
Abstract
Objective:
To evaluate genetic variants within the regulatory regions of interferon regulatory factor 6 (IRF6) and TP63 for the etiology of nonsyndromic oral clefts risk factors.
Design:
We performed allelic transmission disequilibrium test analysis on 5 eligible single-nucleotide polymorphisms (SNPs) and SNP haplotypes using the Family-Based Association Test.
Participants:
The study sample consisted of 334 case–parent trios of nonsyndromic oral clefts from Taiwanese population, separated into nonsyndromic cleft lip/palate (NSCL/P) and nonsyndromic cleft palate only (NSCPO) groups.
Results:
We found all 3 selected SNPs of the IRF6 gene show significant association with nonsyndromic oral clefts (rs2235371, P = 5.10E-07; rs642961, P = .00194; and rs77542756, P = 9.08E-07). Haplotype analyses identified 3 possible SNP combination haplotypes in the IRF6 gene and found that C-G-G showed significant undertransmission (P = .058), whereas 2 other haplotypes, T-G-A and C-A-G (P = 2.71E-06 and P = 5.00E-04, respectively), were significantly overtransmitted to the NSCL/P children but not to the NSCPO children. For the TP63 gene, we failed to detect evidence of nonsyndromic oral cleft association in the 2 SNPs within the TP63 large intron 1 region.
Conclusions:
We used a family-based analysis in 334 Taiwanese case–parent trios to evaluate selected SNPs of IRF6 genes and TP63 genes for a risk of orofacial clefting. This study provides additional evidence for an association between IRF6 and NSCL/P, including the genetic variants within the 5′-noncoding region of the gene. We also confirmed that NSCL/P and NSCPO individuals belong to different groups. For the TP63, our data did not favor the direct involvement of TAp63 isoforms during orofacial development.
Orofacial clefts are common congenital malformations in the human face. Both genetic and environmental factors have been indicated for contributing to its heterogeneous phenotype (Murray, 2002; Murray and Schutte, 2004; Mossey et al., 2009; Dixon et al., 2011). These clefts have commonly been divided into the cleft of the lip only (CLO), cleft of the palate only (CPO), and clefts of the lip and palate (CLP) according to their developmental origin (Dixon et al., 2011; Leslie and Marazita, 2013a). In the past decades, enormous progress has been made in identifying genetic risk variants for nonsyndromic oral clefts, from candidate genes or chromosomal regions (Leslie and Marazita, 2013; Leslie and Murray, 2013) to large-scale genome-wide association studies (GWAS) in different ethnic populations (Beaty et al., 2010; Mangold et al., 2010; Dixon et al., 2011; Ludwig et al., 2012; Beaty et al., 2013; Khandelwal et al., 2013; Sun et al., 2015; Leslie et al., 2016a).
Among the large number of candidate genes thought to contribute to orofacial clefting, interferon regulatory factor 6 (IRF6), which belongs to a family of transcription factors, is the only gene that has shown a convincing degree of consistency across studies. Interferon regulatory factor 6 is a member of transcription factors that consist of a highly conserved helix-turn-helix DNA-binding domain and a less conserved protein-binding domain (Kondo et al., 2002). It may be an important contributor to orofacial development since mutations of the IRF6 gene have been found in Van der Woude syndrome (VWS) and popliteal pterygium syndrome (PPS), 2 disorders that can clinically resemble an isolated cleft lip and palate (Kondo et al., 2002; de Lima et al., 2009; Leslie et al., 2013c). Given the overlapping phenotype of VWS with isolated nonsyndromic oral clefts, in searching the gene for mutations, nonpathogenic variants in the IRF6 gene were significantly associated with nonsyndromic oral clefting in many different populations and ethnic groups (Zucchero et al., 2004; Blanton et al., 2005; Ghassibé et al., 2005; Scapoli et al., 2005; Srichomthong et al., 2005; Park et al., 2007; Vieira et al., 2007; Jugessur et al., 2008; Leslie et al., 2016b).
