
Abstract
Megakaryocytes (MKs), which are differentiated from megakaryocytic–erythrocytic progenitors, generate platelets (PLTs) by expanding and branching their cellular fragments under the influence of cytokines and intercellular mechanisms such as autophagy and release of reactive oxygen species (ROS) in the bone marrow. Autophagy is a self-destructive process that plays a significant role in cell growth and energy maintenance of the cells. In contrast, ROS are toxic products of cellular metabolism that are harmful to human stem cells but have a crucial role in determining lineage commitment at the common myeloid progenitor stage and deriving further maturation progression toward MKs and PLTs production, with an interconnected relationship in the onset and deriving of autophagy. This review summarizes and discusses what has been discovered about the current state of knowledge regarding autophagy effects on MK differentiation, ROS effects on megakaryopoiesis (MKp), and the relationship between these molecules and autophagy initiation. Furthermore, in vitro applications of controlling these external factors on MKp are reviewed according to pertinent studies. Utilizing these regulatory mechanisms supports an improved design of in vitro MKp for introducing artificial PLT sources and might be beneficial in creating novel treatments of PLT-related coagulation disorders.
Introduction
Autophagy is an intracellular cytoplasmic degradation process with the participation of lysosomes, having a key role in maintaining cell survival (Das et al., 2012). Pivotal to sustaining intracellular homeostasis, autophagy is reported to be essential in acclimation to starvation, clearance of excessive or defective internal organelles, antigen presentation, and also regulation of organ development (Allen and Baehrecke, 2020; Klionsky, 2007; Mizushima, 2005). To date, several studies have found links between autophagy and major human diseases such as diabetes (Bhattacharya et al., 2018; Wang et al., 2024), cancer (both in tumor progression and suppression) (Chavez-Dominguez et al., 2020; Debnath et al., 2023; Mulcahy Levy and Thorburn, 2020), gastrointestinal (GI) disorders (such as inflammatory bowel disease, GI cancer, and GI motility disorders (Chang et al., 2024)), infectious diseases (Liu et al., 2022; Niu and Deng, 2022), and neurodegenerative disorders (such as Alzheimer’s disease and Parkinson’s disease; Fujikake et al., 2018; Guo et al., 2018). Considering the significant impact of autophagy as a balancing strategy, it would be predictable to see more and more links between autophagy and the pathophysiology of other diseases (Jing and Lim, 2012). This remarkable interconnection between autophagy and health-disease states implicates an undeniable chance of novel therapeutics aimed at modulating autophagy (Mulcahy Levy and Thorburn, 2020; Zhao et al., 2023). More than 30 autophagy-related genes (ATGs) that take part in this biological process have been discovered up until now. These genes are strongly conserved in eukaryotes, displaying the crucial role of this mechanism (Nakatogawa et al., 2009).
Megakaryocytes (MKs) are generated from hematopoietic stem cells (HSCs), which are located in bone marrow (BM); however, at the early stages of hematopoiesis, they are found in the liver, yolk sac, and spleen (Ogawa, 1993). Several transcription factors such as Fli 1, RUNX1, and GATA1 guide the process of megakaryopoiesis (MKp) (Bartley et al., 1994; Kaushansky et al., 1994; Martin et al., 1993). Following changes in the MK development, size, and intracellular compounds in the MK maturation (Machlus and Italiano, 2013), pro-platelets (PLTs) are extended through sinusoidal blood vessels (Italiano et al., 1999).
Although the role of autophagy in MKp has not been precisely elucidated yet, some compelling evidence indicates that autophagy plays an essential role in the regulation of MKp (Cao et al., 2015; Schwertz and Middleton, 2023). Deficiency in the main factors of autophagy would steer to impairments in PLT production and its ultimate function (Ouseph et al., 2015). GATA1, as an important factor in MKp, is able to activate the transcription of genes encoding the essential autophagy components such as microtubule-associated protein 1 light chain 3B and its homologs (Kang et al., 2012).
In this review, we will further investigate the role of autophagy in the regulation of MK differentiation and its importance in thrombopoiesis. Therefore, MKp physiology and its in vitro induction have been discussed. Followingly, autophagy physiology was summarized, and its implications in the MKp and PLT production were evaluated. Furthermore, reactive oxygen species (ROS) production and possible cross-talks between autophagy during MK maturation and differentiation were investigated.
Megakaryopoiesis
In vivo MKp
MKs, similar to other blood cells, originate from HSCs. Bipotent megakaryocytic–erythrocytic progenitor (MEP) cells deriving from HSC would generate MKs and erythrocytes during differentiation (with the addition/loss of some CD markers). At the precommitment stage, the transcription balance between two key genes, that is, KLF1/EKLF and FLI1, drives MEP differentiation (Kwon et al., 2021). KLF1 and other transcription factors such as TAL1 and GATA1 interact with specific regions of genomes, inducing erythroid lineage maturation (Mukherjee et al., 2021; Tallack et al., 2012). In contrast, FLI1 has an antagonizing effect on KLF1 and triggers the transcription of other genes for MK production, such as ITGA2B, GP1BA, GPIX, MPL, and CXCL4 (Kwon et al., 2021).
MKp, similar to other processes of hematopoiesis, first occurs in the yolk sac during both human and murine embryonic stages, which will be followed by hematopoietic sites at the liver and then in the BM, as the main source of hematological cells in adulthood (Davenport et al., 2022; Kaushansky, 2008; Shivdasani et al., 1995). Interestingly, fetal MKs are more proliferative and less polyploid (N ≤ 8) than their adult equivalents, highlighted by augmented expression of genes involved in DNA replication and cell cycle (Davenport et al., 2022). Both external microenvironmental factors and intrinsic gene expression regulation are essential in this transition from fetal to adult MKp (Davenport et al., 2022). This also caused differences in cell signaling pathways, as the inhibition of mechanistic target of rapamycin (mTOR) signaling will only affect maturation, while it would repress both maturation and polyploidy in adults (Liu et al., 2011). The fact might be partly due to the vital demand in fetuses to pool their BM with progenitor cells at a higher rate than adults.
The process of MKp is tightly regulated by a set of transcription factors, including RUNX1, GATA1, FLI1, NFE2, ETV6, MECOM, and GFI1B (Geddis, 2010; Noh, 2021). These transcription factors get activated in a particular order to guide the MK maturation (Noh, 2021). For instance, GATA1 takes part in the differentiation of MEP and the development of both erythrocytic and megakaryocytic lineages; nonetheless, the interaction between GATA1 and FLI1 is required for the final stages of MK differentiation (Noh, 2021; Xu et al., 2024b). Furthermore, these factors are crucial to thrombopoiesis, since RUNX1, FLI1, and NFE2 control PLT formation, granule biogenesis, and expression of surface PLT markers (Xu et al., 2024b).
MKp is also regulated by external signals, that is, the signals initiated by growth factors and cytokines. Key elements recognized for MK differentiation include thrombopoietin (TPO), erythropoietin, granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-6, IL-3, IL-11, and c-Kit ligand/stem cell factor (KL/SCF) (Avraham et al., 1994; Geddis, 2010; Hacein-Bey-Abina et al., 2020; Metcalf et al., 2005). Among these factors, TPO is the main regulator of MK maturation, MK growth, and endomitosis (Kuter and Gernsheimer, 2009). Conspicuously, the constitutional production of TPO is adjusted according to the number of MKs and PLTs (Jelkmann, 2001; Nagata et al., 1997). Its receptor would activate JAK2 and TYK2, as the cytoplasmic kinases, which finally activate mitogen-activated kinase (MAPK) pathway, phosphoinositol-3-kinase (PI3K) pathway, and cAMP response element-binding protein pathway (Kaushansky, 2024). In addition to the abovementioned pathways, recruitment of STAT3 and STAT5 would also enhance the expression of the genes involved in cell survival and proliferation, for example, cyclin D1, p27, p21, and Bcl-xL (Kaushansky, 2003; Kirito et al., 2002).
