
Abstract
Collective biological systems can exhibit surprisingly rich macroscopic adaptive behaviors from their relatively simple components. These behaviors often resemble sensing, cognition, and ecological interaction, as in higher-order organisms. Unfortunately, a normative model explaining cognitive behaviors from the interactions of simple elements is not yet available. We hypothesize here that minimal self-knowledge can be observed from local interactions between simple components if two conditions are given: (i) self-generated, globally shared information and (ii) higher-order feedback loops emerging from the actions of simple elements. To evaluate the hypothesis, a discrete-time, discrete-space computational simulation is developed, modeling simple particles discharging a chemical-like substance (pheromone). The results indicate that unstructured variations introduced through random movements of particles across the arena enabled the formation of clusters with complex, emergent behaviors. Under the hypothesized conditions, these results indicate that a minimal form of self-knowledge emerges, representing a primitive self-definition.
Highlights
• Particle-pheromone interactions organize into differentiated clusters, enabling the emergence of goal-driven-like behaviors. • Clustered movements of particles are built over blind randomness. • Emergent behaviors require global self-information sharing. • Higher-order feedback loops generated by particle actions enable self-definition.
Introduction
A minimal circuit for self-knowledge refers to the basic, foundational level of self (Limanowski & Blankenburg, 2013). Self-knowledge represents the ability to recognize, respond to, and update based on one’s prior actions. This concept also involves some degree of pre-reflective experience of being an organism distinct from the environment and other individuals.
A minimal self is often linked to both the sense of agency (Di Paolo, 2005) in terms of “feeling like one” given the cause of their actions, and ownership regarding “feeling like a part” or experience belonging to oneself. That understanding of agency and ownership suggests that a minimal self is crucial for knowing how the world is experienced and the interactions within it. We can speculate that even at the extreme of minimal sensory capabilities, a system would still have a rudimentary sense of self, a minimal self. In this respect, the minimal self is the foundation of self-knowledge, providing the basic mechanism for experiencing the world and allowing for more complex self-concepts to develop (Di Paolo, 2005; Forch & Hamker, 2021; Maturana & Varela, 1980; Roush, 1984).
A self-organizing system showing minimal self should show emergence of collective knowledge, which can surpass the knowledge of individual members even when those individuals possess limited information or capabilities. This understanding refers to the “wisdom of the crowd,” which relates to Galton’s observation of crowd estimations at a livestock fair (Surowiecki, 2004). Such wisdom could be seen as emergent from simple elements (Strogatz, 2024). It is unclear yet what mechanisms enable such emergence in relatively simple scenarios because higher-order feedback loops should be implicit in affecting the actions of simple components.
Despite current literature on minimal self (Forch & Hamker, 2021) and collective intelligence in simple populations (Strogatz, 2024), we do not currently have a normative, falsifiable computational model that derives self-knowledge from simple interactions. In this respect, Forch and Hamker (2021) explicitly framed current models, suggesting that the current knowledge of minimal self is incomplete and methodologically diverse, rather than defining a single normative, falsifiable account. Similarly, collective-behavior literature shows mature mathematical models of simple-population collective intelligence with no general, normative account (Strogatz, 2024). Indeed, these literatures rarely converge with minimal-self modeling to derive individual self-knowledge as an emergent, testable prediction of simple interaction rules.
This article takes a first step toward connecting self-knowledge with self-organization. We start modeling finite-state agents (particles) with minimal sensorimotor access to derive emergent collective feedback. The same formalism takes into account both the individual-level and the group-level contributions. This work reports analyses using computational modeling with indirect coordination, defining simple particles that modify a shared environment through the diffusion of pheromone-like substances (Theraulaz & Bonabeau, 1999; Wyatt, 2014). These substances in turn locally constrain/afford the next actions of particles. Thus, the system-level organization is produced via environment-mediated traces rather than internal models of others’ states (Theraulaz & Bonabeau, 1999).
The main goals of this work are: (i) to implement a discrete-time, discrete-space computational model where two types of particles interact by means of pheromone diffusion and (ii) to discuss how emergent collective actions of particles arise from their local interactions through particle clustering. This approach allows us to quantify how local interactions and pheromone dynamics give rise to unexpected global behavior. Systemic entropy and interaction influence between particles are measured to elucidate the mechanisms underlying global behaviors in a simplified but illustrative proposed system.
Related Work
The notion of a minimal self is now fundamental in both the philosophy of cognitive science and social/cognitive neuroscience. According to the most widely shared view, the minimal self designates a pre-reflective, embodied, perspectival sense of being the subject of experience in the present moment (Limanowski & Blankenburg, 2013). The self-knowledge associated with minimal self is not propositional or autobiographical because it denotes the ability of the system to recognize itself, respond to, and update from the traces of its own past activities. To put it differently, it means enabling the distinction between self-produced effects and other-produced effects to improve one’s future performance or predictive ability.
