
Abstract
The coordination of competitively coupled systems is notoriously complex. One-v.-one attacker-defender interactions (1v1s), common in team sports, provide a vehicle for investigating the role of nonlinear interactions between timescales during competitive coordination. This study tracked movements of 20 amateur players in a 1v1 futsal task, modelling the attacker’s leading role and the dyad’s nonlinear interactivity with an anticipatory-synchronisation metric and multifractality, respectively. Anticipatory synchronisation encoded how well an attacker could pre-empt a defender’s movements. Multifractality encoded variability and comparison to linear surrogates encoded nonlinear interactions across scales. Generalised linear mixed-effects modelling showed that attackers scored more points when they anticipatorily synchronised with defenders’ movements and promoted greater multifractal variability in defenders, as well as in a collective variable pooling dyadic variability. Attackers scored more when they themselves constrained their own multifractal nonlinearity to generate targeted attacks. Thus, competitive coordination in invasion games rests on nonlinear cross-timescale interactions, not just linear sequences of perceptual, cognitive, and motor processes. Explanations of 1v1s may need to address both independent-player contributions and shared nonlinear interactivity. Finally, we discuss avenues for future research on the multiscale dynamics of competitive co-adaptation in sport and related contexts.
1. Introduction
Social coordination involves a rich interplay of pursuing goals and evading obstacles – often with other agents, but sometimes against them. In the latter case, the obstacles may even be other agents pursuing their own goals. Sport offers a variety of competitive or ‘antagonistic’ (De Poel, 2016) coordination. For instance, invasion games pit opposing players against each other to manipulate an object, such as a ball, into goals or target areas at opposite ends of a symmetrical playing area. The one-v.-one sub-phase (henceforth 1v1) of invasion games arises when the interpersonal distance between an attacker and a defender decreases, and their movements as a dyad become increasingly coupled (Passos et al., 2016). The 1v1 phase exemplifies antagonistic coupling in which the attacker attracts the defender, and the defender repels the attacker (De Poel, 2016). The present work examined the 1v1 futsal dyad to probe for underlying dynamics of antagonistic coupling. The 1v1 dyad is methodologically simplifying without losing touch with what is fundamentally challenging about antagonistic coupling. In 1v1s, opposing objectives unfold according to a set of selected constraints whose interaction with the movement of participants defines a particular sport (Farrokh et al., 2025).
Antagonistic couplings unfold at many timescales, ranging from predator-prey pursuit to co-evolutionary ‘arms races’ and often complement more affiliative coordination (Dotov & Paxton, 2024). Mutual cooperation between agents is not always guaranteed, so the processes supporting adaptation in antagonistic couplings may be implicated in a variety of the complex intelligent behaviours of evolved organisms. Although the generality of antagonistic couplings provides ample motivation to study them, progress has been a challenge. Antagonistic couplings appear in such a wide variety of organisms that explanation will not find simple grounding in any specific process or structure of a nervous system (Turvey, 2018). Indeed, a comprehensive framework for studying the neurobiology and biomechanics of perceptual and motor processes which underpin antagonistic couplings such as predator-prey interactions remains elusive (Peterson et al., 2021). Despite their ubiquity, antagonistic couplings raise complex epistemological questions. For example, the need to relate the movements of an individual to the dyad within which they participate raises a number of difficult questions. These questions include the locus of intentionality, the structure of temporality, and the nature of unit or self. It is thus advantageous to investigate these couplings in less anatomical and more functional terms, using nonlinear-dynamics tools for modelling interactions across time scales, considering both individual and collective variability (Kelso, 1995). The present work addressed antagonistic coupling in a dyadic sport paradigm, examining how each agent led or followed the other and effected nonlinear interactions across time scales.
Much of the difficulty understanding antagonistic co-adaptation is in the modelling of nonlinear interactions across scale. These interactions appear both at the scale of individual agents and at the scale of the dyad. Any context-independent strategy, defined at the level of the individual (e.g. a pattern of zigs and zags), is susceptible to exploitation by counteracting moves. Behavioural science’s bias towards methodological individualism means that the contribution of independent actors and independent processes has overshadowed interactive, dyadic, and collective actions (Davids & Araújo, 2010; Dunwoody, 2006). The prevailing assumption has been that compiling the discrete and isolated actions of the individual would explain more complex or interactive contexts (Vargas, 2024). Growing interest in extended cognition has entailed growing interest in individual-environment interactions. However, individual-environment interactions have been an acceptable focus of study only ‘[i]nsofar as this thesis requires only that one attend to the interactions of cognitive processes with noncognitive environmental processes’ (Adams & Aizawa, 2008, p. 79). The challenge of explaining how multiple agents might model each other’s models ad infinitum (e.g. recursive theory of mind) usually only appears as a curiosity ‘yet to be unpacked’ (Friston et al., 2012, p. 4).
Explaining antagonistic coupling mainly in terms of individual agents may reflect the broader commitment to explaining behaviour as the sum of independent component factors. Indeed, the foregoing suggestion that antagonistic coupling may depend on nonlinear interactions across scale verges beyond more conventional approaches to sport. Influential models of the ‘mental chronometry’ of processing stages in sport envision chain-like sequences (e.g. perceive-decide-execute) at a single timescale (e.g. Poplu et al., 2003; Ripoll et al., 1995). Such models hinge on assuming independent or decomposable timescales (i.e. ‘vertical separation’) separating, say, the cellular from organismic behaviour and both from longer timescales on which cultures develop games. Orthogonal to this vertical separation, decomposability of timescales gives permission to propose ‘loose horizontal couplings’, that is, the chain-like sequences operating at a single scale (Simon, 1977). Because the stages in horizontal processing-stage models reflect the contributions of pre-defined, independent components, systems operating in this fashion are characterised by component-dominant dynamics (Van Orden et al., 2003; Wallot & Kelty-Stephen, 2018). Fortunately, this assumption of decomposable scales and independent processes is not simply personal preference but generates empirically testable features which we could query in the behaviour in 1v1 competitions. Specifically, independent processes producing component-dominant dynamics should yield classical indicators of linearity: time-symmetry and additivity, built only of independent oscillations, producing often uncorrelated white noise (Kelty-Stephen et al., 2023; Kelty-Stephen & Wallot, 2017).
The possibility of nonlinear interactivity in antagonistic couplings opens the possibility that 1v1 player behaviour is no horizontal chain, but fluid coordination unfolding across many scales. As scale-dependent componential constraints loosen, a process may exhibit interaction-dominant dynamics (Van Orden et al., 2003) in which functions are soft-assembled or fluidly defined within an unfolding multiscale synergy (Turvey, 2007). In the case of 1v1s, fine-scale events include the action potentials and tendon stretching supporting muscle contractions. Medium-scale events might include synergistic muscular co-contractions and scanning eye movements, and longer-scale events might be decisions to dribble the ball right or left or to change tack. Coarser scales would include changes in possession, adjustments to strategy or formation, and even the glacial change of tactical trends (Balagué et al., 2019). Curiosity about and inclusivity of multiple scales may be necessary to avoid ‘scale-blindness’ in our theorising (McGann, 2024) and to prevent scale-dependent logic from trivialising adaptive behaviour (Mangalam et al., 2024). Interaction-dominant dynamics in behavioural data are consistent with the principle of nested organism-environment reciprocity endorsed by ecological psychology (Gibson, 2014; Wagman & Miller, 2003). This principle implicates a stance of ‘methodological direct perception’ in which the contributions of the individual and the surrounding context are ontologically entangled (Van Orden et al., 2003, 2010).
Nonlinear interactions across timescale manifest in our measurements as a power-law relationship between amount and frequency of behaviour. Behavioural science in sport has already begun unearthing these signatures of nonlinear interactions across timescale. For instance, Passos et al. (2009) showed that the fluctuations of 1v1 dyads exhibit power-law scaling. The power-law signature is crucial because the interactions in interaction-dominant systems involve cascades – fluid configurations which build multiplicatively across multiple scales of events (for a comprehensive treatment of cascades see Kelty-Stephen et al., 2013; Kelty-Stephen & Mangalam, 2022). So, invasion game 1v1s between two bipedal humans entail a hierarchy of constraints and multiple scales (Balagué et al., 2019). Gameplay produces fluctuations which flow through this multi-scaled hierarchy of constraints and engender a fractal structure in which small fluctuations are nested within larger fluctuations. The relationship between the magnitude (i.e. amount) and timescale (i.e. frequency) of fluctuation very often appears as a power-law (Kelty-Stephen et al., 2023; Mandelbrot, 1999). The scale-invariant structure of a power law might seem at first like a denial of the heterogeneity and nuance in antagonistic coupling. However, nonlinear cross-scale interactions create unevenness and heterogeneity in the form of multiple such power-law relationships. Each power-law relationship expresses a slightly different parametrisation of the scale-invariant relationship. The dyad is not a zero-sum exchange, and each player is leveraging ongoing variety in their own, and their opponent’s, behaviour to exploit emerging fluctuations and amplify opportunities. In this case, one power law will not suffice, and the variability of play – between or across a dyad – is best understood nonlinearly through a family of power laws.
