
Abstract
Objective
The study evaluated the association of BMP4 tag-SNPs and SNP-SNP interactions involving genes active by BMP4 pathway during craniofacial development in the susceptibility of nonsyndromic orofacial clefts (NSOC) in the Brazilian population.
Design
Case-control study.
Setting
Brazilian Oral Cleft Group.
Participants
The study included 881 healthy controls and 800 patients with different types of NSOC: 232 with cleft lip only (NSCLO), 568 with cleft lip and palate (NSCLP), and 274 with cleft palate only (NSCPO).
Interventions
The genomic DNA was genotyped with allelic discrimination assays for five BMP4 tag-SNPs (rs11623717, rs17563, rs2071047, rs2761887 and rs4898820), and analyzed their allelic and genotypic associations using multiple logistic regression. The interactions of these variants with genes involved in the BMP4 signaling pathway, including FGFR1, GREM1, NOG, VAX1 and the 4p16.2 locus, were explored.
Main outcome measures
BMP4 variants in the NSOC risk.
Results
Although only nominal p values were identified when the whole sample was considered, subgroup analysis including the patients with high African genomic ancestry showed significant associations of rs2761887 with risk for nonsyndromic cleft lip with or without cleft palate (NSCL ± P)[(ORhom: 2.16; 95% CI: 1.21–3.85; p = 0.01) and (ORrec: 2.05; 95% CI: 1.21–3.47; p = 0.006)]. Thirteen significant SNP-SNP interactions involving BMP4 and the SNPs at FGFR1, GREM1, NOG and VAX1 and at locus 4p16.2 for increased risk of NSCL ± P were identified.
Conclusions
Our results demonstrate an increased risk of NSCL ± P in Brazilian individuals with enrichment of African ancestry in the presence of the BMP4 rs2762887 polymorphism, and reveal relevant genetic contribution of SNP-SNP epistatic interactions involving BMP4 variants to NSCL ± P risk.
Keywords
Introduction
Nonsyndromic orofacial clefts (NSOC) are the most common congenital malformations in the craniofacial region, with a worldwide prevalence of 1:1000 live births. 1 As the Brazilian population is highly miscegenated, its prevalence varies from 1:650 to 1:2700 live births according to the state or region studied, with the highest prevalence in the southern region, where the European ancestry is predominant, and the lowest in the northeast region. 2 NSOC are divided into three main subtypes, NSCLO (nonsyndromic cleft lip only), NSCLP (nonsyndromic cleft lip and palate) and NSCPO (nonsyndromic cleft palate only). However, there are supporting literature to consider NSCLO as a variant of NSCLP, allowing their combination in NSCL ± P (nonsyndromic cleft lip with or without cleft palate). 3 The NSOC etiology is multifactorial, resulting from the interactions between genetic and environmental factors. 4
Previous studies aimed to explore the genetic components related to NSOC have revealed several genetic variants, mainly represented by single nucleotide polymorphisms (SNP), in candidate genes in different populations. 5 Among them are variants in the bone morphogenetic protein 4 gene (BMP4), one of the members of the transforming growth factor beta superfamily of genes. 6 Different aspects of the craniofacial development are regulated by BMP4, which are related to the processes of proliferation, apoptosis and differentiation. 7 During palatogenesis, BMP4 is expressed throughout the epithelium and ectomesenchyme of the palatine processes, with exception of the posterior region, coordinating the epithelium-mesenchyme interactions. 8 Furthermore, studies on BMP4 during development reinforce the importance of its signaling during palate fusion and emphasize that the loss of function of Noggin, the main antagonist of BMP4, leads to overexpression of BMP4 causing cleft palate due to loss of epithelial integrity.9,10
In addition to animal models demonstrating that BMP4 is a candidate for the etiology of NSOC, association studies and meta-analyses have supported a close relationship between BMP4 and clefts in humans.11–14 The first human case-control study exploring the association of BMP4 variants and the risk of NSCL ± P analyzed the role of rs17563 in a Chinese population, revealing increased odds for NSCL ± P in the presence of the C allele. 15 Overall, the presence of the rs17563 variant C allele was associated with an increased NSCL ± P risk in populations of different origins, including those from China, Chile, India and Iran,11,14,16,17 whereas in the Brazilian population the effect of the allele was protective. 18 This is a very intriguing fact, but it is important to consider that the studies with Brazilian populations were carried out with small cohorts and did not consider the variability of the genomic ancestry of patients. Currently, three meta-analyses have investigated the association of SNPs in BMP4 and the risk for the development of NSCL ± P.19–21 These meta-analyses confirmed the association of risk for NSCL ± P in the Chinese population, while in the Brazilian population a protective effect was found.
Given the conflicting scenario about the association of BMP4 variants in NSOC development, the present case-control study explored the role of tag-SNPs rs11623717, rs17563, rs2071047, rs2761887 and rs4898820 in BMP4 in the development of NSCL ± P and NSCPO in the Brazilian population. SNP-SNP interactions involving genes activated by BMP4 signaling pathway during craniofacial development and NSCL ± P susceptibility were also explored.
Methods
Samples
The sample size estimation was calculated using the Quanto software (version 1.2.4, https://pphs.usc.edu/biostatistics-software/#quanto). The calculation was based with the following parameters: unmatched case-control design, gene only analysis, additive inheritance model, genetic effect of 1.5, minor allele frequency (MAF) of 0.10 (most conservative found in the literature for the BMP4 selected SNPs), two-sided type I error of 0.0125 (corrected for multiple testing using the Bonferroni correction), and Brazilian population risk (prevalence) of 0.00147. With these parameters, the sample required for a power of 80% was 646 per group.
