
Abstract
Objective
To investigate the association between Wnt signaling pathway genes and non-syndromic orofacial cleft (NSOC) in the Han Chinese population.
Design
Based on a previously published genome-wide association study (GWAS), we performed a discovery phase analysis on 635 QC-passed SNPs (out of 7054 initially extracted from 59 Wnt pathway genes). Significant loci were then validated in an independent replication cohort.
Setting
A specialized craniofacial surgery center within a tertiary care institution.
Patients/Participants
In the discovery phase, we extracted the genotype data of 2512 NSOC cases and 2255 controls from two previous published GWASs. The independent replication cohort included 2724 patients with NSOC and 1263 healthy controls, all of Han Chinese descent.
Interventions
No clinical interventions were applied; the study involved genetic data analysis only.
Main Outcome Measures
SNPs associated with NSOC and its subtypes were identified through allelic and genotypic association analyses, with odds ratios (ORs), 95% confidence intervals (CIs), and P-values calculated.
Results
In the independent replication cohort, rs4821611 in RAC2 was significantly associated with NSOC (P = 5.8 × 10−7, OR = 0.77, 95% CI: 0.70-0.85), NSCL/P (P = 4.4 × 10−11, OR = 0.68, 95%CI: 0.61-0.77), and NSCLO (P = 3.27 × 10−15, OR = 0.6, 95% CI: 0.53-0.68). Genotypic analysis confirmed these associations. rs757190 in WNT3 was also associated with NSCLP (P = 0.00056, OR = 0.78, 95% CI: 0.67-0.90).
Conclusions
rs4821611 in RAC2 and rs757190 in WNT3 are associated with NSOC and its subtypes in the Han Chinese population, supporting a role for Wnt signaling in cleft pathogenesis.
Keywords
Orofacial clefts have an approximate global prevalence of 5.34 per 10,000 population, 1 with notable racial and geographic variations. 2 In China, cleft lip with or without palate ranks among the most common congenital birth defects, with an overall prevalence of 7.4 per 10,000 births. 3 Patients with clefts of the lip and palate face challenges in appearance, speech, feeding, and social integration, imposing significant burdens on individuals, families, and society. 4
Orofacial cleft is categorized into non-syndromic orofacial cleft (NSOC) and syndromic orofacial cleft, with NSOC accounting for approximately 70% of cases. 5 NSOC can be further subdivided based on different phenotypes of cleft lip and cleft palate into NSCL/P (non-syndromic cleft lip with or without cleft palate), NSCLP (non-syndromic cleft lip and palate), NSCPO (non-syndromic cleft palate), and NSCLO (non-syndromic cleft lip) (Figure 1). The development of NSOC involves complex interactions between genetic and environmental factors, among them, genetic factors play a dominant role,6,7 and its underlying mechanisms remain incompletely understood. Elucidating the etiology of NSOC is essential for prevention strategies.
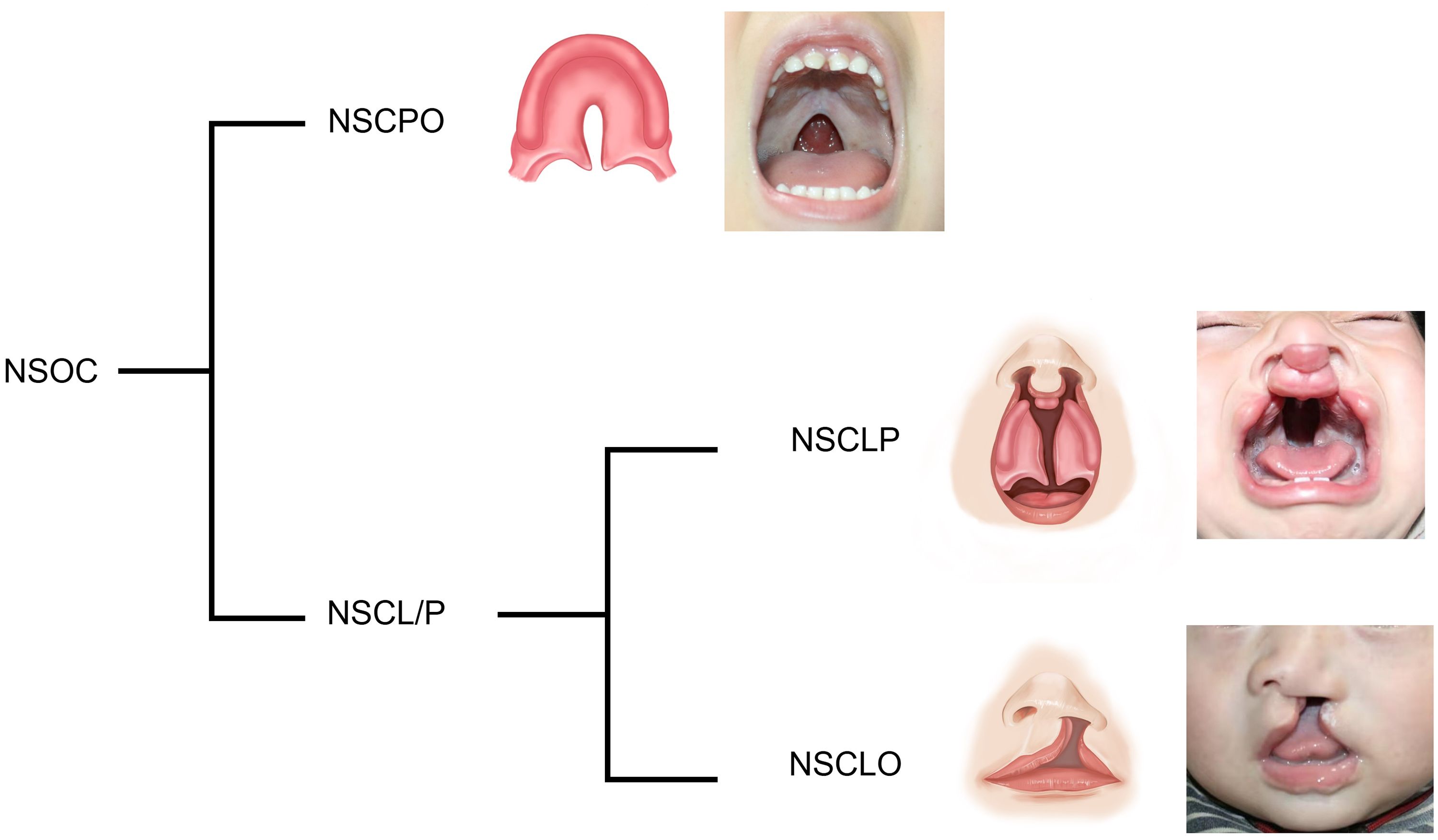
Classification diagram of non-syndromic orofacial cleft (NSOC).
