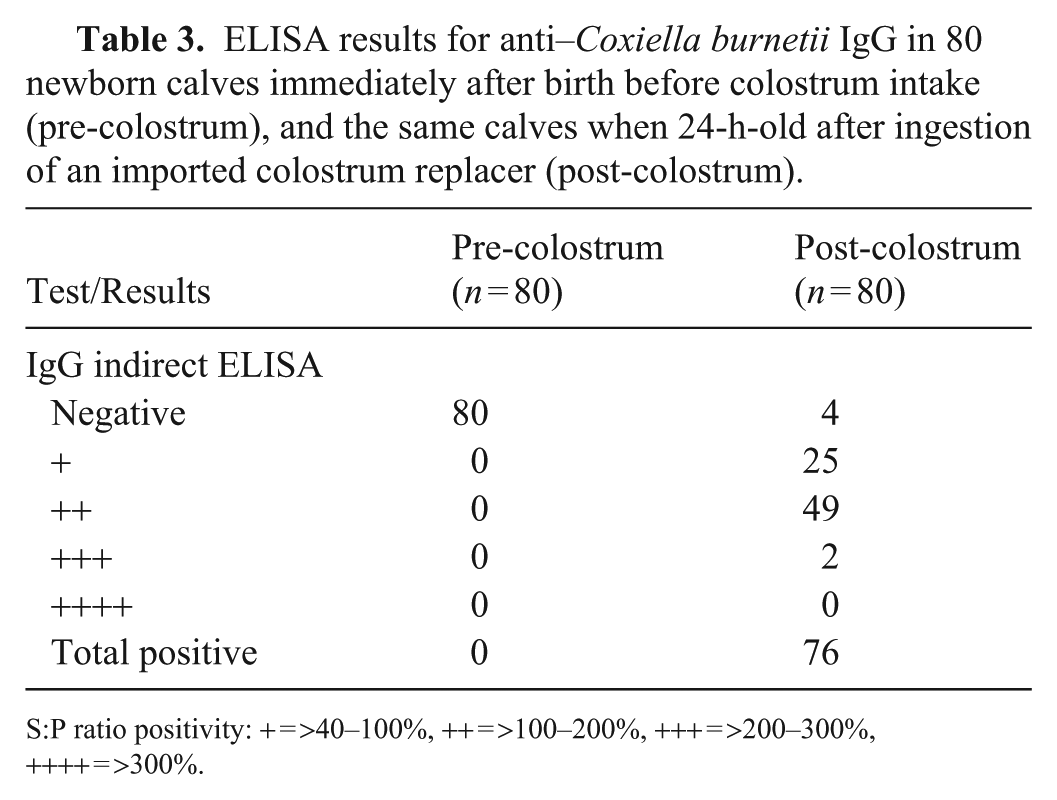
S:P ratio positivity: + = >40–100%, ++ = >100–200%, +++ = >200–300%, ++++ = >300%.
When C. burnetii infection leads to the APSW complex, the fetal immune response is eventually activated. 1 If the placenta has trophoblasts heavily loaded with C. burnetii, the likelihood of transplacental infection increases, leading to detectable antibodies in the fetus, as shown in stillborn calves. 8 Conversely, lower—possibly dormant—C. burnetii placental loads may support full-term pregnancies, with infection restricted to the placenta, no fetal infection, and no development of a fetal immune response. 1 Similarly, others have reported this outcome even in cows with cotyledons confirmed C. burnetii–positive by PCR testing.20,21 Consistent with this, we observed that C. burnetii seropositive cows gave birth to seronegative calves, further supporting that exposure of the dam does not necessarily result in transplacental transmission or fetal seroconversion.
Placental immunosuppression with IL10 production and dendritic cells silencing, essential for fetal tolerance, could also favor evasion of immune clearance by C. burnetii and its persistence within placental tissues.4,10 This placental immune environment can shape the progression of infection and affect pregnancy outcomes.