Multifractal modelling of player and of dyad in the 1v1 format offers three major contributions toward clarifying how nonlinear interactivity in antagonistic coupling translates to the competitive outcome. First, it leans on previous insights (e.g. Passos et al., 2008, 2009) to find a place in the nonlinear-interactivity formalism to express individual differences in player behaviour across a dyad. Multifractal formalism allows player, or dyad, behaviour to have multiple power-laws which are estimable as part of a multifractal spectrum. The width and height of this spectrum reflect the number and strength of those power laws (Chhabra & Jensen, 1989; Ihlen, 2012; Kelty-Stephen et al., 2023). Specifically, wider multifractal spectra should correspond to greater exploratory variability, for example, as a defender tries to search out the most advantageous place to stop the attacker. Second, multifractal modelling allows us to quantitatively test the validity of the horizontal chain-like stage models of 1v1 play. Whereas single power laws or ‘monofractal’ results are suggestive of nonlinear interactivity, they have never been independently capable of distinguishing these interaction-dominant models from linear approximation. We can make this linear approximation using synthetic surrogate-data series representing the best linear model (i.e. iterated amplitude adjusted Fourier transform [IAAFT]; Schreiber & Schmitz, 1996). Comparing multifractal results between original measured movement and surrogate data can clearly demonstrate nonlinearity (Kelty-Stephen & Wallot, 2017; Mangalam et al., 2025).
Blending multifractal modelling with surrogate series allows a third contribution that could substantially strengthen nonlinear-dynamical models of performance. Beyond refuting linear chain-like models of behaviour, this original-vs.-surrogate difference in multifractal spectrum width allows expressing specifically how a player or dyad’s exploratory variability makes use of nonlinear interactions. This specific difference in multifractality from surrogates has been immensely useful in understanding movement control (Deligiannis et al., 2024; Mangalam et al., 2020). More to the point of 1v1 antagonistic coupling, this multifractal nonlinearity predicts perceptual-motor outcomes in learning how to respond to perturbation (e.g. Carver et al., 2017) even under uncertain circumstances (Kelty-Stephen, Lee, et al., 2023; Stephen & Dixon, 2011). Effectively, the growth and attrition of multifractal nonlinearity appears to correspond with the release and constraint of motoric degrees of freedom (Kelty-Stephen & Mangalam, 2024). Variations in multifractal nonlinearity reflect the unfolding of broader, more destabilised variability (i.e. with high multifractal nonlinearity) as well as the more focused, more certain constraint of variability (i.e. with lower multifractal nonlinearity (Bell et al., 2019; Booth et al., 2018; Jacobson et al., 2021; Kelty-Stephen, 2018; Kelty-Stephen et al., 2021, 2024). So, it is possible that multifractal nonlinearity could offer new leverage on modelling and predicting the outcomes of antagonistic coupling in 1v1 games.
Revisiting the format of a successful performance in 1v1 futsal might sharpen hypotheses about how and when multifractal dynamics predict successful task completion. First, the successful futsal attack begins with anticipatorily synchronising with the defender, that is, watching where the defender is moving and creating enough space to get there in advance. More generally, we can expect the defender to (temporally) follow the lead of the attacker. Hypothesis 1 was that classical metrics of anticipatory synchronisation (Rosenblum et al., 1996; Senthilkumar & Lakshmanan, 2007) would show that, between the 1v1 dyad, the attacker was leading the synchronisation and the defender was following. Next, following an ecological dynamics rationale (Araújo et al., 2006), Hypothesis 2 was that movements in a 1v1 task would exhibit interaction-dominant dynamics. In finer detail: Hypothesis 2a was the prediction of non-zero multifractal spectra widths, and Hypothesis 2b was the prediction of significant differences between the spectra widths of original data and the populations of surrogates. The presence of multifractal dynamics and nonlinear difference from surrogates was expected in both players as well as in a collective dyadic variable encoding interpersonal angle (Passos et al., 2009). These hypotheses follow neatly from wide evidence of multifractal nonlinearity in various demonstrations (Kelty-Stephen & Mangalam, 2024).
We also predicted that the attacker score could be associated with a divergence of multifractal nonlinearity between attacker and defender (Hypothesis 3). Without empirical precedent on the role of multifractal nonlinearity in 1v1 contexts, it was not clear whether successful attacking performance would be associated with higher or lower attacker multifractal nonlinearity, relative to defender multifractal nonlinearity. Thus, we entertained two exclusive alternatives regarding the role of this potential divergence in attacker score. In the first alternative, attackers would have higher scores when attackers showed more nonlinearity contributing to multifractality than defenders (Hypothesis 3a). This alternative would follow the heuristic understanding that more multifractality is healthy and potentially more robust than less or no multifractality (Ivanov et al., 2001). The second possibility is rooted in the foregoing description of the antagonistic coupling of a 1v1. That is, the successful attacker will destabilise the defender, encouraging wider variability in their defending movements before exploiting this instability by constraining their own behaviour into an attack. In this latter case, attackers might show less multifractal nonlinearity than defenders (Hypothesis 3b).
2. Method
2.1. Participants
Twenty male football and/or futsal players aged 18 to 38 years (M = 23.55, SD = 4.88) were recruited from student recreational sports activities on campus and by word of mouth to participate in this study on a voluntary basis. The study was approved by the Sheffield Hallam University ethics committee (ER47422746) with each participant providing informed consent before taking part in the study. The specific skill level of the participants was not a primary consideration of the study because the research question concerned the emergence of domain-general coordination dynamics. However, basic experience and an amateur level of performance were required the control of the ball within the capture volume. This requirement was reflected in a selection criterion of three or more years of self-reported experience playing football (M = 11.05, SD = 4.73) or futsal (M = 1.05, SD = 1.23) at any level.
2.2. Design
A 1v1 ‘futsal’ (i.e. an adaptation of football played on a hard surface) task met a number of practical constraints. 1v1 attacker-defender dyads exemplify antagonistic coupling (De Poel, 2016): the attacker attracts the defender, while the defender repels the attacker. The task engaged the experiential qualities of 1v1s in competitive matches, and the futsal format enabled the use of a high-precision motion capture system. All participants completed at least one match (i.e. two trials: one attacking, one defending) and no participants completed more than 12 trials (M = 5.3, SD = 3.06). Pairings were assigned randomly, with no pairings repeated across the data collection, so all dyads were unique.
2.3. Procedure
Player movements were recorded by a motion capture system (Cortex 6.0, Santa Rosa, CA) tracking the trajectory of a sacral marker (120Hz). All participants wore tight-fitting Lycra shorts to ensure consistent and stable fixture of a sacral marker. Video footage of all trials via a video camera (Panasonic HC-V770, Newark, NJ) allowed confirmation of match scores and maintenance of play within capture volume. Sacral-marker position along a transverse axis between target zones would effectively encode task engagement. As explained below, reaching and occupying the target zones with their foot or the ball entailed players moving laterally to fulfil their roles.
The experimenter read task rules and objectives aloud to all participants prior to their first trial. White tape demarcated playing area on the floor. Matches were played with official size and mass Nike Pro futsal balls. Attacking required manipulating a futsal ball into either of two 60 × 60 cm targets marked with white tape 200 cm apart on the floor (see Figure 1(A)). Defending required participants to occupy target zones by maintaining foot contact with the target zone (see Figure 1(C)). Scoring a point involved meeting three conditions simultaneously. First, the attacker had to have foot contact with the ball. Second, the ball contacted the target zone. Third, the defender’s foot had no contact with the target zone (see Figure 1(B)). Attackers could score consecutive points in the same zone if the ball fully exited and then re-entered the zone. The experimenter asked each participant to stay on their side of the white-tape dashed line connecting target zones to avoid physically obstructing their opponent’s movements. The experimenter also asked participants to stay within the larger marked area denoting the motion-capture capture volume. All participants completed an unscored 15-second warm-up in each role, that is, attacker and defender prior to their initial trial. Each match lasted 60 seconds marked by an audible tone synchronised with the start and end of recording. Participants switched attacker/defender roles after 2–4-minute rest periods. Depiction of (A) the basic experimental setup, (B) an attacker scoring a point by controlling the ball in a target zone while the defender is not in contact with the zone, and (C), a defender preventing the attacker from scoring a point by placing their foot in the target zone before the attacker controls the ball in the zone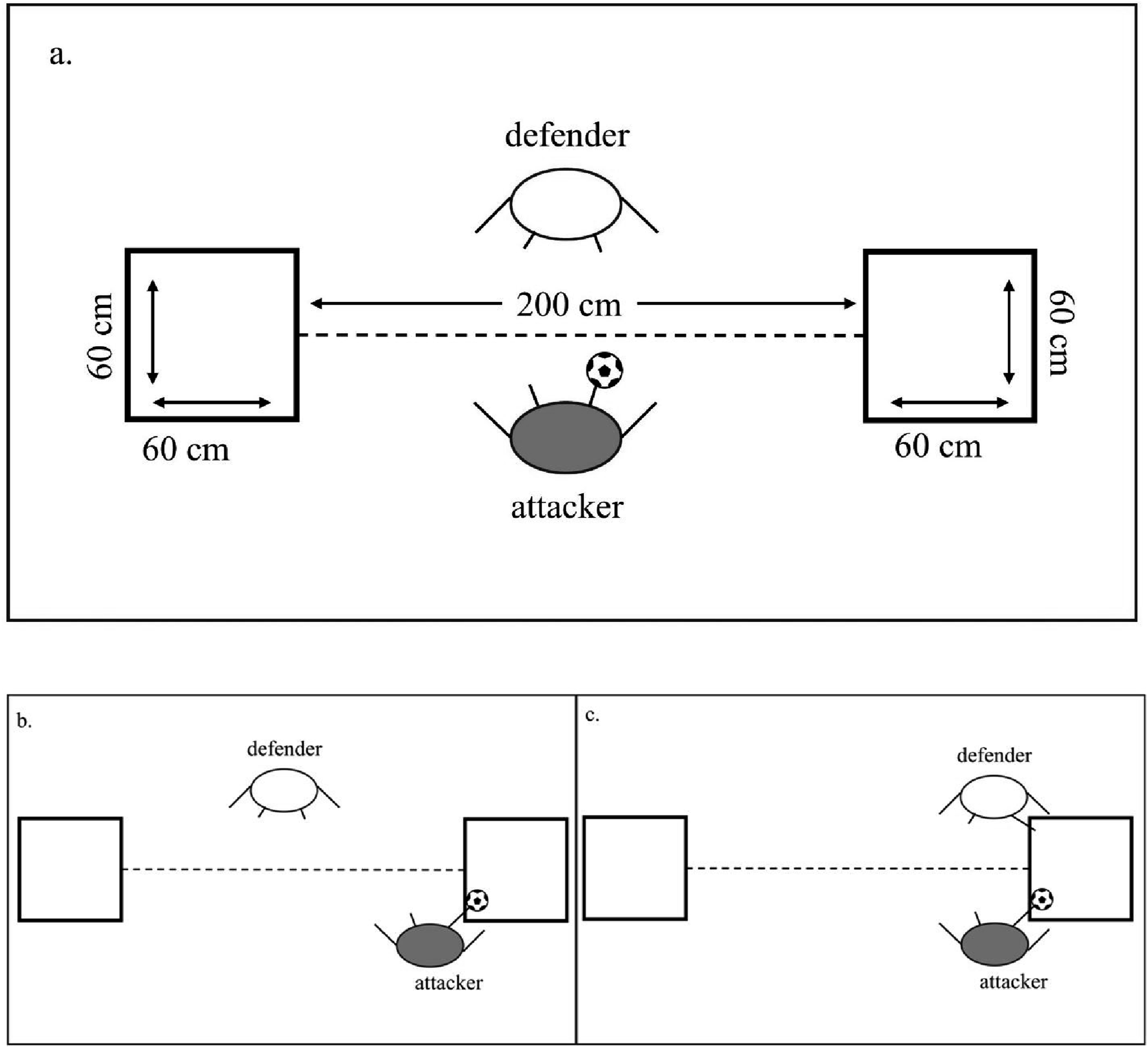
2.4. Data Processing and Variable Calculation
twenty-eight matches (56 trials) were recorded. After excluding six matches due to marker dropout, data analysis addressed sacral-marker trajectories along an axis connecting the two target zones (i.e. the dashed line in Figure 1).