The Wnt signaling pathway, a conserved cellular communication mechanism, plays a crucial role in embryonic development, including axis formation, cell differentiation, proliferation, and migration. It is divided into the canonical (β-catenin-dependent) and non-canonical pathways, the latter comprising the Wnt/planar cell polarity (PCP) and Wnt/Ca2+ pathways.8–10 Studies have demonstrated that the functional state of the Wnt pathway significantly impacts orofacial development, as evidenced in multiple animal models. Several genome-wide association studies (GWASs) and population-based association studies have also linked polymorphisms in Wnt pathway genes to NSOC.11–14 However, prior research has primarily focused on the canonical pathway, with limited exploration of the non-canonical pathways. Moreover, GWAS findings are subject to population heterogeneity. Investigating the association between Wnt pathway-related genes and NSOC in the Chinese Han population is critical for understanding the etiology of NSOC in this population.
Materials and Methods
Samples
This study used two cohorts: (1) a discovery cohort derived from our previously published GWAS of NSOC,14,15 including 2512 NSOC cases and 2255 controls; and (2) an independent replication cohort recruited at Department of Cleft Lip and Palate Surgery, West China Hospital of Stomatology, Sichuan University. The replication cohort involved two groups defined by distinct inclusion criteria. The case group comprised 2724 individuals of Han Chinese descent diagnosed with NSOC by experienced cleft specialists at Department of Cleft Lip and Palate Surgery, West China Hospital of Stomatology, Sichuan University. All patients were confirmed to have no other congenital anomalies or systemic diseases. The control group, also of Han Chinese descent, included 1263 individuals with no history of congenital anomalies, systemic diseases, or familial occurrence of orofacial clefts. These participants were recruited from West China Hospital of Stomatology, Sichuan University. The study protocol was approved by the Ethics Committee of West China Hospital of Stomatology, Sichuan University, WCHSIRB-D-2022-237-R1. Written informed consent was obtained from all participants prior to enrollment. For participants under 18 years of age, consent was provided by a parent or legal guardian. Detailed information on the participants and group assignments, explicitly distinguishing between the discovery and replication cohorts and including the exact number of cases and controls for both phases, is clearly presented in Table 1.
Samples Information of the Discovery Phase and Replication Phase.
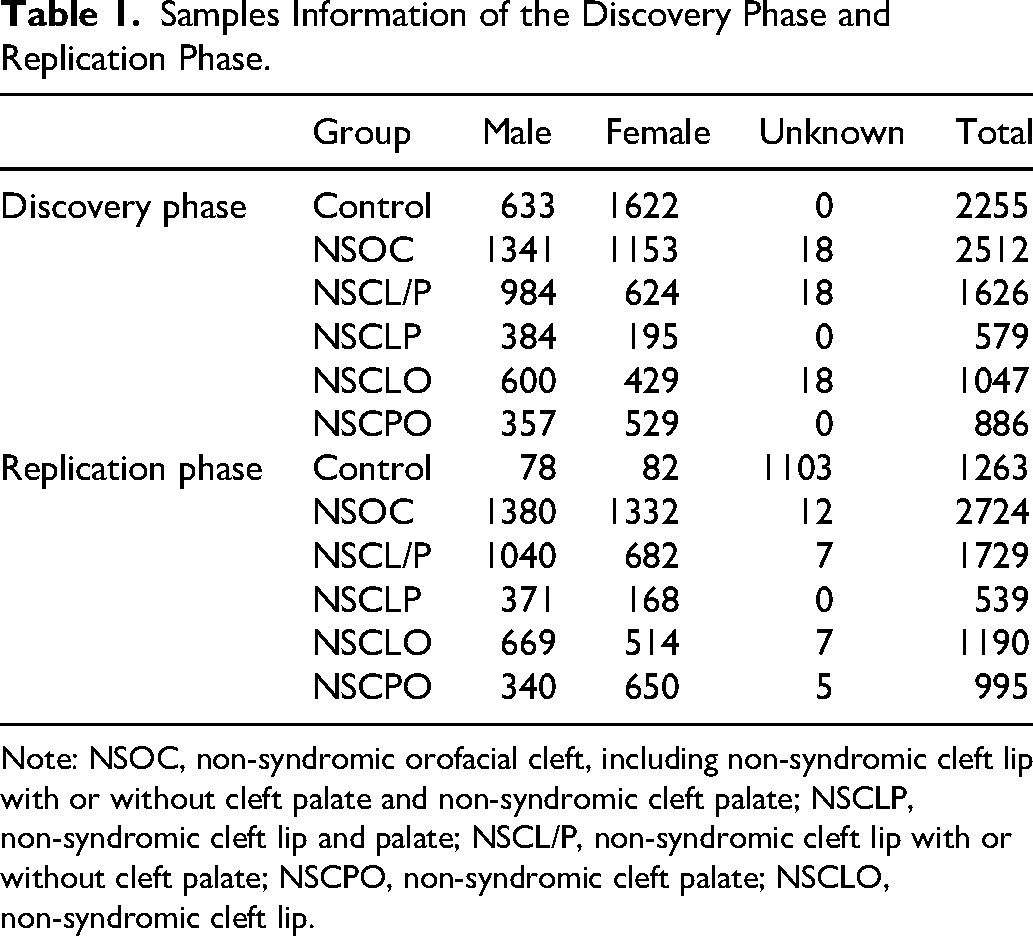
Note: NSOC, non-syndromic orofacial cleft, including non-syndromic cleft lip with or without cleft palate and non-syndromic cleft palate; NSCLP, non-syndromic cleft lip and palate; NSCL/P, non-syndromic cleft lip with or without cleft palate; NSCPO, non-syndromic cleft palate; NSCLO, non-syndromic cleft lip.