Recently, 2 studies have linked between p63 and IRF6 and confirmed that IRF6 is the direct target of TP63 during palate development (Moretti et al., 2010; Thomason et al., 2010) by using an experimental mouse model. The p63 protein, another transcription factor, has been recognized as a key regulator in epithelial development, including the development of epidermis, limbs, and craniofacial tissues (Yang et al., 1999; van Bokhoven et al., 2011). In humans, TP63 mutations have been identified in several overlapping disorders, which are characterized by varying combinations of limb abnormalities, ectodermal dysplasias, and facial clefting (van Bokhoven and Brunner, 2002; van Bokhoven et al., 2011; Sutton and Bokhoven, 2017). A mutation in the sterile alpha motif domain of TP63 (p.Asp564His) also implied patients had a nonsyndromic cleft palate only (NSCPO) or a nonsyndromic cleft lip/palate (NSCL/P; Kantaputra et al., 2011).
In clinical assessment, the genetic variants within noncoding genomic regions may provide genotypic information for the apparent risk associated with oral clefts. We attempt to evaluate 3 single-nucleotide polymorphisms (SNPs) within the IRF6 genes and 2 SNPs within the large intron 1 region of TP63 genes for the etiology of nonsyndromic oral cleft risk factors in a Taiwanese cohort using family-based analysis.
Materials and Methods
Participants
Participants were clinically assessed by the plastic surgeon and diagnosed with nonsyndromic oral clefts based on clinical examination, medical records, and a detailed questionnaire from the Chang Gung Craniofacial Center (Taoyuan City, Taiwan). A total of 334 unrelated Taiwanese patient–parent trios were included in this study. Of these, 83 trios have an index case with CPO, 45 have an index case with CLO, and 206 have an index case with CLP. This study was approved by the institutional review board of Chang Gung Medical Foundation (102-1180A3), and written informed consent was obtained from all patients or their parents prior to participation in this study.
Single-Nucleotide Polymorphism Genotyping
Peripheral venous blood samples were drawn from each affected individual and their parents. Genomic DNA was extracted by standard proteinase K digestion and phenol/chloroform extraction using PUREGENE DNA purification kit from GENTRA (Minneapolis, Minnesota). Single-nucleotide polymorphism markers of interest were obtained from the literature review and the SNP database (http://www.ncbi.nlm.nih.gov/snp/). In total, 5 SNPs (3 for IRF6: rs642961, rs77542756, rs2235371 and 2 for TP63: rs4488809, rs10937405) were selected for genotyping. Three IRF6 DNA variants (rs642961, rs77542756, and rs2235371) were selected based on their potential biologic impact on IRF6 expression and function. Single-nucleotide polymorphism rs2235371 was first found as a missense mutation (V274I) located in exon 7 of the IRF6 gene, which is significantly associated with NSCL/P in Asian and Amerindian populations. The SNPs rs642961 and rs77542756 are located within the upstream enhancer region (MCS-9.7) of the IRF6 gene. For the TP63 genes, 2 SNPs (rs4488809 and rs10937405) within the large intron 1 region were included for distinguishing the possible role of TAp63 isoform from the ΔNp63α isoforms in the cleft etiology. Single-nucleotide polymorphism genotyping was performed by using TaqMan Genotyping Assays (either through the Assay-on-Demand service or through the Assay-by-Design service), with an allele-specific hybridization approach implemented on the ABI PRISM 7900 HT Sequence Detection system. Polymerase chain reaction (PCR) amplification was carried out according to the manufacturer’ instructions. Automated allele calling was performed by allelic discrimination plots with the ABI SDS allelic discrimination software version 2.02 from Applied Biosystems (Foster City, California). All variants identified by allelic discrimination assays were further confirmed by directly sequencing PCR products performed by the DNA sequencing core at Chang Gung Memorial Hospital.
Statistical Analyses
All statistical analyses were performed using SAS software, version 17.0 (SPSS Inc, Chicago, Illinois). The allele and genotype frequencies of each marker among samples were compared with χ2 and Fisher exact tests whenever appropriate. Statistical significance was defined at P values <.05. Adherence to Hardy-Weinberg equilibrium (HWE) was assessed for all SNPs using unrelated parents of index patients in this Taiwanese cohort.
