
Abstract
Objective:
To examine how emotional awareness (the ability to understand and identify one’s own emotions) and neural reward circuitry, processes that are associated with depression and undergo significant maturation during adolescence, prospectively predict depressive symptoms in a community sample of youth at varying risk for affective disorders.
Method:
Youth aged 13–19 years (N = 84), risk-enriched for depression (66% with familial history of affective disorders), completed self-report assessments of emotional awareness and depressive symptoms at baseline. Depressive symptoms were also assessed at 1-year follow-up. Neural reward function, as indexed by functional connectivity of the dorsomedial prefrontal cortex (dmPFC) and nucleus accumbens (NAcc), was assessed via a functional magnetic resonance imaging task of winning versus losing monetary rewards at baseline. Controlling for age, biological sex, familial risk status, and baseline depression, linear regressions examined the interaction of emotional awareness and dmPFC–NAcc functional connectivity to predict changes in depressive symptoms over 1 year.
Results:
Low emotional awareness at baseline predicted more severe depressive symptoms at 1-year follow-up, particularly among youth demonstrating heightened positive dmPFC–NAcc functional connectivity, a pattern potentially indicating over-regulation of reward responding.
Discussion:
Low emotional awareness and stronger frontostriatal functional connectivity during processing of reward relative to loss are relevant early-emerging risk factors that could jointly lead to future depression. Interventions targeting emotional regulation skills and reward processing may mitigate the onset and course of adolescent depressive symptoms.
Introduction
Adolescence is a developmental period marked by profound changes in emotional and neurocognitive systems that jointly shape vulnerability for psychopathology (Dahl, 2004; Forbes et al., 2021; Somerville et al., 2010). Rates of depression rise sharply during this developmental transition, with many youth experiencing their first onset of depressive symptoms between early-to-late adolescence (Cole et al., 2002; Gudmundsen et al., 2019). At least two in five U.S. youth endorsed past-year depressive symptoms (Bommersbach et al., 2025), with prevalence rates of clinical diagnoses of depression increasing by 60% between 2017 and 2021 (Xiang et al., 2024). Notably, depressive symptoms tend to persist beyond adolescence and into adulthood, highlighting the importance of early detection and intervention (Keyes et al., 2024). Given the significant prevalence and socioeconomic burden of adolescent depression, identifying early-emerging emotional and neurobiological processes that confer risk is essential.
Growing research highlights emotion dysregulation as a transdiagnostic process underlying the development of depression and related psychopathology (Aldao et al., 2016; McLaughlin et al., 2020; Milojevich et al., 2019; Seah et al., 2026). Broadly, emotion dysregulation encompasses a range of implicit (unconscious and automatic) and explicit (conscious and effortful) processes. These include, but are not limited to, difficulties identifying, understanding, and/or regulating one’s own emotions (Beauchaine and Cicchetti, 2019; Gratz and Roemer, 2004; Vine and Aldao, 2014), as well as more frequent and inflexible engagement of putatively maladaptive (e.g., rumination, avoidance) versus adaptive (e.g., reappraisal, problem-solving) coping strategies (Aldao et al., 2010; Gross, 2015; Sheppes et al., 2015). Yet, the specific components of emotion dysregulation that predict later depressive symptoms, as well as the neural circuits that may influence these vulnerabilities, remain less understood. Considering that emotional and neurocognitive processes undergo significant maturation during adolescence, it is crucial to examine how they interact to prospectively influence adolescent depression risk.
Emotional awareness, defined as the ability to identify and understand one’s internal feeling states, represents a foundational dimension of emotion regulation pertinent to psychopathology and treatment (Gross, 2015; Mennin, 2006). Low emotional awareness is associated with the reduced capacity to recognize self-relevant emotional cues, difficulty monitoring changes in affect, heightened emotional and physiological reactivity to stress, and reliance on maladaptive coping strategies (Boden and Thompson, 2015; Edwards and Wupperman, 2017; Seah and Coifman, 2022). Moreover, the process model of emotion regulation postulates that emotional awareness is an upstream process that likely precedes the deployment of emotion regulatory strategies such as cognitive reappraisal (Gross, 2015). Emerging evidence suggests that emotional awareness deficits predict the development of depression and other psychopathology (e.g., anxiety, substance use) in youth (Kranzler et al., 2016; Rieffe et al., 2008; Weissman et al., 2020). Even after accounting for other factors that putatively exert influence on affective psychopathology, such as age, biological sex, and other symptoms (e.g., aggression, traumatic stress), low emotional awareness was associated with depression and maladaptive coping behaviors (e.g., rumination, conduct problems), suggesting that it accounts for unique variance in psychopathology.
Adolescence is a sensitive developmental period that demands heightened emotional awareness, as youth navigate increasingly complex social contexts that elicit emotions of varying intensity while also learning to interpret and articulate internal emotional experiences effectively (Blakemore and Mills, 2014; Morie et al., 2022; Wintre and Vallance, 1994). Prior research suggests that aspects of emotional awareness, such as the ability to identify and differentiate one’s own emotions, are diminished in adolescents compared with children and adults (Nook et al., 2018) and prospectively influence the development of internalizing psychopathology (e.g., social anxiety; Seah et al., 2024). Poor emotional awareness may thus function as an early-emerging indicator of depression risk, potentially shaping how adolescents respond to emotionally salient events and interact with their social environments (Van Beveren et al., 2019). However, emotional awareness shows substantial individual variability and may reflect a normative developmental inflection, with only some youth progressing toward elevated depressive symptoms.
