
Abstract
Interfacing robots with real biological systems is a potential approach to realizing truly adaptive machines, which is a long-standing engineering challenge. Although plants are widely spread and versatile, little attention has been given to creating cybernetic systems incorporating plants. Producing such systems requires two main steps: the acquisition and interpretation of biological signals, and issuing the appropriate stimulation signals for controlling the physiological response of the biological part. We investigate an automated physiological recovery of young avocado plants by realizing a closed interaction loop between the avocado plant and a water-control device. The study considers the two aforementioned steps by reading out postural cues (leaf inclination) and electrophysiological (biopotential) signals from the plant, and controlling the water resource adaptive to the drought condition of an avocado plant. Analysis of the two signals reveals time-frequency patterns of increased power and global synchronization in the narrow bands when water is available, and local synchronization in the broad bands for water shortage. The results indicate the feasibility of interface technologies between plants and machines, and provide preliminary support for achieving adaptive plant-based ‘machines’ based on plants’ large and robust physiological spectrum and machines’ control scheme diversity. We further discuss fundamental impediments hindering the use of living organisms like plants for artificial systems.
1 Introduction
Plants, in terms of biomass, are resilient living organisms. They have evolved and have adapted to various environments over time. Plants exploit patches that are rich in resources. They also exhibit light gradient growth (Karban, 2008; Trewavas, 2003) by changing their morphology (root/stem length, leaf growth pattern), or by altering the dynamics of their internal states, for example the number of chloroplasts on the leaves’ surface. Plants’ capacities to sense and compute these environmental influences, along with their ability to produce a goal-oriented response, are regarded as desired characteristics of machines (Mazzolai, Laschi, Dario, Mugnai, & Mancuso, 2010).
In a ‘cybernetics’ sense, as defined by Norbert Wiener (Wiener, 1948), it is beneficial to extend the science of communication and control to a novel class of living systems: the plants. The advancement of interfacing techniques between plants and machines can potentially produce novel sensing and actuation capabilities for machines and thus endow them with unprecedented adaptive abilities. For example, plant signals in plant–machine systems could potentially be useful as a robust sensor to harvest complex information from the environment, and to detect environmental resource distribution. Plants could also be used as alternative means of stretch-locomotion through controlled growth, with the appropriate stimulation input signals. Additionally the advancement of plant–machine interface techniques can support and sustain vegetative life under harsh conditions or provide efficient resource consumption schemes as well, which are critical for a sustainable future agriculture (Chen, 1990).
Limited research has been dedicated to the scope of plant–machine systems. An approach by Kuribayashi et al. has managed to provide a more accessible interaction between plants and humans by transforming plant signals into visual and acoustic feedback (Kuribayashi, Sakamoto, Morihara, & Tanaka, 2007; Kuribayashi, Sakamoto, & Tanaka, 2007). This technically enhanced plant anthropomorphism has enabled people to develop an emotional bond with their pet plants more easily. The observation of various parameters concerning the growth of crop plants (phytomonitoring) on a large scale is of great interest to the agricultural field (Shibayama et al., 2009). The automated visual detection system of leaf inclination was developed in Font, Körösi, and Farkas (2005) and Sakamoto et al. (2012), though the difficulty of monitoring a large number of plants remains. Ton et al. developed a commercial system based on phytomonitoring (Phytech Ltd.) for growers (Kopyt, Ton,Ner, Bachrach, & Zieslin, 2001). The benefits of this system were shown by Gurovich et al., who were able to increase the yield in an avocado orchard (Gurovich, Ton, & Vergara, 2006). They also evaluated electric signal measurements (biopotential) to gain a deeper insight into the processes that take place in every plant. A unique approach can be found in Correll et al. (2009), wherein Correll et al.constructed a garden with plants that were cultivated by robots with visual capabilities. The robots automatically irrigated the plants and harvested mature fruits. Studies of the artificial growth of plants using electrical gradients and the application of a synthetic plant hormone (auxin) are documented in the early work of Schrank (1951), and in the studies of Levetin and McMahon (1999), or Guardiola and Lazaro (1987) respectively.
One of the basic resources that a plant relies on is water. The fundamental functions of the plant, such as photosynthesis and the upkeep of its anatomical state, are heavily dependent on the availability of water. Cell turgor, a hydrostatic pressure inside the vacuole of a plant cell, keeps the plant erect and maintains its rigidity. Typically, stress due to ongoing drought results in visual signs of water shortage, such as declining leaves and wilting. Furthermore, water shortage affects the ability to transport nutrients inside the vascular tissue; the metabolic rate is immediately restricted. Self-recovery in the presence of water is an important property of plants. Note that the recovery does not necessarily promise that the plant regains its initial canopy configuration. Therefore, in soils with scarce water resources, a way of regulating the plant growth is by optimizing the consumption of the limited water resource. Insufficient water availability causes the plant to wilt, whereas excessive water availability leads to water shortage.
In the context of drought, the objective of this work is to investigate means for a bidirectional interface between a plant and a machine, where the machine can play a regulatory role in the water resource consumption. In particular, we examined visual and electrophysiological interfacing methods, in other words, assessing the leaf positions, or recording the electrical potential (biopotential) within the trunk of the plant, in order to evaluate and control drought signs in plants. These measurements can be used by the device counterpart to detect drought and respond by providing the plant with water when needed, which will ultimately maintain its healthy state. This study takes a necessary step toward establishing a closer relationship between plants and machines. We focus on the postural and biopotential changes in an avocado tree, and manage the communication between the plant and the water-control machine by conveying information from the plant to the machine and back again, in order to form a closed-loop interaction and maintain the fitness of the overall system. To our knowledge, combining the measurements of leaf inclination with those of biopotential in order to assess the hydration state and control a watering mechanism has not yet been approached.
The remainder of the paper is organized as follows: in Section 2, we outline the materials and methods; in Section 3, we present our experiments and results; we discuss some challenges in this work in Section 4 and we close with some concluding remarks in Section 5.
2 Material and methods
2.1 Growth and experimental environment
In this section, we present the experimental setup (plant stimuli, control algorithm, irrigation mechanism) and analysis methods. The avocado tree (Persea americana) has several advantages for indoor experiments: it has a rigid stem, exhibits a reasonable growth speed, and the seeds are commercially available almost everywhere; the tree has lance-shaped leaves with large surfaces, and it can grow at room temperature and under typical office lighting conditions (Encyclopaedia Britannica, 2011).
The avocados that we used for the experiments were grown in 1 l capacity pots for six months before the beginning of the experiments in a room as shown in Figure 1A. They reached an average height of approximately 40 cm. The experiments were carried out in a specially designed chamber where the climate conditions were kept at approximately 26°C and the lighting was maintained at a consistent level throughout the experiments (Figure 1B). Cameras took pictures of the entire setup every 10 min to verify the validity of the distance sensors’ values.