The association between each SNP and haplotype with NSCL/P and NSCPO was estimated using Haploview software (http://www.broad.mit.edu/mpg/haploview/). Haplotypes with a frequency <0.03 in these cleft trios were considered rare and thus excluded from statistical analysis. Global P values <.05 were considered significant. Bonferroni corrections for multiple SNPs were applied.
For family-based association analysis, we performed the allelic transmission disequilibrium test (TDT) analysis (Spielman et al., 1993) on every single marker and for haplotypes of multiple SNPs. We implemented the TDT test in PLINK (v1.07;http://pngu.mgh.harvard.edu/purcell/plink/), which provides an allelic TDT assessing whether heterozygous parents transmit the target allele (arbitrarily defined as the minor allele) to the affected child at the expected 50% probability and generating an asymptotic P value for the χ2 statistic (one degree of freedom). The odds ratio and 95% confidence intervals were also calculated. The parent-of-origin analyses were carried out using the transmission asymmetry test (TAT), which is similar to the TDT, but excludes mating between 2 heterozygotes, where transmission can be ambiguous (Weinberg et al., 1998; Weinberg, 1999). Bonferroni corrections were performed for multiple comparisons.
Results
A total of 1002 Taiwanese individuals, including 334 probands and their parents, were recruited for this study. Of these, 83 trios had an index patient with cleft palate (CP), 45 had an index patient with CL, and 206 had an index patient with CLP. We selected 5 SNPs to evaluate these DNA variants for etiology risks of nonsyndromic oral clefts in a Taiwanese cohort using family-based analysis. Three IRF6 DNA variants, rs642961, rs77542756, and rs2235371, were selected based on their potential biologic impact on IRF6 expression and function. Two SNPs, rs4488809 and rs10937405, within the large intron 1 region of the TP63 genes were included for distinguishing the possible role of the TAp63 isoform from the ΔNp63α isoforms in the cleft etiology. The genetic information of each SNP, including the major/minor allele and minor allele frequency, is shown in Table 1. All SNPs had call rates of at least 99.1%, and none of them deviated from HWE genotype frequencies among the Taiwanese population.
Five SNP Information of IRF6 and TP63 Genes for This Study.a
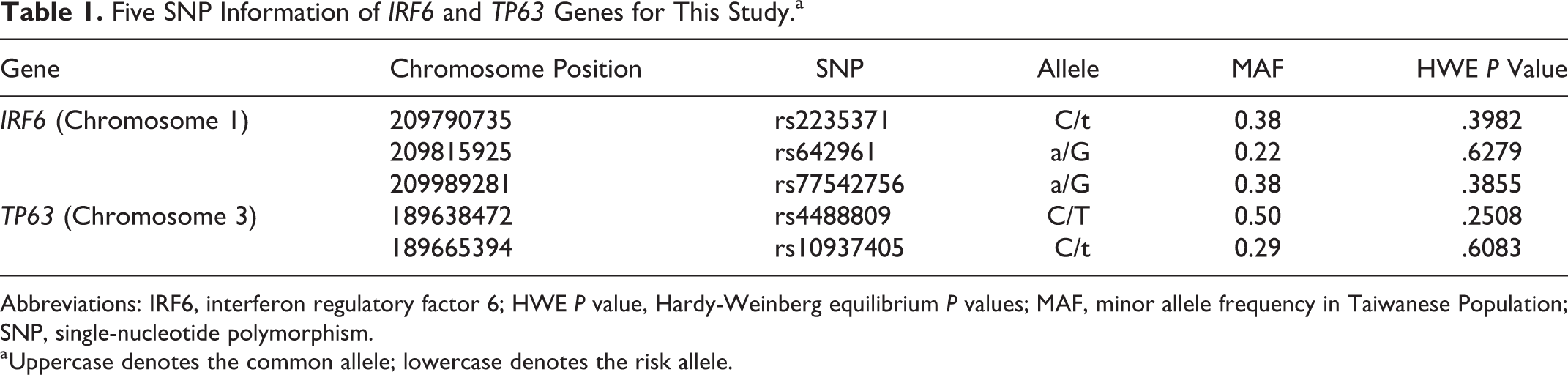
Abbreviations: IRF6, interferon regulatory factor 6; HWE P value, Hardy-Weinberg equilibrium P values; MAF, minor allele frequency in Taiwanese Population; SNP, single-nucleotide polymorphism.
a Uppercase denotes the common allele; lowercase denotes the risk allele.