An interpersonal angle was defined by the line-of-sight between attacker and defender and the goal line (see Figure 2; Passos et al., 2009). This measure encodes variation in coupling, where 90° would entail face-to-face mirroring between defender and attacker. Above and below 90° entails the attacker to the left and right, respectively, of the defender. As the 1v1 experimental task constraints required the defender and attacker to stabilise and destabilise the dyad, respectively, fluctuations in the interpersonal angle related to key task objectives. Interpersonal angle was preferred to other potential collective variables such as interpersonal distance and relative phase because the experimental task constraints created primarily lateral movement towards and away from target zones, and the anticipating synchronisation metric (see section 2.5.1) already addressed the temporal coordination of the dyad. Hence, interpersonal angle was selected to capture important aspects of the dyad’s co-adaptation over time. Data capture process for the interpersonal angle θ for a representative trial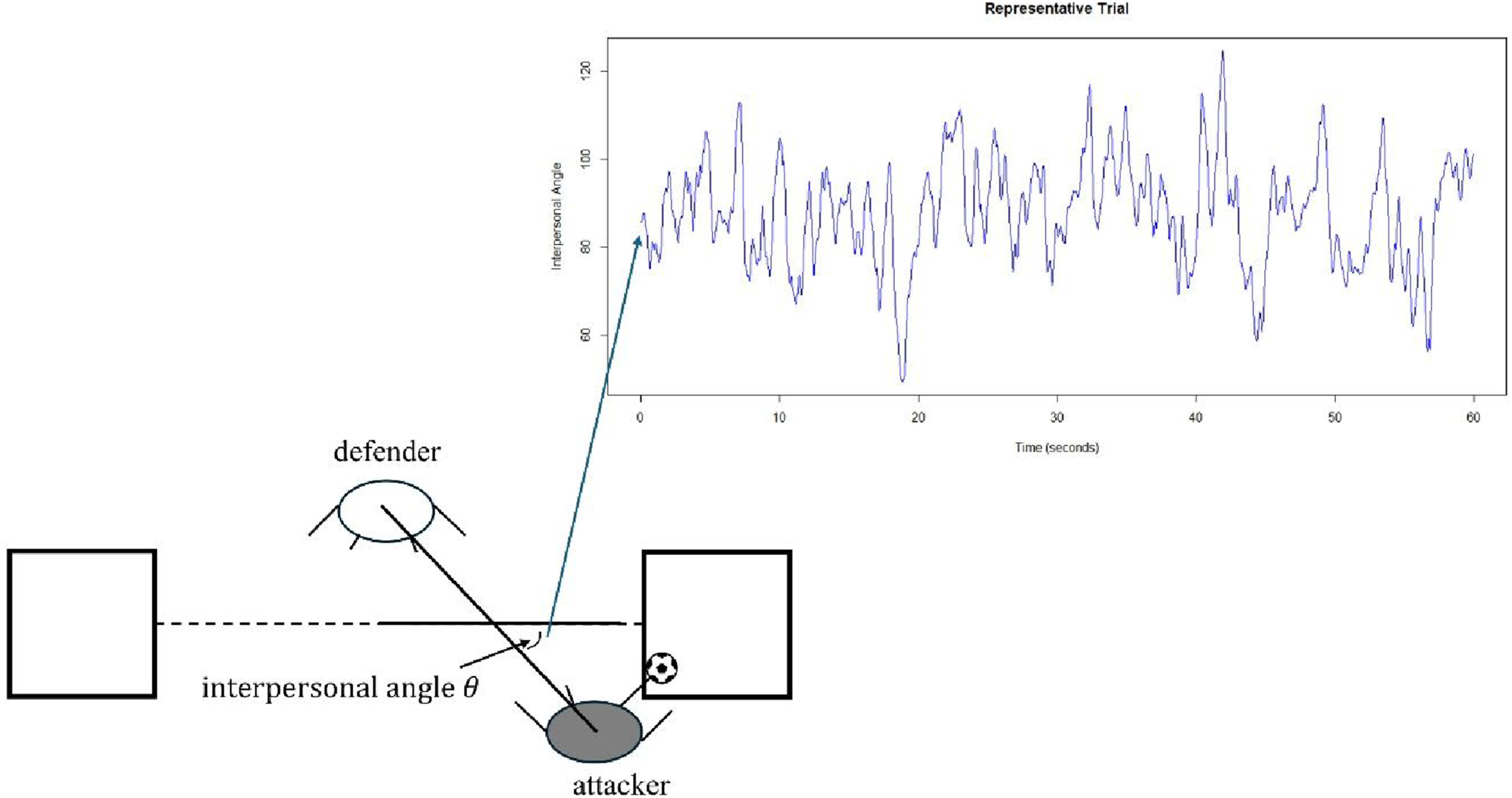
2.5. Data Analysis
All statistical analyses were conducted in R-Studio (Posit Team, 2022; R Core Team, 2021b). To confirm and assess the anticipatory synchronisation of attacker with defender movements, we used Senthilkumar and Lakshmanan’s (2007) measure of intermittent anticipating synchronisation (see supplemental materials), based on previous work by Rosenblum et al. (1996). Multifractal detrended fluctuation analysis (MFDFA; Kantelhardt et al., 2002) was conducted and iterative amplitude adjusted Fourier-transform (IAAFT; Schreiber & Schmitz, 1996) surrogates were produced with the package ‘multifractal’ (Schadner, 2023). We used an ANOVA (R Core Team, 2021a) to address basic differences in multifractal spectra before fitting a regression model for attacker score. The regression model was fit with the package ‘glmmTMB’ using the function ‘glmmTMB’ with family set to ‘nbinom2’ (Brooks et al., 2017). Conceptual outlines of these analyses appear in the subsequent paragraphs.