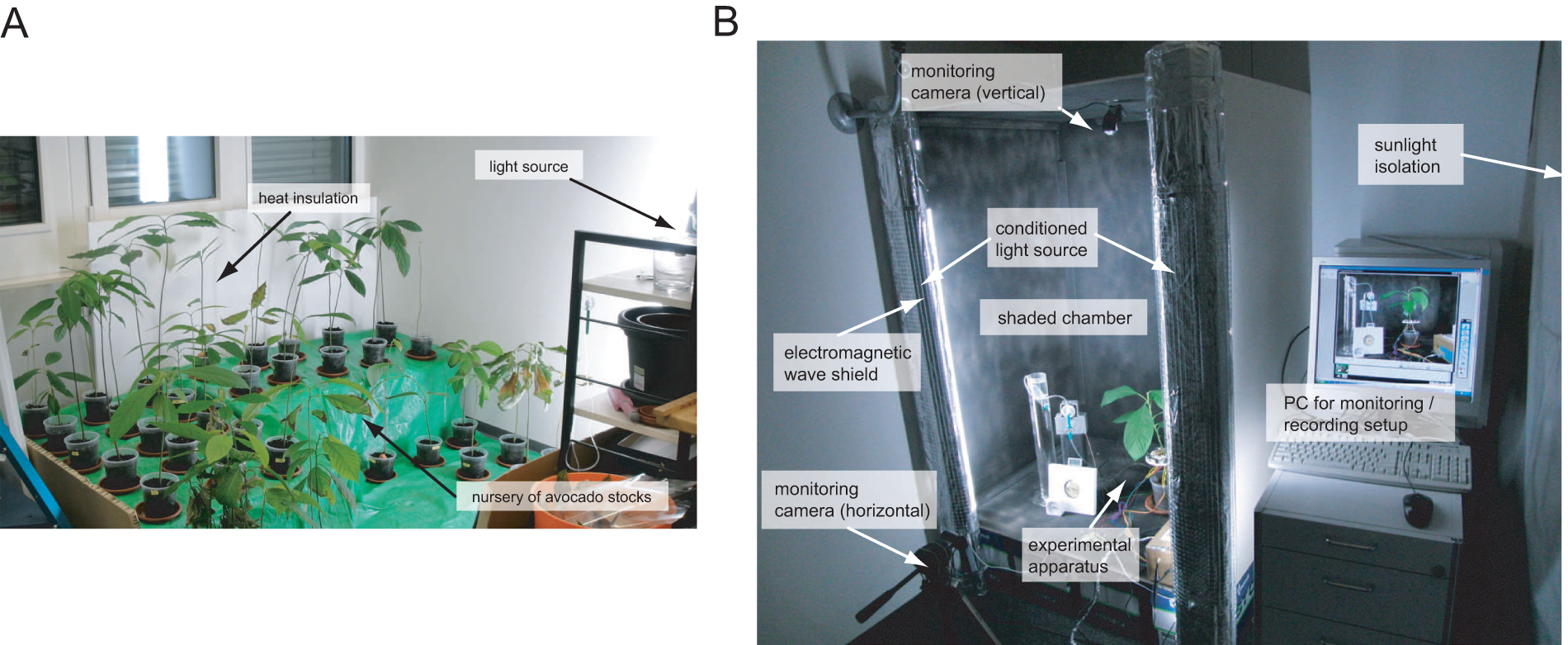
Setup spaces for avocado plants and avocado-based systems: (A) avocado plant nursery; (B) experimental room setup with controlled conditions and monitored processes.
2.2 Leaf inclination
Leaf inclination is an indicator of drought level in many plants (Font et al., 2005). Although leaf inclination may change due to various environmental factors, we reduced these influences by performing experiments with avocado plants in the controlled environment previously described. To ensure the portability of the device for future use, we built a disk by rapid prototyping to support six infrared (IR) distance sensors (Sharp GP2D120 83) arranged in a hexagonal shape (Figure 2A and B). The disk was attached to the stem of the plant 4 to 30 cm away from the canopy, which is also the range of the distance sensors. The distance sensors cover 360°, granting redundancy and allowing for an accurate reading of the postural state of the avocado. The measured values from the sensors were recorded and processed on a controller board (ATmega1280).
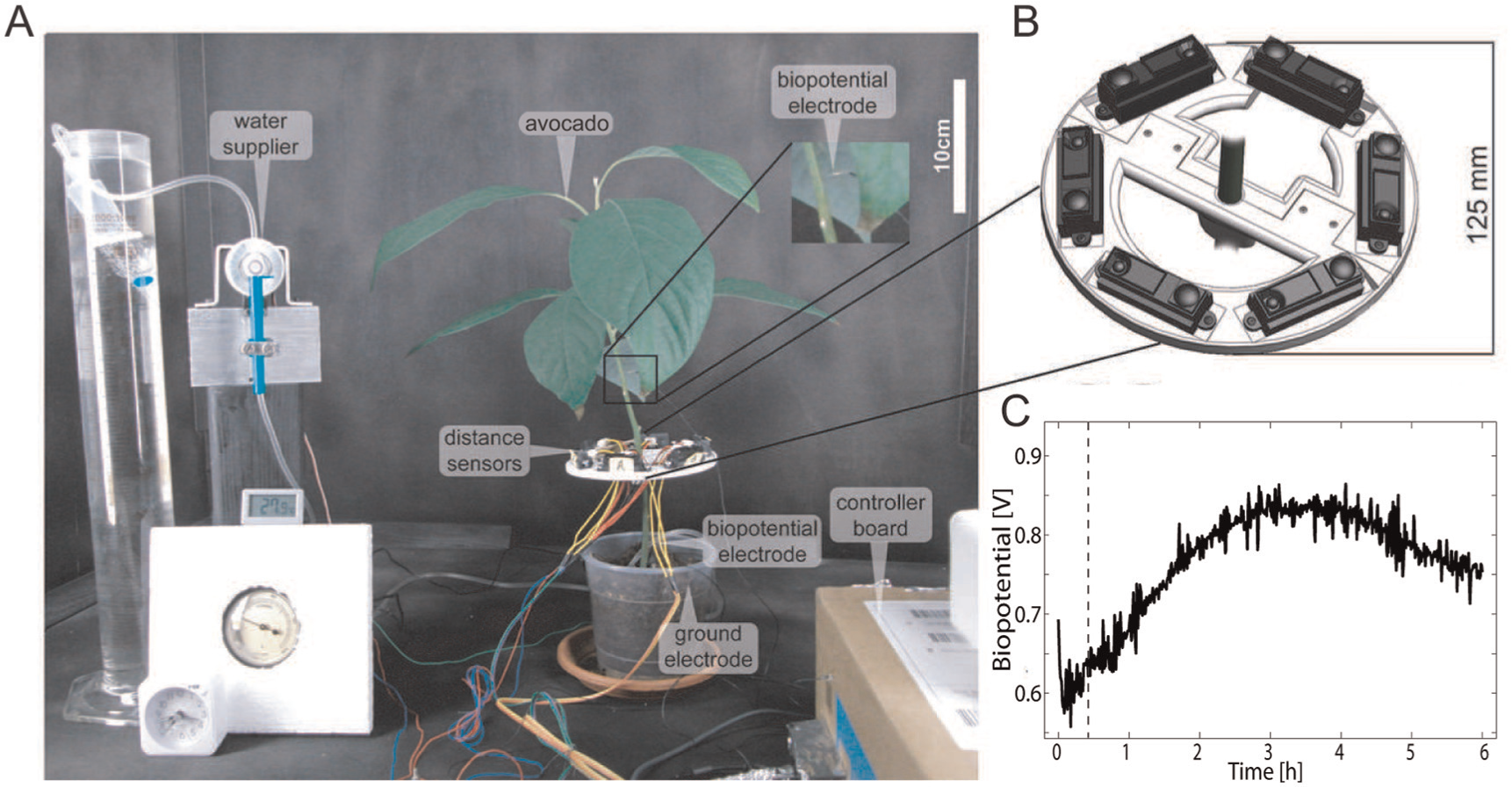
Experimental setup and procedure: (A) snapshot of the experimental setup; (B) the disk that supports the six distance sensors that compose the leaf-inclination detection system; (C) example of a biopotential signal from an avocado plant recorded at 1 kHz. The vertical dashed line represents an event of irrigation.
2.3 Biopotential
Different concentrations of ions between various parts of a plant produce an electric potential which is called biopotential. The difference in ionic concentrations emerge primarily due to the metabolic activities of the living cells (Fensom, 1963; Tattar & Blanchard, 1976). The metabolic activity, and by extension, the bioelectric parameters, are affected by the multiple dynamic equilibria that form the inter-cellular environment in a living organism. The bioelectric signals are also influenced by the organism’s interaction with a multitude of environmental influences, such as water stress, light yield, and the availability of nutrients (Fromm & Fei, 1998; Fromm & Lautner, 2007). In avocado plants, the physiological response to water and light was studied in Oyarce and Gurovich (2010) and their response to injuries such as pruning is found in Oyarce and Gurovich (2011). Since the availability of water is essential for every metabolic activity in plants we investigated the influence of water stress on the physiological state of the avocado plant.