Table 2 summarizes the results of TDT analysis and family-based association of IRF6 and TP63 with nonsyndromic oral clefts in a cohort of Taiwanese case–parent trios. The CL/P subgroup is a combination of the CL and CLP subgroups. For the IRF6 gene, all 3 selected SNPs show significant association with NSCL/P (rs2235371, P = 2.55E-06; rs642961, P = .0097; and rs77542756, P = 4.54E-06 after Bonferroni corrections). However, none of them were associated with NSCPO (rs2235371, P = .5688; rs642961, P = .1011; and rs77542756, P = .6464). For the TP63 gene, we failed to detect evidence of association with nonsyndromic oral clefts in the 2 SNPs within the TP63 promoter region. Both SNPs did not reach significant value for association with NSCL/P (rs4488809, P = .7453; rs10937405, P = .3317) nor with NSCPO (rs4488809, P = .5529; rs10937405, P = .6326) in these TDT analyses.
Transmission Disequilibrium Test and Family-Based Association Test in Taiwanese Case–Parent Trios.a
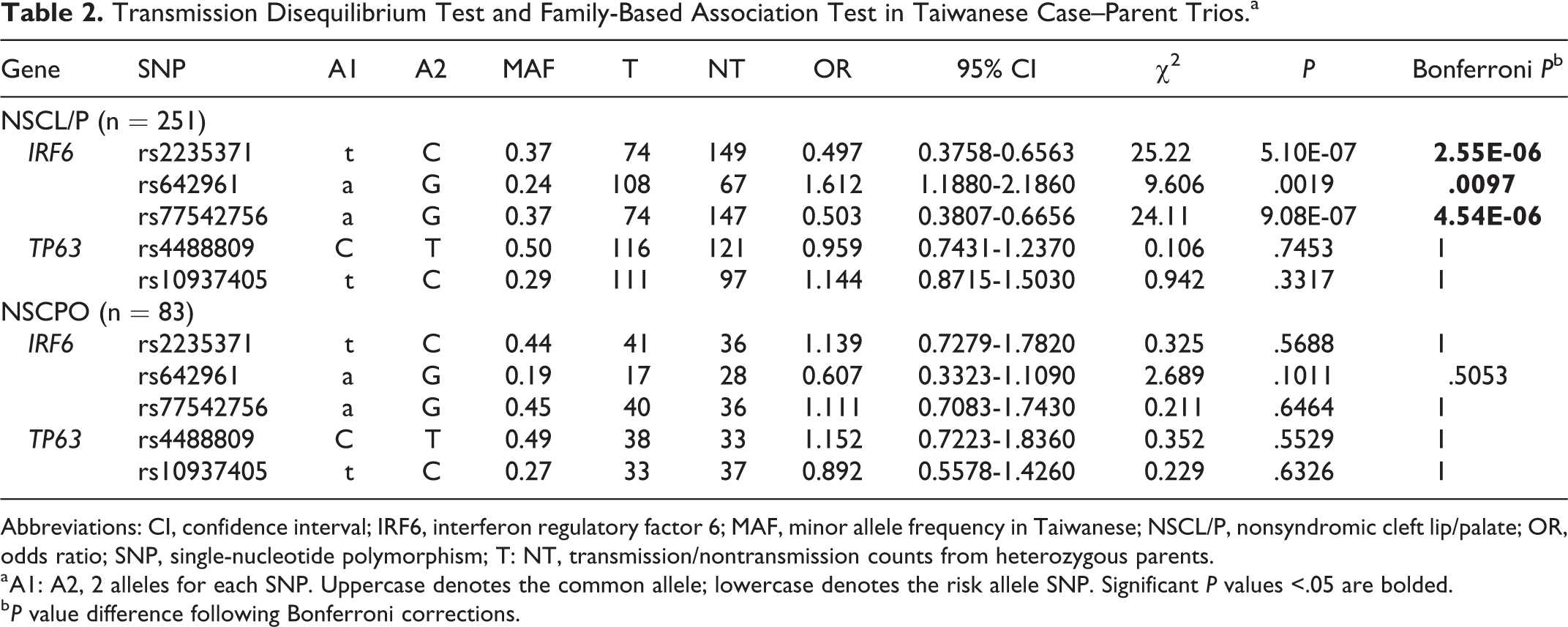
Abbreviations: CI, confidence interval; IRF6, interferon regulatory factor 6; MAF, minor allele frequency in Taiwanese; NSCL/P, nonsyndromic cleft lip/palate; OR, odds ratio; SNP, single-nucleotide polymorphism; T: NT, transmission/nontransmission counts from heterozygous parents.