This case-control study included 800 samples of patients with NSCL ± P (232 with NSCLO and 568 with NSCLP), 274 with NSCPO and 881 healthy controls. All patients included in the NSCL ± P and NSCPO groups were affected by orofacial clefts exclusively (nonsyndromic forms). The patients were enrolled in the Brazilian Oral Cleft Group (BOCG), and samples were collected from four associated Centers, which are located in northeast, southeast and south Brazilian regions. In these centers, all individuals with orofacial clefts were carefully screened to detect and exclude those who had the presence of signs and symptoms of syndromes. The individuals with any other congenital defect or cases with possible suspicion of syndromic cleft were excluded. The healthy individuals belonging to the control group were recruited from the same geographic regions of Brazil, and only phenotypically normal individuals and without family history of orofacial clefting were included. The ethics committees of each Center approved this study (approval number 08452819.0.0000.5418), and each patient's written consent was obtained from their parents or guardians and/or participants.
Tag-SNP Selection
The genomic region located 5 kb upstream and 2 kb downstream of BMP4 was screened for tag-SNPs tracking (https://snpinfo.niehs.nih.gov/snpinfo/snptag.html). With data from the 1000 Genomes Project and the criteria of a MAF >0.1, p value in Hardy-Weinberg equilibrium (HWE) > 0.05 and linkage disequilibrium limit (r2) > 0.8, 5 SNPs were selected (Table 1).
Characteristics of the tag Single Nucleotide Polymorphisms (tag-SNP) Frequency of the minor Alleles, Hardy-Weinberg Equilibrium and Genotyping Call Rates.
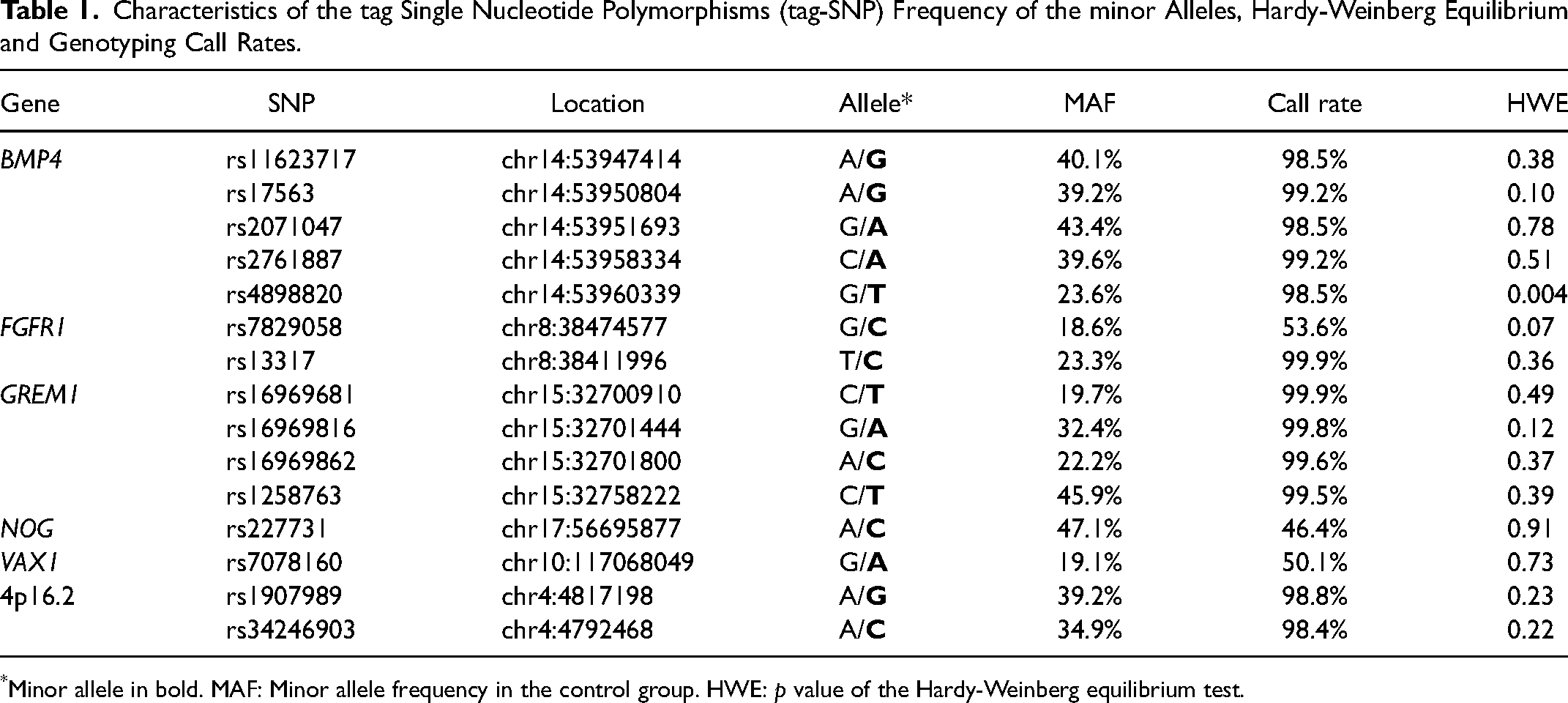
*Minor allele in bold. MAF: Minor allele frequency in the control group. HWE: p value of the Hardy-Weinberg equilibrium test.
