
Abstract
Introduction. As a tumor suppressor, germline and somatic inactivation of BRCA1 associated protein 1 gene (BAP1) is a common finding in mesothelioma, melanocytic tumors, clear cell renal cell carcinoma and several other epithelial, mesenchymal and neural tumors. Incidence of BAP1 genetic alterations and subsequent expression loss has not been well established in non-small cell lung carcinoma (NSCLC) by large-scale studies. Design. After IRB approval, a total of 356 NSCLC cases of our institution between July 2016 and June 2020 were reviewed. The study cohort consisted of 214 (60%) adenocarcinomas, 89 (25%) squamous cell carcinomas, and 53 (15%) diagnosed as “non-small cell lung carcinoma” without specified subtype. All tumors underwent comprehensive target cancer gene next generation sequencing (Oncomine Assay). The protein expression status of BAP1 was subsequently evaluated by immunohistochemistry. Results. BAP1 somatic mutations were detected in 8 NSCLC tumors (incidence: 2.2%). Tumors harboring BAP1 mutations were all diagnosed at advanced stage and carried at least one additional genetic alteration. Immunohistochemically, four tumors showed complete loss of BAP1 protein expression, including two adenocarcinomas which harbored different missense BAP1 mutations and another two with bioinformatically predicated deleterious frameshifting mutations. Conclusion. Compared with known BAP1 loss associated other malignancies, such as mesothelioma, inactivation of BAP1 by somatic mutation is a rare occurrence in NSCLC. BAP1 mutations and loss of expression in NSCLC are accompanied by other complex genetic alternations, suggesting BAP1 mutation maybe a late event NSCLC carcinogenesis.
Introduction
BAP1 (BRCA1-associated protein 1) is a tumor suppressor gene located at 3p21.1–3 BAP1 encodes a nuclear localizing deubiquitinating enzyme, which was first identified as a binding protein to the BRCA1 RING finger domain and believed to cleave ubiquitin and enhance the growth suppressive effects of BRCA1. 4 Later studies showed that BAP1 has a role in BRCA1 mediated processes as well as BRCA1 independent functions.5–8 Currently, the tumor suppressor function of BAP1 is thought to be involved cell cycle regulation, chromatin modulation, transcriptional regulation, ubiquitin-proteasome degradation, and the DNA damage response pathway.2,4,5,6–8
Loss or inactivation of BAP1 may occur as a result of chromosomal deletions involving the BAP1 gene locus, or due to sequence variation in the BAP1 gene.1,2 Several different alterations in the BAP1 gene have been described, including large deletions; frameshift mutations due to insertion or deletions; splice-site mutations, and base substitutions leading to nonsense and missense mutations.1,7,8 Among these, frameshift mutations, missense and nonsense mutations are the most common sequence alterations. 8 Given the complex function of BAP1, it is probable that mutations in different regions of the gene in which they occur lead to different functional consequences.2,7,8 It has been hypothesized that BAP1 mutations may yield different tumor phenotypes, depending on the cell types and the co-existing oncogenic mutations. 7
Germline mutations of BAP1 are known to confer increased susceptibility for a variety of tumors including uveal melanoma,3,9,10 atypical Spitz tumors,3,10 cutaneous melanoma,3,10 mesothelioma,11–17 renal cell carcinoma7,18–20 and possibly other epithelial, mesenchymal and neural tumors.7,21 In addition to germline BAP1 mutations, sporadic, non-familial tumors may harbor somatic mutations. 2 Double-hit inactivation of BAP1 with subsequent loss of expression of the BAP1 protein occurs in about 60% of mesotheliomas.12–17 One study found BAP1 mutations were significantly more common in pure epithelioid mesotheliomas than in biphasic or sarcomatoid variants, 16 while another study failed to confirm such an association. 17
As an important differential diagnosis, non-small cell lung carcinoma (NSCLC) can be morphologically similar to mesothelioma, in particular the epithelioid subtype. Usually a set of immunohistochemical stains including calretinin, WT1, high-molecular-weight keratin 5/6, and D2-40, can help distinguish mesothelioma and NSCLC.22,23 However, about 10% to 20% of tumors exhibit confounding immunohistochemical results or only a portion of tumor cells express certain markers. 23 Since BAP1 inactivation is found in more than 60% of sporadic mesothelioma, BAP1 immunostaining has been suggested to improve the ability to differentiate mesothelioma from NSCLC based on the presumption that BAP1 expression will be frequently lost in mesothelioma.11–17 Loss of expression of BAP1 has also been proposed as a specific marker for mesothelioma diagnosis in cytology specimens.12,24–27 In addition, immunohistochemistry for BAP1 in lung carcinoma has been performed in tissue microarray format.28,29 Nevertheless, the incidence of BAP1 mutation in NSCLC has not been fully explored.