We utilized biopotential needle electrodes to assess the electrical state of the avocado. We produced Ag/Ag-Cl electrodes by electrolyzing a silver wire (
The biopotential in plants is a mV-order weak electric potential (Parkinson & Banbury, 1966). Its measurement is a challenging task because the biopotential demonstrates a low signal-to-noise ratio, being impacted by environmental noise. To overcome this issue, a standard instrumentation amplifier consisting of three op-amps and a potentiometer to adjust the gain was used to increase this signal. Because biopotentials differ from plant to plant, the gains were adjusted to the range of 0 to 5 V. Furthermore, every 10 s, 1000 values from the sensors were sampled at 10 kHz and averaged to minimize high-frequency noise. A sample of a biopotential signal from an avocado plant, recorded with a sample rate of 1 kHz, is shown in Figure 2C, while the plant is irrigated with water after 0.5 hours from the beginning of signal recording. The controller board and the amplifier were placed in a cardboard box that was lined with aluminum foil and subsequently grounded.
2.4 Water supplier
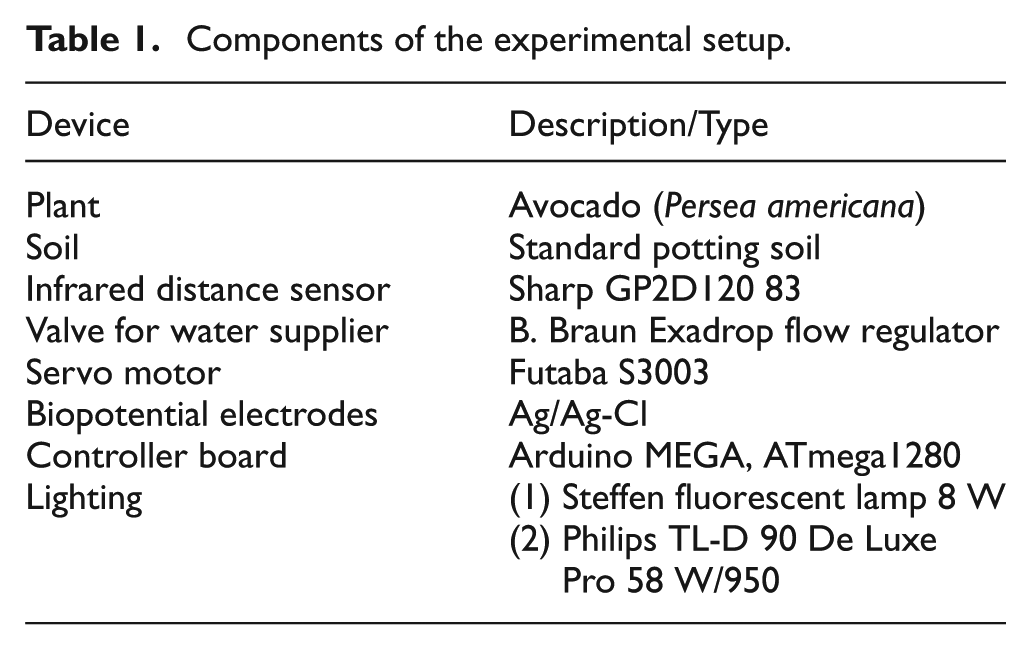
Constant and slowly dripping irrigation was provided by a flow regulator for gravity infusions (B. Braun Exadrop) as seen in Figure 2A. This allowed the automatic regulation of the flow to up to 250 ml per hour from the water tank. The hydrostatic pressure of the water tank was maintained for every experiment by manually refilling the tank with water periodically. The lower end of the tube was buried in the soil near the stem. The Exadrop valve was controlled by a servo motor (Futaba S3003), operated by the same controller board. The core system setup is listed in Table 1.
Components of the experimental setup.
2.5 Time-frequency analysis
The wavelet transformation of waveforms containing rhythmic activity is a strong method to see power and phase information of the activity at a specified time and frequency. The method is thus applied to various natural phenomena including life activity to uncover a state of the system in time-frequency domain. Here, we assumed that leaf inclination and biopotential of the avocado plants are also involved in biorhythmic activities, and the availability of water alters the state of this system. The wavelet analysis was first performed for the manual irrigation data to elucidate the potential state change of the plants as time-frequency trends that could not be seen from the original waveforms. Subsequently, the applicability of the trends to the automatic irrigation data was tested to validate the automated system (note that the automatic irrigation was not transient as the manual case). Hence, the continuous wavelet transformation with the complex Morlet wavelet
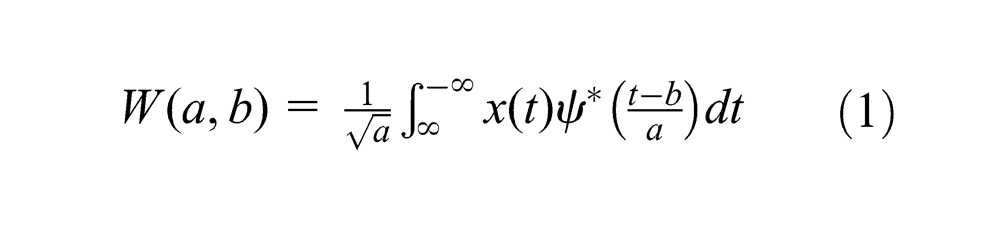
where
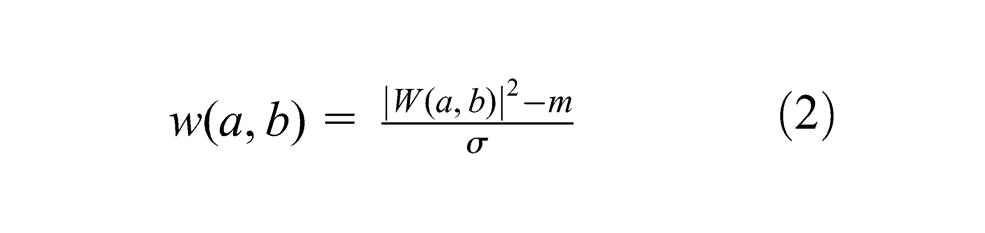
where
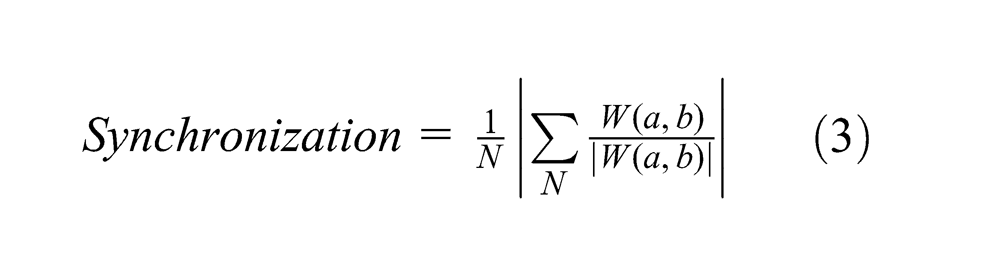
where
3 Experiments and results
In this section, we describe manual (Section 3.1) and semi-automatic (Sections 3.2.1 and 3.2.2) irrigation experiments as a study of control patterns of leaf inclination and biopotential signals. Based on previous experiments, an automatic closed-loop based on both leaf inclination and biopotential was achieved and is described in Section 3.2.3.
3.1 Manual irrigation
3.1.1 Effect on leaf inclination
In this experiment, we evaluated the change in leaf inclination caused by manual irrigation using the distance sensors. A baseline was recorded over the course of two hours, then the plants were manually irrigated with 50 ml of water, and the effect on the leaf inclination was measured for another four hours.
Figure 3 shows the average vertical movement of the canopy of eight avocados. Because the initial height of the canopy differed from plant to plant, the values were normalized to the same level at the time of the irrigation event, by multiplication with a fixed factor such that the average elevation at the time of irrigation was the same for every plant. Hence, the average elevation at the time of irrigation was 8 cm for every plant.