Neural systems implicated in reward and motivation—processes relevant to depression and that undergo significant maturation during adolescence—may be an important moderating factor. Although much of the literature has focused on activation-based indices of reward processing, connectivity-based approaches provide complementary information regarding the coordination of neural systems supporting reward processing (Forbes et al., 2021). Frontostriatal circuitry between the dorsomedial prefrontal cortex (dmPFC) and the nucleus accumbens (NAcc) may be relevant for understanding adolescent neural reward function and depression (Eckstrand et al., 2022a; Forbes et al., 2021; Pizzagalli, 2014). Prior meta-analyses of reward-related fMRI tasks indicate that reward receipt reliably activates the NAcc and PFC (Liu et al., 2011; Oldham et al., 2018). These brain regions are involved in reward processing, valuation, and regulation, which are crucial for reward-driven mood and behavior (Etkin et al., 2011; Haber and Knutson, 2010). The NAcc, which forms most of the ventral striatum, is considered the hub of the neural reward system underlying the motivational aspects and subjective experience and valuation of reward. The dmPFC, which is implicated in self-referential cognitive processing, exerts top-down control over striatal reward signals, regulating how reward influences mood and behavior (Haber and Knutson, 2010). Functional connectivity between these regions may thus reflect the extent to which reward-related signals are integrated into higher-order self-relevant mental representations, allowing better understanding of reward system-level function than activation-based metrics alone.
Converging evidence from functional magnetic resonance imaging (fMRI) studies suggests that altered frontostriatal functional connectivity is characteristic of adolescent depression, particularly in the context of anhedonia and blunted reward sensitivity (Forbes and Dahl, 2012; Healey et al., 2014; Kerestes et al., 2015; Morgan et al., 2016). For example, relative to those without depression history, adolescent boys with past depression demonstrated heightened positive (i.e., stronger) dmPFC-NAcc functional connectivity during a task of winning versus losing monetary rewards (Morgan et al., 2016). Hyper-connectivity, as indicated by a more positive relationship between the dmPFC and NAcc during reward‐related tasks, has been interpreted as greater top‐down regulation of striatal reward responses by the dmPFC (Forbes et al., 2021; Forbes and Dahl, 2012), which may diminish the motivational salience of positive experiences. Such neural patterns have also been observed in healthy adolescents with a familial history of depression, suggesting that atypical reward circuitry may reflect a premorbid vulnerability rather than solely a consequence of current symptoms or impairment (Gupta et al., 2024b). Finally, greater decreases in frontostriatal functional connectivity during transcranial magnetic stimulation have been associated with enhanced positive mood in depressed youth (Gupta et al., 2024a), highlighting the relevance of this neural circuitry for understanding and treating depressive symptoms.
Given evidence linking individual differences in emotional awareness and frontostriatal function with depression, it is crucial to examine how they interact to form a neurobehavioral profile of depression risk during adolescence, when both emotional and neural systems are rapidly developing. Emotional awareness and frontostriatal reward function reflect distinct yet related processes: the former indexing how well youth attend to and understand affective cues, and the latter indexing how effectively such affective cues as reward signals are generated and integrated to influence mood and behavior. Receipt of reward tends to evoke a feeling of “liking,” which refers to a consummatory process associated with subjective pleasure (Berridge et al., 2009), although awareness of such feelings is modulated by frontostriatal function. Therefore, low emotional awareness and altered frontostriatal function may interactively influence the degree to which youth engage with rewarding experiences and use them effectively for positive reinforcement (Hogeveen et al., 2021; Morie et al., 2016). The combination of struggling to pinpoint positive emotions (e.g., pride) in response to pleasant events (e.g., getting a high score on an exam) alongside altered processing of neural reward signals may disrupt adaptive responding to rewarding experiences, precipitating the emergence of youth depression and related symptoms (e.g., anhedonia; Goerlich et al., 2017; Hogeveen et al., 2021; Morie et al., 2016). However, longitudinal studies testing these interactive pathways are few, which limits understanding of pertinent mechanisms that may explain the development of youth depression.
Current study
The present study addresses this knowledge gap by examining how emotional awareness and frontostriatal functional connectivity in response to rewarding outcomes jointly predict depressive symptoms over 1 year in adolescents at risk for affective disorders based upon familial psychiatric history. We focused on functional connectivity during reward outcome, as this stage involves processing and evaluation of reward receipt that may be particularly relevant to depressive symptom development (Fischer et al., 2019; Luking et al., 2016). Functional connectivity between dmPFC and NAcc was examined in order to capture coordinated activity between regions implicated in reward valuation (NAcc) and self-referential cognitive processing (dmPFC). This approach allows for examination of how self-relevant reward-related signals are integrated into higher-order evaluative processes to shape behavior, which may be particularly relevant for understanding depression risk. To assess dmPFC–NAcc functional connectivity during receipt of reward, we used a reward-based fMRI card guessing task that reliably engages the dmPFC and NAcc in adolescents with or at-risk for depression (Delgado et al., 2000; Forbes et al., 2009). Given known links between age, biological sex, and familial psychiatric history with depression, these variables were included as covariates in our analyses. Based on prior work and developmental models of emotion and neurobiology, we hypothesized that low emotional awareness would predict more severe depressive symptoms over time, especially among youth exhibiting heightened positive (i.e., stronger) dmPFC–NAcc functional connectivity during reward processing, potentially indicative of greater top-down regulation of striatal reward responses.
By integrating emotional and neural markers of risk, this study contributes to a multimodal understanding of the mechanisms through which processes related to emotion regulation confer vulnerability to depression, during a developmental period of heightened risk. Findings have the potential to inform early detection and intervention targets to mitigate the emergence of depression during adolescence.