MicroRNAs have an important role in the differentiation of MEPS into MKs. For instance, miR-150 and miR-34A are two important microRNAs that have been shown to increase through the TPO-cMPL signaling, blocking c-MYB signaling, and pushing the MEP differentiation balance toward the MK lineages (Raghuwanshi et al., 2019). Plenty of other microRNAs were also demonstrated to take part in the lineage commitment of MEP, proliferation, and final release of PLTs (Li et al., 2023; Raghuwanshi et al., 2019). For instance, miR-28A, -708, and -151 could induce MPL and MAPK1 activity and stimulate the final stages of thrombopoiesis (Li et al., 2011). Dimerized STATs are translocated into the nuclei and initiate transcription of p21 and BCL-XL genes (Fox et al., 2002; Kirito et al., 2002). Furthermore, TPO-induced MAPK signaling is another inducer of MK differentiation (Matsumura et al., 1997).
MKs go through several nuclear and membranous events during maturation, such as getting polyploid through a process called endomitosis (Geddis et al., 2001). Important cytoplasmic changes during PLT differentiation are the formation of a demarcation membrane system (DMS) and the production of alpha (α), dense (δ)s, and other MK-specific granules (Bluteau et al., 2009; Mattia et al., 2002). The DMS is continuous with the plasma membrane to interact with the surrounding environment (Eckly et al., 2014).
While the exact mechanism of PLT generation has not been fully elucidated, there are three proposed mechanisms at this stage after MK maturation, that is, pro-PLT formation, cytoplasmic fragmentation, and membrane budding (Furniss et al., 2024). According to the proplastid hypothesis, the PTLs are generated from special cellular extensions organized with the involvement of microtubules during steady-state conditions. Furthermore, during stress-related conditions, cytoplasmic fragmentation with the involvement of internally arranged PLT is hypothesized (Furniss et al., 2024). Finally, the most current model is the direct budding off from the MK membrane, as Potts et al. demonstrated that the pro-PLT model might not be thoroughly responsible for the in vivo PLT production (Potts et al., 2020). Nevertheless, Italiano et al. have reported some MK-related microvesicles that might be mistaken for the shedding PLT from MKs, which necessitates more accurate and meticulous imaging approaches for monitoring PLT generation (Italiano et al., 2021).
In vitro MKp
PLTs can be efficiently produced by in vitro culturing of CD34+ cells, derived from various sources, including peripheral blood, umbilical cord blood (UCB), BM, and induced pluripotent stem cells (iPSCs; Izady et al., 2022; Perdomo et al., 2017). Each unit of extracted HSCs must be expanded and primed to differentiate into MKs; nevertheless, the number of produced PLTs is less than one unit of apheresis PLT production practice (Martínez-Botía et al., 2020; Mookerjee et al., 2020). For instance, only 100 PLTs can be gathered from UCB-derived MKs, whereas a PLT unit contains around 500 billion PLTs, and progenitor cells have to be expanded several times to reach the ultimate number of a typical PLT unit (Karagiannis and Eto, 2015). Therefore, expansion and stimulation methods should be improved to establish a more sustainable resource for PLT production.
For expanding the cells, designing an environment with the supply of necessary growth factors and signaling molecules is indispensable (Meric et al., 2020). Strassel et al. reported that the use of human low-density lipoproteins, cytokine cocktail, and SR1 (antagonist of aryl hydrocarbon receptor) would induce the generation of CD34+CD41low cells and the generation of functional PLTs in bleeding (Do Sacramento et al., 2020; Strassel et al., 2016). In studies on embryonic stem cell (ESC)-derived MKs, culture media containing factors and cytokines such as IL-6, IL-11, TPO, and feeder cells OP9 or C3H10T1/2 stromal cells are used with quite satisfactory results (Eto et al., 2002; Fujimoto et al., 2003; Gaur et al., 2006). Moreover, overexpression of FLI-1, GATA-1, and TAL-1 in ESCs directs them toward the MK lineage (Matsubara et al., 2013). OP9 cells, mice BM preadipocytes, are also another source that has been utilized for PLT production. A significant increase in p45NF-E2 gene expression plays a critical role in MK production from OP9 cells (Matsubara et al., 2013; Ono-Uruga et al., 2021).
Overall, it is feasible that around 1011 MKs and finally 1012 PLTs are released, which is one-third of a transfusion unit. However, there are limitations regarding the lifespan of these PLTs (Sugimoto and Eto, 2021). The researchers have focused on producing immortalized MK progenitor cell lines from iPSCs and PLT production. Thus, the expression of genes, such as BCL-XL, BMI-1, and c-MYC, in iPSC-derived MKs could directly enhance the expansion of immature MKs (Borst et al., 2017; Nakamura et al., 2014; Zucker-Franklin and Philipp, 2000). Nonetheless, it is still many influencing variants that should be addressed, and their full domain of effects becomes elucidated to reach standard protocols of in vitro MK expansion and PLT production with an acceptable rate, efficiency, and cost-effectiveness.
Autophagy and Its Role in MKs
Autophagy biology and significance
Autophagy, as a vital biological process, involves mechanisms that bring about the degradation and recycling of intracellular molecules and organelles (Glick et al., 2010). Autophagy is active at a basal level in most cells and can be upregulated in response to extracellular and intracellular signals such as starvation, lack of growth factor, endoplasmic reticulum (ER) stress, and pathogenic infection (Haas et al., 2015). In general, there are two types of autophagy: nonselective mode, which is practical during stress-related conditions and nutrient starvation, and selective mode, which takes part in removing the determined organelles and proteins (Glick et al., 2010). There are more than 32 ATGs that have been recognized to regulate different stages of autophagy, and most of them are conserved in different species (Nakatogawa et al., 2009).
Three types of autophagy include macroautophagy, microautophagy, and chaperon-mediated autophagy (CMA; Wang et al., 2024; Yang and Klionsky, 2020). In macroautophagy, there is one double-membrane vesicle named an autophagosome, formed in the cytoplasm and merged with a lysosome to produce an autolysosome. In microautophagy, cytosolic compounds are directly taken up by invagination through the lysosome membrane. Conversely, CMA is not directly mediated by forming intracellular membranes, but it finds its targets with chaperone proteins (such as HSC70) and gets recognized by lysosomal-associated membrane protein 2A. This would result in the straight delivery of proteins to the lysosomes (Glick et al., 2010; Yang and Klionsky, 2020).