Genotyping and Estimation of the Genomic Ancestry
The genomic DNA was isolated from oral mucosa cells using a purification protocol as previously described elsewhere. 22 Genotyping for rs11623717 (C__31531798_10), rs17563 (C___9597660_20), rs2071047 (C___3113060_20), rs2761887 (C__15930504_10) and rs4898820 (C___3113066_10) were performed using custom TaqMan SNP genotyping assay (Applied Biosystems, USA) according to the manufacturer's instructions. The genomic ancestry of each participant of study was determined with an informative panel of the Brazilian ancestry composed of 40 biallelic insertion-deletion polymorphisms (INDEL), after the methods of Messetti et al. (2017). 23
SNP-SNP Interactions
For the analysis of SNP-SNP interactions, SNPs previously analyzed by our group, including rs13317 and rs7829058 in FGFR1,24,25 rs16969681, rs16969816, rs16969862 and rs1258763 in GREM1 (Gremlin-1), 26 rs227731 in NOG, 27 rs7078160 in VAX1 (ventral anterior homeobox 1), 28 rs1907989 (unpublished data) and rs34246903 in the 4p16.2 locus, 25 were selected because they are located in genes known to interact with BMP4 during the craniofacial development, as revealed by STRING (http:/string-db.org) and literature survey.
Statistical Analysis
The existence of HWE in the samples of the control group was calculated using the x2 test with Yates correction. To verify the existence of differences between the groups for sex, the chi-square test was used, and for the proportions of ancestry, the Mann-Whitney test was applied. Multiple logistic regression analyses under additive, dominant and recessive genetic models, taking the genomic ancestry and gender as possible covariates, were performed using the Rstudio program, with the SNPassoc package. The haplotype analysis was performed with the Haplo.stats package in the Rstudio software. Statistical significance after Bonferroni correction for multiple corrections was defined as p < 0.0125. For SNP-SNP interactions, the mbmdr package in R was used, followed by permutation tests (n = 1000 permutations) to eliminate false-positive interactions.
Results