SNP Screening and Association Analysis in the Discovery Phase
Based on previously published GWAS studies on NSOC by our research team,15,16 genotyping data for all 7054 SNP within 59 Wnt signaling pathway-related genes were extracted for the first-round association analysis. We performed for the SNPs: (1) for sample-level QC: call rate ≥ 98%; (2) for SNP-level QC: Hardy-Weinberg equilibrium (HWE) in controls (P > 10−6); (3) call rate ≥ 98%; (4) minor allele frequency (MAF) ≥ 0.01. A total of 635 SNPs were passed quality controls and enrolled in the association analysis. SNPs with P < 2.62 × 10−5 (0.05/635SNPs/3phenotypes) are considered to meet the Bonferroni-corrected significance level, while SNPs with 2.62 × 10−5 ≤P < 7.9 × 10−5 (0.05/635SNPs) are treated as suggestive. SNPs with P-values meeting the Bonferroni-corrected suggestive level were enrolled in the following replication phase, which was conducted using both case and control samples. All association analyses were performed using PLINK v1.9. Specifically, logistic regression models were utilized for the comparison of allelic and genotypic frequencies between cases and controls, allowing us to calculate the corresponding odds ratios (ORs) and 95% confidence intervals (CIs). Allelic association testing was conducted to evaluate the baseline susceptibility conferred by individual minor alleles. Genotypic association testing was then performed to determine the likely mode of inheritance and to understand how specific allele combinations contribute to the susceptibility of distinct cleft subphenotypes.
DNA Extraction and Genotyping in Replication Phase
Details regarding the collection, processing of biological samples, and DNA extraction are provided in the reference. 15 Genotyping experiments were outsourced to Shanghai Tianhao Biotechnology Co., Ltd., using SNPcan™ technology (http://www.geneskies.com/).
Data Quality Control and Statistical Analysis of Replication
The genotype frequency distribution in the control group was tested for HWE to verify the genotyping quality of the study sample. The MAF was calculated for each group, and SNPs with an MAF below 1% were excluded. In the replication phase, we used PLINK v1.9 to perform allelic and genotypic association analyses using logistic regression models. Allelic association testing was conducted to evaluate the baseline effect conferred by individual minor alleles. Genotypic association testing was then performed to determine the likely mode of inheritance and to understand how specific allele combinations contribute to the susceptibility of the distinct NSOC subphenotypes. The Bonferroni correction significance threshold was set at 0.0024 (P-value = 0.05 / (7 SNPs × 3 phenotypes)).
Functional Prediction and Annotation
The potential effects of candidate SNPs on transcriptional regulation were explored using the HaploReg v4.2 database (https://pubs.broadinstitute.org/mammals/haploreg/haploreg.php). Noncoding SNP-related regulatory elements and chromatin states were annotated using the 3DSNP v2.0 tool (https://omic.tech/3dsnpv2/).
Results
Preliminary Screening of Candidate SNPs in Discovery Phase
From the first-round association analysis of only 635 SNPs that passed QC (out of the 7054 initially genotyped SNPs), candidate SNP meeting the following criteria were selected for the replication phase: (1) Statistical significance of P < 7.9 × 10−5; (2) No linkage disequilibrium among candidate SNP. After screening, rs307369, rs2230773, rs186940875, rs17302049, rs757190, rs118086259, and rs4821611 were identified as candidate SNPs for the replication phase. In the discovery phase, only allelic association analysis was performed to identify preliminary signals. The preliminary screening results are presented in Table 2 and Figure 2a.
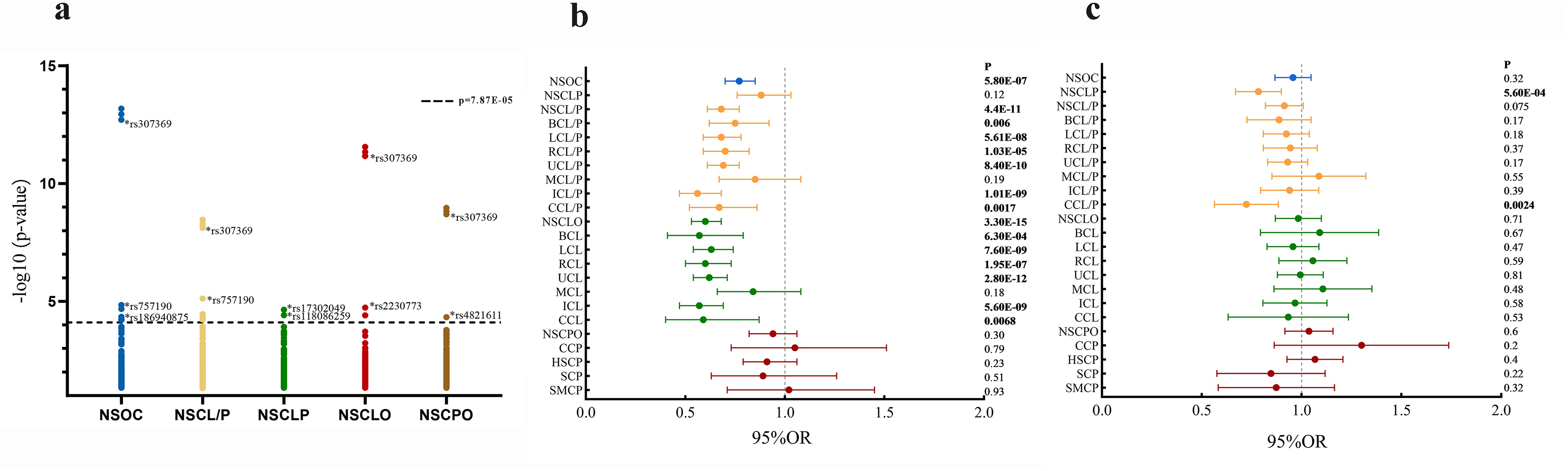
Preliminary screening and allelic association analysis of candidate SNPs in Wnt signaling pathway-related genes. (a) Preliminary screening results of candidate SNPs in Wnt pathway-related genes. (b) Allelic association analysis results of rs4821611 (P < 0.0071). (c) Allelic association analysis results of rs757190 (P < 0.0071).