Among the most crucial autophagy triggers is nutrient starvation. Depletion of all amino acids and serum starvation could strongly induce autophagy in various types of cultured cells. The endocrine system, specifically insulin, regulates autophagy (Mortimore and Pösö, 1987). IL-3 is another regulator that can suppress autophagy through regulating nutrient availability (Lum et al., 2005). Moreover, other factors, such as BCL-2 (Djavaheri-Mergny et al., 2006), ROS (Scherz‐Shouval et al., 2007), calcium, and AMP-activated protein kinase (Høyer-Hansen et al., 2007), and some other factors are considered autophagy regulators. Since the mammalian/mTOR is the master regulator in nutrient signaling, autophagy regulatory signals are influenced by mTOR. Therefore, mTOR inhibitors such as rapamycin or small-molecule enhancers of the cytostatic effects of rapamycin provoke autophagy (Noda and Ohsumi, 1998; Sarkar et al., 2007). The PI3K/AKT class 1 pathway is activated by insulin and growth factors and reacts with mTORC1 to inhibit autophagy (Ge and Schekman, 2014; Yang and Klionsky, 2010). mTOR kinase is suppressed by signals that sense starvation, such as hypoxia and nutrient deficiency. Hypoxia is able to activate autophagy through hypoxia-inducible factor 1 (HIF-1) and also through the HIF-1-independent pathway.
ER facilitates and proofreads the folding of newly synthesized peptides (Braakman and Hebert, 2013; Stevens and Argon, 1999). ER stress, which is caused by various triggers such as protein aggregation, glucose deficiency, hypoxia, and oxidative stress, leads to the accumulation of unfolded proteins in the ER (Haeri and Knox, 2012). Studies show that autophagy is activated in these situations, leading to the deletion of compacted ER and lowering mitochondrial clumps (Hamacher-Brady and Brady, 2016; Kwon et al., 2023; Rashid et al., 2015; Schofield and Schafer, 2021). This will prevent excess consumption of ATP in the ER and ROS production in mitochondria.
Autophagy stages and activation mechanisms
Indeed, there are several successive stages in the process of autophagy. However, these stages can be divided into five main steps: induction, nucleation, elongation and closure, fusion, and finally degradation (Gómez-Virgilio et al., 2022). There are six groups of ATG proteins in these stages, which assist in the formation of isolation membranes, elongation, and lysosomal fusion (Nakatogawa, 2020). Indeed, considering the functional proteins in mammals, effector proteins in autophagy could be classified into 6 groups: (1) the ULK protein kinase, (2) the ATG9 transmembrane proteins, (3) the ATG2/WIPI complex, (4) the PI3K complex, in addition to ubiquitin-like conjugation systems consisting of (5) the ATG12-ATG5-ATG16L complex, and eventually (6) the ATG8 conjugation system (which includes Microtubule-associated protein 1 light chain 3 (LC3) and GABARAP with its ligands in mammals) (Nakatogawa, 2020; Yang and Klionsky, 2010). Key members and regulators of autophagy have been summarized in Table 1.
Key Molecules and Regulators of Autophagy

Colors selected based on the related phase of each molecule.
During normal conditions, mTORC1 maintains an inhibitory effect on the first initiating complex of autophagy proteins through the phosphorylation of its components (Yang and Klionsky, 2010). This complex maintains a kinase activity on the downstream autophagy proteins and is composed of ULK1/2, ATG13, ATG101, and FIP200 (Nakatogawa, 2020; Wong et al., 2013). During starvation and nutrient deficiency, mTORC1 would be constrained in terms of its phosphorylating activity, and this results in higher kinase activity by ULK complex and more accurate localization upon its targets (Wong et al., 2013).
Furthermore, there are other molecules that might regulate the ULK complex, that is, AMP kinase (AMPK), protein kinase A (in yeasts, in vitro studies), and also some likely phosphatases (Wong et al., 2013). Among these factors, the role of AMPK has to be more elucidated, since contradictory findings have been reported for the stimulatory or inhibitory effects of AMPK (Wong et al., 2013). It was considered that under starvation and low energy status, AMPK gets activated and would show its functions via both phosphorylation of mTORC1 (phosphorylation site: S863 on Rptor subunit) and ULK1 (phosphorylation sites: S467, S555, T574, etc.), which would augment autophagy (Alers et al., 2012). Beyond its regulatory effects on ULK, AMPK has been proven to have supporting functions on the more downstream autophagy proteins and to upregulate autophagosome-lysosome fusion (Wang et al., 2022). Nonetheless, glycemic starvation would offset the amino acid starvation and tip the AMPK phosphorylating activity toward the inhibition of ULK complex through the AMPK-liver kinase B1 axis (Kim et al., 2011; Park et al., 2023).
Thereafter, the ATG9 transmembrane proteins, ATG2/WIPI complex, and the PI3K complex help in the progression of the nucleation stage (Lin and Hurley, 2016). This stage is followed by the activity of ubiquitin-like protein conjugation systems (LC3 become processed and activated by ATG4, 7, and 3, then it is recruited by the ATG12/ATG5/ATG16L complex on the surface of autophagosome membrane (Levine and Kroemer, 2019; Van Limbergen et al., 2009)), which drive the expansion of isolation membrane (phagophore membrane) formation and autophagosome cargo loading (Lin and Hurley, 2016). For the trafficking of selected targets of autophagy in mammalian cells, autophagy receptors with ubiquitin-binding domains are recruited, such as p62, TAX1BP1, optineurin, and ND52 (Hurley and Young, 2017; Yim and Mizushima, 2020). The phagophore membrane formation is carried out by the active exchange of lipids and proteins from several intracellular organelles, for example, ER, mitochondria, and Golgi apparatus (Yu et al., 2018).
Autophagy and MKp
Previous reports have shown that knocking out ATG7 in HSCs leads to deficient MKp and eventually deficient hemostasis (Cao et al., 2015). This way, fewer PLTs are produced, and they are often larger than normal and defective in terms of their activity. ATG7 knocking out in MK and PLTs brings out PLT aggregation and hemostasis deficiency, whereas the number and the size of PLTs are normal (Sehgal et al., 2015). K562, a chronic myelogenous leukemia (CML) cell line with the ability of differentiating to the MK lineage, showed loss of this feature after knocking out autophagy genes (Itakura et al., 2008).
In contrast, triggering autophagy is related to higher MK lineage production by these cells. Colosetti et al. described that phorbol-12-myristate-13-acetate (PMA), in addition to SB202190, a p38 MAPK inhibitor, can induce autophagy. The accumulation of LC3-II in the cells treated with PMA and SB indicated the occurrence of autophagy. Meanwhile, the differentiation of cells into MKs was coupled with the upregulation of Beclin-1 and p62/SQSTM1, representing their crucial role in autophagy in this process (Colosetti et al., 2009). Autophagy also operates as a main regulator in the MK maturation and thrombopoiesis. Sun et al. showed that treating the Dami cell line with rapamycin potentially initiated multiple cellular responses, including higher autophagy rate, apoptosis, differentiation, and endomitosis, which is vital for PLT generation (Sun et al., 2020). Baichuan et al. reported the relationship between thyroid hormone level and MK-related autophagy. As they showed, physiological levels of thyroid hormone are necessary for the maintenance of the MKp and thrombopoiesis through PI3K/AKT signaling (Xu et al., 2024a). However, increased levels of the hormone are linked with excessive autophagy and reduced PLT production (Xu et al., 2024a), which might implicate an imbalance in the intracellular pathways of energy production. These preliminary data help to consider an essential role for autophagy in MKp and thrombopoiesis (Fig. 1).