Individual variations in the genetic ancestry proportions were detected, but the groups were not statistically different (Supplementary Table 1). All groups showed a higher prevalence of European ancestry compared to African and Amerindian. Regarding sex, the frequent of males were significantly higher in NSCL ± P (n = 450, 56.2%, p < 0.001) and in NSCLP (n = 326, 57.4%, p < 0.0005) than in the control group (n = 421, 47.8%), whereas in NSCPO the frequency of females was significantly higher (p < 0.01). No significant difference between control and NSCLO was observed. The genotype call rate was higher 98.5% for all SNPs, and, after repeated randomly 5% of the reactions for each SNP, the agreement rate was 100% (Table 1). All polymorphisms respected the HWE, with the exception of rs4898820, which was excluded from further analyzes (Table 1).
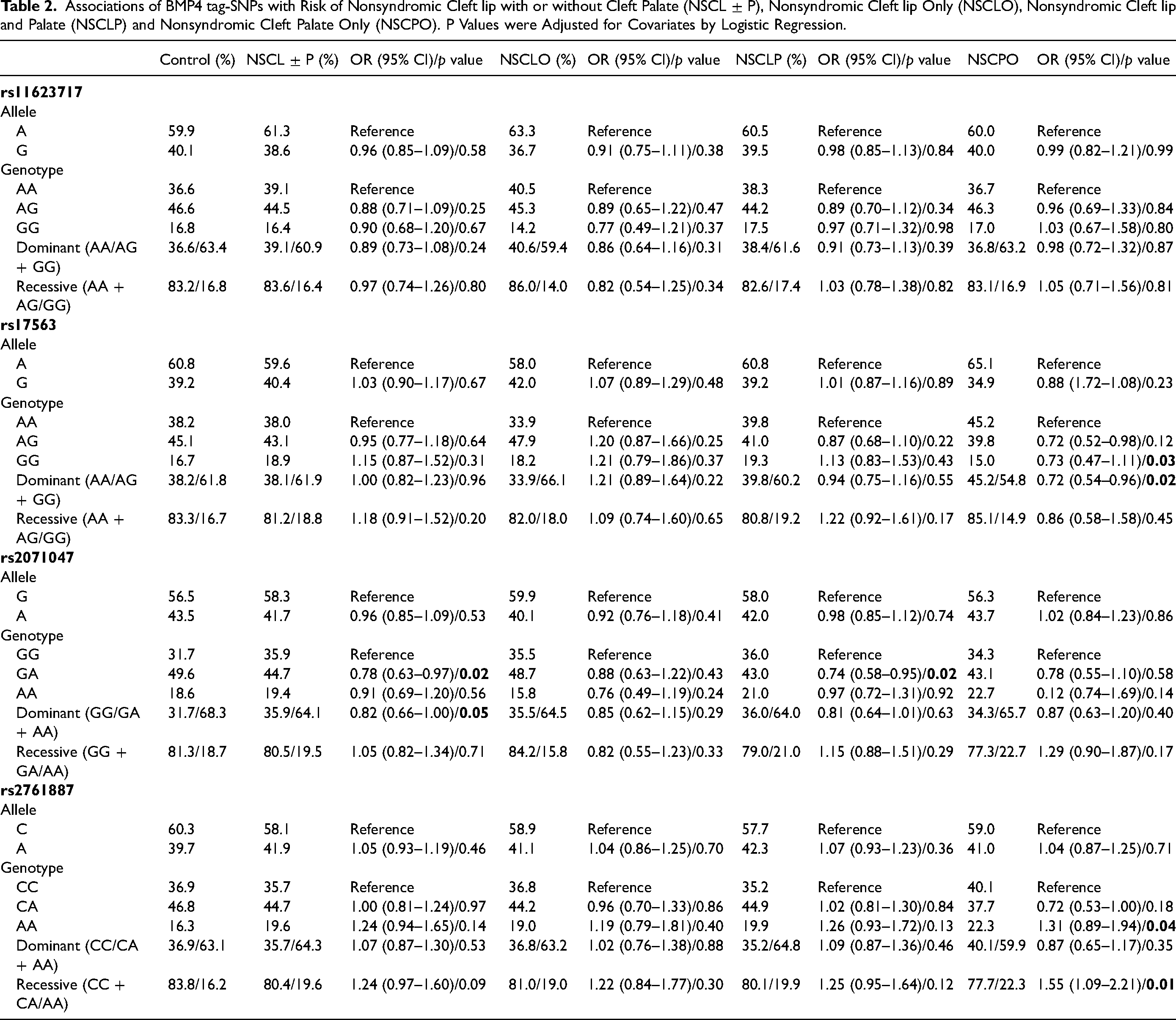
To evaluate the association of the tag-SNPs, allelic distribution and additive, dominant and recessive genetic models were used. In all models, there were no statistically significant associations between SNPs and risk for NSCL ± P, NSCLO or NSCLP (Table 2). However, for the NSCPO group, nominal p values revealed potential association with rs2761887 and rs17563. In the recessive model (CC + CA/AA), rs2761887 generated a NSCPO risk odds of 1.55 (95% CI: 1.09–2.21, p = 0.01), whereas rs17563 showed an OR of 0.72 (95% CI: 0.54–0.96, p = 0.02) in the recessive model and 0.73 (95% CI: 0.47–1.11, p = 0.03) in the dominant model (Table 2).
Associations of BMP4 tag-SNPs with Risk of Nonsyndromic Cleft lip with or without Cleft Palate (NSCL ± P), Nonsyndromic Cleft lip Only (NSCLO), Nonsyndromic Cleft lip and Palate (NSCLP) and Nonsyndromic Cleft Palate Only (NSCPO). P Values were Adjusted for Covariates by Logistic Regression.
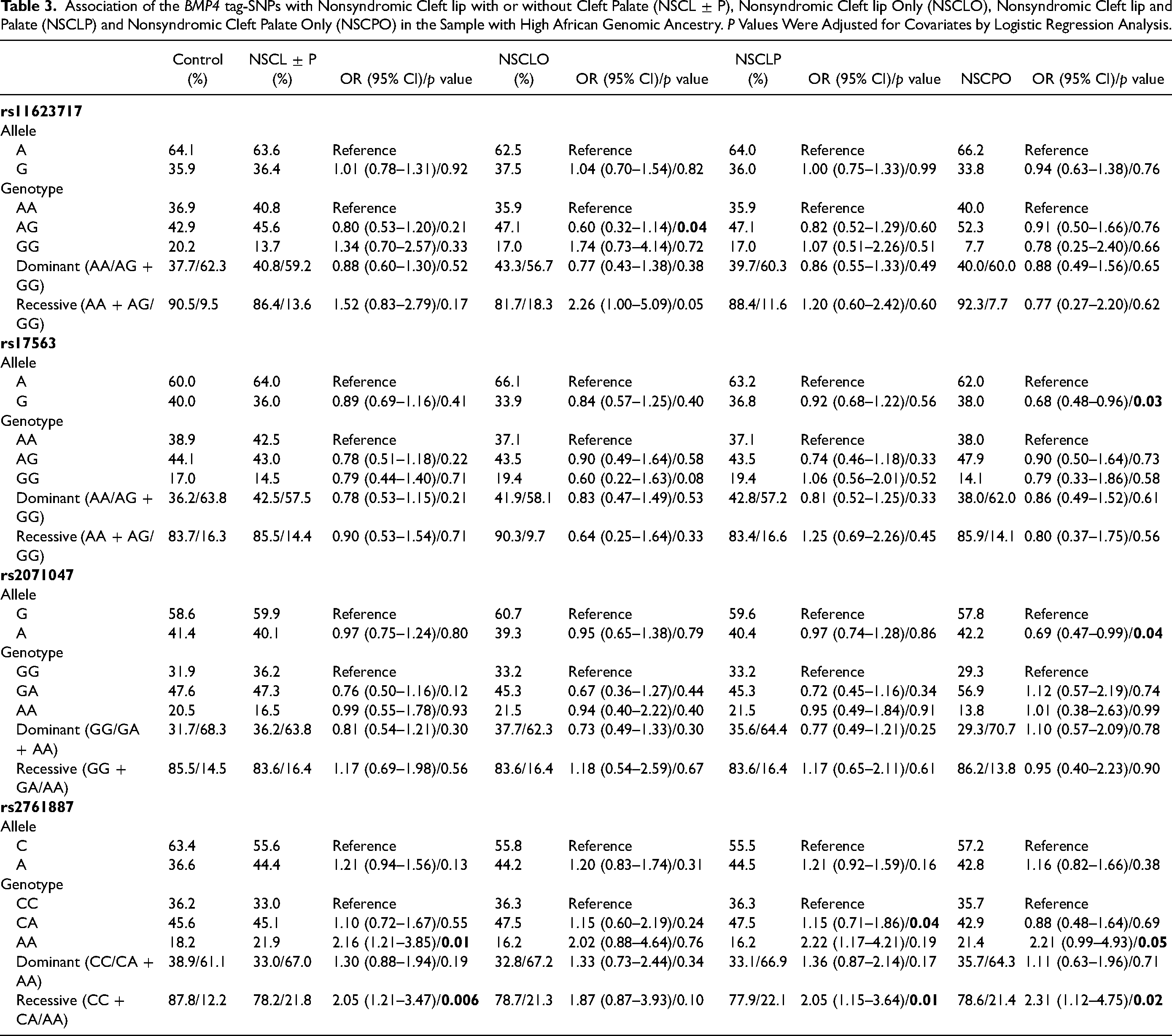
To explore the results according to ancestry, the cohort was split, based on the percentage of European and African ancestry of the samples, in high European ancestry and high African ancestry. No significant associations were found for the sample enriched with European ancestry (Supplementary Table 2). However, for the samples with African enrichment, significant associations were found with the SNP rs2761887. The frequency of the AA genotype was significantly higher in the NSCL ± P group compared to the control group (21.9% and 18.2%, respectively), generating an OR in homozygosis of 2.16 (95% CI: 1.21–3.85, p = 0.01) (Table 3). Using the recessive genetic model (CC + CA vs. AA), the OR for NSCL ± P was 2.05 (95% CI: 