Allelic Association Analysis Results of Candidate SNPs in the Discovery Phase.
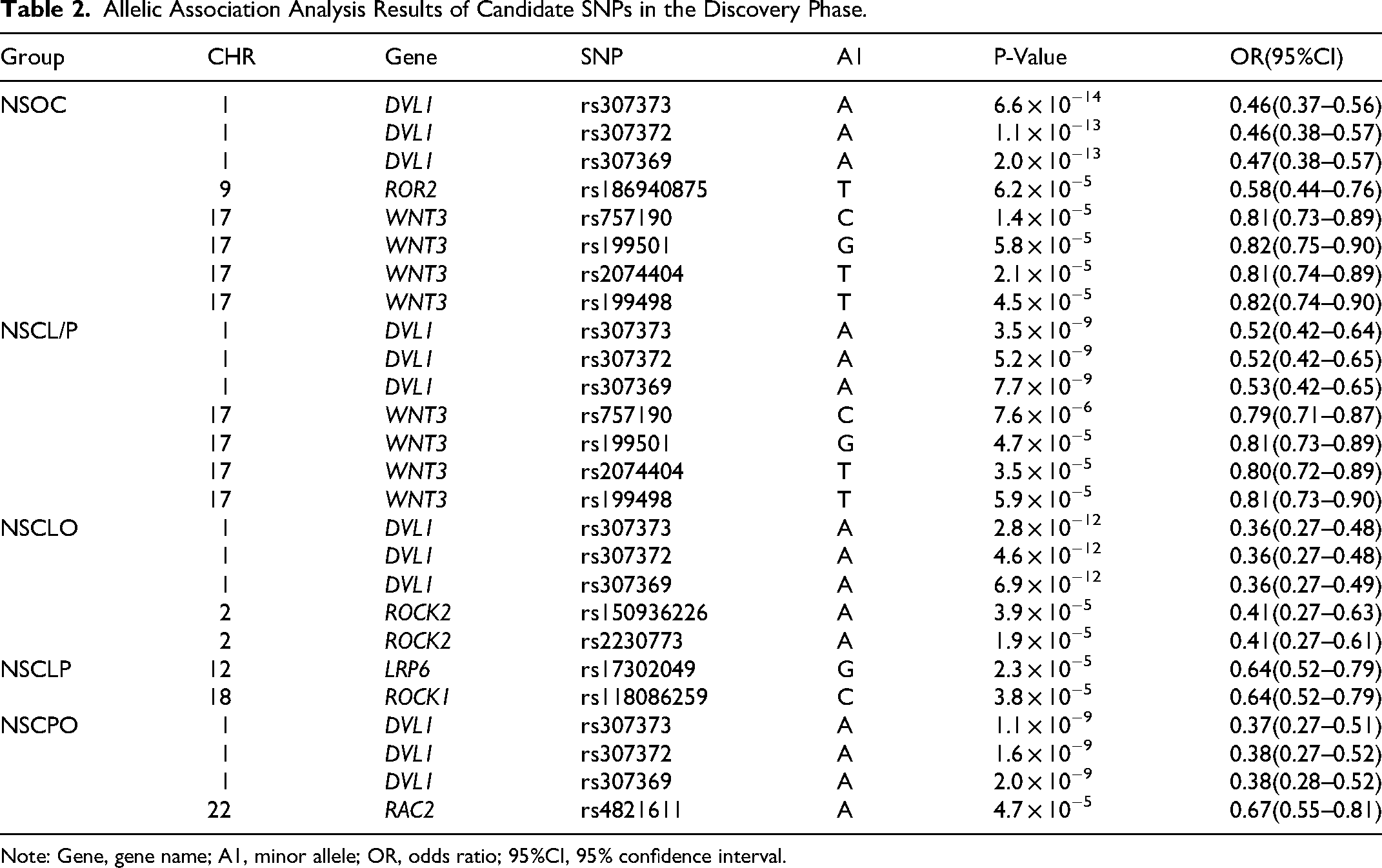
Note: Gene, gene name; A1, minor allele; OR, odds ratio; 95%CI, 95% confidence interval.
HWE Test and MAF in Replication Phase
The genotype frequencies of the seven SNPs selected for validation were tested for HWE in the control group. The results showed that all seven SNPs conformed to HWE (P > 0.05), indicating no significant difference between the observed and expected genotype frequencies, thus confirming the genotype quality of the samples. The MAFs of the seven SNPs in each subtype group met the study criteria (MAF ≥ 0.01). The results of the HWE test and MAF analysis are presented in Table S1.
Results of Allelic Association Analysis in Replication Phase
Allelic association analysis revealed that allele A at rs4821611 in RAC2 gene of the non-canonical Wnt signaling pathway (PCP pathway) was associated with NSOC (P = 5.8 × 10−7, OR = 0.77, 95% CI: 0.70-0.85) and NSCL/P (P = 4.4 × 10−11,OR = 0.68,95%CI:0.61-0.77) and NSCLO (P = 3.3 × 10−15,OR = 0.60,95%CI:0.53-0.68) after multiple testing corrections. Within the canonical Wnt signaling pathway, only the C allele at rs757190 in WNT3 gene showed association with NSCLP (P = 5.6 × 10−4, OR = 0.78,95%CI:0.67-0.90) after multiple testing correction. SNPs exhibiting opposite effect directions across Tables 2 and 3 failed to pass the corrected significance threshold and are therefore strictly classified as non-replicated signals. Detailed results are provided in Table 3.
Allelic Association Analysis Results of NSOC and its Subtypes in Replication.