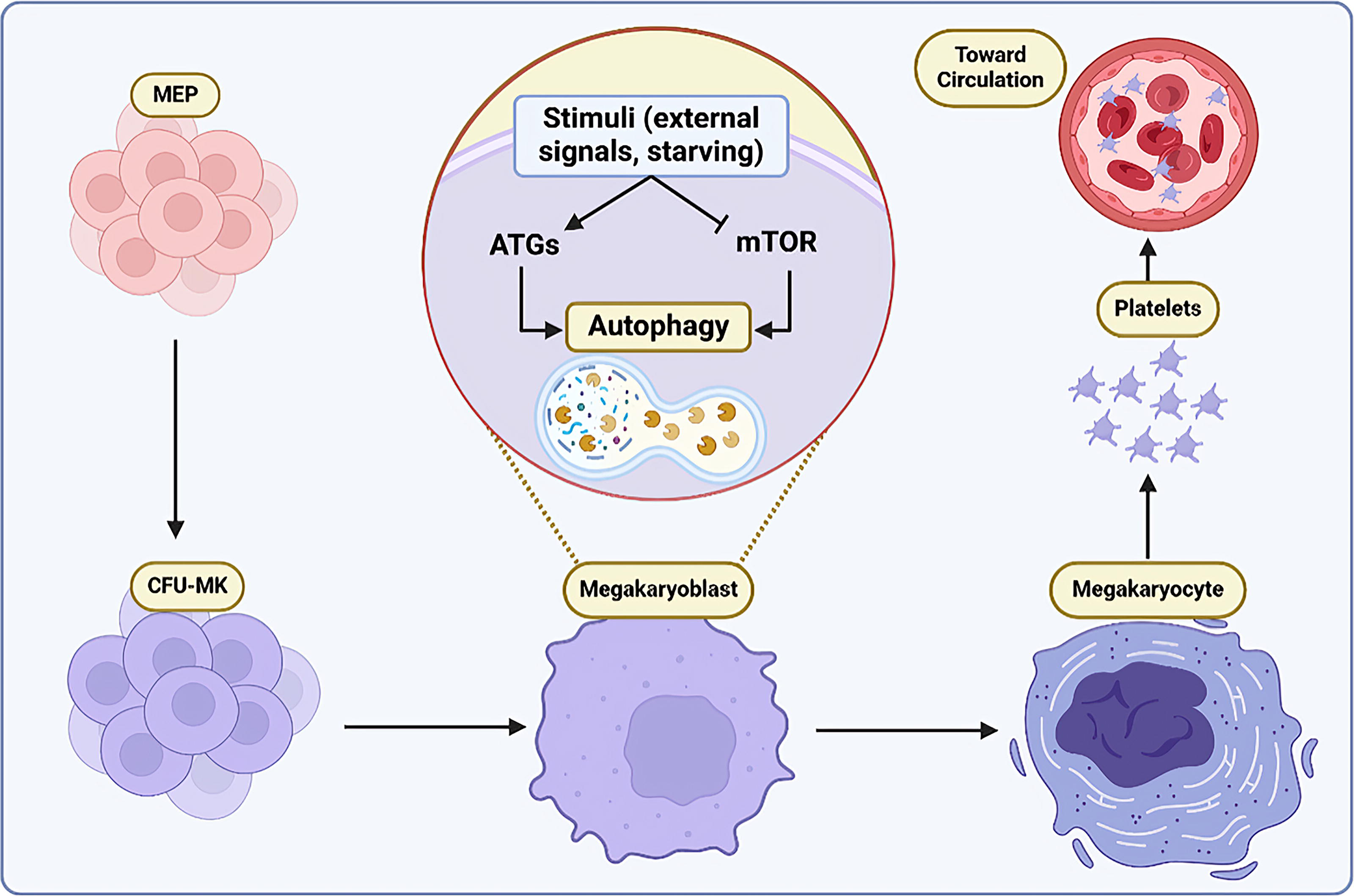
Autophagy effect in MKp: Several stimuli could activate autophagy by inducing ATGs and inhibiting the mTOR pathway. Stimulation of autophagy results in MK maturation and PLT production. ATG, autophagy-related genes; MKp, megakaryopoiesis; mTOR, mechanistic target of rapamycin; PLT, platelet.
Beyond the essential role of autophagy, the presence of an optimal level in MKs is also salient. Wang et al. attested that with regard to this conjecture, both inhibition of autophagy by Bafilomycin A1 and induction by rapamycin in the early stages of MKp were associated with a remarkable decrease in high polyploidy MKs, CD41/CD61 coexpression on the cells, pro-PLT generation, and consequently PLT release (Wang et al., 2017). Nevertheless, Diab et al. assessed the inhibition and activation of the autophagy pathway on the maturation of MKs, produced following the differentiation from human erythroleukemia cells (Diab et al., 2021). They found that autophagy suppression has a noticeable repressive effect on polyploidization and membrane glycoprotein V (GpV) production. In contrast, activation with metformin, while increasing GpV expression, did not induce nuclear maturation (Diab et al., 2021). Based on these findings, it could be supposed that autophagy as a housekeeping function might have versatile effects on MKs, and modulating this pathway should be meticulously assessed in different conditions.
Autophagy and PLT function
Autophagy might have indirect impacts on the PLT activity and function. The function of PLT is a heterogeneous concept, and therefore, it is not possible to draw conclusions easily from the results of various studies due to differences in the methodology. Nonetheless, according to Cao et al. study, impaired autophagy pathway is linked to deficient PLT production and the generation of dysfunctional PLTs (Cao et al., 2015). Murine MK progenitors, BM Lin-cells, had significantly lower markers of MK activity (measured by the activity of acetylcholine esterase), and a smaller amount of differentiated MK (assessed by comparison of CD41+ forward scatter in flow cytometry) from common myeloid progenitor (CMP) was detected in ATG7-deficient mice (Cao et al., 2015). The effect of ATG7 knockout cells might be through the increased mass of mitochondria and resultant uncontrolled internal energy balance, in addition to alteration of the cell cycle with less apoptosis and polyploidy in the process of differentiation (Cao et al., 2015). Further, heterozygote or homozygote knockout of the ATG7 gene was ascertained to be associated with lower levels of P-selectin (CD62P) and limited activity of integrin αIIbβ3 (Cao et al., 2015). This could demonstrate the reduced PLT function following the ATG7 deletion. Aging has also negative impacts on ATG7 (Liu et al., 2024), as Chen et al. discovered that PLTs in the elderly showed significantly lower levels of ATG7 expression than younger individuals (Chen et al., 2024). In another study, Bessa de Sousa et al. found that pathways related to autophagy and its regulation would get dysregulated by aging (de Sousa et al., 2023).
Regarding the relationship of autophagy and PLT activity, Balduini et al. showed that MKs derived from COL6α1−/− mice developed an accumulated population of COL6 polypeptides, which enhanced intracellular stress, apoptosis, and simultaneously restricted autophagosome formation (Abbonante et al., 2023). This would be justified by the increased activity of the PI3K-AKT-mTOR pathway associated with the accumulation of impaired collagen chains in the ER (Abbonante et al., 2021; Grumati et al., 2010). Nonetheless, unlike the effect of ATG deletion on the PLT function, due to the simultaneous elevated function of the store-operated Ca2+ entry (SOCE) mechanism and enhanced activity of the mTOR pathway, the PLTs were more susceptible to activation (Abbonante et al., 2021). The SOCE mechanism might be linked to the ER stress and is recruited to manage the intracellular calcium stores and related signaling pathways (Makio et al., 2024; Zhang et al., 2023; Zhang et al., 2020). Interestingly, starvation and rapamycin could restore the MK autophagy in COL6α1−/− mice model, removing the intracellular stress and accumulated COL6 (Abbonante et al., 2023).