1.21–3.47, p = 0.006) (Table 3). The analysis stratified by NSCLO, NSCLP and NSCPO in African enrichment group showed only borderline associations for the SNPs rs11623717 and rs2761887 (Table 3). Association analyses of the haplotypes with NSCL ± P and NSCPO susceptibility were performed with the combination of 2, 3 and 4 SNPs, but no statistically significant associations were found (Supplementary Table 3). The stratified analysis of the samples by sex showed no significant results (data not shown).
Association of the BMP4 tag-SNPs with Nonsyndromic Cleft lip with or without Cleft Palate (NSCL ± P), Nonsyndromic Cleft lip Only (NSCLO), Nonsyndromic Cleft lip and Palate (NSCLP) and Nonsyndromic Cleft Palate Only (NSCPO) in the Sample with High African Genomic Ancestry. P Values Were Adjusted for Covariates by Logistic Regression Analysis.
We performed pairwise SNP-SNP interaction analyses between the SNPs in BMP4 and SNPs in FGFR1, GREM1, NOG and VAX1 and with a SNP at the 4p16.2 locus, which is a potential regulator of genes such as EVC, EVC2 and MSX1 (Ingersoll et al., 2010). Thirteen pairs of SNPs demonstrated significant interactions after correction of the p value with 1000-permutation test (Table 4). Our results found highly significant interactions between 3 BMP4 SNPs with VAX1 after correction: rs11623717-rs7078160 (p < 0.001), rs2071047-rs7078160 (p < 0.001), rs2761887-rs7078160 (p < 0.001). Furthermore, the interactions that occurred between VAX1 and BMP4 generated the highest numbers of risk genotypes (from 4 to 5). Significant interactions were also detected between rs11623717 (BMP4) and rs227731 (NOG) (p = 0.006), rs2761887 (BMP4) and rs13317 (FGFR1) (p = 0.004) (Table 4). The interactions involving rs11623717 (BMP4) and rs7829058 (FGFR1) (p = 0.033) and rs17563 (BMP4) and rs3741442 (12q13.13) (p = 0.046) resulted in borderline p values (Table 4). The interactions that occurred between the polymorphisms rs16969816 (GREM1), rs1907989 (4p16.2), rs7829058 (FGFR1) and rs11623717 in BMP4 were only between risk genotypes, but only the interaction between rs7829058 (FGFR1) and rs11623717 (BMP4) was able to be remain significant after 1000-permutation correction.
SNP-SNP Interactions among BMP4, FGFR1, GREM1, NOG, VAX1 and 4p16.2 Assessed by the Model-Based Multifactor Dimensionality Reduction (mbmdr) test.