Enactive (Froese, 2012) and autopoietic (Varela et al., 1974) literature go further by locating the minimal self not in representations but in organizational closure. The system should be a self-producing and operationally closed dynamical entity whose identity emerges from ongoing sensorimotor coupling with its environment (Di Paolo, 2005; Maturana & Varela, 1991). On this view, the minimal self is a mechanistic consequence of a closed loop: action → world → sensed consequence → internal update. Computational models compatible with this view have been put forward to some extent. Though most extant models either assume continuous-valued state spaces, rich sensory channels, or internal architectures large enough to effectively presuppose the self in their design (Forch & Hamker, 2021). What is still missing is a truly minimal, formally transparent, and discrete implementation in which the transition from action traces to self-knowledge can be observed and quantified directly from the dynamics.
Consider also that a literature running in parallel explores how collective intelligence can arise within systems comprising many individually simple components, surpassing any level of information that may be accessible to individuals (Surowiecki, 2004). In this, dynamical systems work shows emergent high-level competencies arising out of local positive/negative feedback interactions and phase transitions in networks of simple units (Strogatz, 2024). Unfortunately, the bridge from the minimal-self mechanisms at the individual level to higher-order feedback loops at the group level is unclear. Yet, in all those domains, we only have either (i) phenomenology-first philosophical accounts of minimal self (Gallagher, 2000) or (ii) empirical/algorithmic models of simple distributed collectives (Sumpter, 2006), but not a formally minimal, finite-state, discrete-time, executable circuit that expresses the minimal self and can be multiplied to test scaling.
There is accordingly a clear gap at the intersection of these literatures. We lack explicit, minimal computational models that demonstrate: (1) how the minimal self, defined operationally as the ability to recognize, respond to, and update on one’s own prior actions, can emerge from the smallest possible circuit operating in discrete-time and state and (2) how ensembles of such minimal selves can give rise to higher-order feedback loops at the group level that exist in none of its individual components. Bridging this gap is more than a stylistic oversight because it presupposes a mechanistic, low-level description and the conceptual commitments about the foundations of self-knowledge. If the minimal self is truly foundational, then even in extreme sensory impoverishment, we should still be able to implement it in a minimal circuit, and we should be able to measure some consequences of it. Conversely, collective intelligence in simple populations depends on the existence of such minimal self-mechanisms within individuals. In that case, it will become visible only when the minimal circuits are implemented and tested at scale (cf., (Couzin, 2009)). This understanding goes against a purely statistical view of aggregation. In other words, collective computation in biological collectives is not simply a by-product of sampling or averaging but is known to depend on agent-level stateful feedback (Franks et al., 2002; Sumpter, 2010).
The work presented here is an attempt to fill this gap by implementing a discrete-time, discrete-state minimal selfcircuit to see whether higher-order feedback emerges. In this way, we render the conceptual claims about the minimal self computationally falsifiable, and we directly test whether group-level competence does indeed require individual-level minimal self-mechanisms.
Methods
Pheromone Diffusion
Space is a two-dimensional
Stigmergy
These patches enable particles to search for pheromones to transform, where yellow particles transform blue pheromones into green, and red particles transform green pheromones into blue. Changes in concentrations of pheromones may stigmergetically (Marsh & Onof, 2008) influence other particles through the environment, modifying locations and consequently variations of gradient sensing. In this context, stigmergy refers to a mechanism of indirect coordination through the environment, where particles influence each other’s actions by modifying shared surroundings. Note that the purpose of transforming pheromones is to create potential fields that guide the collective behavior of the same particle type. This transformation enables spatial communication and coordination between particles of the same type. The diffusion process, together with pheromone laying by particles, creates concentration gradients to locate pheromones to transform. The pheromone transformation encourages nearby particles of the same type to explore the vicinity of successful paths (exploitation).
Pheromone Concentration
A specific number of simple particles (8000 in total) or pheromone is contained in each patch. Pheromones diffuse to neighboring patches in equal (100%) or different (50% higher or lower) proportions relative to the other pheromone type at each time step. The concentration level of red pheromones is kept in [−20; 20], and for yellow is [−10; 10]. These pheromone concentration levels are chosen based on empirical tuning. The difference between particle-type concentration levels prevents pheromone steadiness and induces particle motility. The concentration gradient of pheromones is calculated for each patch and for neighboring grid patches where particles are located. Diffusion takes place at one patch per time step. A patch containing no particles is visualized as a combination of normalized concentrations of green and blue pheromones. If a patch contained one particle, its color depends on the type of particle (Figure 1). Spatial density distribution maps across the arena for the combined pheromone concentrations. Plots are shown every 100 simulation time steps. Yellow and red dots represent particles, and pheromones are represented by green and blue patches. Particles search for pheromones to transform, where yellow particles transform blue pheromones into green, and red particles transform green into blue pheromones. The experimental configuration is based on pheromone with equal diffusion for yellow and red particles (20 a.u.), and equal gradient sensing for both particle types (5 a.u.) (see Methods)
Pheomone Gradient
Random movements of particles are modeled as an arbitrary deviation in [−1; 1] for
Pheromone Diffusion Process
The process determines the dynamics of the sensed gradient, which increase the pheromone area over time around a cluster of particles. Note that the pheromone gradient of one type of pheromone is defined as going from the cluster’s high concentration (typically at the center of the cluster) towards lower concentrations represented as the boundary of the cluster. In this respect, particles move for clustering towards the same direction to the perceived maximum concentration of their produced pheromone (positive influence). Also, particles move towards the opposite direction to the consumed pheromone gradient (negative influence). No new particles appear while the simulation is running, and the sum of pheromones (green and blue) is conserved between 0 and a control parameter in the model.