a A1: A2, 2 alleles for each SNP. Uppercase denotes the common allele; lowercase denotes the risk allele SNP. Significant P values <.05 are bolded.
b P value difference following Bonferroni corrections.
Parent-of-origin effects were also examined using the TAT by separating paternal and maternal alleles. This result only shows evidence of excess parental transmission for SNPs in the IRF6 gene, but not for the 2 SNPs within the large intron 1 region of the TP63 gene. Among the NSCL/P group, our data revealed that almost all 3 IRF6 variants were overtransmitted from both the paternal side (rs2235371, P = .0087; rs642961, P = .0506; and rs77542756, P = .0059 after Bonferroni corrections) and the maternal side (rs2235371, P = 3.61E-04; rs77542756, P = .0011), except rs642961 (P = .3239) shows slightly less significant transmission from mothers (Table 3). In concordance with Table 2, none of these 5 SNPs exhibit association with NSCPO.
Parent of Origin Likelihood Ratio Test for Inequality Between Maternal Versus Paternal Transmission in Taiwanese Case–Parent Trios.a
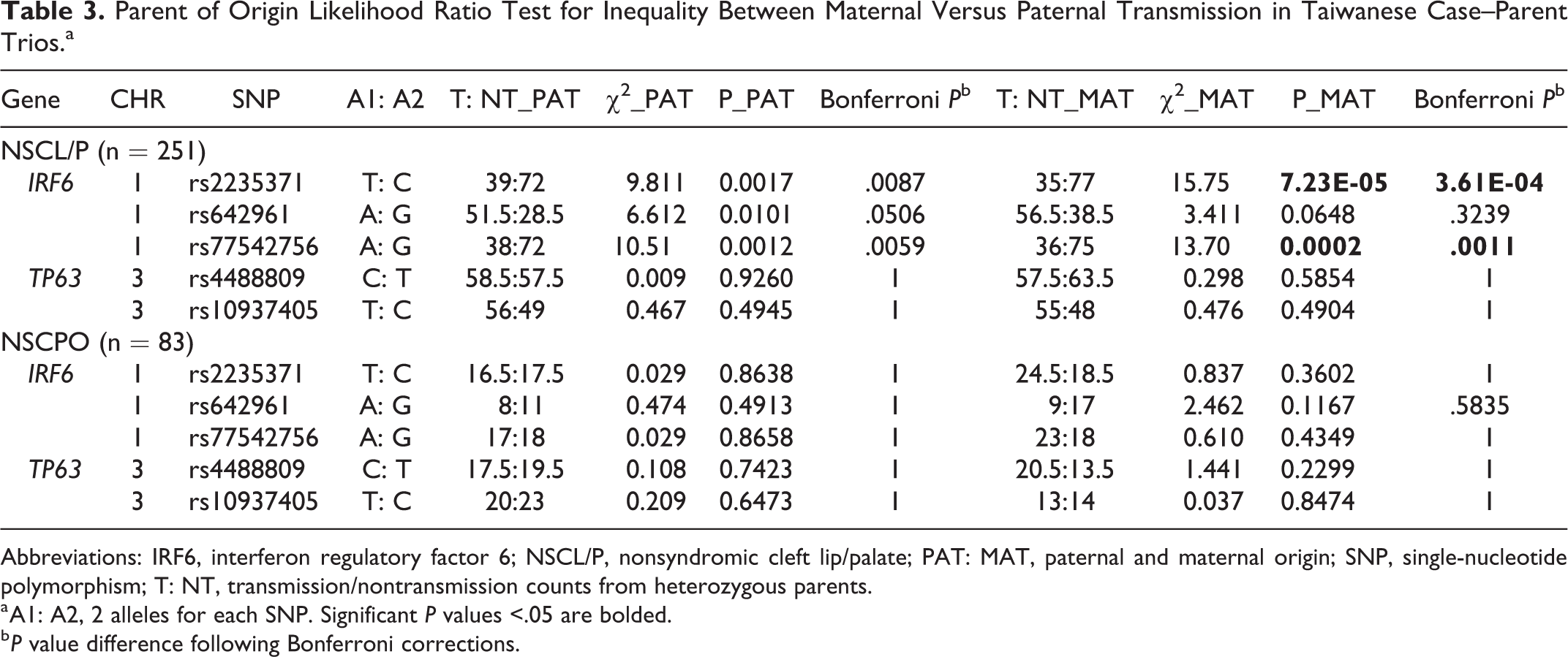
Abbreviations: IRF6, interferon regulatory factor 6; NSCL/P, nonsyndromic cleft lip/palate; PAT: MAT, paternal and maternal origin; SNP, single-nucleotide polymorphism; T: NT, transmission/nontransmission counts from heterozygous parents.
a A1: A2, 2 alleles for each SNP. Significant P values <.05 are bolded.
b P value difference following Bonferroni corrections.
After noticing an association with haplotype-based transmission disequilibrium analyses, we identified 3 possible SNP combination haplotypes (T-G-A, C-A-G, and C-G-G) of 3 SNPs (rs2235371, rs642961, and rs77542756) in the IRF6 gene (Table 4). We found that the most common haplotype (C-G-G) showed significant undertransmission (P = .1743), whereas 2 other haplotypes, T-G-A and C-A-G (P = 8.12E-06 and P = .0015, respectively, after Bonferroni corrections), were significantly overtransmitted to the NSCL/P children but not to the NSCPO children. For the TP63 gene, again, we failed to detect evidence of association with nonsyndromic oral clefts in the 3 haplotypes (C-T, C-C, and T-C for SNPs rs4488809 and rs10937405) within the TP63 promoter region.