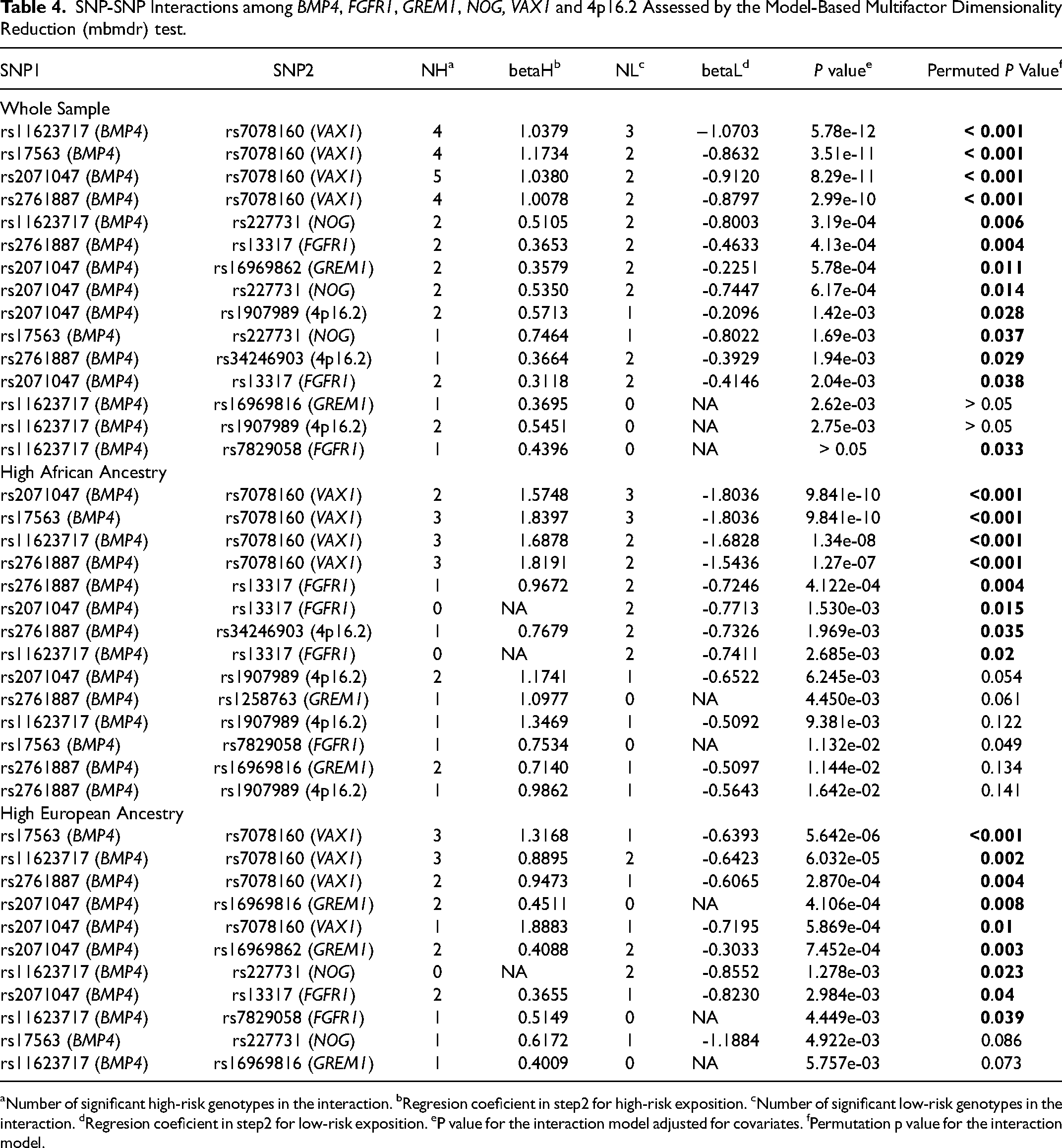
Number of significant high-risk genotypes in the interaction. bRegresion coeficient in step2 for high-risk exposition. cNumber of significant low-risk genotypes in the interaction. dRegresion coeficient in step2 for low-risk exposition. eP value for the interaction model adjusted for covariates. fPermutation p value for the interaction model.
Exploring the ancestry differences, we observed nine significant interactions in the sample with high European ancestry and other nine in the high African ancestry. In both, the most significant interaction involved rs7078160 in VAX1 (Table 4). To the group with high African ancestry, significant interactions between rs2761887 (BMP4) and rs13317 (FGFR1) and borderline p values to SNP-SNP interactions between BMP4 and FGFR1 and between BMP4 and 4p16.2 were detected. In addition, significant interactions involving rs2071047 in BMP4 and GREM1 (rs16969816 and rs16969862) were found only in high European ancestry (p < 0.008).
Discussion
The miscegenation of a population can influence different aspects that involve the studies with NSOC, so it is necessary to consider the impact of miscegenation of the participants in the design of the studies. For example, the variation in the prevalence of different cleft types can vary in the population when comparing an immigrant group versus the local ethnicity of the country, as the immigrant group shows NSOC prevalence in similar rates of the ethnicities of their countries of origin. 29 Another influence driven by miscegenation is related to the genetic aspects of NSOC susceptibility, as different genes/alleles may cause both protective or risk effects depending on the ethnic ancestry of the evaluated population. 26 In the present study, the polymorphisms rs11623717, rs17563, rs2071047 and rs2761887 did not show relevant associations with NSCL ± P, NSCLO and NSCLP in an analysis without division by ethnic ancestry, but taking into account the miscegenation data, a significant association between rs2761887 and the risk of NSCL ± P was detected in patients with high African ancestry. In addition, this same SNP revealed a nominal p value for the risk of NSCPO in patients with high African ancestry, revealing the importance of ethnicity in studying the association of genetic variants and NSOC.
The gene-gene interaction analyses can provide important predictive information about the formation of highly complex multifactorial diseases such as NSOC. 30 It is well described in the literature that the effect produced by SNPs in isolation on the phenotypes of complex diseases can be masked by the results of epistatic alterations resulting from the dynamics of interactions between genes, in addition to the influence of non-genetic confounding factors. 31 Our results show strong interactions between the BMP4 rs11623713, rs17563, rs2071047 and rs2761887 and rs7078160 located in the VAX1. VAX1 encodes a transcriptional regulator, which exerts essential functions for human craniofacial development, and has already been associated with the development of NSCL ± P. 32 In cultured cells derived from the optic vesicle of mice silenced for the Shh gene, a negative regulation of BMP4 on VAX1 was observed. SHH is a gene that acts antagonistically to BMP4 during development. 33 Like rs2761887 in BMP4, the rs7078160 variant of VAX1 was also associated with the formation of NSCL ± P in a Brazilian population with enrichment of African ancestry. 28
After the genome-wide association studies by Beaty et al. (2010) 34 and Mangold et al. (2010), 35 which originally identified a NSCL ± P risk locus on chromosome 10q25.3 that encompasses an intergenic region with suggested regulatory effects on adjacent genes, specifically VAX1, many other association studies have demonstrated the association of VAX1 genetic variants with NOSC susceptibility in different populations.36–40 However, functional studies connecting VAX1 with orofacial clefting are scarce. Hallonet et al. (1999), 41 after the generation of VAX1 knockout mice, demonstrated that homozygous mutants exhibited craniofacial malformations, including cleft palate. A recent study revealed additional VAX1 variants in association with NSCL ± P in the Western Han Chinese population, which are located in expression quantitative trait loci in multiple tissues and affect VAX1 binding ability to DNA. 42 However, Geoghegan et al. (2017) 43 brought new insights to the connection between VAX1 and cleft palate by demonstrating that though VAX1 knockout mice develop cleft palate, VAX1 is not expressed in the second palate during palatogenesis, and the cleft palate phenotype is indeed indirectly associated with VAX1 due to a reduction in VAX1-induced sonic hedgehog (SHH) pathway activation. It is reported that SHH pathway can downregulate BMP family members during craniofacial development, 44 suggesting a loop among VAX1-BMP4-SHH during palatogenesis.