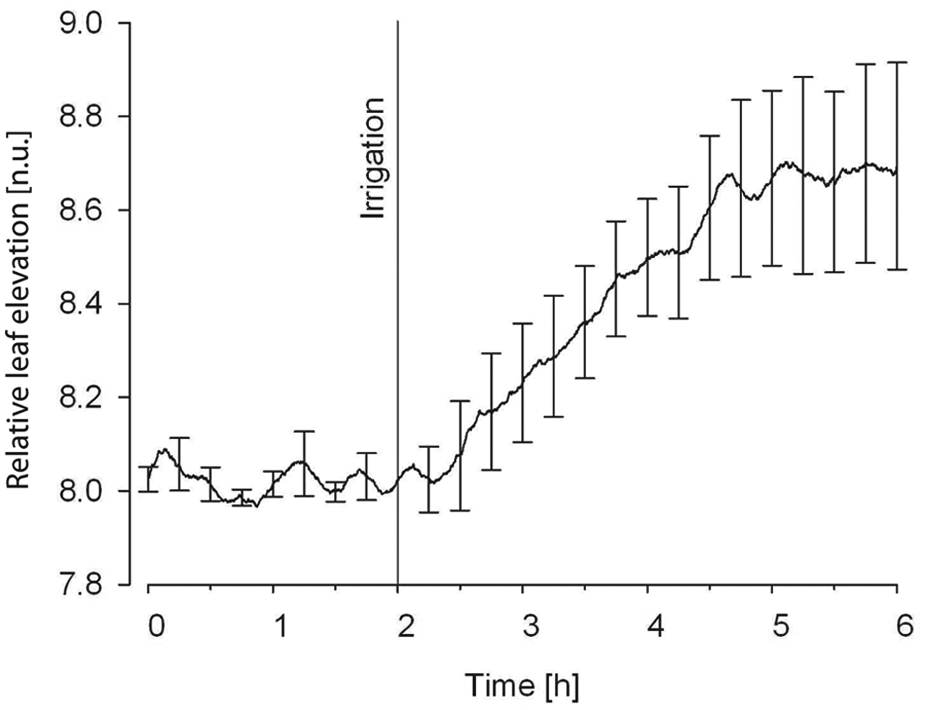
The average normalized vertical movement of the canopy of eight avocados after an irrigation event with 50 ml of water, measured with the distance sensor disk.
When the plant suffered from drought, the leaves withered slowly. As soon as the irrigation event occurred, a sharp rise of the canopy was seen in the figure within 30 min. This indicates that the leaves almost reached their regular height two hours after the irrigation event. Differences between the plants may occur due to phenotypic variations and particular stages of drought.
For some plants, the signal from a signal sensor occasionally registered sudden drops or jumps. This may be caused by a switch in the leaves’ position due to the kinematics of some interacting leaves. We counteracted this effect by selectively rejecting sensors that produced a variance higher than 0.7. This precaution usually dismissed one to three sensors out of an array of six. The redundancy of the sensors enabled the assessment of leaf changes with sufficient precision.
3.1.2 Effect on biopotential
This experiment evaluated the effect of manual water addition on the biopotential in an avocado plant. Plants that showed mild signs of drought were initially equipped with electrodes. After 30 min of recording a baseline, 50 ml of water were added manually.
Figure 4 presents the average normalized change in biopotential of 12 avocados. Because the absolute biopotential differed from plant to plant and because only the relative change was of interest, the measurements of all plants were normalized to the same level at the point of the irrigation event. Normalization was obtained by multiplication of the biopotential with an individual factor for every plant so that the biopotentials were identical at the time point of irrigation. This enabled us to compensate for the varying strengths of different biopotentials. In general, the biopotential tended to drop more or less slowly as soon as the recording began. We recorded data from eighteen different avocado plants, six of which had to be rejected because the biopotential signal was lost during the recording. If the biopotential did diverge from its initial trend, it did so by a sudden increase shortly after the irrigation event. The height of the biopotential in the 12 plants two hours after the irrigation event increased on average by
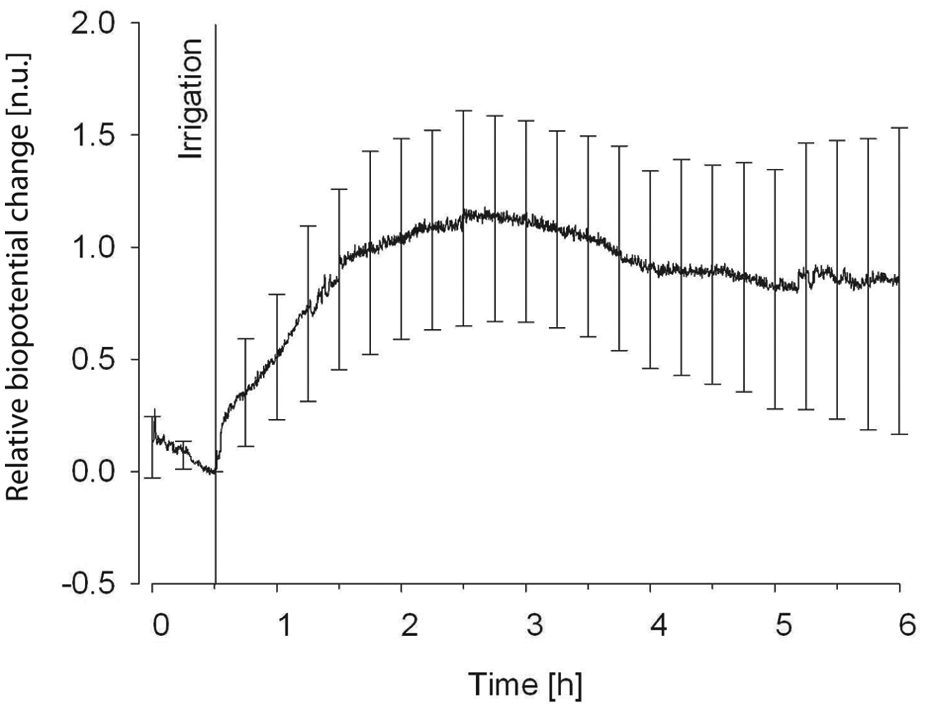
The average normalized change in the biopotential of 12 avocados after a manual irrigation event with 50 ml of water.
Although the pattern of the biopotential signal in avocados was consistent across our experiments, it is difficult to model or predict such a signal due to the variability of each plant feature. At irrigation, both between plants and within one plant, the amplitude, time offset from irrigation, and the slopes of the signal’s rise and fall can be very different. Generically, the signal between two irrigation events can roughly be modeled as a piecewise function such as:
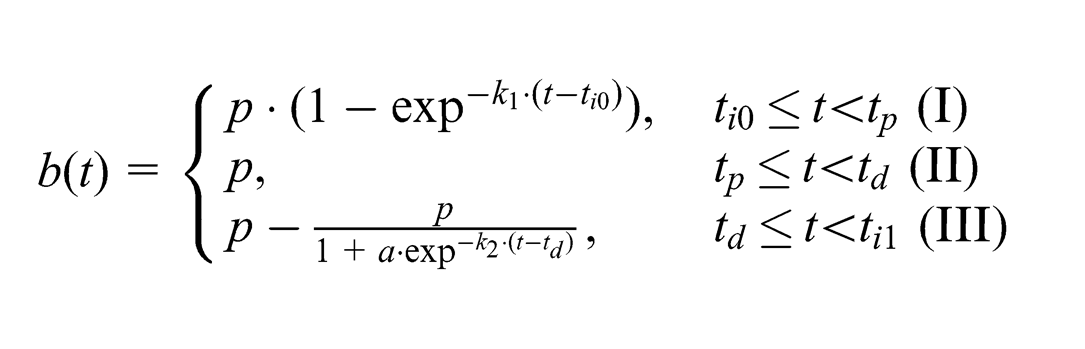
The function describes the biopotential rise after irrigation until it reaches its peak (I), the saturated signal until it starts to fall (II), and the decay of the signal until the next irrigation event (III). As such,
3.1.3 Time-frequency characteristics
The time-frequency analysis described in Section 2.5 was applied to compute normalized wavelet power change for leaf inclination and biopotential in manual irrigation experiments. Figure 5 shows the time-averaged power for two hours after the irrigation for a wide frequency range, as well as time-frequency patterns of power for leaf inclination and biopotential in a frequency range of interest. Since time-averaged power for two hours after the irrigation showed noticeable change in the range of 1 to 5 mHz commonly for leaf inclination and biopotential (Figure 5A), we focused on this frequency range to analyze the time-frequency characteristics of power. Before the irrigation event, while little power change was observed for leaf inclination, momentary power increase appeared for biopotential. Since the pre-irrigation increase in biopotential could not be time-locked to the forthcoming event, this might reflect a decrease in stability and increase in sensitivity to noise. After the irrigation event, however, both leaf inclination and biopotential patterns showed intermittent enhanced power at about 2.5 mHz (highlighted by rectangles in Figure 5B and C). Noticeably, this enhancement began only within 30 min after the irrigation and was time-locked to the onset of irrigation, suggesting that it might be an initial postural response due to the water supply.