Haplotype Analysis of IRF6 and TP63 Gene in Taiwanese Case–Parent Trios.a
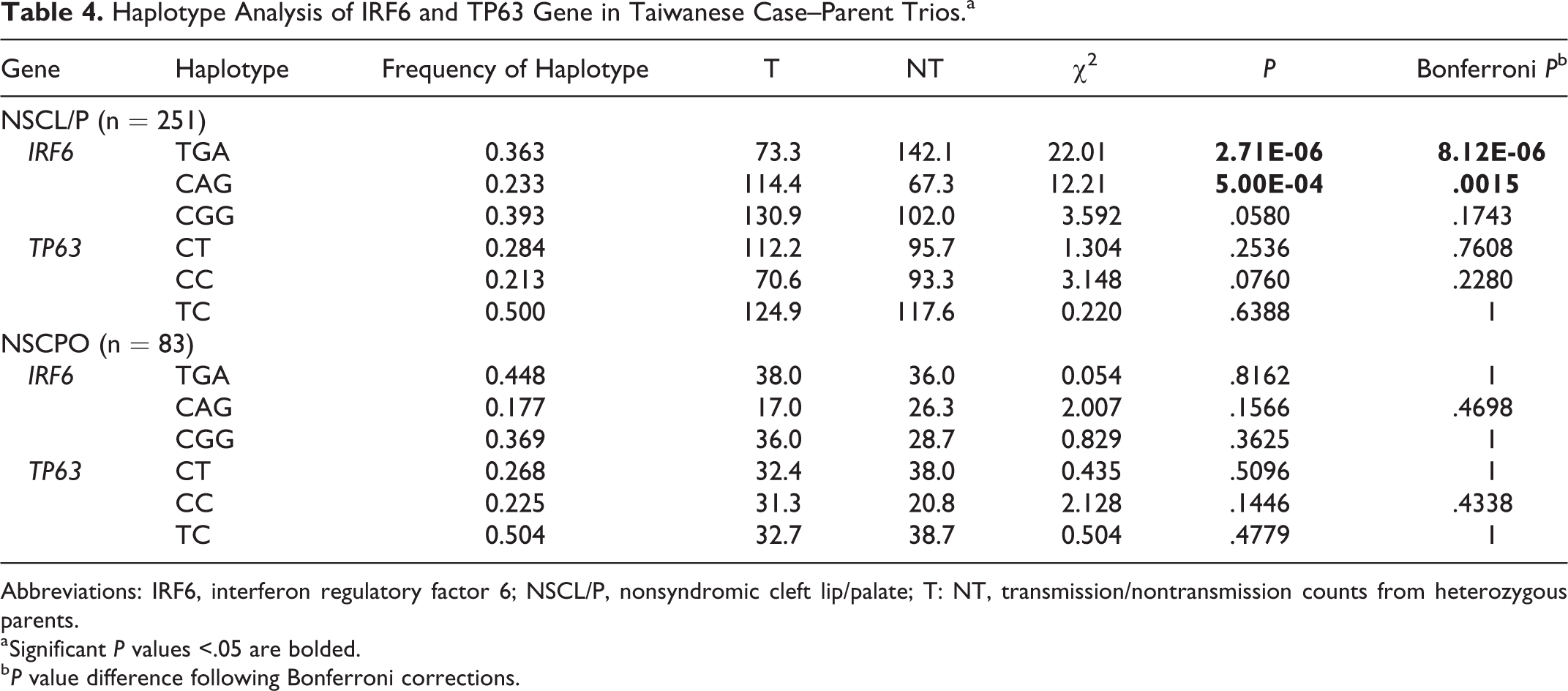
Abbreviations: IRF6, interferon regulatory factor 6; NSCL/P, nonsyndromic cleft lip/palate; T: NT, transmission/nontransmission counts from heterozygous parents.
a Significant P values <.05 are bolded.
b P value difference following Bonferroni corrections.
Discussion
Genetic studies of complex traits in different populations are quite important, both for defining common and population-specific risk alleles and also for understanding the underlying biology of organisms (Dixon et al., 2011). As a part of replication of previously reported findings, we investigated 5 SNPs in the IRF6 and TP63 genes to validate these genetic variants to the risk of nonsyndromic oral clefts in a series of case–parent trios from the Taiwanese population. Association studies based on individual SNPs were generally used to identify novel genetic risk associated with a disease trait. However, the haplotype-based approaches can significantly improve the power of mapping disease genes and provide insight into the dependency among genetic markers (Akey et al., 2001; Shi et al., 2007). In this study, our data provide additional evidence for the association between IRF6 and NSCL/P (rs2235371, rs642961, and rs77542756), especially the genetic variants (rs642961 and rs77542756) located in the upstream noncoding regulatory sequence beyond its transcription start site. The C-G-G haplotype is strongly protective, while the T-G-A and C-A-G are risk haplotypes that show an independent contribution of rs642961 over the 2 other variants. Meanwhile, our study shows no evidence for this association with NSCPO. For the TP63 gene, we failed to detect an association with nonsyndromic oral clefts in the 2 SNPs (rs4488809 and rs10937405) within the TP63 promoter region.