Systemic Entropy
To quantify the relative organization and adaptation of clusters across the arena for each type of particle, the spatial entropy of the system featuring the spatial distribution of components is measured. This measure is based on Shannon entropy computed over the grid of patches (Batty, 1974; Batty et al., 2014). The temporal variation of this spatial entropy follows the standard entropy interpretation of spatial order (Gleiser & Stamatopoulos, 2012):
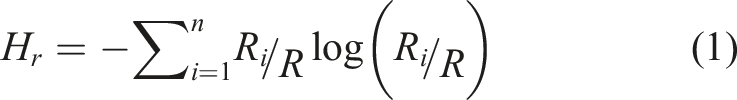
Particle Interaction Entropy
This measure characterizes the influence of particle interactions regarding cluster formation (Cover & Thomas, 2005; Gleiser & Stamatopoulos, 2012):
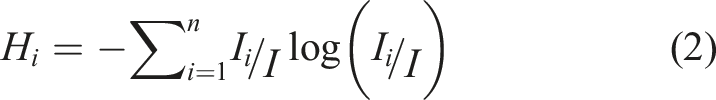
Statistical Comparisons
RM one-way ANOVA, and RM one-way ANOVA-Multiple comparisons (p < 0.01), GraphPad Prism 8; see Suppl. Information.
Results
Autonomous particles in the proposed model are capable of movement, interaction, pheromone sensing, and deposition. The environment, as a medium for interaction, is not passive but dynamically co-constituted by particles and pheromone gradients. The particle-pheromone system is operationally closed because every process (movement, deposition, and sensing) is internally generated and maintained by other processes within the system.
The example snapshots shown in Figure 1 indicate that the dynamics of the model exhibit two phases: (i) interactions among particles of the same type within and between near clusters and (ii) dispersal of particles between patches, generating a change in the clusters’ area and their distribution across the simulation. In the first phase, the interaction of particles within each cluster is described by an independent and non-randomly oriented search when the pheromone gradient is sensed, without any usual feature, rather than following the pheromone concentration (see Methods). Clustering is surrounded by their produced pheromones that represent a heterogeneous environment across the arena due to the different levels of pheromone concentration in each cluster, but are homogeneous inside clusters. In the second phase, the dispersal occurs when the sensed pheromone concentration is higher outside a cluster, producing the movement of particles passing from one cluster to another, which redistributes particles to immediately neighboring clusters. Particles that cannot sense the gradient remain in their native clusters, and the dispersal is spatially symmetrical for all types of particles.
The observed spatial patterns and their stability, dynamically speaking, can be assimilated to a basin of attraction due to perturbations in the face of competition for space from the other particle type. Here, we have a key for understanding the dynamics of the observed organizations: a cluster maintains a stable organization through self-organization. Note that during the simulation, boundaries appear between clusters, one type of particle from another (Figure 1). This coexistence represents a balance between produced and consumed pheromones, which is a trade competitive against the dispersal abilities of the other type of particle.