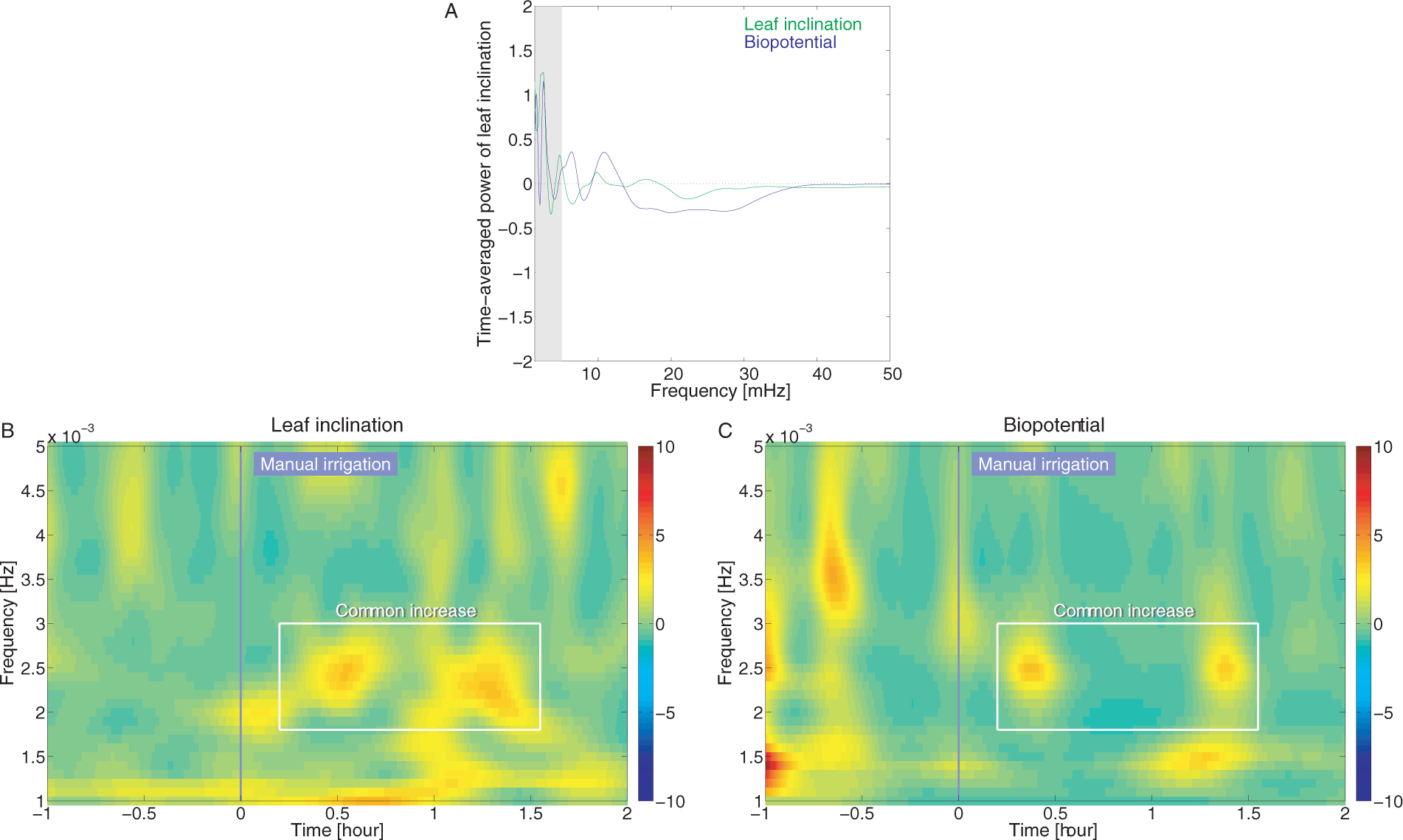
Normalized wavelet power for leaf inclination and biopotential after manual irrigation (
Figure 6 shows synchronization of fluctuation of all leaves in manual irrigation experiments. Time-averaged synchronization from one hour before to two hours after the irrigation also showed noticeable change in the range of 1 to 5 mHz (Figure 6A). We thus focused on the same frequency range as in the power case to analyze the time-frequency characteristics of synchronization. As shown in Figure 6B, leaves randomly fluctuated before water supply. However, after irrigation, they synchronized with each other at approximately 1.5 and 3 mHz, lasting for about an hour.
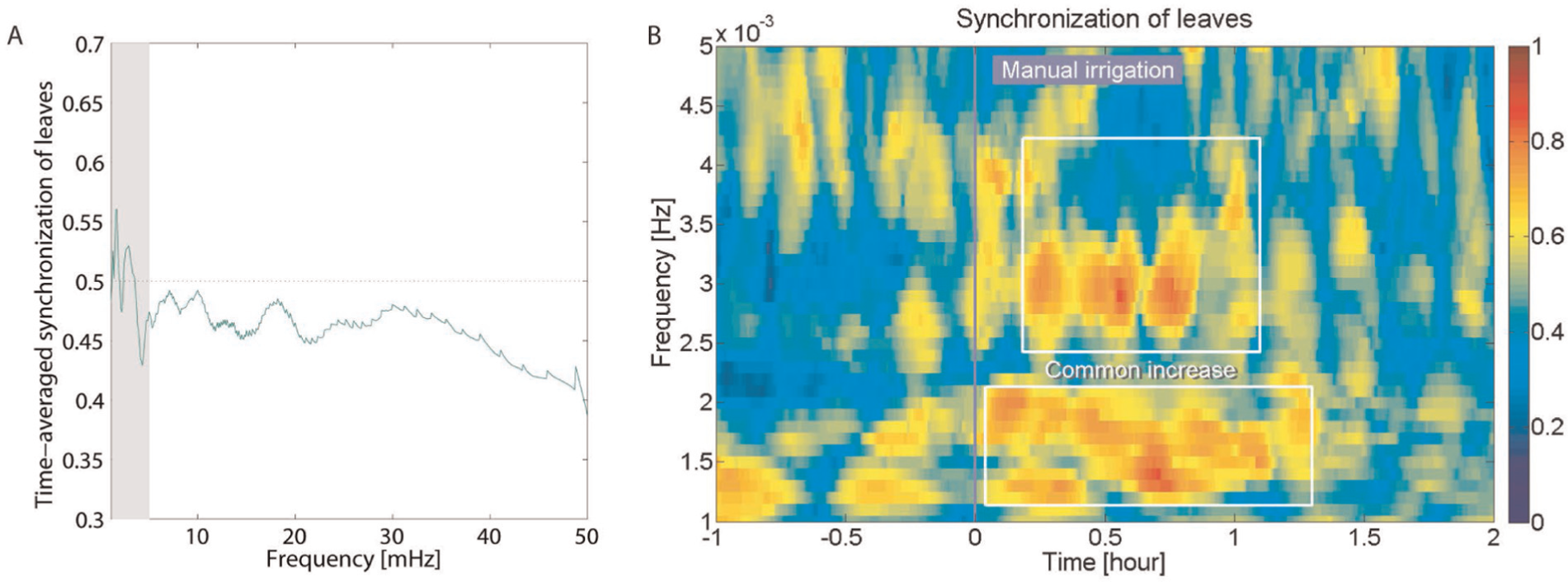
Synchronization of fluctuation of all leaves around manual irrigation (
The synchronization between a leaf inclination close to an electrode and biopotential is shown in Figure 7. Unlike the patterns of synchronization of all leaves, synchronization values were relatively high within the range of 1 to 5 mHz before irrigation. After an irrigation, however, the values decreased and patterns similar to Figure 6, though more complex, appeared at approximately 1.5 and 3 mHz (note that the number of components for the calculation, or
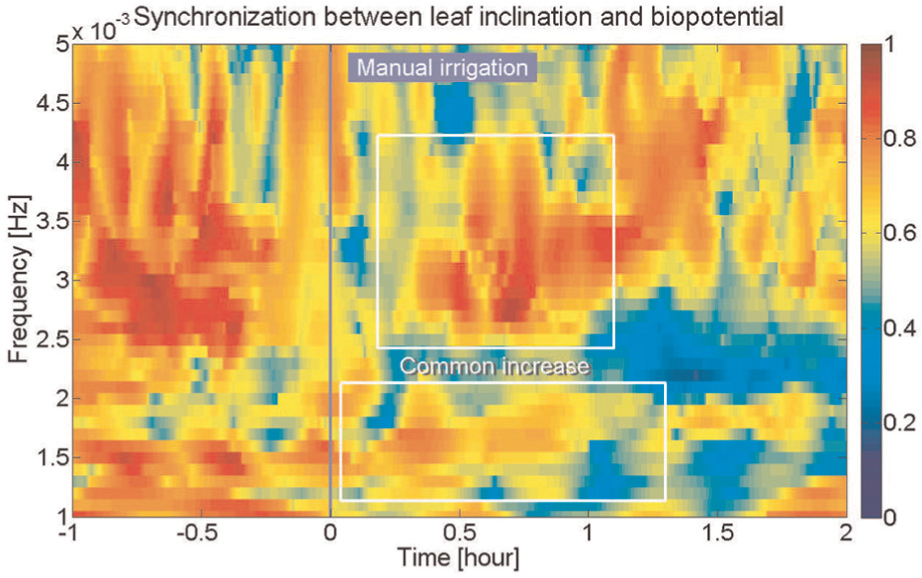
Synchronization of fluctuation between inclination of a leaf close to an electrode and biopotential around manual irrigation (
3.2 Automated irrigation
3.2.1 Automated irrigation by leaf inclination
An initial attempt toward automatic plant-controlled irrigation was conducted based on leaf inclination. The signal from the distance sensors was used to trigger the water supply. The irrigation was stopped automatically by the controller board after two hours.
