
Abstract
Mycobacterium avium subsp. paratuberculosis (MAP) prevalence estimates vary widely across Colombian regions, with no prior longitudinal assessment of Colombian dairy systems across ecologic life zones. We assessed the temporal dynamics of MAP infection in Colombian cattle across 6 distinct ecologic life zones and explored age-related differences in MAP detection using combined serologic and molecular approaches. We longitudinally sampled 129 cattle from 6 municipalities in distinct Holdridge life zones. The animals were stratified by age group and sampled 3 times at 2-mo intervals during 2023 (Aug–Dec). Serum and fecal samples were tested by indirect ELISA and IS900 real-time PCR (rtPCR), respectively. The apparent ELISA prevalence remained stable between 37.2% and 38% across sampling rounds (adjusted true prevalence = 29.9%). Seropositivity increased consistently with age, from 11.9% in animals <1-mo-old to 49% in animals ≥3-y-old. All of the fecal samples tested negative by rtPCR. No consistent seroprevalence clustering was observed across ecologic life zones on descriptive assessment, although management and host factors were not evaluated. We found longitudinal evidence of stable, age-related MAP seroprevalence across ecologic zones in Colombia, highlighting subclinical persistence and diagnostic limitations.
Keywords
Mycobacterium avium subsp. paratuberculosis (
MAP is enzootic in more than half of all European countries, with herd prevalence varying widely (10–80%), depending on the production system and test used.7,8 In North America, herd-level infection rates are estimated to be 68–91%.2,14 In South America and the Caribbean, PTB incidence ranges from 18.7–100% based on the test employed, including ELISA, fecal culture, skin testing, and PCR. 10
In Colombia, PTB was first reported in 1924. Since then, prevalence studies have found marked heterogeneity across regions, production systems, and tests. 6 Herd-level seroprevalence estimates in Colombia via ELISA range from 3.6% in specialized dairy herds from northern Antioquia 3 to >50% in southern Nariño 1 and the eastern plains of Villavicencio. 11 When molecular methods have been applied, environmental qPCR surveys have revealed herd-level prevalence rates of 4.1–14.9% in Antioquia.4,5 Such variability likely reflects differences in management intensity, ecologic conditions, and test methodology. Importantly, all prior longitudinal studies have been restricted to a single farm,9,13 highlighting the absence of multiregional, time-based assessments.
To address this variability, we evaluated temporal MAP infection dynamics via a longitudinal design combining indirect ELISA and real-time PCR (rtPCR) across 6 ecologic life zones in Colombia. We conducted a prospective longitudinal study via nonprobabilistic convenience sampling of 129 cattle from 6 Colombian municipalities, representing distinct Holdridge life zones (