To quantify the relative organization and adaptation of clusters across the arena, and to understand the influence of particle interactions regarding cluster formation, it is measured the entropy and particle interaction in a function of pheromone diffusion for the yellow and red populations of particles (see Methods). Figure 2 represents the mean systemic entropy ( Results obtained with yellow and red particles when changing their relative pheromone diffusion level. Organization measure as (A–B) systemic entropy (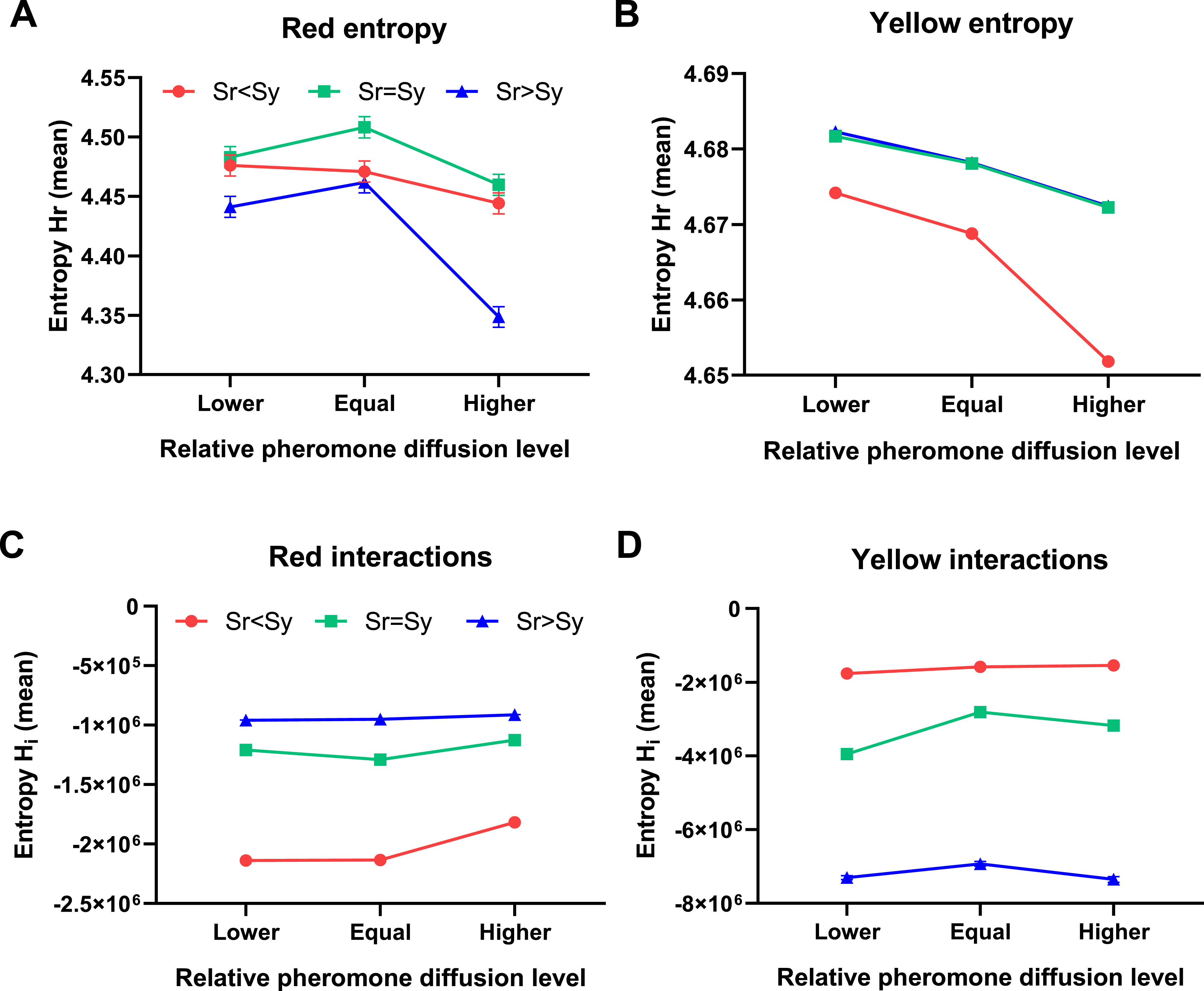
Coexistence, in the sense of a relatively high level of organization (
Interestingly, when the pheromone diffusion of the two types of particles differs by two times, the coexistence produces relatively faster movements (Figure 3). This phenomenon could resemble ‘attacks’ from clusters with faster particles to clusters with slower ones, as observed occurring for a wide range of relative levels of organization (Aon & Cortassa, 1997). Such coexistence (Hassell et al., 1994) tends to self-organize clusters between particle types with less spatial separation, which in turn helps to connect clusters of slower particles (yellow) given perturbations from clusters with faster (red) particles as observed in Figure 3. Example of faster (red) motion exhibiting a form of rudimentary goal-directed, red-to-yellow motility resembling an attack-like display. Each panel represents a small section of the arena. Experimental configuration based on pheromone equal diffusion for yellow and red particles (20 a.u.), and equal gradient sensing for both particle types (5 a.u.) (see Methods)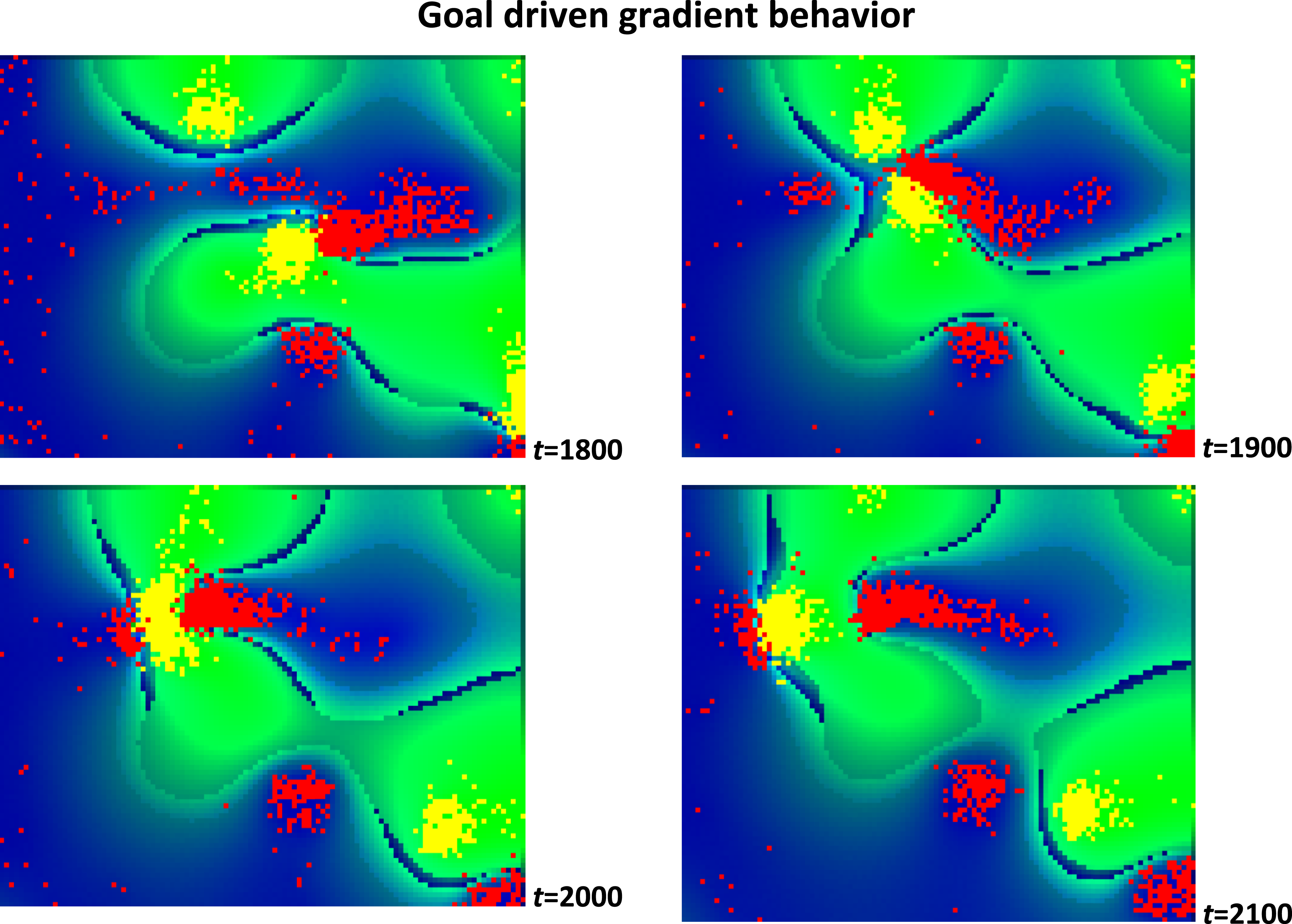
These observations indicate that the two competitive types of particles with relatively different pheromone diffusion rates enable the emergence of unexpected behaviors. For example, experimental conditions defining relatively slower particles tend to remain initially in small areas of the arena till the produced pheromones