There is also some remarkable evidence of the autophagy role in the pathogenesis of PLT-related disorders. One of the most commonly cited diseases in correlation with autophagy is immune thrombocytopenia (ITP). ITP is defined by abnormal PLT counts and is speculated to stem from autoimmune mechanisms and impaired MK function (Audia et al., 2021; Zufferey et al., 2017). Insufficient thrombopoiesis is reported during ITP and might be caused by the relatively restrained ability of MKs to generate pro-PLTs and lower amounts of TPO (Mititelu et al., 2024). Further, being more prone to apoptosis might also underlie the diminished ability of MKs to stimulate and induce immune tolerance mechanisms (Mititelu et al., 2024).
Some considerable evidence has been published highlighting the upregulated autophagy pathways in ITP. Among these studies, Liu and Mei reported autophagy abnormalities in ITP patients (Liu and Mei, 2018). They compared the effect of adding serum of 14 ITP patients with 23 healthy individuals to the culture media of the MEG-01 cell line, a megakaryoblast cell line, and found that Bax expression was limited, whereas BECN1 expression was upregulated. This denotes an increase in autophagy and lower apoptosis in MKs (Liu and Mei, 2018). Similar findings were reported by Shan et al. as who reported higher BECN1 levels and significantly lower SQSTM1/p62 proteins in the peripheral blood mononuclear cells (Liu et al., 2021). The dysregulated levels would get back to their normal values, whereas the patient achieves remission, implicating a dysfunctional augmented process of autophagy in the ITP-related cells (Liu et al., 2021).
Besides, the mTOR pathway gets upregulated in several immunological diseases such as ITP, and inhibition of this pathway has an ameliorating effect on the disease and PLT production (Shan et al., 2016). It was reported that sirolimus and rapamycin are able to counteract the mTOR inhibitory effect on the MKs and result in positive clinical outcomes (Lv et al., 2022; Shan et al., 2016). Although these findings have recorded an increased level of autophagy indicators, this might be a compensatory response to meet the need for more PLTs. Further, these findings might also denote an upregulated but impaired autophagy, which should become under more scrutiny to find any possible downstream factors lowering the autophagy efficiency and enhancing PLT longevity (Wang et al., 2019).
It should be underscored that part of the autophagy-related effects on thrombopoiesis might be mediated through the role of other cells in the BM microenvironment, including macrophages (Mφ; Wu et al., 2023). Impaired IFI17/TFEB autophagy pathway in ITP-Mφ was indicated to be associated with higher expression of LC3-II/LC3-I, Beclin1, and P62, accumulated autophagosomes, and lower autophagolysosomes, whereas the coculture of MK and treated Mφ restored MK maturation and lysosomal function (Wu et al., 2023). Moreover, it has been reported that the use of sitagliptin would activate the AMPK/mTOR/ULK1 in Mφ, enhancing their autophagy and strengthening the thrombopoiesis. This therapeutic approach showed clinical effectiveness in steroid-resistant and relapsed ITP patients (Wu et al., 2022).
Remarkably, mitophagy also has a key role in the regulation of the PLT function. In the DM patients, plasma glucose levels, circulating lipids, and ROS production result in mitochondrial dysfunction. Thus, normal functioning of this special type of autophagy protects PLTs from greater oxidative stress and rescues them from inappropriate thrombosis (Lee et al., 2016). This might also help in the control of inflammation of the adjacent tissues. Accordingly, Zhang et al. demonstrated that mitophagy within PLTs during ischemic/reperfusion injury would protect the cardiac muscle, as measured by changes in the ejection fraction (Zhang et al., 2016).
Supporting the crucial role of autophagy, excessive enhancement of autophagy stimulation could reduce PLT aggregation to agonists, despite its effects on the maintenance of PLT aggregation and hemostasis (Banerjee et al., 2019). Starving as the main trigger of PLT autophagy can lead to a decline in intracellular calcium, ɑ-and δ-granule release, lower adhesion of PLTs to fibrinogen, and turning off major signaling pathways due to diminished phosphorylation of extracellular signal-regulated kinase (ERK), AKT, and PI3K pathways (Paul et al., 2019).
ROS and MK Maturation
ROSs are applied to a heterogeneous group of small molecules and free radicals, including oxygen atoms with unpaired electrons. ROSs are toxic products of cellular metabolism that are harmful to human stem cells because they damage DNA. However, these molecules play a pivotal role in hematopoiesis (Shinohara et al., 2014). These species are among the main regulators of autophagy.
HSCs and other progenitor cells engage in active cell division for proliferation and subsequent differentiation in the more oxygen-rich vascular niches (Eliasson and Jönsson, 2010; Mas-Bargues et al., 2019). The dynamic oxygen gradient that spans from <1% oxygen concentration in the most hypoxic niches to 6% oxygen in the sinusoidal cavity creates a unique microenvironment that enables the distinct regulatory processes of self-renewal and differentiation to occur (Mohyeldin et al., 2010). These cells and other progenitors are constantly expanded and differentiated in more oxygenated vascular niches (Adams and Scadden, 2006; Ghaffari, 2008). Jang et al.’s study revealed that HSCs producing lower levels of ROS exhibited a more primitive state, characterized by enhanced self-renewal capacity (Bedard and Krause, 2007; Jang and Sharkis, 2007). In contrast, high ROS demonstrates substantial exhaustion upon serial transplantation, accompanied by elevated levels of p38 MAPK and mTOR. Notably, treatment with either a p38 inhibitor or rapamycin effectively restored HSC function in the ROS-high HSC population (Jang and Sharkis, 2007).
MKs are matured in sinusoidal spaces of the BM where the oxygen pressure is higher. Since MKs go through maturation in high-oxygenated environments in the BM niche, it could be concluded that the oxygen level directly correlates with MK maturation (Eliades et al., 2012). This precise spatial arrangement ensures that developing MKs are exposed to the appropriate oxygen conditions for proper maturation and differentiation while also permitting the release of PLTs into the bloodstream via the BM sinusoids (Drayer et al., 2006; Eliasson and Jönsson, 2010). In the Shinohara et al. article, it was declared that a low level of ROS is needed for CMP differentiation into MEP, whereas a higher level of ROS leads to an increase in the expression of CSF1R and granulocyte–monocyte progenitor (GMP) differentiation (Shinohara et al., 2014). In contrast, ROS has an undeniably essential role in the differentiation of MEP to MKs, whereas accumulation of these molecules would result in hemolysis and shortened function of erythrocytes (Shinohara et al., 2014).
Furthermore, an increase of ROS and LPA3, in turn, could result in directing the MEP toward the MK lineage. This event is in coordination with the upregulation of Fli-1, the specific transcription factor of MK lineage (Ho et al., 2015). Similarly, limiting the expression of cytoprotective enzymes in the late stages of MK maturation and PLT production is associated with lower ROS levels and constrained expression of PLT genes, such as signaling lymphocytic activation molecule family member 1, selectin-P, factor 5, and glycoprotein 6 (Motohashi et al., 2010). This is in agreement with Sonia Poirault-Chassac et al. study (Poirault-Chassac et al., 2021), as they found three stages in the mature MKs, starting from a lower ROS level, and followed by the changes in the mitochondrial shape, from round to tubular mitochondria, and activation of Drp1 as the fission activator of these organelles (Poirault-Chassac et al., 2021). Hence, it could be concluded that lower ROS is a programmed step toward MEP generation from CMP. Moreover, following this step, a compensatory ROS elevation is required for MK differentiation and maturation events, such as polyploidy (Fig. 2).