Methods
Participants
Participants were recruited as part of a longitudinal study at a U.S. research university examining the development of anhedonia in at-risk youth based on the following eligibility criteria: (1) aged between 13 and 19 years at baseline, (2) had a first-degree relative with lifetime history of a depressive disorder, bipolar disorder, or schizophrenia/schizoaffective disorder (classified as “high-risk;” 66%) or without familial history of any mood or psychotic disorder (classified as “low-risk;” 34%), (3) not currently taking psychotropic medication for at least 2 weeks prior to study participation (except use of stimulant medication which was discontinued for 36-hours prior to the MRI scan), (4) did not meet criteria for lifetime mood or psychotic disorder or moderate/severe substance use disorder diagnosis based upon the Diagnostic and Statistical Manual, Fourth Edition, Text Revision (DSM-IV-TR; American Psychiatric Association, 2000) at study entry.
Of the total sample that completed fMRI scanning at baseline (N = 119), N = 24 had unusable fMRI data for analyses (e.g., excessive motion during scanning and/or ≤50% coverage of the NAcc) and N = 11 had data issues (e.g., corrupt output files, data management error), leaving a final sample of N = 84 available for analyses. Of these 84, N = 2 had missing data on baseline emotional awareness, N = 8 had missing dmPFC–NAcc connectivity values, and N = 17 had missing data on 12-month depressive symptoms. There were no missing data on baseline depressive symptoms, age, sex, and risk group. This left a final sample of N = 65 without any missing values. 1
The Structured Clinical Interview for DSM-IV Disorders (SCID-IV; First et al., 1996) was used to assess first-degree relatives (e.g., parents) diagnoses, while the Kiddie Schedule for Affective Disorders and Schizophrenia, Present and Lifetime Version (KSADS-PL; Kaufman et al., 1997) was used to assess and confirm psychiatric diagnoses in youth participants and potentially affected first-degree relatives (e.g., sibling) <18 years old.
Procedures
At baseline, participants completed diagnostic interviews and questionnaires assessing demographics, emotional awareness, and depressive symptoms. Following, participants underwent fMRI scanning during a card guessing task involving winning and losing small monetary rewards (see Measures). At one-year follow-up, self-reported depressive symptoms were assessed again. All study procedures were approved by the University of Pittsburgh IRB, and participants (and their parent/guardian if age <18) provided informed consent and/or assent.
Measures
Emotional awareness
A 26-item questionnaire (Boden and Thompson, 2015) was used to assess different facets of emotional awareness. Questionnaire items assessed emotional attention, clarity, and differentiation. Item examples: “I am often confused about what emotion I am feeling,” “I don’t pay much attention to my feelings,” and “I am usually very clear about my feelings.” Participants rated each item on a five-point Likert-type scale (“1-Strongly Disagree” to “5-Strongly Agree”). Items were scored accordingly and summed, such that lower scores indicated poorer emotional awareness. The sample’s mean (SD) emotional awareness at baseline was 96.96 (14.91). The scale demonstrated high internal consistency (Cronbach’s α = 0.91).
Depressive symptoms
The 20-item Center for Epidemiological Studies-Depression Scale (CES-D; Radloff, 1977) scale was administered at baseline and at 1-year follow-up to assess past-week depressive symptoms. Participants responded using a 4-point Likert-type scale (“0-Rarely or none of the time (less than 1 day)” to “3-Most or all of the time (5–7 days)”). Items were summed to create an index of depression. The sample’s mean (SD) at baseline and one-year follow-up was 11.85 (SD = 9.00) and 10.80 (SD = 8.75), respectively. At baseline and 1-year follow-up, 28% and 24% scored above the clinical cut-off of 16. The scale demonstrated high internal consistency (Cronbach’s α = 0.87/0.90 at baseline and 1-year follow-up).
Monetary reward fMRI task
At baseline, participants completed a card guessing fMRI task (see Eckstrand et al., 2022b for task diagram). The event-related task consisted of three outcome contexts (win, loss, and neutral outcome) and four anticipation 2 contexts (possible win, possible loss, mixed win/loss, and neutral). Each outcome was associated with different monetary values ($1.00/win; deduction of $0.50/loss; $0 for neutral). The trials were presented in the same pseudorandom order for all participants. Each trial, participants used a button-press to guess whether a card number would be greater or less than 5 (4 s) and they were informed of the trial type using an expectancy cue (2–6 s, jittered; anticipation phase). Then, participants were shown the actual card number (selected randomly; 500 ms), followed by a symbol (500 ms) indicating whether they won or lost money, or had no change in earnings (outcome phase). Between each pair of trials, there was a jittered 0.5–1.5 s inter-trial interval. The paradigm was administered in an 8-min block, with 48 trials and 12 trials per trial type. Participants were unaware that the outcomes of trials were fixed, such that all would experience the same pattern of outcomes and win the same amount of money. They were debriefed regarding the predetermined trial outcomes upon study completion. Although task-specific reliability estimates were not computed for the present dataset, prior work examining the reliability of this task suggests that the reliability of neural activation and connectivity metrics is typically modest but may still be meaningful for individual differences research (e.g., Baranger et al., 2021; Lenniger et al., 2026; Noble et al., 2019).
fMRI acquisition and preprocessing
Functional neuroimaging data were collected at the University of Pittsburgh using a 3.0T Siemens Prisma MRI scanner. Blood-oxygenation-level-dependent (BOLD) images were acquired with a multi-band gradient echo EPI sequence (18 slices, three-factor multiband; 2.3 mm isotropic voxels; TR = 1500 ms, TE = 30 ms; FOV = 220 × 220 mm; matrix 96 × 96; flip angle 55°, bandwidth 1860 Hz Px–1). Structural 3D axial MPRAGE images (TR = 1500 ms, TE = 3.19 ms; flip angle 8° FOV = 256 × 256 mm; 1 mm isotropic voxels; 176 continuous slices) and field maps (2.3 mm isotropic voxels; TR = 500 ms, TE1 = 4.92 ms, TE2 = 7.38 ms; FOV = 220 × 220 mm; flip angle 45°, bandwidth 1302 Hz Px–1) were acquired in the same session.