Previous investigations suggested that the absence of pre-colostral anti–C. burnetii antibodies in healthy calves in infected herds could be the result of resistance of the fetoplacental unit to vertical transmission,3,20 or possibly because of the development of immunotolerance after early C. burnetii exposure in utero, as seen with other pathogens (e.g., bovine viral diarrhea virus). 16 Elucidating whether immunotolerance to C. burnetii exists could significantly impact the epidemiology of coxiellosis, as seronegative shedders infected in utero could be undetectable using conventional serologic methods. 19 If C. burnetii infection occurs early during gestation, when fetal immunocompetence is underdeveloped or when immune tolerance is high, the offspring may fail to recognize the bacterium as an exogenous antigen. It remains to be determined whether the outcome of C. burnetii infection during pregnancy depends on its intrauterine dynamics, and whether pregnancy outcome is influenced by infection severity and the gestational stage at the time of infection.
Although cows in our study were enrolled at parturition using non-randomized sampling, this strategy may still provide a fair estimate of the seroprevalence of C. burnetii in the population of pregnant, non-aborting cows in this herd in 2017, given that we assessed 133 of 356 (37%) cows. All 213 calves sampled in 2017 (n = 133) and in 2021 (n = 80) were born seronegative for C. burnetii, and 76 of the 80 (95%) calves that received the imported commercial colostrum replacer in 2021 turned seropositive when 24-h-old. In all these calves, anti–C. burnetii antibodies were acquired passively through the colostrum. 19 The serologic status of the dams of calves sampled in 2021 were not evaluated at calving; however, 9 of 80 (11%) and 27 of 80 (34%) were seropositive for C. burnetii in 2020 and November 2021, respectively (data not shown). Previous studies showed that calves with maternal C. burnetii antibodies remained seropositive for the first 30 d of life, then titers started to decrease gradually until 90–103 d of life, by which time all calves were seronegative.8,21 The negative results for C. burnetii DNA in pre-colostral blood samples in the calves born in 2017 mitigate the possibility of persistent infection resulting from immunotolerance induced by early in utero exposure, which otherwise results in infection without seroconversion.
We detected total anti–C. burnetii IgG using an ELISA based on a mix of phase I and II antigens. However, ELISAs employing mixed-phase antigens may have limited sensitivity for detecting phase II anti–C. burnetii-specific antibodies, potentially leading to under-detection of recent infections. 7 This potential limitation could have been mitigated by concurrent use of PCR testing, enabling highly sensitive detection of bacterial DNA. Regarding the performance of the commercial kit used in our study, validations conducted by the Institut National de la Recherche Agronomique and Life Technologies reported a sensitivity of 87% and specificity of 100%. 6 A subsequent evaluation using cattle sera estimated a similar specificity of 97%, but a much lower sensitivity of 61%. 12
The colostrum replacer used in 2021 was made from natural bovine colostrum sourced in Scotland (https://thecalfcompany.com/product/sccl-calf-choice-total-bovine-colostrum/), where C. burnetii is endemic in dairy cattle. 17 The manufacturer does not publish details of the production process. We found that, as expected, anti–C. burnetii IgG antibodies in the imported colostrum replacer withstand the production process—presumably involving freeze- or spray-drying methods—retaining the antigen-binding capacity to produce detectable signal in the ELISA. Positive serologic results for bovine leukemia virus (BLV) antibodies have been reported in calves fed the same colostrum replacer, 13 and anti-BLV antibodies were detected in calves fed this colostrum in our study (data not shown; Caroline da Silva Silveira, pers. comm., 2025). Our findings raise concern about the potential interference of colostrum replacer antibodies in seroprevalence studies for different pathogens, particularly in young animals.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Our study was funded by grants PL_27 from the Instituto Nacional de Investigación Agropecuaria (INIA), by grant FMV_1_2019_1_155861 from the Agencia Nacional de Investigación e Innovación (ANII), by funds from Unidad Mixta Pasteur + INIA (UMPI) and Fondo para la Convergencia Estructural del Mercosur (COF 03/11), Institut Pasteur de Montevideo, Uruguay. A. Rabaza, C. da Silva Silveira, D. Caffarena, A. Mendoza, M. Fraga, and F. Giannitti acknowledge support from ANII through the “Sistema Nacional de Investigadores” (SNI). A. Rabaza, C. da Silva Silveira, M. Fraga, and L. Zarantonelli are researchers of the “Programa de Desarrollo de las Ciencias Básicas” (PEDECIBA).