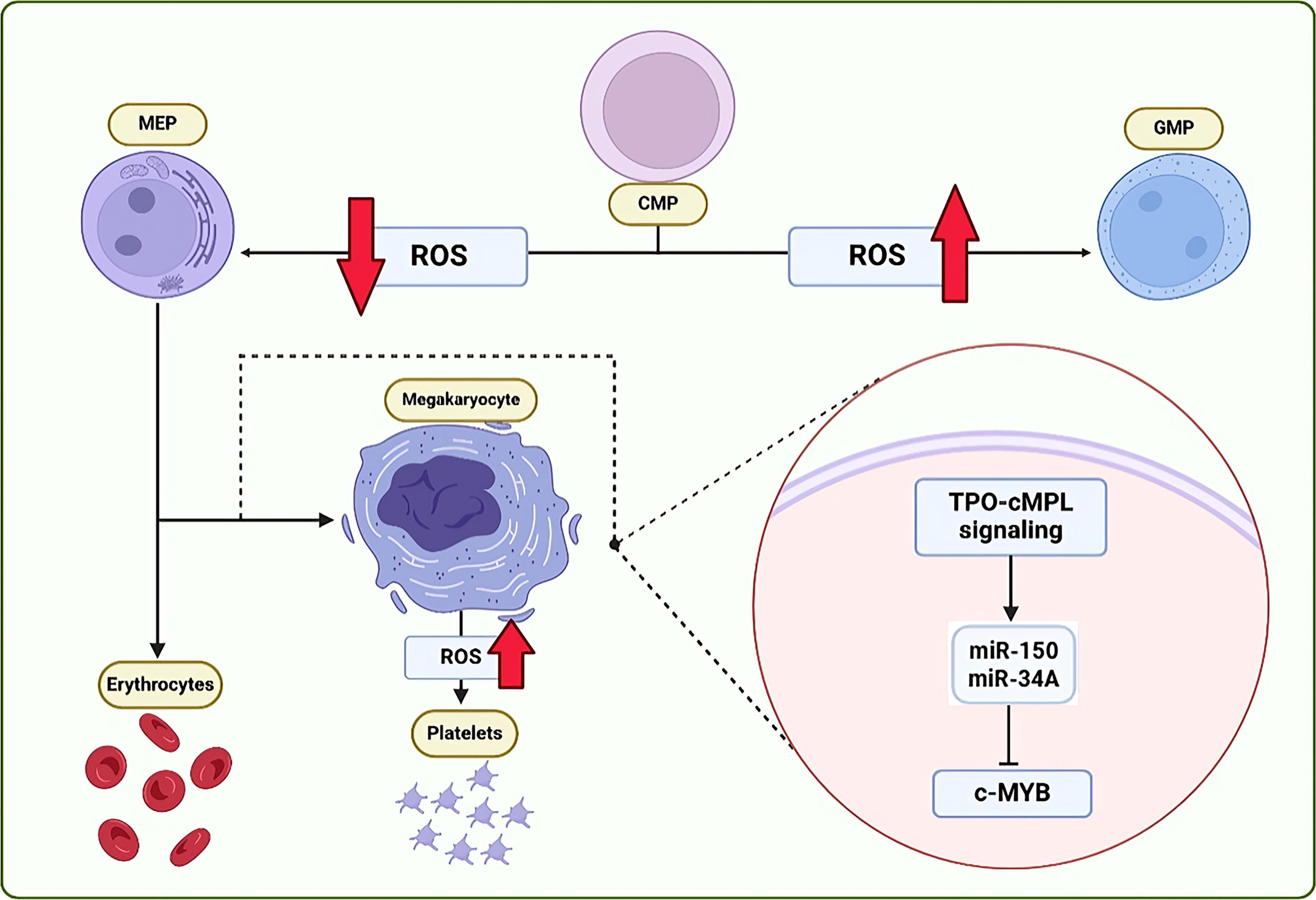
ROS and TPO signaling effects in MKp: A low level of ROS affects the CMP cells to differentiate into MEP cells, whereas a high level has the reverse effect. When TPO-cMPL signaling is induced in the MEP cell, it causes the expression of gene miR-150 and miR-34A to increase, which in turn suppresses the expression of c-MYB gene and leads to cell differentiation toward megakaryocytes. CMP, common myeloid progenitor; GMP, granulocyte-monocyte progenitor; MEP, megakaryocytic-erythropoietic progenitors; MKp, megakaryopoiesis; ROS, reactive oxygen species).
Autophagy and ROS Cross-Talks in MK Physiology
Numerous studies have demonstrated that elevated levels of ROS, a by-product of cellular metabolism, can trigger the activation of autophagy (Chang et al., 2022; Filomeni et al., 2015). Exploring the interactions between oxidative stress, autophagy regulation, and their reparative effects is now an active field of research. Han et al. produced a PLT membrane-based nanoparticle able to scavenge ROS molecules and enhance autophagy in MLE12 cells (Han et al., 2025). This regulation was possibly through the MAPK/BCL2/adenovirus E1B interacting protein 3 (BNIP3) pathway, due to significantly upregulated levels of p38. Subsequent activation of BNIP3 during oxidative stress would also play role in the regulation of cell survival, proliferation, and apoptosis inhibition, in addition to modulating autophagy (Field and Gordon, 2022; Li et al., 2022). This has also been in the process of wound healing and mesenchymal stem cell (MSC) differentiation. Li et al. described a wound coverage based on MSCs and Indocyanine green, a photothermal agent, prohibiting oxidative stress. The coverage was able to enhance autophagy biomarkers through the HSP90 pathway and PI3K inhibition, resulting in MSC differentiation (Li et al., 2025). Anti-inflammatory effects of PLT-related products, including PLT lysate or PLT-rich plasma, could also prove the counterbalance between oxidative stress and autophagy (Ding et al., 2025; Huang et al., 2025; Wang et al., 2025). Fabulously, Yang et al. have also shown that disrupted autophagy through deficient SNAP29 protein would result in severe mitochondrial damage, increased ROS production, and consequent cell death (Yang et al., 2025).
Given that nearly 90% of ROS arises from the respiratory chain within the mitochondrial inner membrane, autophagy emerges as a pivotal player in orchestrating the cellular response to oxidative stress and safeguarding mitochondrial quality control (Kurihara et al., 2012; Redza-Dutordoir and Averill-Bates, 2021). In contrast, several key molecular effectors have been declared to be influenced by increased ROS levels, including Forkhead box O3 (FOXO3), LC3/BNIP3, Nuclear factor erythroid 2-related factor 2 (NRF2), hypoxia-inducible factor 1 (HIF1), BNIP3, and TP53-induced glycolysis and apoptosis regulator (TIGAR) (Li et al., 2015). The abovementioned agents would eventually trigger damaged organelle-targeted autophagy, especially mitophagy, and CMA (Sakurai and Kuwahara, 2025). Hence, it would selectively eliminate damaged mitochondria, thereby ensuring the proper functioning of this vital cellular organelle. Besides, under stress conditions and ROS generation, AMPK, which was discussed earlier as a significant inducer of autophagy, might get activated, resulting in autophagy induction via mTOR inhibition and ULK1 stimulation (He and Klionsky, 2009; Zhang et al., 2025).