Preprocessing was performed using Statistical Parametric Mapping (SPM) software, version 12 (http://www.fil.ion.ucl.ac.uk/spm). For each participant, BOLD images were realigned to the mean volume in the time series and co-registered with the participant’s structural image. Field maps were used to correct for image distortion. Structural images were normalized using a nonlinear transformation to the standard MNI/ICBM 152 tissue probability maps and segmented into gray matter, white matter, cerebrospinal fluid, and other tissues (e.g., bone). BOLD images were transformed into the same space using the structural image and resampled at 2 mm3 isotropic voxel size. BOLD images were normalized and spatially smoothed (FWHM 6 mm).
Data analytic strategy
Neuroimaging analyses
In the current study, the main contrast of interest in our functional connectivity analyses was win outcome > loss outcome. Functional connectivity was examined using generalized psychophysiological interaction (gPPI) analysis, which measures the degree of coactivation between neural regions. We chose to contrast win trials vs. loss trials as gPPI analyses require that coordination between physiological regions of interest depend on correlation with psychological states as inferred from active task conditions (O’Reilly et al., 2012). Therefore, subtracting participants’ responses to loss trials would isolate their responses to win trials specifically from responses to feedback in general.
First-level analyses were completed in SPM12. A fixed effect general linear model (GLM) was performed for each participant, which included the main contrast of interest: win outcome > loss outcome. To identify volumes with high motions and artifacts, the Artifact Detection Toolkit (ART) was utilized and censored volumes with movement of > 3SDs from participants’ mean, > 0.5 mm scan-to-scan translation, or > 0.01◦ of scan-to-scan rotation (Chai et al., 2014), and were entered as covariates in the subject-level GLM to reduce motion-related noise. Lastly, the six motion realignment parameters were entered as covariates to control for head movement. A 128 s high-pass filter and autoregressive modeling were implemented during fitting.
To compute dmPFC–NAcc functional connectivity, we used the built-in PPI toolbox in SPM12. We extracted the generated seed region BOLD time series for the bilateral NAcc, which were then deconvolved using the standard deconvolution algorithm in SPM to estimate the neuronal time series for the given volume of interest. The neuronal time series was then multiplied by the psychological variable (task contrast: win > loss outcome) to generate an interaction term and then convolved with the hemodynamic response function (HRF) to generate a gPPI interaction term. The physiological variable (original BOLD time series of the volume of interest), psychological variable (stimulus onsets convolved with the HRF), and gPPI interaction term were then included into the PPI design matrix. Region-of-interest (ROI) masks for the seed region (bilateral NAcc) and the dmPFC were derived from the Neurosynth meta-analytic database (using thresholded versions of the association test map, thresholded at 50%). Supplementary figure S1 illustrates these ROIs. Beta weights for the gPPI interaction term were then extracted for use in subsequent regression analysis.
To examine task-related activation in NAcc and dmPFC, first-level contrast images (win > loss outcome)—reflecting neural reward activity relative to loss—were entered into a second-level GLM for data extraction in SPM. The same NAcc and dmPFC ROI masks derived above were used in these analyses. Marsbar (http://marsbar.sourceforge.net/) was used to extract contrast values representing BOLD activity in these regions for subsequent supplemental analyses.
Preliminary analyses
To examine the pattern of missingness in our primary outcome variables, Little’s Missing Completely at Random (MCAR; Little, 1988) test was conducted. Additionally, we compared whether the original sample of 65 participants without any missing data systematically differed in baseline depressive symptoms, age, sex, and risk group from the 19 participants with partial missing data using nonparametric tests.
The distribution of primary outcome variables was then examined. All outcome variables were normally distributed. Demographic differences in outcome variables were assessed using t-tests or one-way ANOVAs. Given the skewed distribution of individuals identifying as Hispanic, the nonparametric independent-samples Mann–Whitney U Test was used to examine group differences. Bivariate correlations between variables were examined to identify multicollinearity and potential covariates to be included in subsequent analyses.
Primary analyses
Moderation analyses were conducted using the Hayes PROCESS macro in SPSS. Regression analyses were conducted by entering dmPFC–NAcc functional connectivity as the moderator of the relationship between baseline emotional awareness predicting depressive symptom severity at one-year follow-up. Age, biological sex, familial risk status, and baseline depressive symptoms were included as covariates. The Johnson–Neyman technique was used to probe and plot significant moderation effects by determining the region(s) of significance, that is, value(s) of dmPFC-NAcc functional connectivity at which baseline emotional awareness was significantly associated with future depressive symptom severity (Bauer and Curran, 2005; Finsaas and Goldstein, 2021).