diffuse, connecting nearby clusters of the same type (Figure 1). This phenomenon indicates spanning clusters of pheromone diffusion that percolates between clusters of the other type of particle (Rivier, 1987; Stauffer & Aharony, 1994). Furthermore, isolated clusters can persist with a particular density very much lower than that of the more dispersed clusters. This occurrence happens because the more dispersed a cluster is, the more particles have transforming pheromones.
Discussions
The results indicate that when a cluster depletes the pheromone in its immediate vicinity, it tends to move towards areas with higher concentrations. This can be seen as an adaptive behavior (Fantino & Staddon, 1985), allowing the cluster to seek out more dominant gradients. Furthermore, clusters may also exhibit a form of emergent path finding. Although individual particles are only sensing local pheromone concentrations, the cluster as a whole is able to move towards a distant pheromone source. This observation implies that a rudimentary, but collective decision-making process is at work. In this view, the decision on the direction of movement is not held by any single particle. Instead, it is continuously created through local particle interactions that bias the flow of the entire population, resulting in a coherent choice of direction at the level of the population.
Clusters can merge with other clusters or break apart, depending on the distribution of pheromones. This could give rise to adaptive behavior in the light of changing conditions and spatial distribution of particles. We emphasize again here that the “wisdom” of these cluster movements is not preprogrammed into the individual particles. Rather, it results from the interplay between particle behavior and pheromone dynamics. This represents a first important feature of complex systems: simple interactions can give rise to complex and adaptive collective behaviors even when behavioral structure emerges over blind randomness.