The plot in Figure 8 represents five independent experiments of autonomous irrigation by leaf inclination. We sampled leaf inclination every 10 s and evaluated the drought condition by comparing the position of the leaves with an offset of 1 cm from their initial (healthy) position. When the plant began to suffer from drought, it lost its rigidity and the leaves inclined slowly. Thus, when the signal from the distance sensors exceeded the threshold associated with the drought condition, the water supply was turned on; it irrigated the plant with 50 ml of water per hour. As soon as the irrigation was triggered, the leaves ascended rapidly and nearly reached their initial state after two to four hours. This experiment demonstrates that leaf inclination detection is able to assess signs of drought in the avocado plant and can be utilized as a control signal for irrigation.
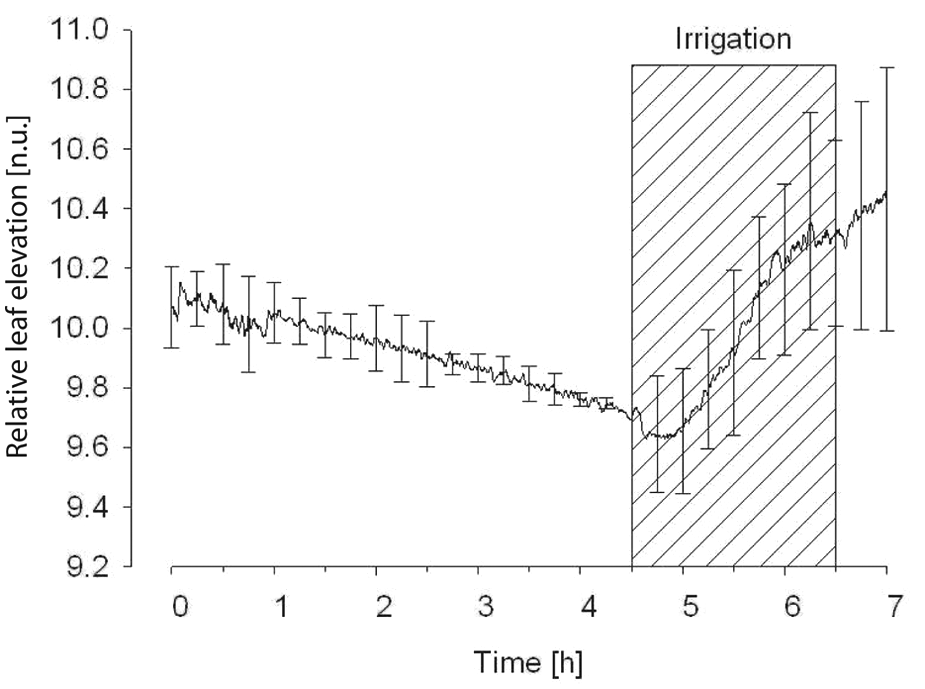
The average normalized leaf inclination signal of five avocados in autonomous irrigation. The shaded rectangle denotes the irrigation period.
3.2.2 Validation of automated irrigation
In order to validate the automated irrigation system, we tested the applicability of the trends revealed in Section 3.1 to the automated case. In contrast to the transient manual irrigation, the automated irrigation lasted for two hours. Time-frequency characteristics of power, which seem less sensitive to the continuous input of water than phase, were thus analyzed for leaf inclination and biopotential, as shown in Figure 9A and B, respectively. Consistent with the manual case, an increase in power change before irrigation was not observed for leaf inclination but appeared for biopotential. Similarly, soon after the irrigation event, both leaf inclination and biopotential patterns showed enhanced power at 1–2 mHz. Though the active frequency was different from that in the manual experiments, these results clearly suggest that the time-frequency power trends are also applicable to the automatic experiment irrespective of the irrigation duration as well as the amount of water, and confirm that our automated irrigation system worked properly with correct feedback from the plant. Hence we used information of leaf inclination as a cue to trigger water supply. The general applicability of the trends, however, suggest that postural changes of plants can be estimated by measuring biopotential in a specific frequency range and vice versa. It means that although leaf inclination could be utilized to fully control the hydration state of an avocado plant, because the biopotential carries information about the metabolic state of the plant, leaf inclination readings can be substituted by biopotential signals. There exist plants that are not suitable for the assessment of leaf inclination, whereas biopotential can theoretically be recorded in every plant. The latter is a universal integrator of environmental influences (Gil et al., 2008) and is more robust for signal acquisition. Therefore, in the future, it could not only replace the tracking of the leaves but could possibly provide enriched information in combination with information of leaf inclination.