Results
Preliminary analyses
Results from Little’s MCAR test revealed that the data was not missing in a systematic way, as indicated by a non-significant p value χ2 = 6.54, p = 0.835. Following, the expectation-maximization algorithm (with 25 iterations; Dempster et al., 1977) was used to iteratively impute missing values accordingly in the sample of 19 participants with partial missing data, yielding a final total sample of 84 participants for analysis. Comparing between N = 65 without missing data with N = 19 with partial missing data, results from independent-samples Mann-Whitney U Tests indicated no significant differences in baseline depressive symptoms (p = 0.872) and age (p = 0.671). Results from chi-square tests of independence indicated no significant differences in risk group (p = 0.392) although there was a sex difference (p = 0.039), where a greater proportion of male participants had partial missing data than female participants.
Table 1 describes the final sample’s (N = 84) sociodemographic characteristics. Results from independent samples t-tests indicated that youth classified as “high-risk” (M = 12.44, SD = 9.35) reported more severe depressive symptoms at one-year follow-up compared with “low-risk” youth (M = 7.69, SD = 6.55), t(82) = 2.44, p = 0.017. No other demographic differences in depressive symptoms at baseline or follow-up were observed (ps > 0.05).
Demographic Characteristics of the Sample (N = 84)

Table 2 describes results from bivariate correlational analyses. Lower emotional awareness at baseline was associated with higher depressive symptoms severity at baseline and at 1-year follow-up. Depressive symptoms at baseline and 1-year follow-up were positively associated. No other significant correlations were observed.
Bivariate Correlations Between Primary Outcome Variables (N = 84)
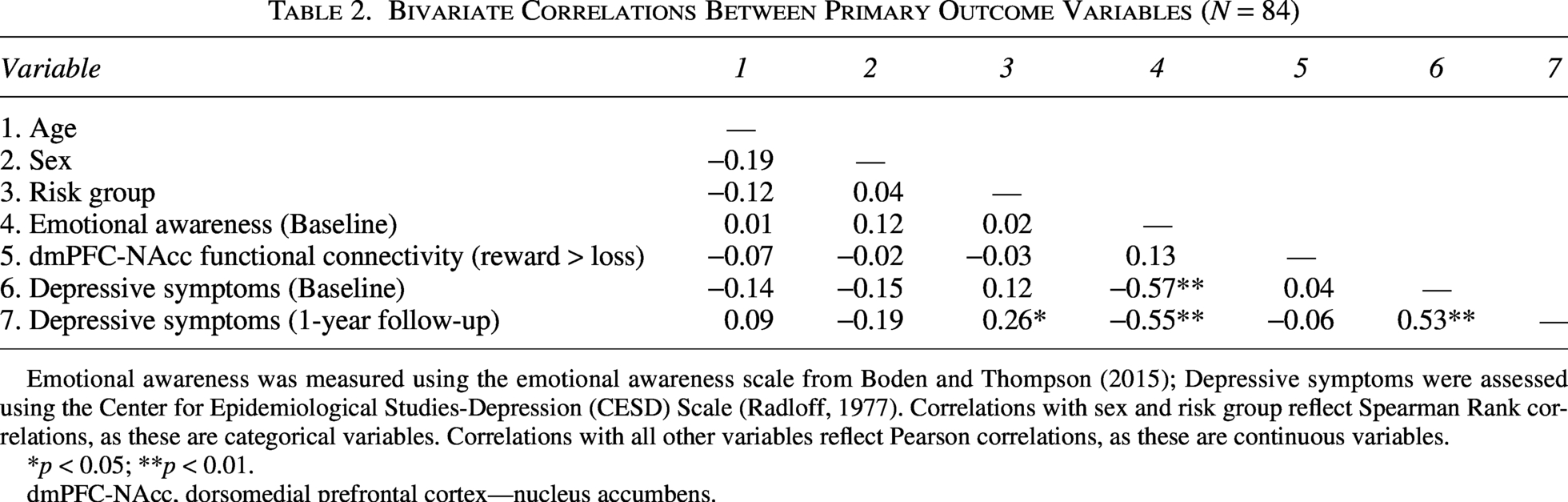
Emotional awareness was measured using the emotional awareness scale from Boden and Thompson (2015); Depressive symptoms were assessed using the Center for Epidemiological Studies-Depression (CESD) Scale (Radloff, 1977). Correlations with sex and risk group reflect Spearman Rank correlations, as these are categorical variables. Correlations with all other variables reflect Pearson correlations, as these are continuous variables.
*p < 0.05; **p < 0.01.
dmPFC-NAcc, dorsomedial prefrontal cortex—nucleus accumbens.
Primary analyses
Moderation analyses revealed a significant interaction between baseline emotional awareness and dmPFC–NAcc functional connectivity during processing of reward (relative to loss) in predicting future depressive symptoms, p = 0.027 (Table 3). Specifically, lower emotional awareness at baseline predicted a greater increase in depressive symptoms at follow-up, particularly among youth demonstrating stronger positive dmPFC–NAcc functional connectivity during reward (vs. loss) receipt. The Johnson-Neyman technique revealed that low emotional awareness predicted more severe depressive symptoms when dmPFC–NAcc functional connectivity was more positive (Johnson-Neyman threshold ≥ −1.04; 82% of all values, p < 0.05). Descriptive statistics of dmPFC–NAcc connectivity values were: M = −0.19, SD = 1.31, range: −2.65–6.28 3 . To illustrate the moderation effect, we plotted the relationship between baseline emotional awareness and future depressive symptoms as a function of dmPFC-NAcc functional connectivity (Fig. 1).