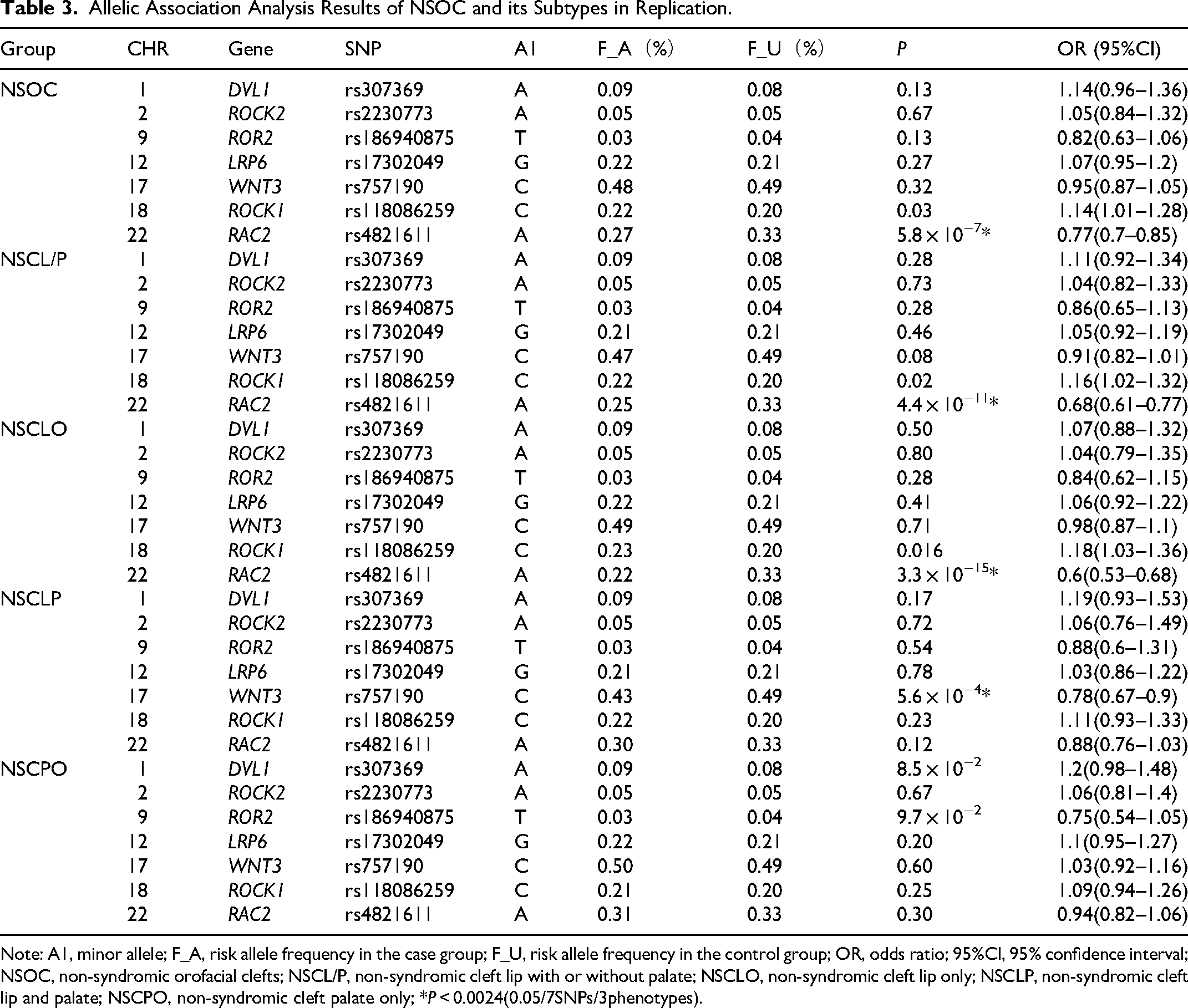
Note: A1, minor allele; F_A, risk allele frequency in the case group; F_U, risk allele frequency in the control group; OR, odds ratio; 95%CI, 95% confidence interval; NSOC, non-syndromic orofacial clefts; NSCL/P, non-syndromic cleft lip with or without palate; NSCLO, non-syndromic cleft lip only; NSCLP, non-syndromic cleft lip and palate; NSCPO, non-syndromic cleft palate only; *P < 0.0024(0.05/7SNPs/3phenotypes).
No SNP was associated with NSCPO. To further investigate, NSCPO was subdivided into four subtypes: complete cleft palate, hard and soft cleft palate (HSCP), soft cleft palate (SCP), and submucous cleft palate (Fig. S1; sample information in Table S2). Further analysis showed that rs307369 was significantly associated with HSCP (P = 0.043, OR = 1.27,95%CI:1.01-1.61), and rs2230773 was significantly associated with SCP (P = 0.040, OR = 1.85,95%CI:1.02-3.37). However, these associations did not remain statistically significant after correction for multiple testing. Detailed results are provided in Table S3.
Given the associations of rs4821611, rs118086259, and rs757190 with lip-related subgroups, further analyses were conducted to assess these loci's associations with lip-related subtypes. Based on the location of clefts, lip-related phenotypes were categorized into: bilateral cleft lip with or without palate (BCL/P), unilateral cleft lip with or without palate (UCL/P), right cleft lip with or without palate (RCL/P), left cleft lip with or without palate (LCL/P), bilateral cleft lip (BCL), unilateral cleft lip (UCL), right cleft lip (RCL), and left cleft lip (LCL) (Fig. S2; sample information in Table S2). Subtype analyses revealed that allele A at rs4821611 was associated with all lip-related subtypes after multiple testing corrections. C allele at rs118086259 was marginally associated with UCL/P (P = 0.029, OR = 1.16,95%CI:1.02-1.33), RCL/P (P = 0.032, OR = 1.21,95%CI:1.02-1.44), and UCL (P = 0.024, OR = 1.18,95%CI:1.02-1.36), but these associations were not statistically significant after multiple testing corrections. rs757190 showed no association with lip-related subtypes.