Another positive SNP-SNP interaction involved rs227731 in NOG. This SNP, which was reported as a risk factor for NSCLP in a Brazilian population with enrichment of European ancestry, is located in an intergenic region, with regulatory effects on NOG. 27 Noggin, the NOG gene encoded protein, is recognized as a specific antagonist of BMP4, as it prevents BMP4 binding to its cellular receptor. 45 He et al. (2010) 10 investigated the role of Noggin and BMP signaling during palatogenesis in NOG mutant mice, revealing that Noggin inactivation leads to cleft palate, as the inhibition of BMP4 is essential for maintaining palatal epithelium integrity. Olivares-Navarrete et al. (2014), 46 when evaluating NOG silencing in human osteoblast-like MG63 cells, reported a strong increase in BMP2 and BMP4 levels promoting both osteoblast and osteoclast differentiation.
The rs2761887 and rs2071047 polymorphisms in BMP4 showed significant interactions with rs13317 and rs7829058 of FGFR1. The FGF-signaling pathway is part of the multiple signaling pathways related to palate development, and inadequate signaling in this pathway can contribute to up to 5% of NSOCs. 47 During osteogenic differentiation, FGF1 and FGF2 expression markedly reduces early markers of osteogenic compromise, such as BMP4, acting in antagonistic ways to BMP4. 48 Fgfr1−/− mice are characterized by delay on proliferation of neural crest cells, impairing the development of nasal and medial processes and the elevation and fusion of the palatal crests. 49 The FGFR1 rs13317 was associated with protection in uneventful bone repair in the Brazilian population, while haplotypes containing rs17563 are linked to failures in adequate bone repair by interfering with bone homeostasis. 50 Furthermore, SNP-SNP interaction between haplotypes of rs2761884 in BMP4 and rs4631909 in FGF3 was associated with protection from the occurrence of peri-implantitis in the Brazilian population. 51 Previous results revealed that rs13317 in FGFR1 reduces the risk of NSCL ± P in the Brazilian population, 25 however no association was found for rs7829058 in the same population. 24
The GREM1 rs16969862 interacted significantly with rs2071047 in BMP4. Although GREM1 did not demonstrate craniofacial effects during embryogenesis in functional studies with Grem1-deficient mice, this gene is reported to be a classic antagonist of BMPs. 52 GREM1 may show different levels of binding affinity with BMPs in renal epithelial cells, for example, with GREM1 having a preference for BMP2 and BMP4 over the other BMPs. However, it is worth mentioning that this effect was only visualized in vitro. 53 In a previous study in the Brazilian population, the GREM1 rs16969862 was associated with a reduced risk for the development of NSCL ± P. 26
The 4p16.2 locus containing rs1907989 and rs34246903 showed significant associations with rs2071047 and rs1761887 in BMP4. The 4p16.2 locus, which contains EVC and EVC2 genes, was previously associated with syndromes that affect the craniofacial complex in humans, 54 and the rs34246903 variant has already been associated to a lower risk of developing NSCLO in the Brazilian population. 25 Functional interactions between EVC and EVC2 with BMP4 have not yet been described in the literature, however, 4p16.2 locus is located close to the MSX1 gene (Msh Homeobox1). 55 The importance of MSX1 for occurrence of orofacial clefts is well recognized, and MSX1 has already been shown to activate BMP4 expression in the mesenchyme during dental morphogenesis. 56 Loss-of-function mutations in MSX1 in humans lead to tooth agenesis of molars and second premolars. 57 Interestingly, our study found a significant interaction between 4p16.2 and BMP4, but the same was not true for the interaction between rs62636562 in MSX1 and BMP4 (data not shown). Machado et al. (2022) 25 did not rule out the possibility that the 4p16.2 rs34246903 is in linkage disequilibrium with other polymorphisms, segregating together with a still unknown causal variant for orofacial clefts or presenting regulatory effects on nearby genes, such as MSX1. More efforts are needed to investigate this interaction between SNPs at loci 4p16.2 with tag-SNPs in BMP4 to clarify the mechanisms by which it is related to the development of NSOC.