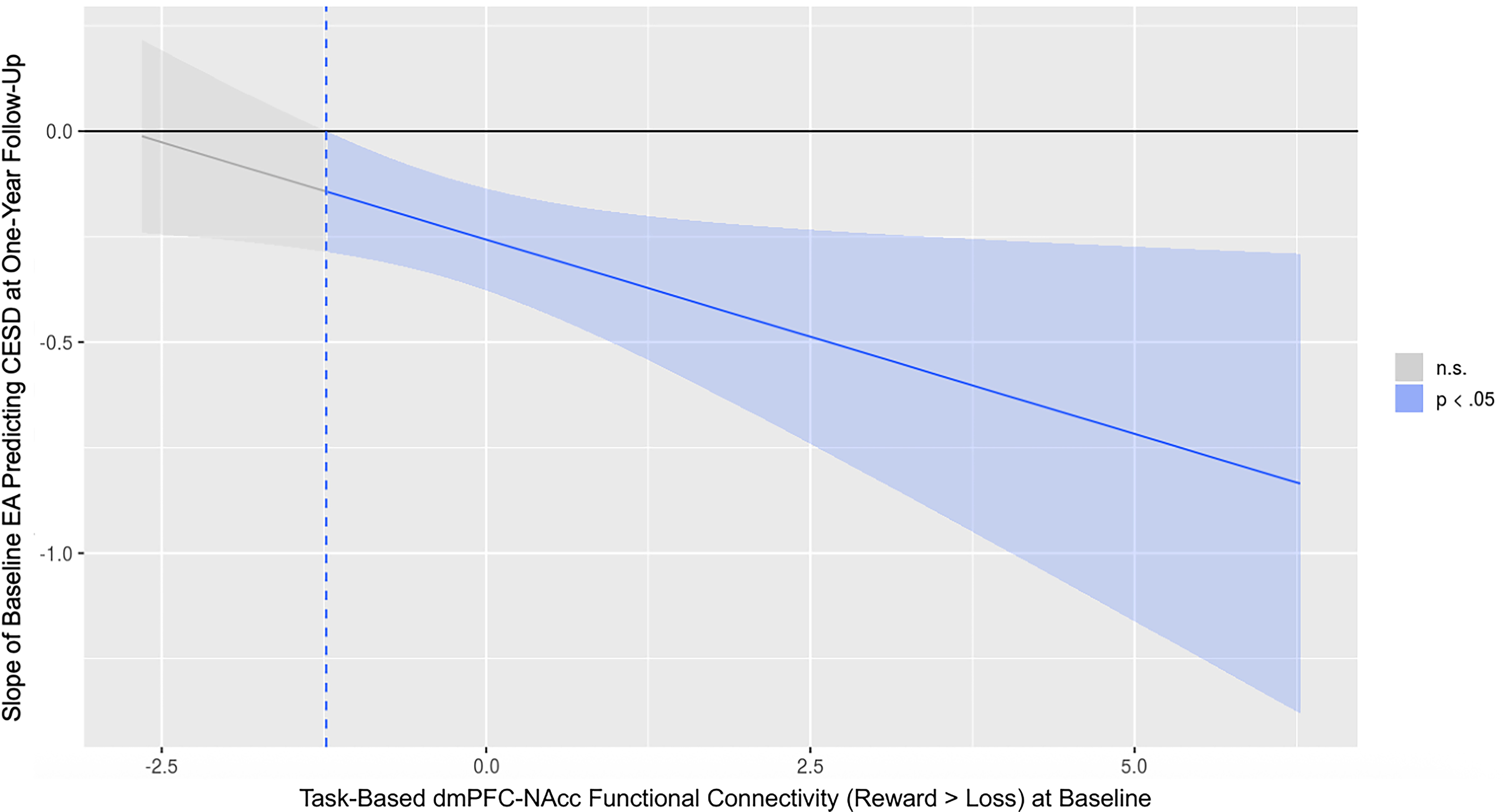
Johnson–Neyman plot of two-way interaction between emotional awareness and task-based dmPFC-NAcc functional connectivity (reward > loss) at baseline predicting depressive symptom severity at 1-year follow-up. Regions shaded in blue indicate values of dmPFC-NAcc functional connectivity where low emotional awareness was significantly associated with worse depressive symptoms (i.e., 82% of all values). Values indicated on x-axis reflect raw values of dmPFC-NAcc functional connectivity of the sample (N = 84; range: −2.65–6.28). CESD, Center for Epidemiological Studies-Depression Scale; dmPFC-NAcc, dorsomedial prefrontal cortex—nucleus accumbens; EA, emotional awareness.