2.5.1. Anticipatory Synchronisation for Testing Hypothesis 1
The similarity function
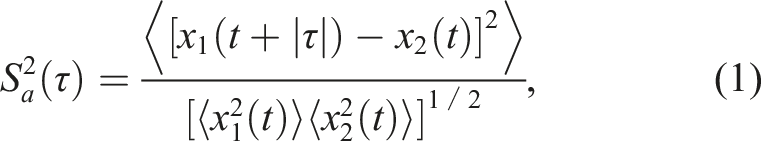
This function assessed the average subtractive difference between the sacral-marker lateral-position series for the attacker x
1
and for the defender x
2
. The value of τ that minimises Plot of the similarity function 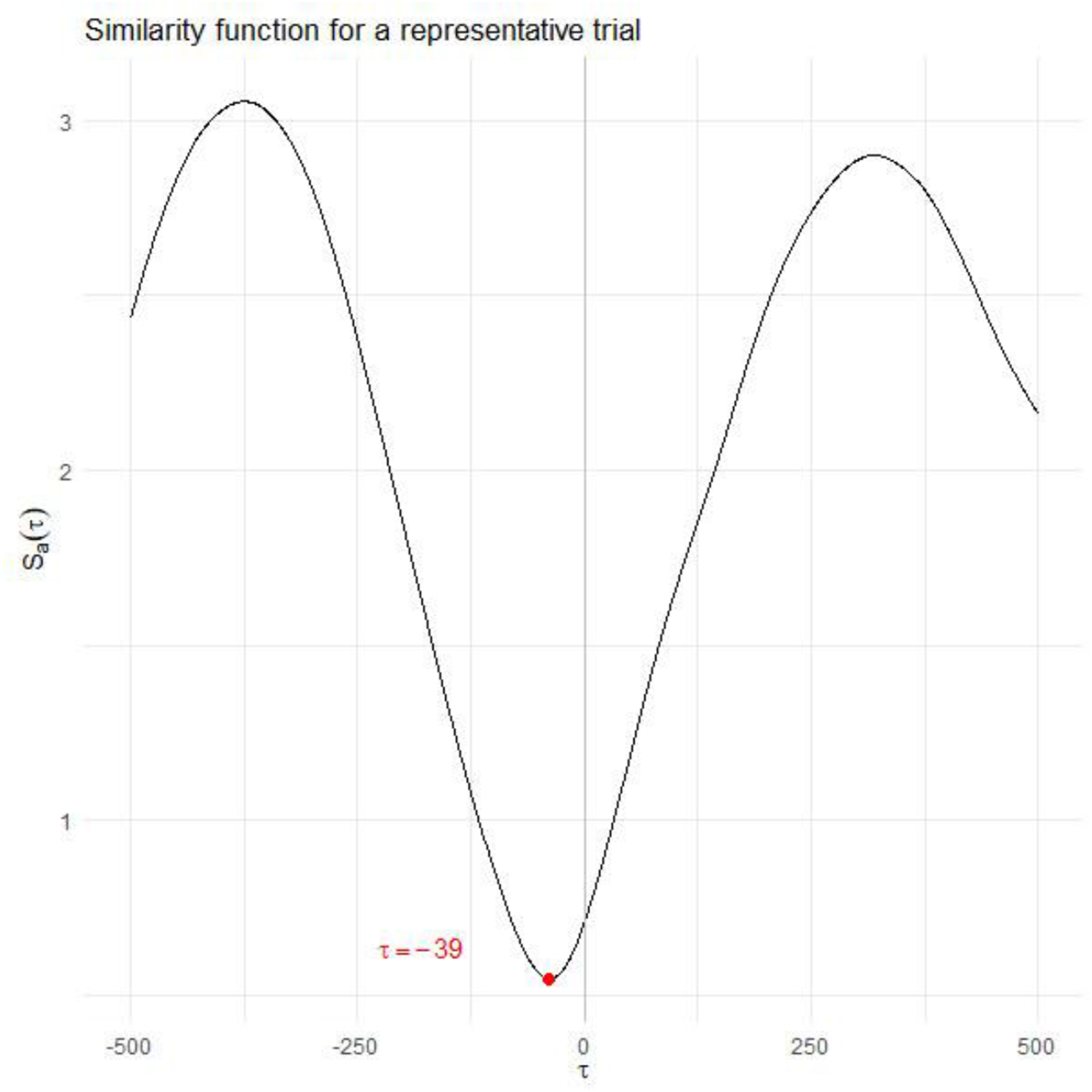
2.5.2. Multifractal Analysis for Testing Hypothesis 2a
For each trial, MFDFA estimated multifractal-spectrum width for three raw time series, that is, attacker and defender’s sacral-marker lateral movement series (Figure 4) and the interpersonal angle series (Figure 2). MFDFA quantifies the scaling behaviour of fluctuations in nonstationary time series (Kantelhardt et al., 2002). Standard, non-multifractal DFA divides the time series into segments, and, within each segment, it applies polynomial detrending and computes the residual standard deviation. The slope of the relationship between residual standard deviation F(s) and segment size s on double-logarithmic plots is an estimate of the Hurst exponent h. DFA’s use of the second moment (i.e. standard deviation) hinges on the use of the exponent 2. For instance, standard deviation involving first squaring of difference scores and then taking the square root of their sum. MFDFA generalises this approach by explicitly manipulating moment as a parameter q. Whereas non-multifractal DFA quantifies a single power law using the second moment (i.e. standard deviation; q = 2), MFDFA generalises standard deviation F(s) from its canonical q = 2 to a continuum qth-order standard deviations, that is standard deviations F (q, s) reflecting the raising of difference scores to the qth power and taking the qth root of their sum Data capture process for the position of the sacral marker of an attacker and defender in a representative trial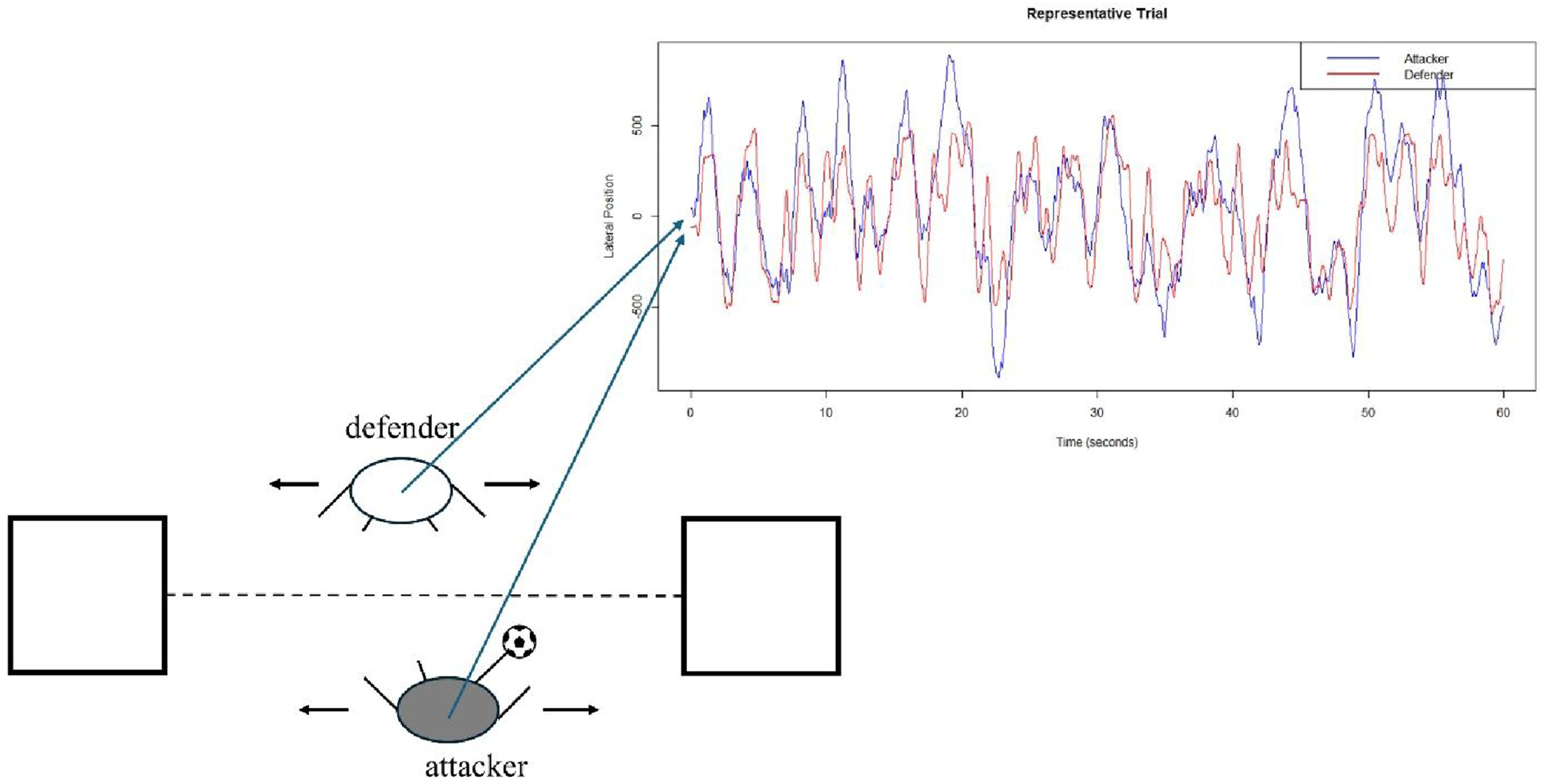
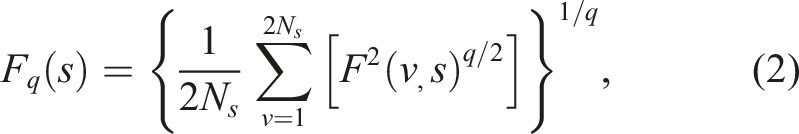
Positive values of q emphasise large-amplitude fluctuations while negative values of q emphasise small-amplitude fluctuations. Consequently, MFDFA uses the slope of the relationship F (q, s) and s as an estimate of The multifractal spectrum α(q), f (α(q)) representing the range of power-law exponents α(q) for different qth-order moments and the Hausdorff dimension f (α(q)) of the subset of the time series characterised by α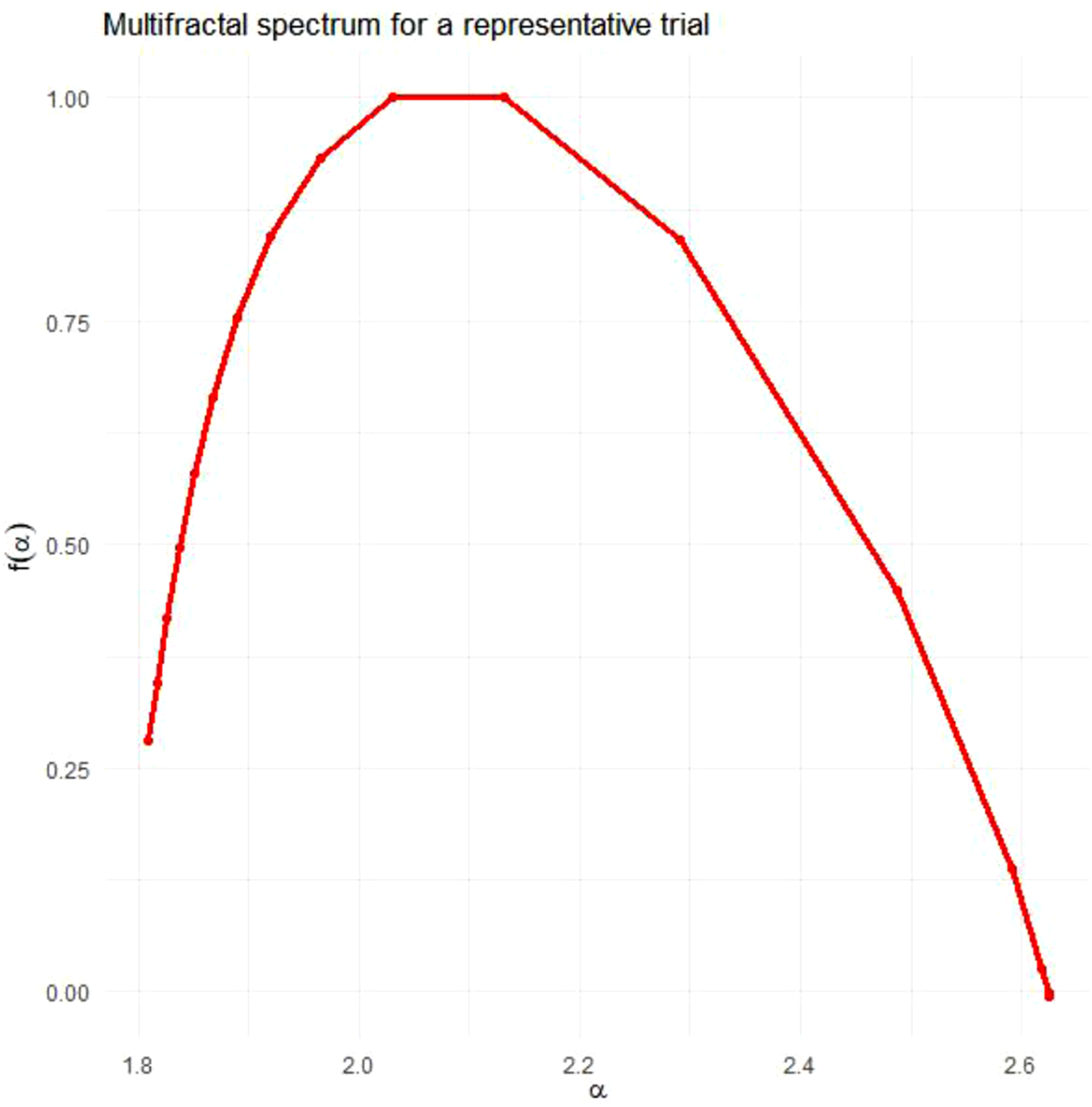
MFDFA requires selecting three primary parameters: range of scales (i.e. minimum and maximum segment sizes), polynomial order of detrending, and range of q moments. Minimum scale must exceed polynomial order of detrending to prevent overfitting. Maximum scale is typically N/4 or N/8 where N is the time-series length. Present analysis used scale range of 24 to 899 (N/8) and quadratic detrending to avoid both small-sample bias at larger scales and overfitting at short scales, respectively (Ihlen, 2012). We decided q-range on trial-by-trial basis beginning with −10 ≤ q ≤ 10 and only used q values for which the fluctuation function on double-logarithmic axes had
2.5.3. Surrogate Analysis for Testing Hypothesis 2b