Regarding cleft severity, lip-related phenotypes were categorized into: complete cleft lip with or without palate (CCL/P), incomplete cleft lip with or without palate (ICL/P), microform cleft lip with or without palate (MCL/P), complete cleft lip (CCL), incomplete cleft lip (ICL), and microform cleft lip (MCL) (Fig. S3; sample information in Table S2). Subtype analyses showed that the A allele at rs4821611 was associated with CCL/P (P = 0.0017,OR = 0.67,95%CI:0.52-0.86), ICL/P (P = 1.0 × 10−9,OR = 0.56,95%CI:0.47-0.68),CCL (P = 6.8 × 10−3,OR = 0.59,95%CI:0.40-0.87), and ICL (P = 5.64 × 10−9,OR = 0.57,95%CI:0.47-0.69) after multiple testing corrections. C allele at rs118086259 was marginally associated with ICL/P (P = 0.032, OR = 1.23,95%CI:1.02-1.49). rs757190 was associated with CCL/P (P = 0.0024, OR = 0.71,95%CI:0.57-0.89) after multiple testing corrections. Detailed results are provided in Table S4. The allelic association analysis results of rs4821611 and rs757190 are shown in Figure 2b and c.
Genotypic Association Analysis Results in Replication Phase
Genotypic association analysis revealed statistically significant associations between the genotypic distributions of rs4821611 and the susceptibility of NSOC (P = 9.7 × 10−1⁰), NSCL/P(P = 3.5 × 10−15), NSCLO(P = 1.1 × 10−2⁰), LCL(P = 6.2 × 10−14), LCL/P(P = 5.8 × 10−13), RCL(P = 6.4 × 10−1⁰), RCL/P(P = 2.3 × 10−7), UCL(P = 1.4 × 10−18), UCL/P(P = 5.3 × 10−15), CCL(P = 0.0041), CCL/P(P = 0.00088), ICL(P = 2.3 × 10−1⁰), and ICL/P(P = 3.7 × 10−11). For rs757190, significant associations were observed with NSCLP (P = 0.00029) and CCL/P (P = 0.0053). Further statistical analysis of the homozygous and heterozygous genotypes in the two sample groups showed that the rs4821611 G/G genotype (Figure 3a) was associated with increased disease susceptibility in NSCL/P (P = 0.0058, OR = 1.46, 95% CI: 1.12-1.90), NSCLO (P = 8.0 × 10−4, OR = 1.65,95%CI:1.23-2.20), ICL(P = 0.0023, OR = 1.99,95%CI:1.26-3.13), ICL/P(P = 0.0011, OR = 2.03,95%CI:1.30-3.15), and BCL (P = 0.0068, OR = 3.22,95%CI:1.21-8.53). The rs4821611 G/A genotype (Figure 3b) was associated with a reduced susceptibility of UCL (P = 0.0071, OR = 0.65, 95% CI: 0.48-0.88), consistent with the results of the allelic association analysis. Both the A/A and A/C genotypes of rs757190 (Figure 3c,d) were significantly associated with NSCLP and CCL/P; carrying the A allele at this locus increased the susceptibility of disease. Detailed genotypic association analysis results are presented in Table S5.
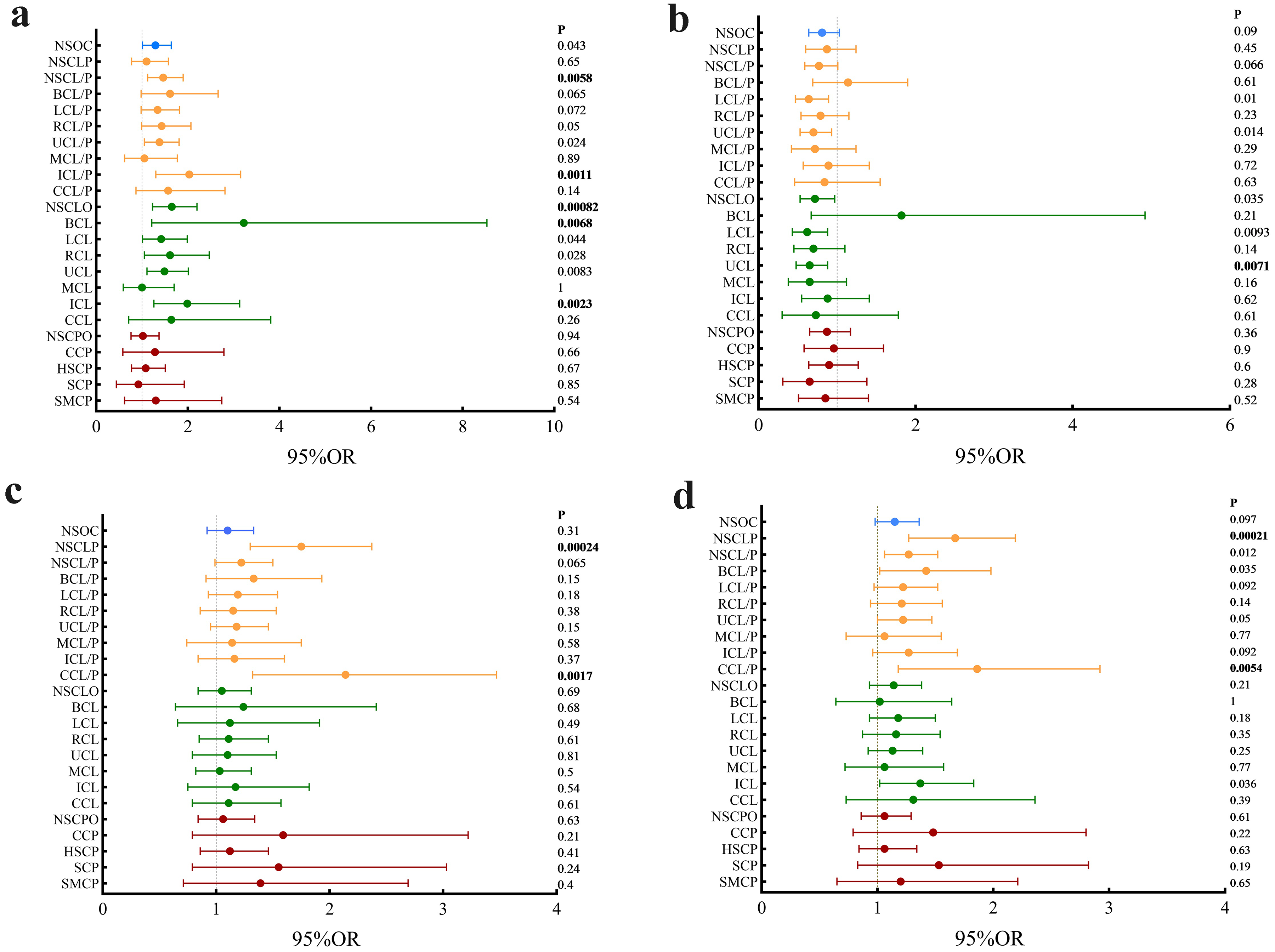
Genotype-based association analysis of rs4821611 and rs757190. (a) The homozygous genotypes association analysis results of rs4821611. (b) The heterozygous genotypes association analysis results of rs4821611. (c) The homozygous genotypes association analysis results of rs757190. (d) The heterozygous genotypes association analysis results of rs757190.