Two-Way Interaction between Baseline Emotional Awareness and dmPFC-NAcc Functional Connectivity (Reward > Loss) Predicting Depressive Symptoms at One-Year Follow-up (N = 84)
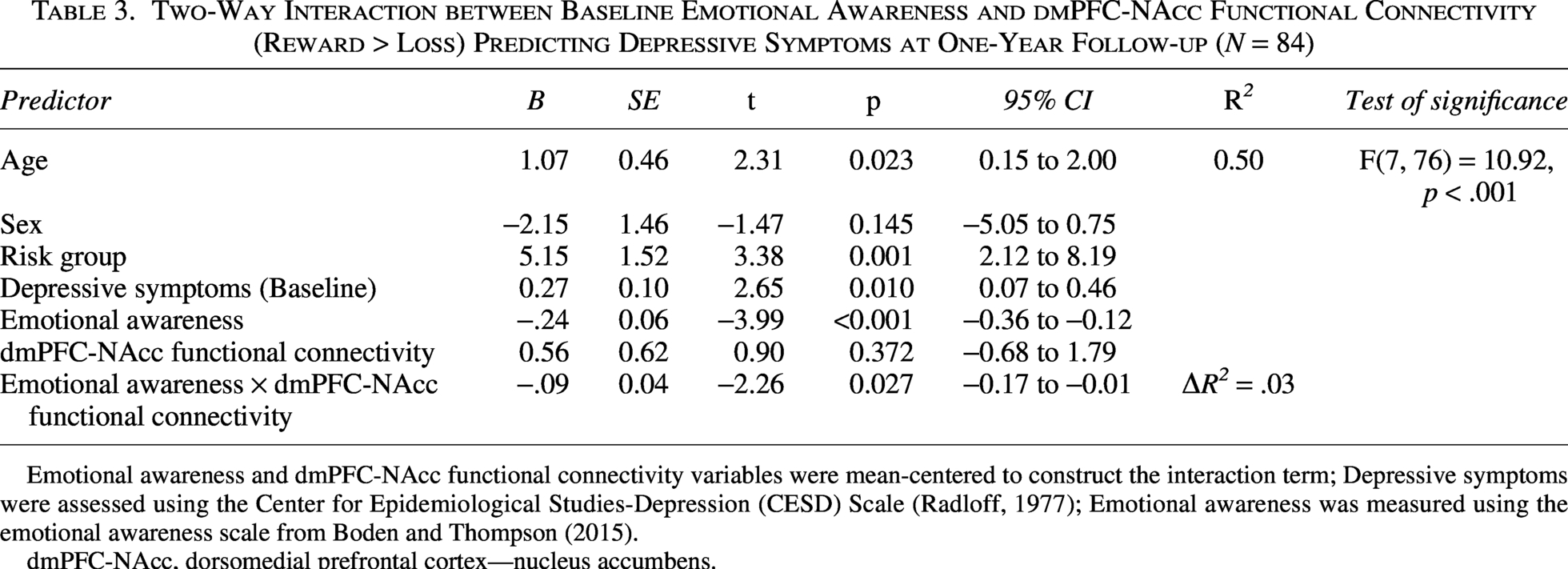
Emotional awareness and dmPFC-NAcc functional connectivity variables were mean-centered to construct the interaction term; Depressive symptoms were assessed using the Center for Epidemiological Studies-Depression (CESD) Scale (Radloff, 1977); Emotional awareness was measured using the emotional awareness scale from Boden and Thompson (2015).
dmPFC-NAcc, dorsomedial prefrontal cortex—nucleus accumbens.
Supplemental analyses
Moderation by dmPFC–NAcc functional connectivity based on reward > neutral contrast
To clarify if the moderation effect observed above was driven primarily by reward or loss trials, or their contrast, we conducted follow-up moderation analysis using dmPFC–NAcc functional connectivity that contrasted reward trials with the neutral condition (reward > neutral). Results indicated no significant moderation effect based upon this alternate contrast, B = −.03, SE = .04, p = .401, 95% CI: [−.11; .05] (Supplementary Table S3). These findings suggest that the moderation effect by dmPFC-NAcc functional connectivity was driven by responses to reward relative to loss, rather than by just reward-related processes alone.
Moderation by dmPFC or NAcc task-related activation
Besides functional connectivity, we conducted supplemental analyses examining if task-related dmPFC and NAcc activation during reward vs. loss trials moderated the relationship between baseline emotional awareness and follow-up depressive symptoms. Results indicated no significant moderation by task-related dmPFC (B = −0.04, SE = 0.03, p = 0.143) or NAcc (B = −0.07, SE = 0.07, p = 0.277) activation (Supplementary Table S4).
Discussion
The present study is the first to examine how low emotional awareness and altered frontostriatal circuitry during processing of reward relative to loss may constitute a neurobehavioral profile of depression risk during adolescence, a vulnerable developmental period encompassing emotional and neurocognitive maturation. Consistent with prior research, we found heightened severity of depressive symptoms over time among youth with familial psychiatric history (e.g., Van Dijk et al., 2021). In addition, deficits in emotional awareness were associated with more severe depressive symptoms at baseline and 1-year follow-up (Kranzler et al., 2016; Weissman et al., 2020). However, unlike prior research, dmPFC-NAcc connectivity was not directly associated with depressive symptoms. Rather, low emotional awareness was prospectively associated with more severe depressive symptoms over time, particularly among youth demonstrating relatively heightened positive dmPFC-NAcc functional connectivity—a pattern suggestive of over-regulation of reward responses. This interaction effect was present even after controlling for key covariates such as age, biological sex, familial risk status, and baseline depressive symptoms, highlighting that both factors accounted for unique variance in depression. Note that task-related dmPFC and NAcc activation were not significant moderators. These findings suggest that emotional awareness may confer risk for depression, particularly in the context of altered frontostriatal connectivity during processing of reward (relative to loss), highlighting the importance of considering the interplay of neurobiological and emotional processes in predicting youth depressive symptoms.
Our findings extend growing research by pinpointing neurobiological mechanisms that influence associations between dimensions of emotion regulation and the development of youth depression. We focused on emotional awareness, which is a foundational and upstream process underlying effective emotion regulation outcomes (Boden and Thompson, 2015; Seah et al., 2020). The study results indicate that variability in neurobiological characteristics, such as frontostriatal connectivity during receipt of reward (relative to loss), may amplify the pernicious effects of poor emotional awareness, impacting trajectories of depressive symptom severity. The observed range of dmPFC–NAcc connectivity values in our study was consistent with the modest magnitude typically observed in other task-based frontostriatal connectivity studies of youth (e.g., Parr et al., 2021; Poon et al., 2019). Prior work examining NAcc connectivity with prefrontal regions has reported values centered near zero, reflecting small but meaningful individual differences rather than large absolute deviations. Accordingly, the term “heightened connectivity” in the present study refers to relatively greater connectivity within the observed sample rather than abnormally elevated connectivity in general.