IAAFT surrogate analysis involves preserving the linear features (mean, variance, autocorrelation) of the data while shuffling the temporal order and thus removing the interactions between scales (Schreiber & Schmitz, 1996). The multifractal spectra width is computed for each of a sample of surrogates (see Figure 6). A one-sample t-test (henceforth tMF) quantifies the difference in multifractality due to nonlinearity absent from the linear structure embodied by the surrogates. This multifractal nonlinearity statistic tMF is a robust estimator of nonlinear interactivity across scales (Kelty-Stephen et al., 2023; Mangalam et al., 2025). Multifractal nonlinearity tMF also serves as significant predictor in regression models of adaptive behaviour across a wide range of perceptual, cognitive, and motoric domains (Kelty-Stephen & Mangalam, 2024). For each trial, we generated 32 IAAFT surrogates for the attacker lateral-position series, defender lateral-position series, and interpersonal angle series (e.g. Figures 2 and 4) and computed tMF for each. As the task objectives encouraged participants to explore primarily lateral movements, significant differences between IAAFT surrogates and original time series would justify going beyond oscillation-based forms of analysis. Including both multifractal analysis and the measure of anticipating synchronisation (τAS) enabled analysis to address both multiscale coordination and oscillatory synchronisation, respectively. Multifractal spectra for original time series (red) and 32 IAAFT surrogate time series (grey) for a representative trial. Note. Original and IAAFT surrogate time series were found to exhibit a similar range of Hurst exponents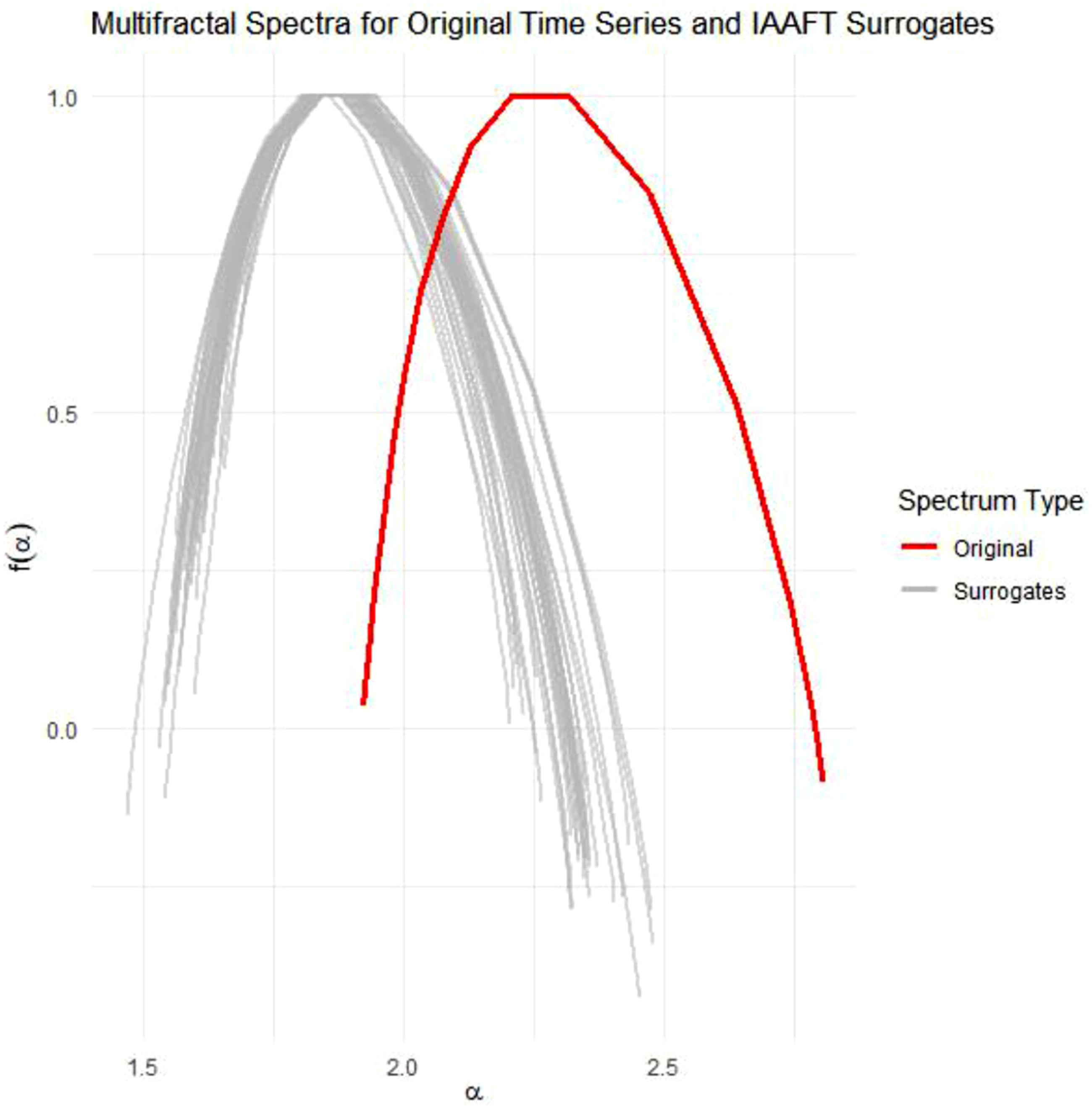
2.5.4. Generalised Linear Mixed-Effects Model to Test Hypothesis 3
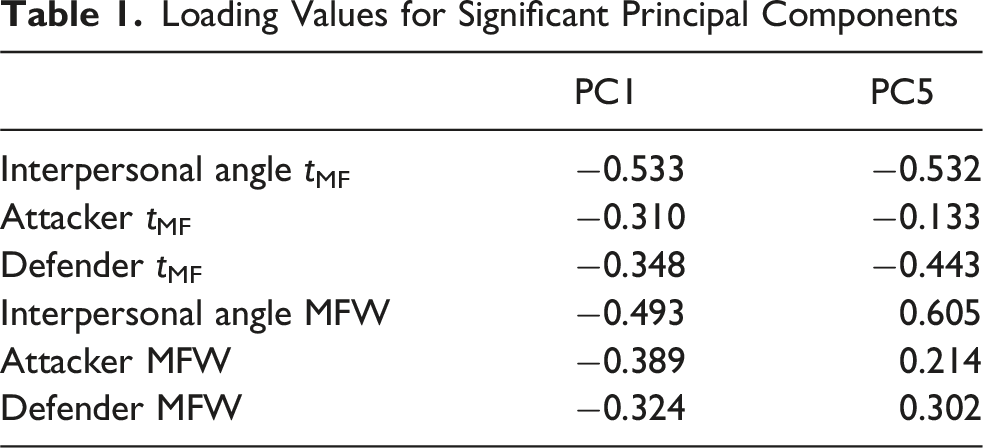
Loading Values for Significant Principal Components
Negative Binomial Mixed-Effects Model for Attacker Scores
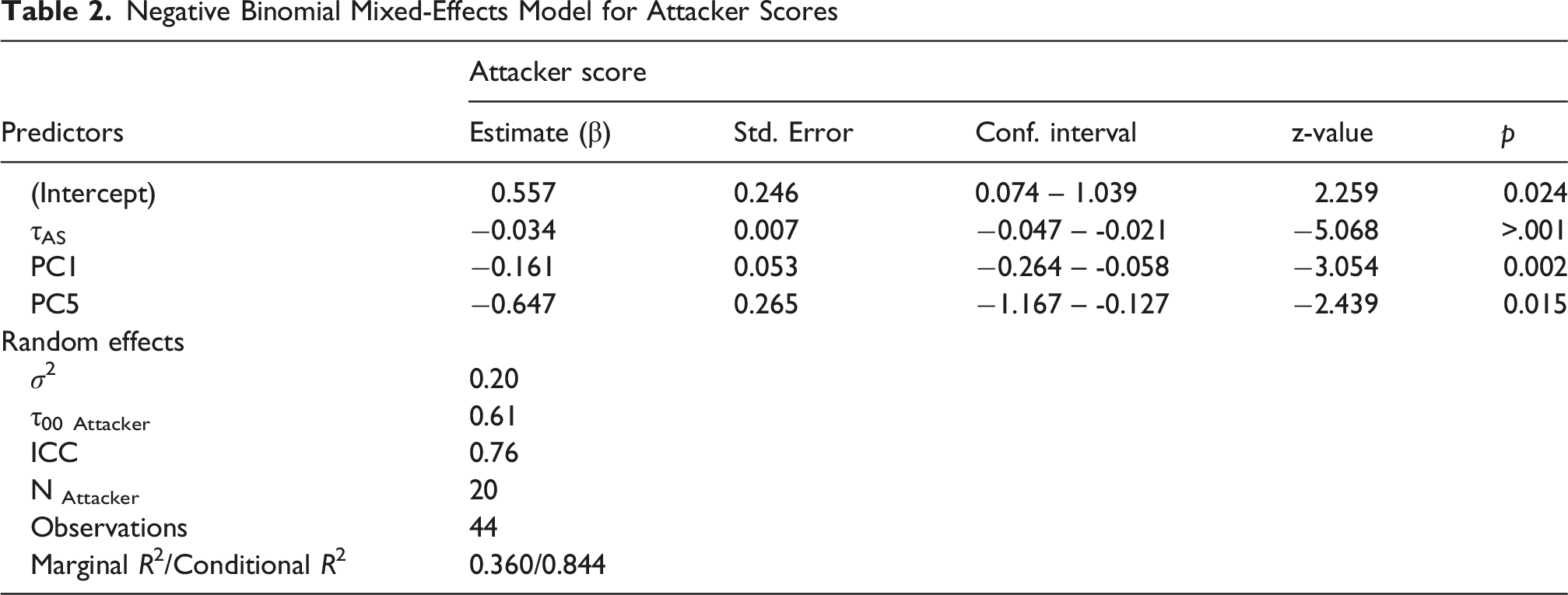
Mixed effect modelling is a departure from standard ANOVA treatments of our dependent measures. Whereas ANOVA estimates fixed effects for balanced designs using ordinary least-squared modelling, mixed-effect modelling uses maximum likelihood to estimate fixed effects for covariates while nesting response behaviours and covariate values by participant even for unbalanced designs. We included an ANOVA to detail the group differences in multifractal spectra before presenting the regression model.
3. Results
3.1. Hypothesis 1: Attackers Lead Anticipating Synchronisation
The analysis of anticipating synchronisation showed that attackers’ movements tended to lead or anticipate defenders’ movements, based on the values of τ that minimised the similarity function
3.2. Hypothesis 2a: Non-Zero Multifractal Spectra Widths
All roles in all trials exhibited non-zero multifractal spectra widths (attackers M = 0.869, SD = 0.312; defenders M = 1.151, SD = 0.419; interpersonal angles M = 0.851, SD = 0.337). Non-zero multifractal spectra widths entail the presence of a range of power-laws at work in contrast to the single power-law exhibited by monofractal processes.
3.2.1. Hypothesis 2b: Significant Differences Between Original and Surrogate Spectra Widths
IAAFT surrogate analysis revealed MFW for the original data differed significantly from MFW of 32 linear surrogates in all trials and roles
3.3. Hypothesis 3: Diverging Multifractality Relating to Attacker Score
3.3.1. Analysis of Variance