Moreover, it was stated in the first part that plenty of microRNAs have been reported to regulate the final stages of MKp and PLT release. Since it was found that miRs have specific effects in regulating autophagy with pathogenic changes in some diseases (Akkoc and Gozuacik, 2020; Xu et al., 2012; Zhao et al., 2019), studying the role of these molecules in the light of making the connection between ROS and autophagy is necessary. Figure 3 summarizes the interactions between autophagy and ROS, regarding PLT generation.
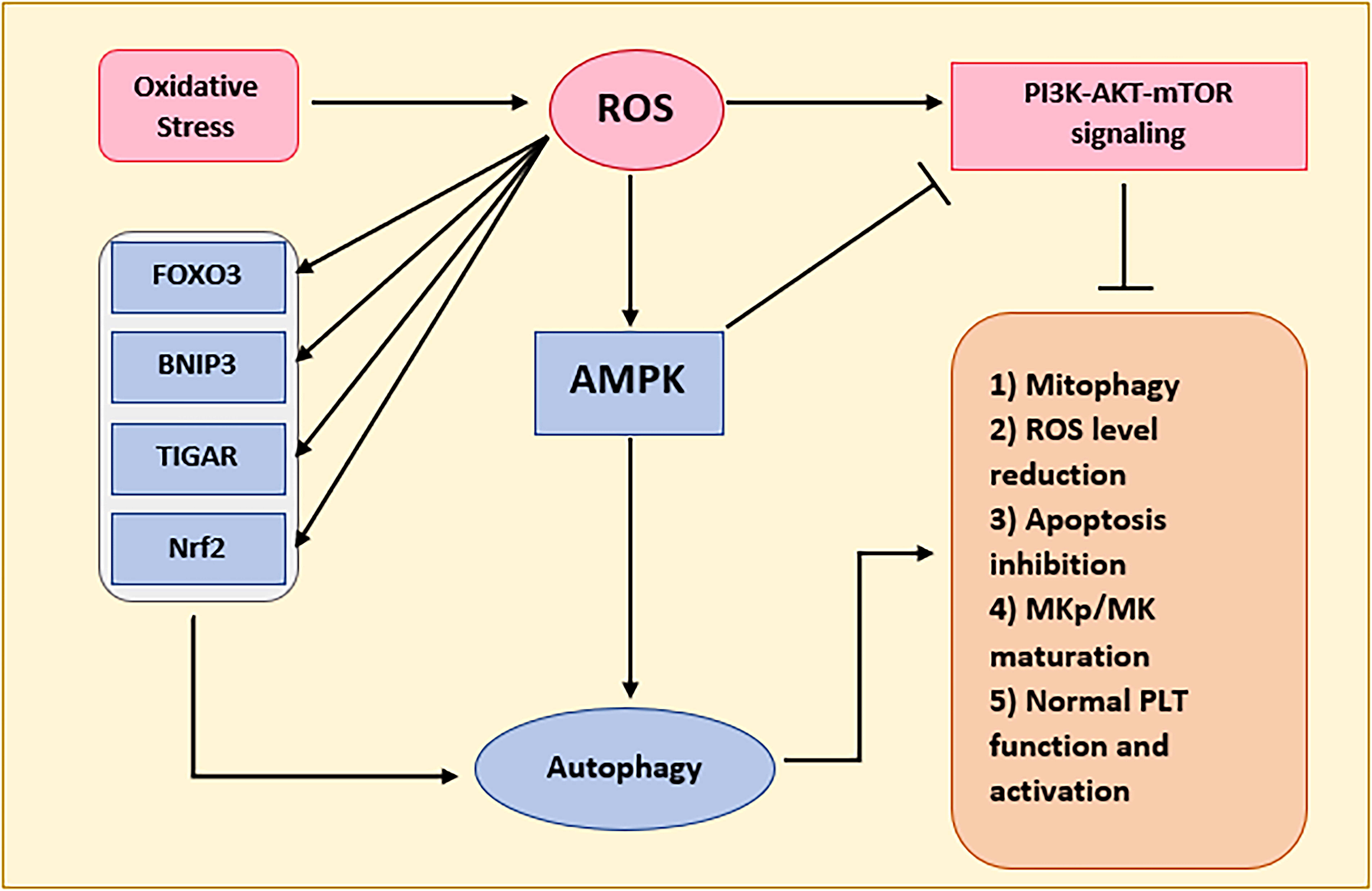
Diagram of the ROS autophagy cross-talk in PLT generation. PLT, platelet; ROS, reactive oxygen species.
Medical Applications for PLT Disorders
Regarding the role of autophagy in the maintenance of a healthy state and also its probable dysregulation in a host of diseases, it has been speculated to use autophagy as a potential target for diseases. In recent investigations, it has been demonstrated that targeting autophagy could play a role in the treatment of several PLT disorders. mTORC1 is pivotal for differentiation and regulation of T cells, and its high expression is seen in MK-related disorders such as ITP. In ITP, activation of the mTOR pathway is increased. It has been observed that this pathway could be an effective approach in ITP patients (Liu et al., 2021; Sun and Shan, 2019).
For example, rapamycin, as the inhibitor of mTOR, could lead to the formation of autophagosomes. In a clinical study, it was observed that rapamycin could be regarded as effective in treating ITP patients (Shan et al., 2016). AZD8055, another inhibitor of mTOR, is used for the treatment of SLE. This medication is capable of inhibiting mTORC1 and mTORC2 and is regarded as a feasible future treatment of ITP (Boya et al., 2005). Decitabine, another inhibitor of the mTOR pathway, with a low dosage, could lead to enhanced polyploidy in MKs and PLT release (Wang et al., 2011; Zhou et al., 2015). Furthermore, imatinib has been shown to cause ATG-dependent autophagy and MK differentiation in CML cells (Yu et al., 2004). Moreover, in another investigation, Feng et al. showed that autophagy disorders could cause impaired aggregation and PLT adhesion (Feng et al., 2014). In addition, Torisu et al. showed that endothelial autophagosomes contain a significant amount of von Willebrand factor (vWF). Thus, knocking down ATG5 and ATG7 could cause inhibition of vWF secretion in vitro, and therefore, it can be considered as a potential treatment for thrombosis (Torisu et al., 2013). This goal could also be achieved through inhibition of autophagy negative regulators. Ming et al. have described that Famciclovir was able to suppress AhR expression and significantly constrained oxidative stress and mitochondrial damage in PLTs. This was linked with lower inappropriate PLT activation and thrombosis (Ming et al., 2025). In summary, studying autophagy and devising agents to modulate this process could be of highly clinical benefit in PLT generation/function disorders.
Discussion and Future Perspective
MKs differentiate from MEPs of BM cells in oxygen-rich regions of the BM under the control of several transcription factors, such as FLI1, GATA1, RUNX1, TPO, GM-CSF, IL-6, IL-3, IL-11, and KL/SCF (Kwon et al., 2021; Lu et al., 2018). In addition to the cytokines within the niche, other intracellular mechanisms such as autophagy and ROS level are also involved in the MK maturation and PLT generation (Eliades et al., 2012; Liao et al., 2025; Schwertz and Middleton, 2023).