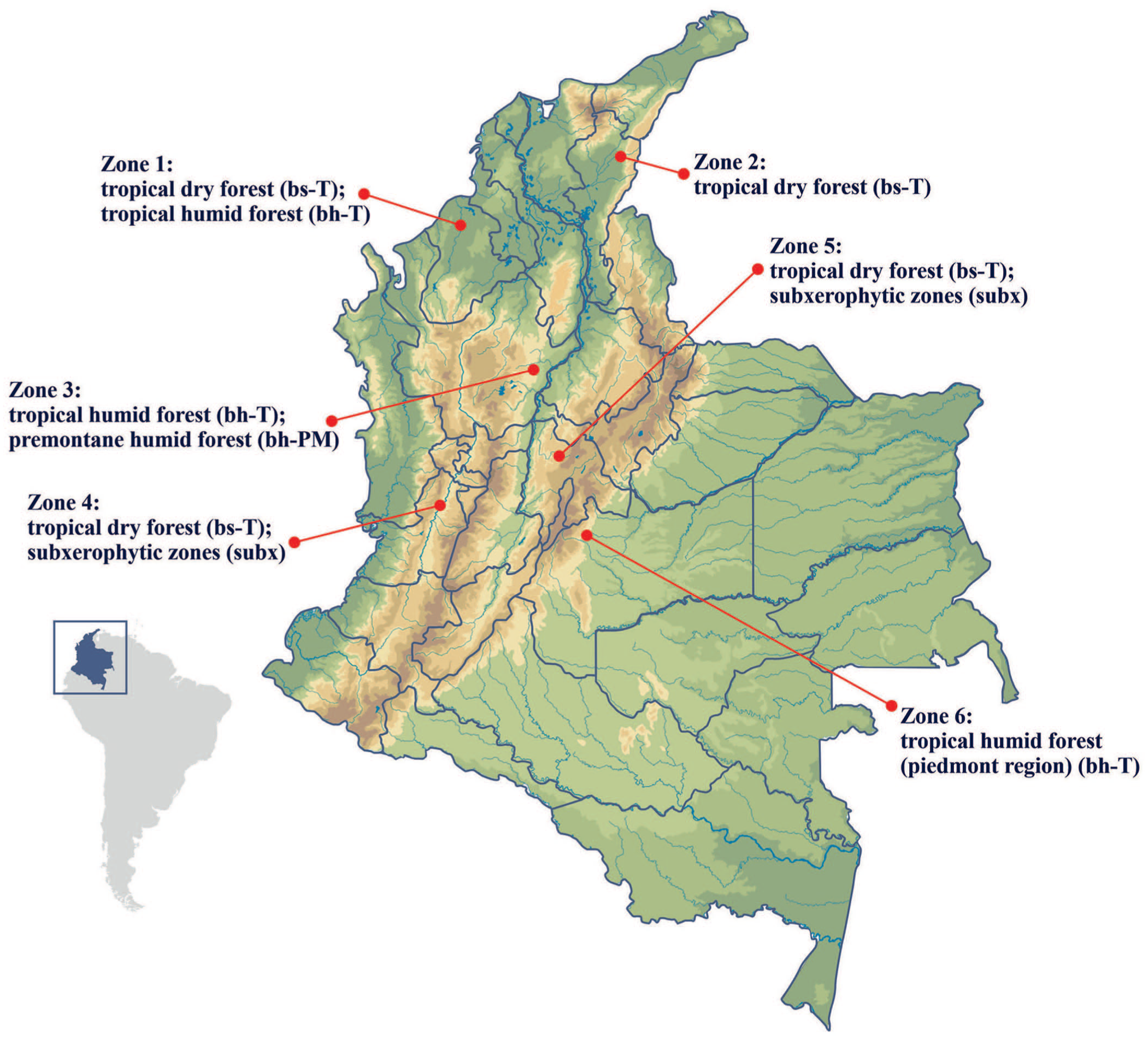
Locations of the 6 distinct life zones studied in Colombia. Subx = subxerophytic, which is not a formal Holdridge life zone code but rather a descriptive term for dry transitional areas with semi-arid characteristics.
Herd-level identifiers were unavailable; therefore, each animal was treated as the unit of analysis, with ecologic classification assigned by the municipality of origin. The ecologic classification followed the HLZ system defined by the Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM; Bogotá, Colombia, https://www.ideam.gov.co/ecosistemas?utm), which stratifies Colombia based on the mean annual biotemperature, precipitation, and potential evapotranspiration ratios. The HLZ categories considered in our study included tropical dry forest (bs-T), tropical humid forest (bh-T), premontane humid forest (bh-PM), and subxerophytic (subx) transition zones. The updated climatic layers (temperature and precipitation, 1981–2020) from the official IDEAM database were integrated with municipal boundaries via QGIS 3.44.0 “Solothurn” (June 2025). This intersection produced 6 ecologic units that corresponded to the municipalities sampled, allowing standardized ecologic stratification for descriptive comparisons of MAP prevalence across life zones. This ecologic framework offers an environmental context but was not intended as an explanatory variable, as prevalence differences may also reflect herd-level management, biosecurity, and animal density.
In our longitudinal MAP surveillance study, 129 cattle were proportionally allocated to 6 life zones classified by altitude, mean temperature, and vegetation type: zone 1 (tropical dry forest, bs-T; tropical humid forest, bh-T; 18 meters above the sea level (m.a.s.l.); 28°C; n = 31, 24.0%); zone 2 (bs-T; 131 m.a.s.l.; 29°C; n = 22, 17.1%); zone 3 (bh-T; premontane humid forest, bh-PM; 800 m.a.s.l.; 23°C; n = 22, 17.1%); zone 4 (bs-T; subx; 2,516 m.a.s.l.; 16°C; n = 11, 8.5%); zone 5 (bs-T; subx; 1,006 m.a.s.l.; 25°C; n = 32, 24.8%); and zone 6 (bh-T; piedmont; 336 m.a.s.l.; 25°C; n = 11, 8.5%). Within each life zone, animals were further distributed across age groups: <1-mo-old, 1 to <6-mo-old, 1 to <2-y-old, 2 to <3-y-old, and ≥3-y-old ( Table 1 ). These groups were defined based on physiologic and management stages relevant to MAP exposure and immune response. The 1 to <6-mo-old group reflects weaning timing and initial contact with adult fecal material in Colombian dairy systems, when the risk of MAP exposure increases. Breed information was unavailable; however, unique animal identification codes allowed longitudinal monitoring. Each animal was sampled 3 times over 4 mo at 2-mo intervals (2023 Aug, Oct, Dec).