Prior to testing more specific hypotheses about the role of attacker and defender multifractality in attacker score, we utilised one-way Analysis of Variance (ANOVA) to compare spectra widths and multifractal nonlinearity in attackers, defenders, and the collective variable, interpersonal angle. The results indicated a significant effect of role on spectrum width F (2, 137) = 10.52, p < .001 (see Figure 7). The effect size ( Violin plot comparing multifractal spectra widths for attackers, defenders, and interpersonal angles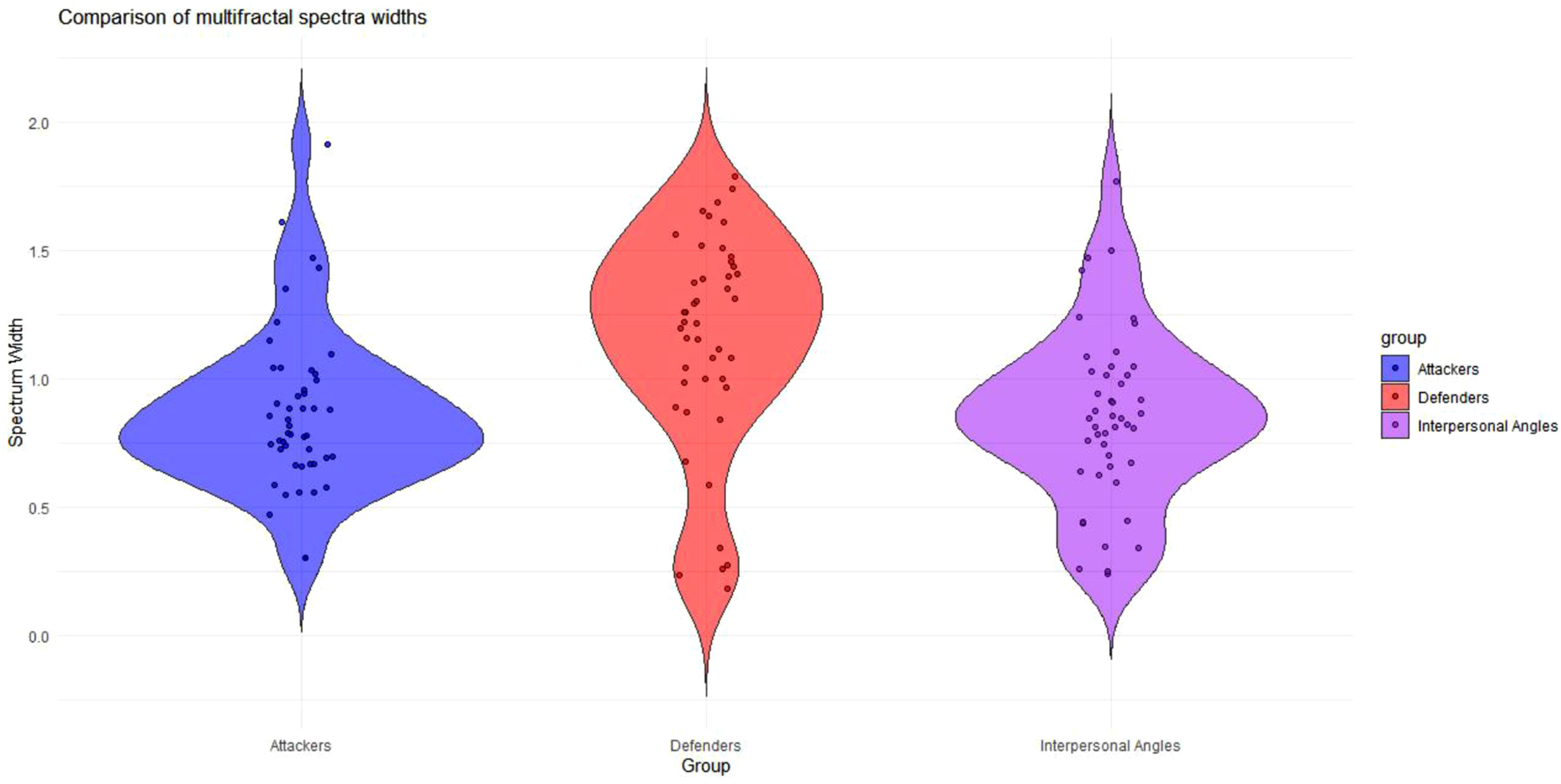
Multifractal nonlinearity ( Violin plot comparing multifractal nonlinearity (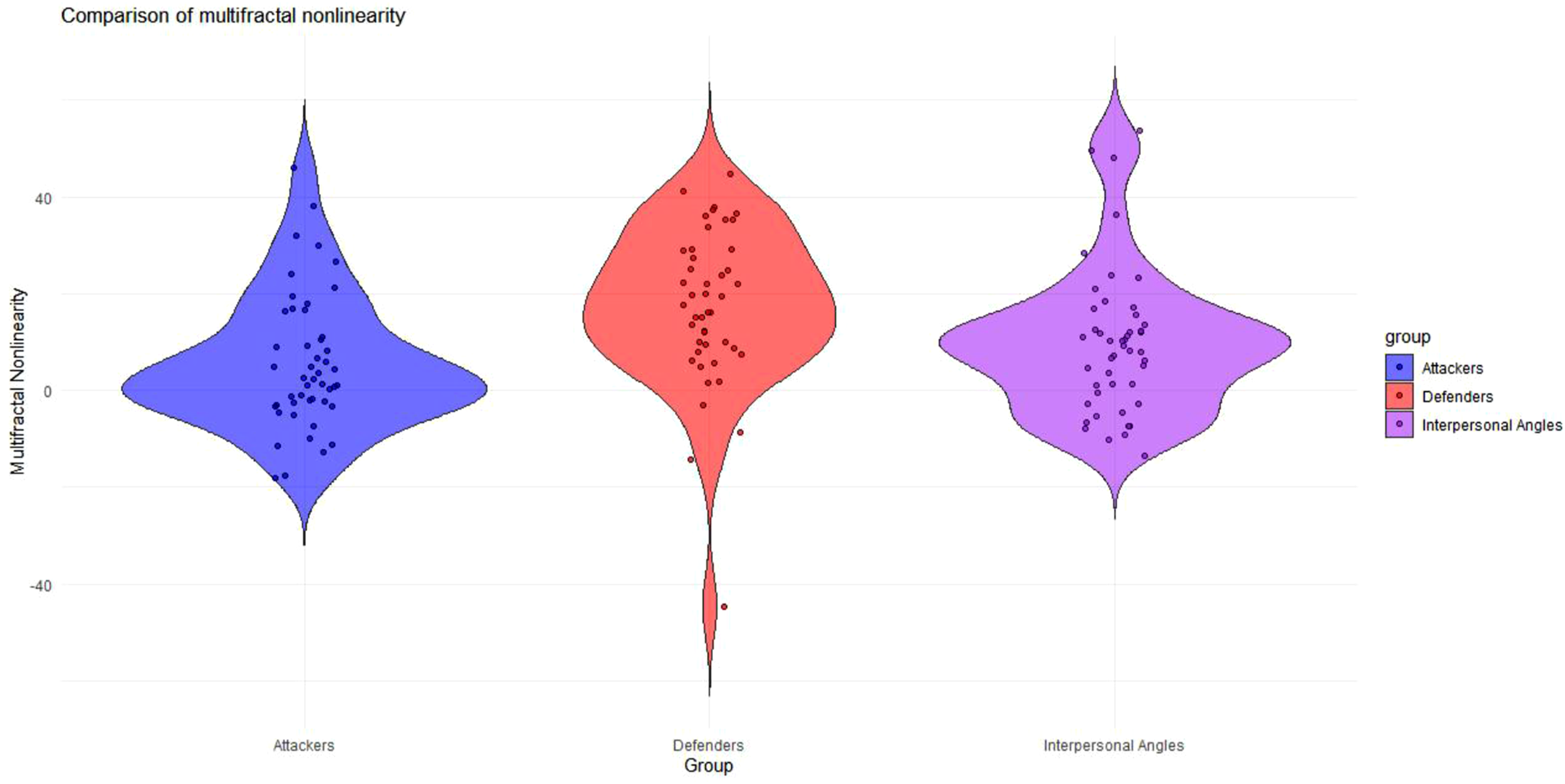
3.3.2 Hypothesis 3a and 3b: Multifractality and Attacker Score
The generalised linear mixed-effects model revealed main effects of anticipating synchronisation (τ), global multifractality (PC1), and the contrast between MFW and
4. Discussion
The present study tested three primary hypotheses. The results supported the predictions of all three. First, attackers exhibited temporally anticipating synchronisation with defenders (Hypothesis 1). Second, all trials for all players and dyads exhibited non-zero multifractal spectra widths (Hypothesis 2a) and significant multifractal nonlinearity (Hypothesis 2b), confirming the presence of interaction-dominant dynamics. Two exploratory hypotheses considered how multifractal nonlinearity would be associated with attacker score. Results failed to support the possibility that attacker score would be associated with greater attacker multifractality than defender or dyad multifractality (Hypothesis 3a). Instead, results supported the alternative hypothesis, that is, that attackers scored more points when they fostered increased multifractal nonlinearity in the defender and interpersonal angle (Hypothesis 3b). The present findings develop on work by Passos et al. (2009) which found power-law distributions in the interpersonal angle values in antagonistically coupled 1v1 dyads while adding confirmation of nonlinear interactions between scales (i.e. multifractal nonlinearity) and modelling its role in attacker scores. Passos et al. (2009) interpreted power-law distributions in fluctuations in 1v1s as a form of self-organised criticality (SOC) (Bak, 1996) which arises when multiple agents co-adapt to ‘the edge of chaos’ (Kauffman & Johnsen, 1991). The present results confirm the presence of power-law behaviour in the fluctuations of 1v1s and lend support for the general interpretation of self-organisation under nested constraints. However, the role of multifractal nonlinearity
These results suggest sport performance may rely on a lawful fluidity allowing players to act and respond on multiple time scales in a way that carries the events at one scale into adjustments and corrections at another. Classically, scholars have attempted to model sport as unfolding at two distinct levels: a local space of immediate interactions governed by superficial action-perception processes and a global ‘form space’ of rules and tactics governed by representational knowledge structures (Ripoll, 2009). Such claims fall back on an older tradition of cognitive psychology that presumes a mental ‘knowledge level’ housing and operating upon mental representations about events unfolding on a physical level