Therefore, the emergent circuit-like loop for minimal self-knowledge can be summarized as follows:
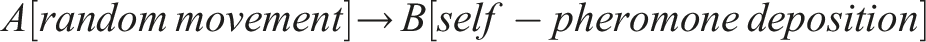
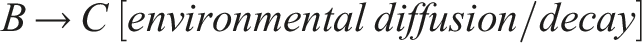
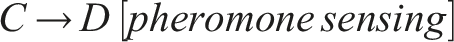
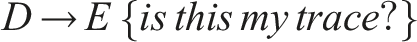
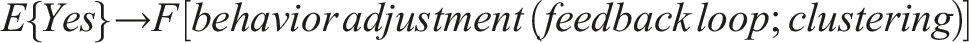
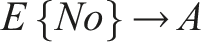
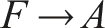
This circuit does not require internal memory and utilizes the externalized environment as working memory for robust behavior (see also Fernandez-Leon, 2011, 2014), enabling temporal self-distinction.
These results both add to our knowledge of collective complex behavior in the simulated systems and provide potential insights into a range of real-world phenomena. The particle-pheromone interaction model, for example, might find counterparts in self-organization dynamics concerning neural-based systems: systems aiming at an integrated global map of space from local sensory inputs, such as described in (Buzsáki, 2009; Dong & Fiete, 2024; O’Keefe & Nadel, 1979), or internally representing space as suggested in (Kropff & Treves, 2008; Zilli, 2012). Above all, this quest underlines a general principle of complex systems: intelligent behavior can self-organize from the bottom up by simple rules and local interactions alone. In this perspective, traditional approaches based on top-down control are contrasted with the ability of autonomous systems to self-organize into complex and adaptive patterns.
Global vs. Local Order
The consumed pheromones determine the movement of a cluster (global order) based on changes in the pheromone gradient landscape (local order). The movement of particle clusters in this simulation warrants further discussion. While individual particles follow simple rules, their collective motion exhibits a degree of coordination that can be interpreted as a form of rudimentary goal-directed behavior (Figure 2). In other words, there is a pheromone-driven coordination in which clusters move in response to pheromone gradients. This means that the particles within a cluster are not moving randomly, despite the definition of their motility, in which they are responding to a global signal in their environment. The signal is, in fact, a product of their collective activity, creating a feedback loop that guides the particles’ movement (Figure 4). Schematics representing the emergence of minimal self-knowledge in the face of goal-driven behavior. The particle-pheromone interactions serve as a powerful organizing principle, leading to the formation of differentiated clusters that exhibit goal-directed-like behaviors. Complex patterns of movement and organization emerge not from centralized control but are built upon blind randomness, where unstructured particle motion lays the groundwork for adaptation. Crucially, global information facilitated by self-generated pheromone fields enables the system to maintain coherence and coordination across space. As particles of the same type interact, they generate higher-order feedback loops, reinforcing local structures. These loops effectively guide future actions, allowing the collective to adapt dynamically and exhibit behavior that mimics goal-driven actions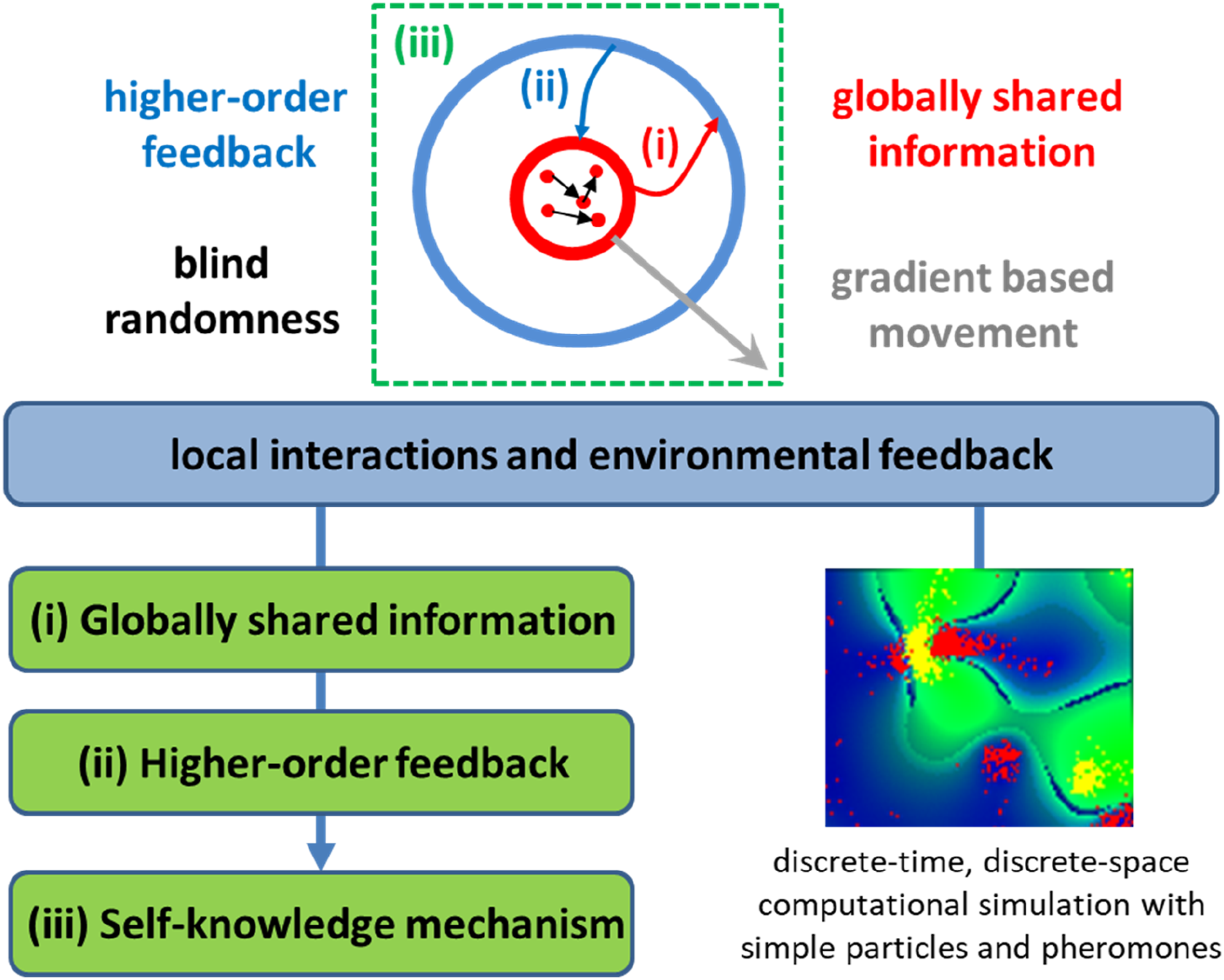