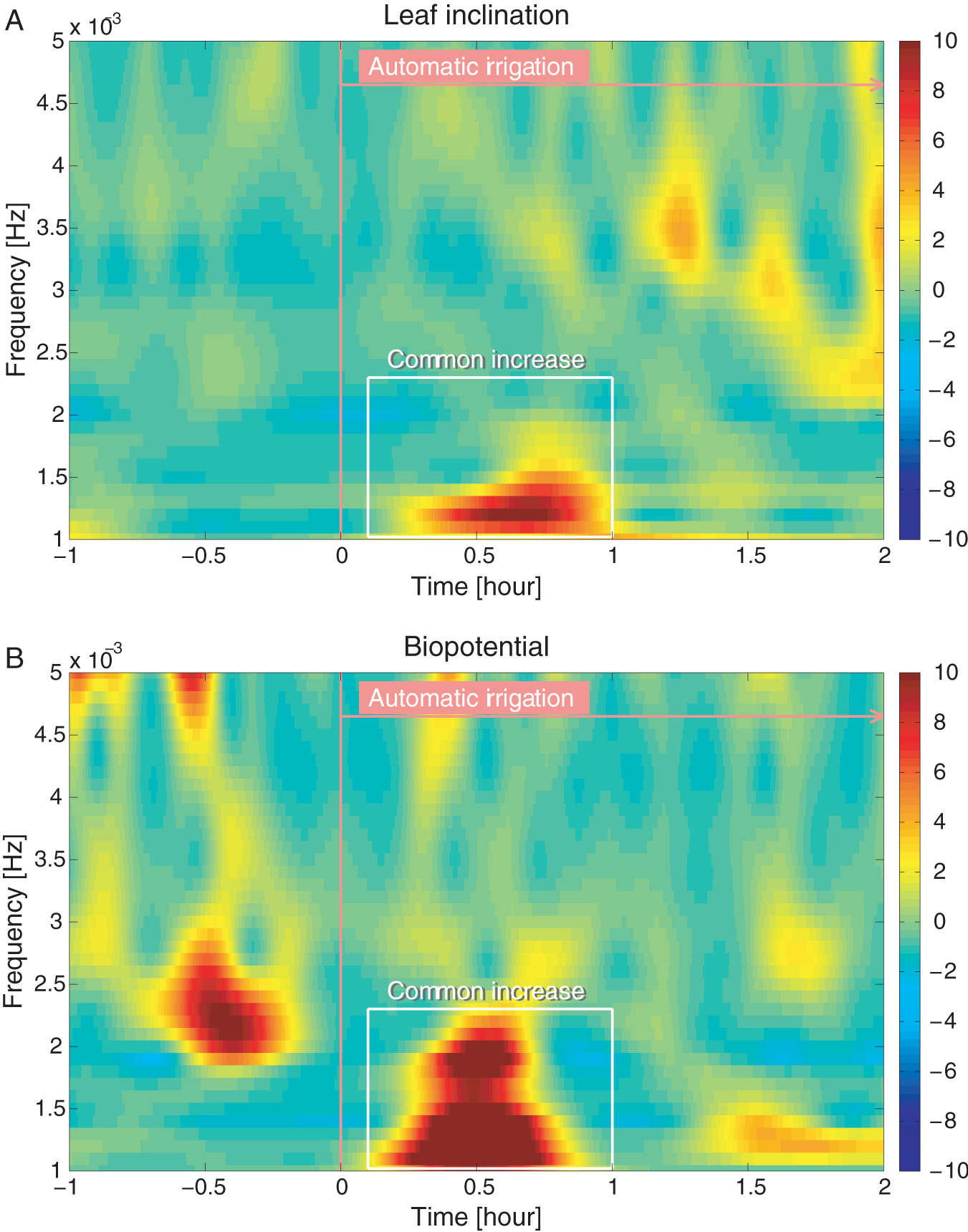
Normalized wavelet power for (A) leaf inclination and (B) biopotential after manual irrigation (
3.2.3 Closed-loop interaction
The results of the experiments described in the previous sections reveal that patterns in the monitored signals exist and can be used to automatically close the loop in plant–machine interfaces and to autonomously control plant resources, for example water. According to the power and phase analyses in particular, these patterns are represented by the decline of leaves that may be used to trigger irrigation and by the rise in biopotential that may be used for stopping irrigation. The algorithm that was developed to control the automatic irrigation of the plant depending on these two variables is depicted in Figure 10. Let
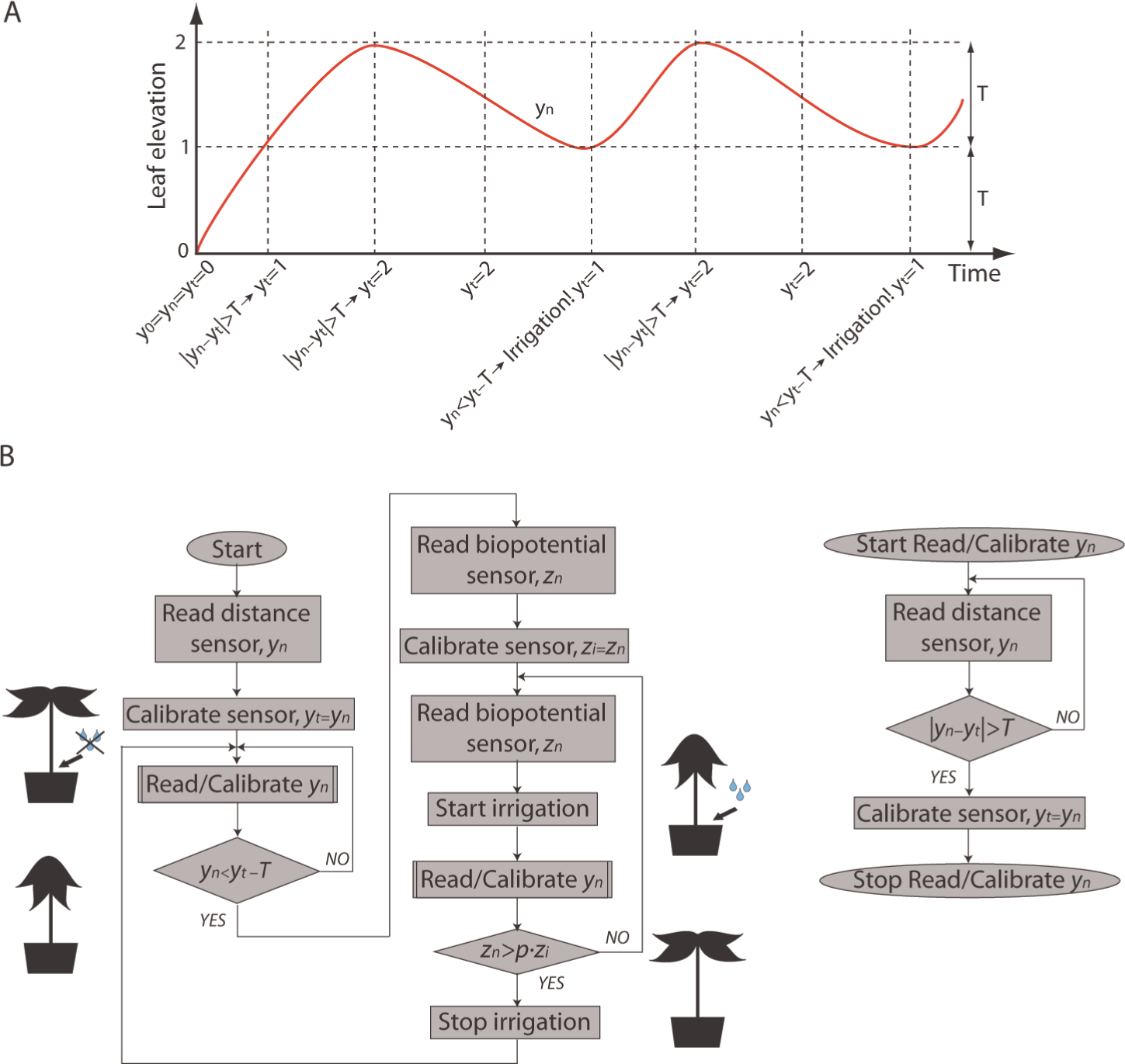
Control of the closed-loop plant–machine interaction. (A) Example of threshold-renewing for leaf elevation.
To stop irrigation, changes in the plant’s biopotential were considered. Our observations pointed to an increase in the biopotential after the avocado received water. This cue was used to stop the irrigation. If
A representative series of pictures in Figure 11A shows the postural changes of the plant during a closed-loop experiment. Figure 11B is a plot of the closed-loop interaction between an avocado plant and the control device. The irrigation increases the leaf elevation as well as the biopotential after two hours. The plot shows that the concept was successfully verified, that is, after one cycle the plant recovers and the process repeats.