of sensorimotor process (Newell, 1982). In such a framework, the antagonistically coupled dyad becomes a drama of duelling representational systems each trying to predict and outwit the other. Independence appears both within each player, separating knowledge from sensorimotor levels, and between each player, with each intentionally misleading the other. Consequently, from this perspective, it has made sense to address skill development in 1v1 contexts strictly through the movement patterns of the individual (Gearing & Bridge, 2024; Parry et al., 2025).
The present results show that there is more that warrants modelling in dyadic antagonistic coupling than the representational account can address. Understanding attacker success requires reckoning with multiple spatial (i.e. individual and dyadic) and temporal scales that overlap and thus prevent such clean fracturing of the antagonistically coupled dyad. In a 1v1, an individual’s intention to successfully attack or defend refers to an outcome at the level of the dyad. To underscore the challenge in barer form: the player must respond to their own body and the dyad at the same time. For example, when an attacker A intends to evade a defender D, A’s actions must refer to multiple scales of events, not just their current bodily state but to the dyadic set {A, D}. The dyad is a coordinative structure that becomes potentially another synergy in which A’s movement system must participate. Crucially, the dyad constitutes another scale beyond A’s body in which A must achieve an outcome O{A, D}. Hence, the dyadic-level definition of the goal must include an entailment that has the logical structure A ({A, D}) →O{A, D}. Mathematical theory has attempted resolve how to define independent members of a set that act according to the set they belong to. A century later, this program appears to be logically impossible (George, 1987; Russell & Whitehead, 1910; Turvey, 2018). Atomic components cannot approximate a distributed fluid process in which a single event has forking entailments that diverge across scales (Dewey, 1896; Gibson, 1960; Mandelbrot, 1999; Van Orden et al., 2010). The present evidence of cross-scale interactivity in 1v1 dyads suggests that this issue is more than a logical abstraction.
By contrast, advancing Passos et al.’s (2009) work to go beyond SOC toward the broader class of cascades, fluid dynamics affords a means of articulating lawful regularities in a way that avoids scale-bound approximations (Buckingham, 1914; Mangalam et al., 2024; Weyl, 2015). The lawful fluidity of the cascades provides a ready link between scales spanning both the individual and the dyad. Cascade-like interplay between laws and constraints governing and shaping flow, respectively, relaxes the need for a local, centralised point of control and suggests that intentionality may be expressed through an ecology of distributed constraints, rather than chain-like efficient causes (Juarrero, 1999; Kloos & Van Orden, 2010; Turvey, 2007). Eminent movement scientist, Nikolai Bernstein, observed that movement was better analysed as an integrated meshwork rather than a chain of independent details (see Bernstein, 1967, p. 69). The fluid adaptivity of movement manifests in intermittent, sweeping reorganisations stemming from non-obvious, non-local causes.