The experimental implications convey some possible observables: (a) clustering where particles follow their traces; (b) avoidance patterns where self and other traces are distinguished; and (c) looping behavior, even without explicit memory. As the gradient sensing of the two types of pheromones becomes less similar (Figure 2), the faster they dominate progressively larger areas around the arena, mainly when its pheromone diffusion rate is slow. This observation suggests that a stable balance between production and dispersion of pheromones within clusters (spatial patterns) is not possible without the interaction between types of particles in competition for spreading. In addition, the local dispersal combined with inherently unstable local particle dynamics can lead to self-organized persistent spatial structures in space despite spatio-temporal differentiation between types of particles. These results indicate that the coexistence and differentiation between strategies of clustering are promoted by a trade-off between local competition and global dispersion, and also convey a balance between the persistence of clusters and the sensitivity of particles to move to different clusters with higher pheromone concentrations.
Goal-Driven Behavior
The emergence of complex behavior from simple particle interactions can be loosely understood as a higher-order behavior. In other words, the particles operating with simple rules collectively exhibit a higher level of organization that goes beyond the capabilities of any single particle. This collective behavior can be interpreted as a primitive form of reflection or computation on the system’s state, analogous in some sense of its own state (Dahaene et al., 2017). More specifically, the simulation demonstrates an emergent pathfinding behavior, where clusters of particles navigate towards a pheromone source. This could be seen as a rudimentary form of goal-directed behavior, which aligns somehow with the concept of higher-order cognitive processes (Dehaene, 2014; Grossberg, 2021).
While individual particles do not have a global map, the cluster as a whole demonstrates a capacity to reflect on the environment and find a more probable path. The global sharing of information between clustered particles is then necessary for the emergence of cluster movements based on the interaction of simple particles. Movements of a cluster toward a common gradient sensing become the focus of additional processing for sensing the gradient towards where to move. Thus, high-level information is made available to the entire cluster for “deciding” where to move (see also (Froese, 2012)).
Note also that the gradient information and the repertoire of self-knowledge available by the cluster’s pheromone have enough relevant information to enable the cluster to move, but this knowledge should be constantly updated. This phenomenon implies that the system has hidden information, where particles in each cluster cannot see the whole pheromone map at once, but the cluster should explore it locally. So it is not a perfect information system based on partial information situations, which is more like the real world. Parts of the system (cluster) have to infer everything else to come up with a more probable gradient. It remains unclear however, how the randomness of the interaction between particles in a cluster acting as an “irruption” phenomenon (Froese, 2023) enables the emergent behavior in terms of unpredictability and entropy in clusters as the material basis of behavior.