The study has limitations and strengths. The limitations include those related to sample size, particularly the reduced number of NSCPO, though rs2761887 and rs17563 in BMP4 demonstrated potential associations for increased risk of NSCPO at nominal p values, and the reduction in the power during stratification analyses. We can also highlight as possible limitations the interaction tests of only a few SNPs in the genes belonging to BMP4 signaling pathway, the lack of characterization of impact of SNPs on function of the encoded proteins, and the absence of environmental factors, which could exert important roles under gene-environment interactions. Among the strengths we can highlight its multicenter design, enrolling samples from distinct regions of Brazil, which brings a better representation of the Brazilian population, the application of Tag-SNP strategy, covering BMP4 hot-target SNPs, and the use of robust statistical approaches with control for confounding effects including sex and ancestry proportions and application of correction for multiple comparison tests such as Bonferroni threshold and 1000 permutation, which reduce spurious results. The present study offers many potential directions for future research. For example, expanding the analysis by including other SNPs within BMP4 and variants in other genes of the pathway could provide a more comprehensive understanding of the genetic underpinnings of craniofacial development. Additionally, exploring genetic interactions in other signaling pathways involved in craniofacial development could help elucidate the complex mechanisms at play. Moreover, investigating the functional significance of the genetic variants identified in the study and examining gene-environment interactions could provide further insight into the etiology of NSOC. Lastly, exploring similar patterns of genetic variation in a larger sample size, particularly from NSCPO, could confirm the findings of the present study and contribute to the development of more of more effective strategies of prevention and diagnosis. More comprehensive understanding of the genetic and environmental factors and molecular mechanisms that contribute to NSOC could be gained by integrating the recent advances of bioinformatics with the multi-omics datasets and functional studies.
Conclusion
Our results demonstrate the association of rs2761887 in the susceptibility of NSCPO, and the increased risk for NSCL ± P and NSCPO in the presence of rs2761887 variant in Brazilians with high African ancestry. Interactions between BMP4 and FGFR1, GREM1, NOG, VAX1 and the 4p16.2 locus may also be related to the pathogenesis of this common craniofacial malformation.
Supplemental Material
sj-docx-1-cpc-10.1177_10556656231180086 - Supplemental material for Ethnic Differences in the Brazilian Population Influence the Impact of BMP4 Genetic Variants on Susceptibility of Nonsyndromic Orofacial Clefts
Supplemental material, sj-docx-1-cpc-10.1177_10556656231180086 for Ethnic Differences in the Brazilian Population Influence the Impact of BMP4 Genetic Variants on Susceptibility of Nonsyndromic Orofacial Clefts by Lilianny Querino Rocha de Oliveira and Hellen Carolliny de Souza Nicolau, Daniella Reis Barbosa Martelli, Hercílio Martelli-Júnior, Rafaela Scariot, Ana Lúcia Carrinho Ayroza Rangel, Silvia Regina de Almeida Reis, Ricardo D. Coletta, Renato Assis Machado in The Cleft Palate Craniofacial Journal
Supplemental Material
sj-docx-2-cpc-10.1177_10556656231180086 - Supplemental material for Ethnic Differences in the Brazilian Population Influence the Impact of BMP4 Genetic Variants on Susceptibility of Nonsyndromic Orofacial Clefts
Supplemental material, sj-docx-2-cpc-10.1177_10556656231180086 for Ethnic Differences in the Brazilian Population Influence the Impact of BMP4 Genetic Variants on Susceptibility of Nonsyndromic Orofacial Clefts by Lilianny Querino Rocha de Oliveira and Hellen Carolliny de Souza Nicolau, Daniella Reis Barbosa Martelli, Hercílio Martelli-Júnior, Rafaela Scariot, Ana Lúcia Carrinho Ayroza Rangel, Silvia Regina de Almeida Reis, Ricardo D. Coletta, Renato Assis Machado in The Cleft Palate Craniofacial Journal
Supplemental Material
sj-docx-3-cpc-10.1177_10556656231180086 - Supplemental material for Ethnic Differences in the Brazilian Population Influence the Impact of BMP4 Genetic Variants on Susceptibility of Nonsyndromic Orofacial Clefts
Supplemental material, sj-docx-3-cpc-10.1177_10556656231180086 for Ethnic Differences in the Brazilian Population Influence the Impact of BMP4 Genetic Variants on Susceptibility of Nonsyndromic Orofacial Clefts by Lilianny Querino Rocha de Oliveira and Hellen Carolliny de Souza Nicolau, Daniella Reis Barbosa Martelli, Hercílio Martelli-Júnior, Rafaela Scariot, Ana Lúcia Carrinho Ayroza Rangel, Silvia Regina de Almeida Reis, Ricardo D. Coletta, Renato Assis Machado in The Cleft Palate Craniofacial Journal
Footnotes
Author Contributions
Conceptualization, H.M-Jr., R.D.C., R.A.M.; data curation, L.Q.R.O., H.C.S.N., R.A.M; formal analysis, L.Q.R.O., H.C.S.N., R.A.M; funding acquisition, D.R.B.M., H.M-Jr., R.D.C.; investigation, L.Q.R.O., H.C.S.N., R.A.M; methodology, R.A.M., R.D.C.; resources, A.L.C.A.R., S.R.A.R., R.S.; writing-original draft, L.Q.R.O., R.D.C., R.A.M.; writing-review and editing, L.Q.R.O, H.C.S.N., A.L.C.A.R., S.R.A.R., R.S., D.R.B.M., H.M-Jr., R.D.C., R.A.M. All authors have read and agreed to the published version of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was partially supported by grants from Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) for D. R. B. Martelli. H. Martelli-Junior and R. D. Coletta are research fellows of The National Council for Scientific and Technological Development (CNPq), and L. Q. R. de Oliveira is a receipting of a fellowship from The Coordination of Superior Level Staff Improvement (CAPES)
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.