SNP Functional Annotation Results
We found that rs4821611 (RAC2) and rs757190 (WNT3) were located in intronic regions. The SNP located within the intron may influence gene expression by altering transcription factor binding motifs and regulatory chromatin states. 17 Functional annotation results from HaploReg v4.2 indicated that variations at rs4821611 and rs757190 might have altered the binding abilities of multiple transcription factors to DNA (see Table S6). According to 3D SNP v2.0 annotation (visualized in Fig. S4), rs4821611 scored the highest in the “motif” category. For rs757190, its interacting gene was WNT9B, which had been identified in GWAS studies as associated with non-syndromic cleft lip and palate.17–19 In mice, WNT9B was involved in regulating facial morphogenesis, 20 and knocking out WNT9B led to cleft lip and palate phenotypes. 21
Discussion
Craniofacial development involves the coordination of multiple signaling pathways, including the Bmp/Tgf-β signaling pathway, Fgf signaling pathway, Wnt pathway, and Shh signaling pathway. 22 Among them, Wnt signaling as a central, mechanistically substantiated contributor that plays distinct roles at multiple stages of orofacial cleft pathogenesis. Specifically, canonical Wnt/β-catenin signaling is required for facial process outgrowth and lip fusion.23–25 Conversely, non-canonical WNT5A signaling via ROR2 governs directional cell migration in the secondary palate mesenchyme and is strictly required for palate closure. 26 Furthermore, non-canonical pathways operating through JNK and PCP mechanisms heavily regulate facial prominence morphogenesis.27,28
In this study, 7054 SNPs from the canonical Wnt, Wnt/Ca2+, and PCP pathways were screened, of which 635 SNPs passed quality control and were subjected to statistical analysis. We found that A allele at rs4821611 in RAC2 gene (Wnt/PCP pathway) was associated with NSOC (P = 5.8 × 10−7, OR = 0.77,95%CI:0.70-0.85), NSCL/P (P = 4.4 × 10−11, OR = 0.68,95%CI:0.61-0.77), and NSCLO (P = 3.3 × 10−15, OR = 0.60,95%CI:0.53-0.68). Additionally, A allele at rs4821611 was associated with all lip-related subtypes (BCL, BCL/P, LCL, LCL/P, RCL, RCL/P, UCL, UCL/P). In analyses related to cleft lip severity, A allele at rs4821611 also exhibited associations with CCL/P (P = 0.0017,OR = 0.67,95%CI:0.52-0.86), ICL/P (P = 1.0 × 10−9,OR = 0.56,95%CI:0.47-0.68), CCL (P = 0.0068,OR = 0.59,95%CI:0.40-0.87), and ICL (P = 5.6 × 10−9,OR = 0.57,95%CI:0.47-0.69). Genotypic association analysis revealed that the G/G genotype at rs4821611 was associated with NSCL/P (P = 0.0058, OR = 1.46,95%CI:1.12-1.90), NSCLO (P = 8.0 × 10−4, OR = 1.65,95%CI:1.23-2.20), ICL (P = 0.0023, OR = 1.99,95%CI:1.26-3.13), ICL/P(P = 0.0011, OR = 2.03,95%CI:1.30-3.15), and BCL (P = 0.0068, OR = 3.22,95%CI:1.21-8.53). The G/A genotype was associated with UCL (P = 0.0071, OR = 0.65,95%CI:0.48-0.88), aligning with the allelic association findings. Furthermore, both the A/A and A/C genotypes of rs757190 were significantly associated with NSCLP and CCL/P.
rs4821611 in RAC2 of the Wnt/PCP pathway, which encodes a GTPase protein and a member of the Ras superfamily. 29 While direct studies on RAC2 in facial development are limited, its integral role in the Wnt/PCP pathway provides strong biological plausibility for its implication in craniofacial anomalies. The Wnt/PCP pathway is essential for the cellular orientation and directional migration requisite for proper palate morphogenesis. Disruptions in Wnt/PCP components, such as PRICKLE1, SAPCD2, CELSR3, and CCDC88C, severely impair cell migration and cause cleft palates in both animal models and human cohorts.30,31 WNT11, another crucial component, acts through JNK/PCP signaling to dynamically regulate cell behavior and spatial orientation during lip fusion.12,27 Given the function of RAC2 in cellular polarization and motility, 32 the genetic variants at rs4821611 could critically disrupt the highly coordinated cell migration processes required for facial prominence outgrowth. However, further studies are needed to explore its molecular mechanism in regulating cellular processes during lip and palate formation.
rs757190 is located in WNT3 gene, a component of the canonical Wnt signaling pathway. The WNT3 protein plays a critical role in regulating cell differentiation and germ layer formation during embryonic development. 33 Specifically, WNT3 functions within a highly conserved regulatory module (Pbx-Wnt9b-Wnt3-p63-Irf6) that controls apoptosis and epithelial seam degradation during lip fusion. 34 Its functional necessity is highlighted by the fact that WNT3 nonsense mutations cause Tetraamelia syndrome (involving severe clefts 35 ), and WNT3/WNT9B protein levels are significantly decreased in human cleft tissues. 36 Besides, the WNT3/WNT9B cluster at 17q21 is a well-established susceptibility locus for clefting. Large-scale multi-ethnic GWAS37–39 have consistently linked various WNT3 SNPs to NSOC susceptibility worldwide. Building upon this distributed genetic architecture, our study was the first to report the association of rs757190 (WNT3) with non-syndromic cleft lip and palate in a Chinese population. However, the functional significance of rs757190 is still unknown, and additional mechanistic studies are required to determine how variation at this locus may influence Wnt signaling and craniofacial morphogenesis.