Complex Behaviors from Blind Randomness
Particles in the model rely on blind randomness as unstructured variation introduced through random movements to explore the arena. In contrast, it is possible to think that clusters enable emergent behaviors using built-in knowledge or biases in the movements of particles that guide the search of the cluster in the arena. This paper argues that the cluster movements over blind randomness are not only more aligned with biological intelligence but also observable in some complex adaptive systems. The limits of randomness become increasingly apparent in the proposed model, mainly because the prior action effects are proposed in the model definitions. In this, each particle or cluster of particles in the modeled system is not able to reason, generalize, or anticipate the trajectory but just senses the more dominant gradient. Thus, the system cannot simulate and plan the next movements using internal models of the particle or cluster, which implicitly require structure but not randomness.
The reported results exemplify that clusters do not explore moves at random. It is possible to speculate that blind randomness as an unstructured, unguided variation in the model has several further limitations, making random search infeasible to find the pheromone gradient, and that random changes are not informed by past actions. Structured prior actions seem then a way to explore the arena in that the sensing of the gradient by a cluster constrains and eliminates invalid candidate movements. The joined actions of particles within a cluster proposed thousands of outcomes using the interaction of particles ending up in unexpected, emergent behavioral strategies that outpace human intuition.
Conclusions
The present research explores how the joint interaction of simple particles might appear much more sophisticated and coordinated than anything possible by an individual in the system. Minimal self-knowledge emerges when clusters use environmental feedback (pheromones) based on their past actions, leading to implicit modeling of the cluster’s actions vs. the actions of other clusters in the environment. Clusters of particles emerge in forms that stabilize over time, in which pheromone dynamics regulate where new clusters form or dissolve. Each cluster of particles and its trail network maintains itself and responds to perturbations (e.g., dispersal and reformation). The within-cluster concentration is locally unstable due to the activity of particles inside clusters. Any global persistence of particle population results from the spatially distributed nature of interactions (see (Hassell et al., 1994) for similar arguments).
Results indicate three main mechanisms in the system. First, particles’ reactions are observed given the defined model, which does not directly influence the behavior of the cluster of particles. Second, shared pheromone distribution information is accessible by clusters of particles and acted upon deliberately, which involves global broadcasting of gradient information to elements within the cluster. These mechanisms also enable a cluster to reflect on its organizational state, which involves basic self-monitoring supporting goal setting. Discussions are also focused on the particles in the model that rely on blind randomness to move across the arena. Further, clusters enable emergent behaviors using structured priors that build in knowledge or biases in the movements of particles that guide the search behavior of the cluster.
The seemingly complex behavior of particle clusters, such as coordinated movement, emerges from particles sharing information via generated pheromones. In essence, both features illustrate that complex, higher-level coordinated movement in the simulation results from the collective exchange of information as self-generated knowledge among simpler units. This observation is contextualized in the existence of higher-order processes enabling the ability to reflect information on the cluster’s own dynamics and actions; see also the enactive concept in (Froese, 2012; Froese et al., 2024; Rupert, 2016). The proposed discussions contrast with traditional views emphasizing centralized control or complex individual programming to explain organized behavior. Instead, it is demonstrated here that coordinated movement can arise from decentralized interactions and environmental feedback loops. These observations suggest that a minimal form of self-knowledge under the hypothesized conditions can emerge as a primitive self-definition by means of a recognition loop.
Supplemental Material
Supplemental Material - A Self-Knowledge Minimal Circuit Emerges From Blind Randomness and Environmental Feedback
Supplemental Material for A Self-Knowledge Minimal Circuit Emerges From Blind Randomness and Environmental Feedback by Jose A. Fernandez-Leon in Adaptive Behavior
Footnotes
Acknowledgments
Thanks to Alexander Klyubin for defining the conditions in an early version of the model. JAF is currently supported by The National Scientific and Technical Research Council (CONICET), Argentina, through the Scientific and Technological Researcher Career program (DI-2019–2516-APNGRH-CONICET).
Ethical Considerations
This article does not contain any studies with human participants performed by the author.
Consent for Publication
The author affirms that human research participants or related data were not included.
Author Contributions
JAF-L: Conceptualization, Investigation, Methodology, Software, Data curation, Data and Formal analysis, Validation, Visualization, Writing—original and final draft, Writing—review and editing, and funding acquisition.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: JAF-L is supported by The National Scientific and Technical Research Council (CONICET), Argentina, through the Scientific and Technological Researcher Career program.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Clinical Trial Registration
The author affirms that there is not clinical trial information associated to this manuscript.
Code Availability
The custom code used in this paper is available upon reasonable request from the corresponding author.
Permission to Reproduce Material From Other Sources
Permission should be obtained in writing from the author if all or part of any of the figures or text is to be used in a publication.
Supplemental Material
Supplemental material for this article is available online.
Author Biography
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.