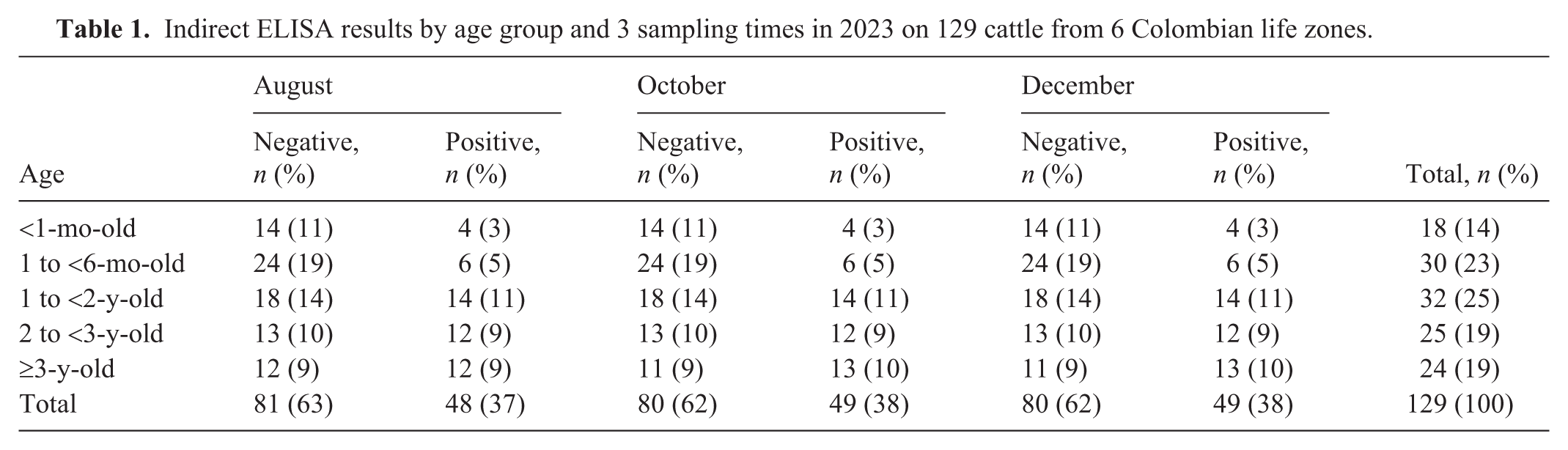
Indirect ELISA results by age group and 3 sampling times in 2023 on 129 cattle from 6 Colombian life zones.
Blood and fecal samples were collected from all the animals at each time via standard procedures to minimize pain and stress. All sampling was conducted as part of routine veterinary testing. Blood was drawn from the coccygeal or jugular vein into EDTA and red-top evacuated tubes (Vacutainer; Becton Dickinson). Fecal samples were obtained directly from the rectum using single-use clean gloves. Both sample types were transported under refrigeration at 4°C. The blood was centrifuged at 1,008 × g for 5 min and the serum was stored at −20°C until processing; fecal samples were maintained at 4°C until analyzed.
Serum indirect ELISA was performed with a commercial kit (INgezim Paratuberculosis; Gold Standard Diagnostics), according to the manufacturer’s instructions. The assay, which is based on direct immunoenzymatic detection of MAP-specific antibodies, includes a preabsorption step with Mycobacterium phlei to reduce cross-reactions. Serum samples were diluted 1:20, and kit-supplemented positive and negative controls were included in each run. Animals were classified as ELISA positive if the S:P ratio was >0.3, negative if <0.2, and inconclusive if 0.2–0.3; inconclusive samples were reanalyzed to confirm their status.
Each fecal sample was homogenized by vortexing for 5 min to ensure a uniform matrix before DNA extraction. To prevent cross-contamination, the samples were processed individually using sterile, single-use materials, aerosol-resistant pipette tips, and standard molecular laboratory practices, including physical separation of pre- and post-PCR workflows and routine surface decontamination.