Heightened positive dmPFC–NAcc connectivity during receipt of reward has been interpreted as excessive prefrontal modulation of striatal reward signals, potentially dampening reward responsiveness and positive affect (Forbes and Dahl, 2012; Healey et al., 2014; Morgan et al., 2016). Such over-regulation may interfere with adolescents’ ability to fully experience or learn from positive outcomes, thereby contributing to anhedonia and motivational deficits that characterize depression. Our findings suggest that the detrimental effects of reward dysregulation are especially significant when contrasted with responses to loss. Contrary to prior research, dmPFC–NAcc connectivity was not directly associated with depressive symptoms. This could be due to a combination of factors such as task-specific effects, developmental timing, and sample characteristics. For example, our sample consisted mostly of middle-to-late adolescents, a developmental stage where other forms of reward (e.g., social) may be more salient than monetary rewards (e.g., Kilford et al., 2024). This suggests that functional connectivity may not function as a standalone risk marker, but rather as a context-dependent moderating characteristic that influences developmental vulnerability for depression in the presence of other factors, such as lower emotional awareness. When paired with low emotional awareness, this neurobehavioral profile may further undermine the reinforcing value of positive events and decrease approach motivation, precipitating depressive symptoms over time. Both emotional awareness and neural reward circuitry may thus function as crucial processes that shape how affective cues, such as reward signals, are interpreted and translated into subjective experience and behavior.
The present findings also have important implications for extending emotion dysregulation frameworks for understanding psychopathology. Emotional awareness is often conceptualized as an early-stage emotion regulation trait and skill that supports downstream regulatory processes, including strategy selection and flexible responding to emotional stimuli (Gross, 2015; Van Beveren et al., 2019). Adolescents with poor emotional awareness may have difficulties recognizing and labeling positive affect, misattribute its source, or struggle to implement adaptive emotion regulation strategies effectively. When paired with neural activity indicative of over-control of reward signals, positive reinforcement learning is likely disrupted, leading to decreased motivation and heightened withdrawal consistent with depressive phenotypes (Pizzagalli, 2014). This aligns with developmental psychopathology models emphasizing multifinality, where pre-existing emotional vulnerabilities may lead to different mental health outcomes depending on individual differences in neurobiological functioning (Forbes et al., 2021; Nolen-Hoeksema and Watkins, 2011).
Clinical Significance
Clinically, these findings underscore the potential utility of interventions that target emotional awareness and reward processing early in development, particularly for youth at elevated risk (e.g., familial psychiatric history). Skills-based interventions that explicitly train emotional identification, labeling, and differentiation may help adolescents to engage with emotional experiences more effectively (Matt et al., 2024; Seah and Coifman, 2025). For example, these skills are already available in interventions such as dialectical behavior therapy and mindfulness-based interventions (Linehan, 1993; Mennin, 2006; Seah et al., 2025b). Similarly, targeting reward processing, such as savoring of pleasure associated with rewarding outcomes, could strengthen reward sensitivity and positive reinforcement, buffering depression risk (Craske et al., 2023). Our findings also highlight frontostriatal circuitry as a potential mechanistic intervention target. Emerging work suggests that neuromodulatory approaches and behavioral interventions can influence reward-related neural functioning in adolescents and adults (Gupta et al., 2024a; Nagy et al., 2020). Integrating emotional regulation skills training with neuromodulatory interventions may be particularly beneficial for youth exhibiting both emotional awareness deficits and frontostriatal dysfunction (e.g., Neacsiu et al., 2022).
Strengths and Limitations
The present study had several strengths. We recruited a risk-enriched youth sample with varying familial psychiatric history and incorporated multimodal neurobehavioral data. Importantly, the longitudinal study design allowed testing of prospective relationships. Yet, the study also has limitations. The sample size was relatively modest, which may limit statistical power and generalizability. Replication in larger samples is needed to confirm the robustness of these interaction effects. While the longitudinal design strengthens causal inference, we examined depressive symptoms at only two time points. Additional follow-up assessments would enable further understanding of long-term developmental trajectories. The present study focused on reward-related frontostriatal connectivity during a well-validated monetary reward paradigm. Although task reliability estimates were not directly computed for the present dataset, prior work suggests that the reliability of this task is modest and varies across regions, task stages, and analytic approaches (Baranger et al., 2021; Lenniger et al., 2026). These considerations underscore the importance of interpreting neural findings cautiously and highlight the need for future studies to incorporate explicit reliability assessments when possible. Furthermore, given that social forms of reward (e.g., peer acceptance) are particularly salient during adolescence (Blakemore and Mills, 2014; Schriber and Guyer, 2016), future work should examine if a similar pattern of findings emerges in response to social reward or peer feedback (Eckstrand et al., 2022b; Seah et al., 2025a).
Conclusion
In summary, the present study provides novel evidence that emotional awareness and frontostriatal functional connectivity interact to prospectively predict youth depressive symptoms. Low emotional awareness predicted worse depressive symptoms, particularly among youth showing altered dmPFC–NAcc functional connectivity during reward receipt. These findings highlight the importance of integrating emotional and neural markers of risk within developmental models of depression. Early interventions targeting emotional awareness and reward-related processes may help mitigate the emergence of youth depressive symptoms.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.