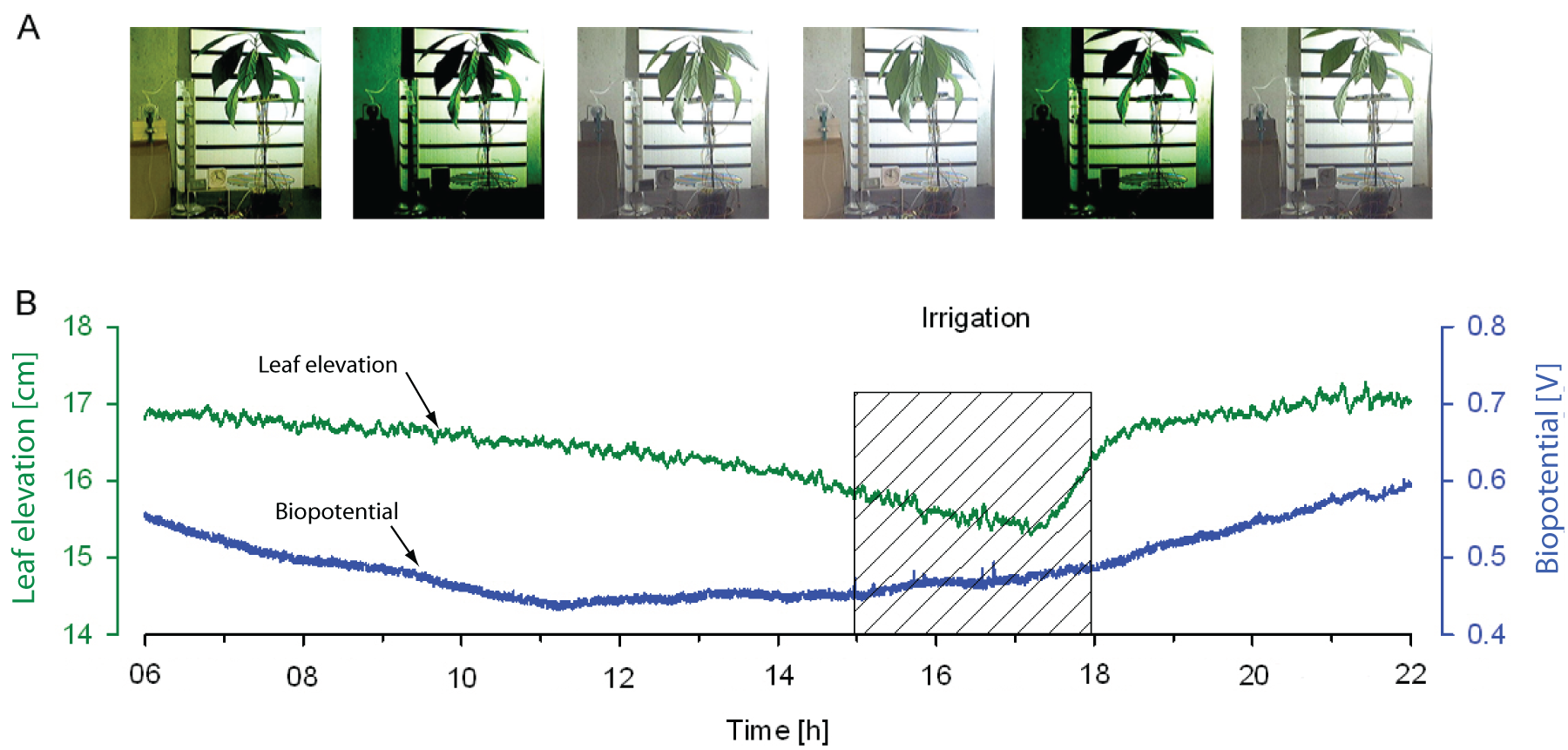
Closed-loop experiment: (A) series of pictures showing temporal posture changes of an avocado plant during an automatic irrigation experiment with increased detection threshold; (B) plot of the average leaf elevation and the biopotential.
4 Discussion
This work took an avenue toward the control of an adaptive biological creature, for example plant, as a black box for an adaptive machine by specifically focusing on the read/out communication aspect. Although the realization of these types of interfaces becomes more and more feasible, there are many challenges to face due to the complexity of the biological component in the system. Some of these challenges are discussed in this section. One of these difficulties is the reliability in the expected biological response of the avocado plant itself. Different avocado plants often displayed distinctive reactions, and even the same sample regularly behaved inconsistently in different trials. The fact that the obtained results frequently showed high variances or even a counter result advocates the difficulty of handling such a black box in an open environment. By averaging trials we reached some consistent reactions of the plant. Moreover, the advantage of avocado (fast-growing) sometimes confronted us with an undesirable situation of iterating experiments under the same conditions. Choosing a rapid growth plant often worked negatively especially when we attempted to set identical conditions in a series of long-term investigations, therefore we resorted to short period experimental trials. However, we note that the steady metabolic processes within a plant produce continuously changing conditions in the plant–machine interface. This uniqueness of each plant condition, the continuously transforming closed-loop system between the plant and machine, and the underlying complexity of physiological computations within the plant become main features of the biological–artificial system, which greatly differentiates it from pure artificial systems.
The growth of plants is heavily dependent on water, nutrients, space (soil and air amount within the plants’ population) and light resources. During a long series of preliminary experiments, we encountered unpredicted variances of the data. The identification of the causes of these variances entails a vast series of experiments to reduce and isolate potential influential factors in a complex system encompassing a biological entity; it could be the humid air due to the rain outside, could be due to the past three (but not two!) days of rain, or even could be because the air-conditioning settings of the building where the plant resides is different during weekends or because the soil was acquired from different supermarket slots and contained different types of nutrient! Experiencing a series of failures in these pre-experiments made us be more conservative, and brought us to develop the ‘caged’ condition of the plant. In this regard, creating the experimental/measurement condition in Figure 1B was not within our intention when the project started. As our goal is for more general environmental situations, restricting the environmental condition did not match our scope. The control strategy may also need to consider the plant’s internal characteristics such as its growth period or fluctuations due to season, in order to sustain an ontogenetic development of plant-based machines in either artificial or natural environments. From the technological viewpoint, while the leaf inclination recording proved stable, the biopotential signal decreased consistently over a long period of recordings. This may be a consequence of plant’s response to a foreign body (the biopotential needle). This condition made it difficult to pursue recording over a period of more than a week. Nonetheless, an alternative method for long-term biopotential recording that avoids inflicting damage to the plant tissue is surface electrodes, which have been used in Gil et al. (2008).
The obtained experiences made us mindful of the importance of acquiring deep knowledge about the complex communication signals of the plant related to its internal state. With this knowledge, we could potentially find improved solutions related to how much water should be supplied to the plant, for how long, in which condition, and so on. Although our future work is oriented toward making the plant learn and realize its optimal conditions, investigations such as those described in this work are necessary. Although we plan to develop automated plant-based adaptive machines for universal use in various plants, the acquisition of in-depth knowledge about the plant component by conducting experimental procedures in controlled conditions is required. This work is therefore meant to provide insightful information about the potential and the challenges in the development of such interfaces.
5 Conclusion
In this paper, prompted by the ontogenetic potential of realizing plant-based adaptive machines, through the realization of automated physiological recovery of avocado plants, we designed a protocol for plant–machine communication by assessing the postural and intrinsic biopotential changes of an avocado plant for the interface with the machine. First, we focused on the fact that postural changes in the plant represented by leaf inclination were able to detect water stress in the avocado plant, and employ this change as a trigger signal for irrigation. Second, having noticed the abrupt change in biopotential after an irrigation event, by analyzing time-frequency behavior, we employed it as a signal for abandoning irrigation. Hence, by carefully assessing patterns in leaf inclination and biopotential signals, we devised an automated irrigation scheme to control the metabolism of the plant and regulate the amount of water available for the plant’s recovery. The mixed control method based on both leaf inclination and biopotential permitted the exploration of communication channels between the plant and the water-control device. The method enabled the comparative study of these two signals and facilitated the interpretation of biosignals based on existent knowledge of leaf dynamics under drought conditions. The results of this closed interaction loop study warrant innovative possibilities for developing adaptive technologies that autonomously regulate plant resources, or that may utilize the intrinsic computational capabilities of plants in conjunction with stimulation techniques for the artificial growth of plant-based machines. At the same time, the study also brings forth the difficulties of plant–machine interfaces that arise from the complex and continuously changing processes in a plant.
Footnotes
Acknowledgements
We would like to thank Kohei Nakajima and Matthias Böller for their advice and assistance.
Funding
This work is supported by the Swiss National Science Foundation within the project #CR23I2 132702/1.