Several limitations should be noted. First, this study was conducted solely in Han Chinese participants, which may limit the generalizability of the findings to other populations. As observed in the wider literature, Wnt pathway polymorphisms frequently exhibit population-specific linkage disequilibrium patterns; for example, WNT5A rs566926 and WNT3A rs752107 show markedly heterogeneous susceptibility effects across different ancestral groups.40,41 Second, environmental factors and potential gene-environment interactions were not assessed, which could influence the susceptibility of NSOC. This is a critical consideration given that environmental teratogens often converge on cleft phenotypes via Wnt pathway disruption. For example, retinoic acid completely inhibits canonical Wnt/β-catenin signaling in the early developing palate, 42 and maternal lead exposure has been shown to induce WNT3A hypermethylation, mediating a significant portion of NSCL/P susceptibility. 43 Furthermore, stochastic epigenetic events, such as the hypomethylation of a metastable epiallele near Wnt9b, have been shown to drive multifactorial clefting in mouse models. 44 Third, we lacked both demographic sex information and XY genotyping data for the samples used in this study. Consequently, we could not perform specific analyses by adding sex as a covariate or by stratifying the data by sex. Given the well-established sex biases in orofacial cleft subphenotypes, this inability to adjust for sex differences could potentially lead to spurious associations or mask critical association signals. Therefore, strictly sex-matched or sex-adjusted analyses are an absolute necessity in future genetic studies of cleft subphenotypes.
In summary, this study contributes to a better understanding of the role of Wnt signaling pathway-related genes in the development of non-syndromic cleft lip and palate. Future studies with fully harmonized, multi-center datasets are warranted to perform comprehensive meta-analyses and further validate these findings. Moreover, capturing detailed environmental exposures and sex-stratified data will be vital for unmasking the complex, distributed genetic architecture of the Wnt pathway. And functional studies are required to explore the pathogenic mechanisms, which could ultimately pave the way for pathway-targeted therapeutic strategies, as in vivo studies have already demonstrated the potential for pharmacological rescue of cleft phenotypes via localized Wnt agonism. 45
Conclusion
rs4821611 in RAC2 and rs757190 in WNT3, both involved in the Wnt signaling pathway, are associated with NSOC and its subtypes in the Han Chinese population. These findings provide further evidence supporting the involvement of Wnt signaling in the pathogenesis of NSOCs.
Supplemental Material
sj-docx-1-cpc-10.1177_10556656261464115 - Supplemental material for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese
Supplemental material, sj-docx-1-cpc-10.1177_10556656261464115 for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese by Sixuan Jia, Meilin Yao, Wenqi Ba, Sidi Zhang, Yue You, Shijun Duan, Bing Shi and Zhonglin Jia in The Cleft Palate Craniofacial Journal
Supplemental Material
sj-xlsx-2-cpc-10.1177_10556656261464115 - Supplemental material for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese
Supplemental material, sj-xlsx-2-cpc-10.1177_10556656261464115 for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese by Sixuan Jia, Meilin Yao, Wenqi Ba, Sidi Zhang, Yue You, Shijun Duan, Bing Shi and Zhonglin Jia in The Cleft Palate Craniofacial Journal
Supplemental Material
sj-xlsx-3-cpc-10.1177_10556656261464115 - Supplemental material for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese
Supplemental material, sj-xlsx-3-cpc-10.1177_10556656261464115 for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese by Sixuan Jia, Meilin Yao, Wenqi Ba, Sidi Zhang, Yue You, Shijun Duan, Bing Shi and Zhonglin Jia in The Cleft Palate Craniofacial Journal
Supplemental Material
sj-xlsx-4-cpc-10.1177_10556656261464115 - Supplemental material for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese
Supplemental material, sj-xlsx-4-cpc-10.1177_10556656261464115 for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese by Sixuan Jia, Meilin Yao, Wenqi Ba, Sidi Zhang, Yue You, Shijun Duan, Bing Shi and Zhonglin Jia in The Cleft Palate Craniofacial Journal
Supplemental Material
sj-xlsx-5-cpc-10.1177_10556656261464115 - Supplemental material for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese
Supplemental material, sj-xlsx-5-cpc-10.1177_10556656261464115 for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese by Sixuan Jia, Meilin Yao, Wenqi Ba, Sidi Zhang, Yue You, Shijun Duan, Bing Shi and Zhonglin Jia in The Cleft Palate Craniofacial Journal
Supplemental Material
sj-xlsx-6-cpc-10.1177_10556656261464115 - Supplemental material for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese
Supplemental material, sj-xlsx-6-cpc-10.1177_10556656261464115 for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese by Sixuan Jia, Meilin Yao, Wenqi Ba, Sidi Zhang, Yue You, Shijun Duan, Bing Shi and Zhonglin Jia in The Cleft Palate Craniofacial Journal
Supplemental Material
sj-xlsx-7-cpc-10.1177_10556656261464115 - Supplemental material for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese
Supplemental material, sj-xlsx-7-cpc-10.1177_10556656261464115 for Association Study of Wnt Pathway Genes with Non-syndromic Orofacial Cleft in Han Chinese by Sixuan Jia, Meilin Yao, Wenqi Ba, Sidi Zhang, Yue You, Shijun Duan, Bing Shi and Zhonglin Jia in The Cleft Palate Craniofacial Journal
Footnotes
Acknowledgements
The authors thank all the participants who donated samples in this study. This project was supported by the National Science Funds of China (No.81271118 and No. 81600849), the Research and Develop Program, West China Hospital of Stomatology Sichuan University (No. RD-03-202301).
Ethical Approval and Informed Consent Statements
This study was approved by the Ethics Committee of West China Hospital of Stomatology, Sichuan University (WCHSIRB-D-2022-237-R1). All participants were informed about the study and provided written consent prior to enrollment. For participants under 18 years old, consent was signed by their legal guardians.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Research and Develop Program, West China Hospital of Stomatology Sichuan University, Sichuan Province Science and Technology support program, National Science Funds of China, (grant numbers: No. RD-03-202301, 2024NSFSC0649, No. 81600849, No. 82170919).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
Data available upon request.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.