Beyond the intuition that discretised, piecewise analyses fail to capture this form of system (re)organisation, multifractal cascade modelling provides a robust contemporary framework for the quantitative analysis of the nonlinear structures constraining and releasing degrees of freedom in dexterous behaviour (Kelty-Stephen & Mangalam, 2022). The current results build on previous research showing that multifractal cascades may be the mechanism which enables attunement to this lawful, global structure and supports anticipatory coordination (Stephen & Dixon, 2011; Stephen et al., 2008). Rather than ‘weak’ contingence on private predictive models (e.g. Ripoll, 2009; Rosen, 2012) this anticipation would be ‘strong’ (Dubois, 2003). It would ground the prospectivity of adaptive behaviour in both the lawful invariant ecological structure and the exploratory behaviour which supports the discovery of this structure (Arzamarski et al., 2010; Mangalam et al, 2024; Stephen et al., 2012; Turvey, 2018).
Nested in the ecology of a sport dyad, the struggle between attacker and defender unfolds as a competition for affordances, that is, the possibilities for action (Gibson, 2014; Mangalam et al., 2024). For instance, the distance between targets in the present 1v1 task presses upon the classical affordance of reachability (Carello et al., 1989; Choi & Mark, 2004), a tradition which might cast the reachability affordance in 1v1 as some ratio of leg length to inter-target distance (e.g. Warren, 1984). However, the 1v1 task objective nested reachability within the added constraints posed by antagonistic coupling. The classical ratio of bodily to context lengths thus accrue a dizzying number of time-varying denominators. To score a point, an attacker needed to perceive where they could contact the target with foot on ball when the defender could not reach the target first. So, it is not just reachability but a murkier ‘reach-first-ability’ affordance which is highly specific to ebbing and flowing task constraints of 1v1 contexts in competitive sports.
The crucial role of anticipating synchronisation in attacker success suggests that this temporal relationship is a primary concern for both players. The ‘reach-first’ affordance is not simply the nesting of the body within a static task. Reaching first abuts and indeed conflicts with the defender’s leg length: a scoring opportunity is equally a defending inopportunity. Another denominator looms as attacker or defender’s previous movements constrain their future possibilities. The classical sense of affordance-as-single-ratio falters as the nesting relationships change the constraints on reachability. Furthermore, these time-varying constraints on reachability likely only compound with other affordances like controllability of the ball. The attacker’s search for a strategic marginal difference beyond the defender’s affordances are most likely nested sequences unfolding amidst the ebbing and flowing constraints rather than discrete movements. The non-local nature of the nested sequences involved in the generation of reach-first affordances also makes it difficult to explain the findings in terms of internal states or predictive models. Internal predictive explanations of performance would also need to contend with the non-ergodic nature of the task implied by the significant multifractal nonlinearity in each trial (see Mangalam et al., 2023).
Multifractality offers new traction on understanding 1v1 because it addresses the nested complexity of affordances which support ‘strong’ anticipating synchronisation in competition. With fluid accumulation of constraints, the affordance as a body-context ratio quickly yields to a cascade of successive divisions – that is, multiplicative relationships – among constraints at multiple scales. If 1v1 coupling complicates the single body-context ratio sense of affordance, it may be high time to consider multifractality as means to formalise the affordance relationships supporting competitive coordination in sport. Multifractality of head movement has been shown to increase near the action boundaries in judgements of the stand-on-ability of slopes (Hajnal et al., 2018). So, multifractality may offer a generic means to model the movement variability allowing players to explore the nesting relationships supporting affordance perception. Multifractal-spectrum width and multifractal nonlinearity are ways of indexing how much and why, respectively, the fluid meshwork of the Bernsteinian movement system releases its degrees of freedom. Attackers may thrive by cultivating proximity to the defender’s action boundaries, provoking defenders to widen their exploratory variability to maintain poise. Deceptive moves executed near the boundary of the defender’s affordance to reach a target may generate uncertainty and support the attacker’s objective of destabilising the dyad. Near a defender’s affordance boundaries, the ‘leverage’ of an attacker’s movements to destabilise the system is amplified nonlinearly. In these regions, a subtle asymmetry may generate nested ‘counter-affordances’ as a defender commits momentum to recover. One v Ones may require attackers to narrow and, therefore, direct nonlinearity in movement towards the marginal excess of attacking affordances beyond a defender’s affordances. The relative narrowing of attacker’s nonlinear multifractal variability may reflect a transition from exploring for this marginal excess to zeroing in on and exploiting it. Notably, achieving this aim required the attacker to act with respect to the dyad and cultivate multifractal nonlinearity in the interpersonal angle.
Several implications for sports practitioners can be gleaned from the interaction-dominant dynamics of antagonistically-coupled dyads. First, the role of nested affordances in competitive coordination suggests that the design of training activities which ‘represent’ the performance environment should consider not only the spatial layout of the practice area, but also how the specific constraints of a task shape its temporal structure. Activities constrained to reset after the execution of discrete movement patterns may not expose athletes to the nesting structures present in more extensive competitive interactions, crucial for the development of ‘strong anticipation’. Second, practitioners can observe players’ behaviour (e.g. approach, avoidance, hesitancy, decisiveness) around co-determined affordance boundaries to evaluate awareness, fluidity, rigidity, and perhaps even confidence in the competitive context.
5. Limitations and Directions for Future Research
The present work represents a first step towards understanding the dynamical flows of antagonistic couplings in invasion games. We were able to measure and model an extended period of antagonistic coupling with precision. However, this precision came at the cost of limiting play to a single axis in 1v1 exchanges divorced from any team coordination and from any broader match context. Our very rhetoric about the force of interactions across scales entails that future research should be ready to extend the present modelling to contexts of game play across broader scales. That is, future research might investigate the 1v1 dynamic between specific pairs of players within the context of a full match, potentially examining how specific dyads develop their antagonistic coupling with successive encounters. Future research including ball tracking could also investigate progressively higher-order collective variables encoding player-ball-goal angles (e.g. Carrilho et al., 2020).
Future work might also address a more distributed network of coupled agents. This distributed-network approach could expand beyond 1v1 to whole-team dynamics, for example, towards a multifractal portrayal of a ‘group tensegrity’ (Caldeira et al., 2020). This network approach might also focus inward to within-player dynamics. Longer-term data collections across full-body marker sets allow the modelling the flow of fractal-fluctuations across the body (Carver et al., 2017; Mangalam et al., 2020), reflecting body-wide tensegrity organisation (e.g. Turvey & Fonseca, 2014). The present limited data collection could only find that local Hurst exponent values for attackers, defenders, and interpersonal angles appeared to covary closely within trials. Similar network modelling has shown that multifractality helps to classify social coordination across groups or swarms of organisms as attractive or repulsive for resources (Carver & Kelty-Stephen, 2017; Dixon & Kelty-Stephen, 2012). The nonlinear interactions across scale may entail consonant cascades both within and among organisms in social interactions. Multifractality offers a potentially common framework for understanding coordination within and among organisms (Kelty-Stephen, 2017). Sport offers a diverse array of progressively more complex circumstances and coordinative modes for understanding pursuit-evasion interactions.
The present work was exploratory of one sport-specific pursuit-evasion interaction, but experimental manipulation of the affordance layout would generalise this work towards wider ecological frameworks for pursuit-evasion interactions. Specifically, experimental manipulation of inter-target distances as ratios of attacker’s or defender’s leg length could clarify the role of reachability. Explicit manipulation of the affordances could reveal how attackers might cultivate proximity to the defender’s action boundaries. Crucially, attackers might be exploiting the pursuing defenders’ action boundaries to encourage more exploratory variability. Changes in the affordance layout might change the evading attacker’s capacity to fake out the pursuing defender. For instance, the constraints of the present task shaped the competitive interaction unfolding primarily between to two target zones with near-constant proximity to co-determined affordance boundaries. The addition of more attackers, defenders, and targets would create more complex nesting and branching in the affordance layout and demand new forms of exploratory variability in performance. If indeed multifractal modelling can formalise affordance relationships, then experimental manipulations might influence the multifractal results and their relationships with antagonistic outcomes.
Multifractal modelling could also enable empirical exploration of the claim that sports emerge as sets of replicable task constraints that players select for their ability to foster enjoyable experiences (Farrokh et al., 2024, 2025). Multifractality has already demonstrated leverage into the dynamics of experience in computer games (Bennett et al., 2022; Dotov et al., 2017), and we anticipate that similar analyses could help inform the constraint selection process in competitive contexts such as sports. Future work might aim at optimal or ‘flow’ experiences (Csikszentmihalyi, 1990) more precisely by triangulating between constraint manipulation, flow measures, and cascade modelling. Flow experiences might reflect precisely those fluid aspects of cognitive process that cascade dynamics are best suited to predict. Multifractal analysis might show flow and cascades to be more than a coincidental pun.
6. Conclusion
Multifractal modelling of 1v1s suggests that competitive co-adaptation rests on the nonlinear interaction across time scales and not a simple sequence of independent events. Attacker success increased with greater anticipatory synchronisation and less multifractal nonlinearity than for the defender and interpersonal angle. More broadly, present findings encourage future study of the nesting in affordance layouts and network relationships unfolding across individuals, teams, and competitive interactions. Multiscale approaches to ecological dynamics approach might allow stepping beyond performance analysis towards querying the values (Hodges & Rączaszek-Leonardi, 2022) guiding the selection of constraints for athletic participation and the self-organisation of group-tensegrity structures. This strategy might allow a more active and responsible role in shaping the future of sport.
Footnotes
Author’s Note
For the purpose of open access, the author has applied a Creative Commons Attribution (CC BY) licence to any Author Accepted Manuscript version arising from this submission.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Author Biographies