The IRF6 has been suggested to be an important contributor to orofacial development since mutations of the IRF6 gene have been found in VWS and PPS. A role of IRF6 in oral clefts was also supported by analysis of animal models showing that Irf6 mutant mice exhibit a hyperproliferative epidermis that fails to undergo multiple epithelial adhesions for closing the oral cavity, resulting in a cleft palate (Ingraham et al., 2006; Richardson et al., 2006). Single-nucleotide polymorphism rs2235371 was first found as a missense mutation (V274I) located in exon 7 of the IRF6 gene, which is significantly associated with NSCL/P in Asian and Amerindian populations (Zucchero et al., 2004). Regarding the possible function of noncoding regulatory sequences, SNP rs642961 located within AP-2α DNA binding site in the upstream enhancer region (MCS-9.7) of the IRF6 gene has been identified and associated with CL/P in populations of European and Filipino origin (Rahimov et al., 2008). Similar analyses for this noncoding SNP of the IRF6 gene and NSCL/P susceptibility were confirmed in the Central European population (Birnbaum et al., 2009) and in the Chinese Han population (Pan et al., 2010; Zhou et al., 2013); however, analyses showed mixed susceptibility in the Brazilian population present in Brito et al. (2012), but not in Paranaíba et al. (2010) and de Souza et al. (2016). No susceptibility was seen in a non-Hispanic white group study (Blanton et al., 2010), in a Swedish/ Finnish origin group study (Pegelow et al., 2014), or in Iran (Kerameddin et al., 2015). The conflict of replication among these studies may still be due to the difference in patient’ origin and recruiting criteria. Nevertheless, our results favor that DNA variations in the IRF6 noncoding region play a role in NSCL/P development.