Autophagy has different roles in the cells; impaired selective autophagy of the mitochondria might result in aberrant energy balance and ROS production, which could interfere with the maturation of MKs as a result (You et al., 2016). Obviously, flawless autophagy within PLTs is associated with accurate control of intracellular oxidative stress, which evidently becomes influenced by some pathological states (Bedard and Krause, 2007; Lee et al., 2016). It has been shown that the expression of numerous genes such as ATG5, ATG7, Beclin-1, SQSTM1/p62, and LC3 actively participates in different stages of autophagy, whereas dysregulated expression would be linked with autophagy impairment and a significantly reduced PLT release and function (Cherra et al., 2010; Feng et al., 2014; Kim et al., 2016; Liu et al., 2024; Liu et al., 2021; Schwertz et al., 2022; Sharma et al., 2019). Not only the autophagy activation but also its inhibition could have therapeutic effects on major coagulation disorders, as it was found that autophagy inhibition by chloroquine can be applied to prevent hypoxia-induced PLT activation and improper thrombosis (Bandyopadhyay et al., 2025).
ROS has a crucial role in the regulation of stem cells’ growth, proliferation, and differentiation into various cell lines (Bedard and Krause, 2007; Chen et al., 2013; Holzwarth et al., 2010; Mas-Bargues et al., 2019; Mohyeldin et al., 2010; Tiwari et al., 2016). ROS production could activate the AKT signaling pathway, which stimulates HSC differentiation (Herrera et al., 2024). Higher ROS in the niche of BM affects CMPs to differentiate into GMP, whereas lower ROS directs CMPs toward MEP (Eliades et al., 2012; Shinohara et al., 2014). Furthermore, elevated ROS would activate several intracellular signaling pathways and cell cycle-effector proteins, such as AKT, STAT3, STAT5, MAPK, and ERK (Eliades et al., 2012). In contrast, counteracting ROS and oxidative stress within MK progenitors would downplay cyclin D2 and E, while enhancing p27 as the cell cycle inhibitor (Eliades et al., 2012). Studies show that higher oxygen pressure (20%) in the lungs compared with lower oxygen pressure (5%) in the vascular niche causes a higher amount of MK maturation factors to be phosphorylated and activated, and more PLTs are produced (Mostafa et al., 2001; Poirault-Chassac et al., 2021). Indeed, ROS concentration should have a dynamic nature within HSCs to finally promote the release of PLTs. It has been stated that ROS accumulation is necessary for the generation of PLTs from fully matured MKs (Eliades et al., 2012; Poirault-Chassac et al., 2021). By inducing or inhibiting any of these pathways, PLT production can be affected, and it can be effective in the treatment of some PLT-related diseases such as ITP.
While a host of miscellaneous factors could bring about autophagy, oxidative stress and increased ROS are among the main mediators (Hasan et al., 2022; Redza-Dutordoir and Averill-Bates, 2021), which seem to have a physiological role in both MKp, MK maturation, and PLT generation (Chen et al., 2013; Sim et al., 2016; Yang et al., 2022). ROS production is related to the activation of autophagy via several signaling pathways, including ROS-FOXO3-LC3/BNIP3-autophagy, ROS–NRF2–P62–autophagy, ROS–HIF1–BNIP3/NIX-autophagy, and ROS–TIGAR–autophagy (Li et al., 2015). Reciprocally, defective autophagy can also bring about surpassing production of ROS, which might ultimately compromise the function of PLTs (Carresi et al., 2021). Therefore, it could be ascertained that part of the maturation effects of ROS is exerted through the function of autophagy-related molecular effectors.
Nonetheless, it should be admitted the role of autophagy in MKp and PLT function is nuanced, with evidence supporting both its essential and potentially detrimental effects depending on context. Studies show that basal autophagy is crucial for normal PLT production and function, as demonstrated by Atg7-deficient mice exhibiting impaired PLT aggregation, granule cargo packaging defects, and bleeding diathesis despite normal PLT counts (Cao et al., 2015). Conversely, some models reveal that autophagy impairment can paradoxically increase PLT activation susceptibility, as seen in COL6α1−/− mice, highlighting a complex interplay between autophagy, intracellular stress, and signaling pathways such as mTOR and SOCE (Abbonante et al., 2021; Abbonante et al., 2023). In addition, as analyzed in the section “Autophagy and PLT function,” some indicators of autophagy might be increased in ITP patients (Shan et al., 2016; Wang et al., 2019), which could pose a misleading effect in the evaluation of the autophagy role. Resolving these conflicts requires recognizing the autophagy context-dependent role, where both insufficient and excessive autophagy disrupt PLT homeostasis. Thus, to deeper analyze autophagy role in PLT generation, a more standardized approach has to be considered. This includes exclusively revealing methodology, definitions, and assessment tools in different studies (Furniss et al., 2024). Moreover, using a multifaceted analysis regarding different regulators and indicators of autophagy and in different cellular level, that is, gene transcription, translation, and protein synthesis, would elucidate greater and avoid possible confounded results in the analysis and seem to be quite necessary due to the presence of various regulators and multiple steps.
Notably, regarding current advances in cellular and molecular methodologies, it could be supposed that future studies would reveal more and more evidence on the relationship between PLT generation and autophagy. For instance, the application of clustered regularly interspaced short palindromic repeats- and lentiviral-mediated gene editing in MKs has paved the way for rapid screening and identification of essential PLT-related genes (Montenont et al., 2021; Zhu et al., 2018). In addition, the clinical relevance of autophagy within PLTs seems to be flourishing. Aging has been found with meaningfully reduced autophagy flux and altered ATG expression pattern within PLTs, which was also related to lower clot formation and contraction (Chen et al., 2024; Rojas-Sanchez et al., 2024). Besides, in a mouse model of thrombosis, aged PLTs with wild-type mTOR, the negative regulator of autophagy, had lower survival than knocked-out mTOR cases (Portier et al., 2024). These findings bring new hope to consider autophagy as a potential target for PLT disorder (Bandyopadhyay et al., 2025; Lee et al., 2016) or improving the quality of PLT-related products by changing their storage conditions (Zhao et al., 2022). Collectively, it should also be emphasized that gene-editing methods regarding autophagy and ROS cross-talk might go beyond diagnostic and analytical applications toward therapeutic utilization.
The clinical and scientific importance of this subject is to underscore the mutual relationship of autophagy and ROS in physiological states, which could assist in devising novel targets for certain thrombopoiesis and coagulation disorders. Furthermore, producing an in vitro source of perfectly functional PLTs requires a precise recognition of physiological moderators of thrombopoiesis to simulate them in artificial platforms. Future studies should follow a better recognition of these moderators in the artificial PLT production, which could eventually facilitate in vitro PLT production.
Authors’ Contributions
A.K. and R.M.: Designed and conceptualized the article. A.K., R.M., and A.H.: Critically revised the article. A.G., M.P., P.M., M.D., and M.K.: Did the literature review and drafted the article. All authors revised the article and confirmed the final version of the draft.
Footnotes
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Confirmation Statement
All authors confirmed that they are affiliated with an institution that is primarily involved in education or research. It was also highlighted that this research did not receive any form of financial support from any institution/organization for implementation or publication.
Data Availability Statement
This review article does not report new data. All information discussed is derived from previously published sources, which are appropriately cited within the text and reference list. No new datasets were generated or analyzed for this study.
Disclosure Statement
The authors declare that they have no conflict of interest.