DNA was isolated via a commercial kit (ZR fecal DNA kit; Zymo Research), following the manufacturer’s protocol for bacterial and yeast nucleic acids. A mechanical disruption step was included using an automated sample lyzer (Disruptor Genie 120 V; Thomas Scientific) to improve cell lysis. DNA yield was assessed (NanoDrop 2000 spectrophotometer; Thermo Scientific) at 260/280 nm, and integrity was confirmed by agarose gel electrophoresis after 10% extraction. The DNA extracted was stored at −20°C until rtPCR analysis.
The AdiaVet Paratb real-time kit (ADI045-100; BioX Diagnostics) was used. The PCR assay targets the IS900 gene with a hydrolysis probe (exonuclease technology), enabling simultaneous detection of MAP (FAM-labeled probe) and an external amplification control (VIC/HEX-like probe). Each 25-µL reaction mixture contained 20 µL of A5 master mix (including Taq DNA polymerase, 1× reaction buffer, 3 mM MgCl2, and 200 µM of each dNTP), 1 µL of EPC-Amp (internal amplification control), 1 µL of EPC-Ext (extraction control), and 5 µL of template DNA (50–100 ng). Positive and negative controls supplied with the kit were included in each run, along with a field control obtained from a MAP-positive bovid previously confirmed by Sanger sequencing and BLAST alignment in the NCBI database. The reaction mixture was homogenized by gentle vortexing and briefly centrifuged to ensure proper mixing and collection of contents at the bottom of the tube. The sample then was dispensed into optical-grade tubes and run on a real-time PCR detection system (CFX96 Touch; Merck). The thermal cycling conditions were 50°C for 2 min and 95°C for 10 min, followed by 45 cycles of 95°C for 15 s and 60°C for 1 min, with fluorescence signals (FAM and HEX channels) acquired at the end of each cycle. The results were analyzed (CFX Maestro software v.2.3; Bio-Rad).
Data were managed in Excel (v.19.5; Microsoft) and analyzed in Stata (v.19.5; StataCorp). Descriptive statistics were calculated for all variables. The true prevalence was estimated using the Rogan–Gladen estimator, adjusting the apparent ELISA prevalence for test sensitivity (98%) and specificity (88%). All prevalence estimates were calculated at the individual animal level.
Of the 129 animals sampled, 48 (37.2%) tested positive by indirect ELISA at the first sampling (Table 1). Two months later, one additional animal was positive (total 49 of 129; 38%), and the same result was maintained in the third sampling. All blood and fecal samples were negative by rtPCR, with positive and negative controls performing appropriately. No animals were lost in the study.
With respect to age group, 4 animals <1-mo-old and 6 animals 1 to <6-mo-old were consistently positive across all samplings. Among animals 1 to <2-y-old, 14 remained positive, and among animals 2 to <3-y-old, 12 were positive in all rounds. Among animals ≥3-y-old, 12 tested positive in the first round, and 13 tested positive in both the second and third rounds (Table 1).
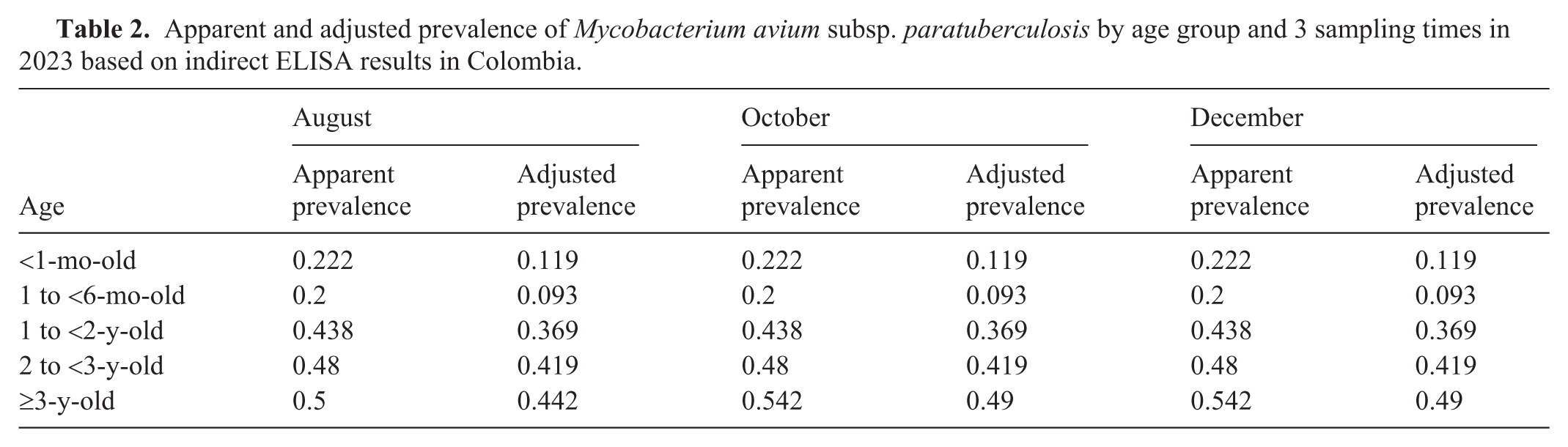
The overall apparent animal-level prevalence across the 3 sampling rounds was 37.7% after the Rogan–Gladen adjustment; the true animal-level prevalence was 29.9% ( Table 2 ). The adjusted prevalence remained low and stable in animals <1-mo-old (11.9%) and 1 to <6-mo-old (9.3%), increasing consistently in older cattle: 36.9% in animals 1 to <2-y-old, 41.9% in animals 2 to <3-y-old, and 49% in animals ≥3-y-old, demonstrating a clear age-related trend across all rounds. Herd-level prevalence could not be estimated; herd identifiers were unavailable because of confidentiality agreements. Seroprevalence did not cluster consistently by ecologic life zone; however, given the