Like IRF6, TP63 is also required for the development of the lip and palate in humans and mice (Yang et al., 2006). Two studies have linked an intriguing relationship between p63 and IRF6 and confirmed that IRF6 is the direct target of TP63 during palate development (Moretti et al., 2010; Thomason et al., 2010) by using an experimental mouse model. The TP63 gene is transcribed from 2 alternative promoters, producing TAp63 isoforms with an N-terminal transactivation (TA) domain or ΔNp63 isoforms without the TA domain. The TP63 gene contains versatile regulatory elements in its large intronic regions (Yang et al., 2006; McDade et al., 2012). A very detailed investigation of p63 isoform expression during epidermal development has been carried out in the mouse (reviewed by Mikkola, 2007; Koster et al., 2007). Their experiments have demonstrated that ΔNp63 isoforms are the main isoform expressed at all embryonic stages during epidermal, tooth, and hair development, whereas TAp63 isoforms are not detected until the very late embryonic stage (Candi et al., 2006, 2007, 2008). In order to distinguish the possible role of the TAp63 isoform from the ΔNp63α isoforms in the cleft etiology, we first included 2 SNPs, rs4488809 and rs10937405, within the large intron1 region of the TP63 genes (Miki et al., 2010) to assess the risk of nonsyndromic oral clefts and found no significant association. The data presented here indicate that the TAp63 isoform might not be involved in the craniofacial development directly. Recently, a novel association of SNP rs76479869 within the third intron of TP63 to CL/P has been reported in the genome-wide meta-analyses of nonsyndromic orofacial clefts (Leslie et al., 2017). This might provide another insight into the role of TP63 in the developmental etiology of oral clefts.
Orofacial clefting is a complex birth defect exhibiting significant genetic heterogeneity. Cleft lip and/or palate (CL/P) and CP are 2 distinct groups of orofacial clefts based on their developmental origin (Jiang et al., 2006). Several GWAS and meta-analyses have identified common variants associated with CL/P (Birnbaum et al., 2009; Beaty et al., 2010; Mangold et al., 2010; Ludwig et al., 2012; Leslie et al., 2017) across different populations, which shows different association with CPO (Koillinen et al., 2005; Birnbaum et al., 2009; Beaty et al, 2011; Ludwig et al., 2012; Böhmer et al., 2013; Leslie et al., 2016a; Leslie et al., 2017; Ludwig et al., 2017).
We tested the IRF6 susceptibility of the NSCL/P and NSCPO groups separately and also confirmed that they had different genetic etiology. Both CL and CLP were usually combined a cleft lip with or without cleft palate (CL/P). However, some data suggest that they might have separate genetic pathways (Grosen et al., 2010; Ludwig et al., 2012; Rahimov et al., 2012).
In conclusion, we have performed a family-based analysis of NSCL/P and NSCPO in 334 Taiwanese case–parent trios with the selected SNPs on IRF6 and TP63 genes. This study provides additional evidence for the association between IRF6 with NSCL/P, including the genetic variants within the 5′-noncoding region of the gene. Consistence with previous reports on genetic etiology, we also confirmed that NSCL/P and NSCPO belong to different groups. For TP63, our data did not favor the direct involvement of TAp63 isoforms during orofacial development.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Chang Gung Medical Foundation (CRRPG5C151-3, YHWC).