nonprobabilistic sampling design and unequal subgroup sizes, formal statistical comparisons were not performed. In Colombia, published herd-level rates range 3.6–50% based on the production system and test method,1,3,11 which is consistent with the 29.9% intermediate prevalence that we found. Given our nonprobabilistic voluntary sampling design, our findings should be interpreted as descriptive rather than inferential. Selection bias cannot be excluded, given that herds with prior PTB suspicion may have been overrepresented. Farm-level confounders—including confounders of biosecurity, animal density, calving management, and milking hygiene—could not be controlled. Future studies incorporating standardized herd management data would allow for better adjustment of these variables.
Apparent and adjusted prevalence of Mycobacterium avium subsp. paratuberculosis by age group and 3 sampling times in 2023 based on indirect ELISA results in Colombia.
All our rtPCR results were negative; internal and external controls performed appropriately. This outcome aligns with the expected low sensitivity of direct molecular detection in subclinical animals, in which MAP shedding is intermittent and bacterial loads are low and remain below the detection threshold.2,16,18 This pattern is consistent with the known pathophysiology of PTB, in which molecular detection has limited sensitivity in subclinical animals. In contrast, ELISA can detect long-lasting antibody responses that may precede or persist during periods of intermittent fecal shedding, which explains the apparent discrepancy between the serologic and molecular findings. The minimal variation in ELISA results across sampling rounds is consistent with the chronic, antibody-stable nature of MAP infection. Seropositivity in animals <1-y-old should be interpreted cautiously, given that positive results may reflect maternal antibody transfer, early environmental exposure, or false-positive reactions rather than active infection. Taken together, our results reinforce the limitations of single-method surveillance in subclinical MAP infection.
HLZ classification provided an ecologic framework for interpreting MAP distribution; however, these zones represent broad categories rather than farm-specific exposures, and climatic variables (such as temperature and humidity) were not formally analyzed. Seroprevalence did not show consistent clustering by life zone, and formal statistical comparisons were not conducted, given our nonprobabilistic design and unequal subgroup sizes. Management-related factors, including hygiene, animal density, feeding practices, and replacement policies, likely have stronger effects on infection dynamics than do ecologic zones alone.
Limitations of our study include convenience sampling, modest subgroup sizes, and the absence of fecal culture as a confirmatory test. The lack of herd-level information—including management practices, herd size, biosecurity measures, and animal movement data—constrained interpretation at each phase of the study, which limited our ability to contextualize infection dynamics or identify management-related risk factors. Information on clinical status was unavailable, which limits our interpretation of infection stage and its relationship to test performance. Moreover, our 4-mo study does not capture seasonal fluctuations in antibody responses or shedding patterns. Our study highlights the need for integrated surveillance systems that combine serologic, molecular, and herd-level management indicators.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Our work was supported by the Ministry of Agriculture and Rural Development of Colombia through Interadministrative agreement MADR334-2023.