In the current study, we sought to assess BAP1 mutation leading to expression loss in a large cohort of lung cancer that underwent comprehensive molecular profiling (Oncomine) in our institution.
Materials and Methods
Non-Small Cell Lung Carcinoma Patients Cohort and Tissue Collection
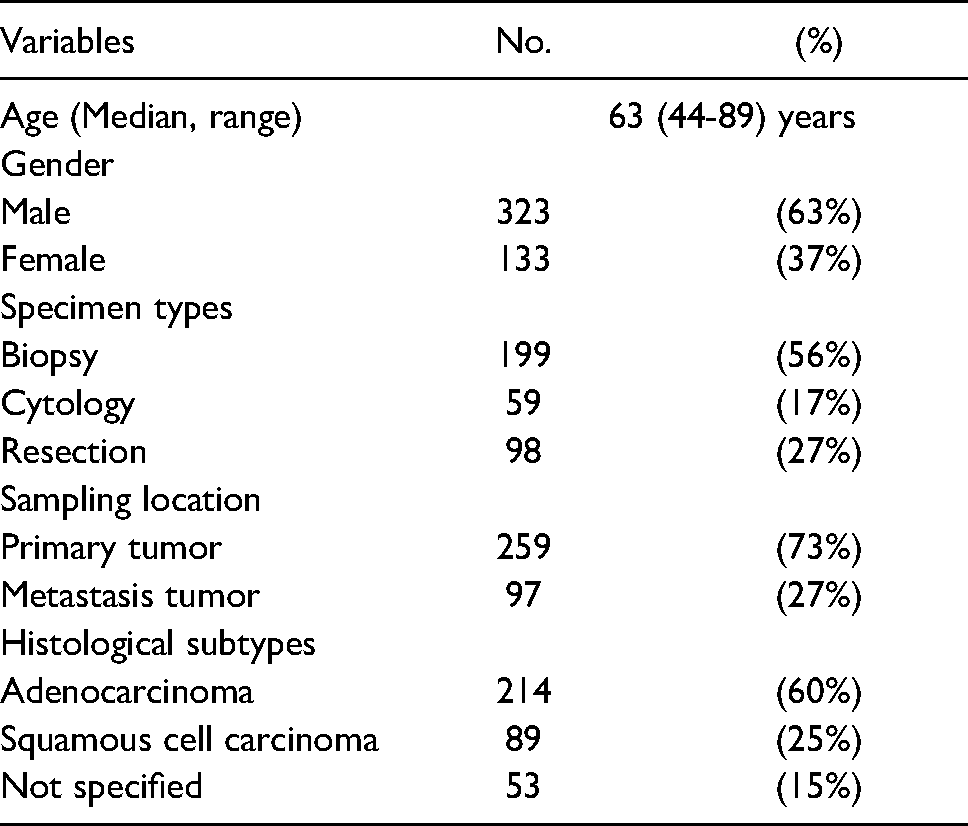
Following Institutional Review Board approval of this study, we collected tumor slides and blocks from 356 patients with NSCLC accessioned to our department pathology files between July 2016 and June 2020. All hematoxylin and eosin-stained slides and immunohistochemistry for all study tumors were reviewed and representative areas were selected for additional testing. Clinicopathologic information from patients was collected from clinical records and pathology reports. A description of the cohort is summarized in Table 1.
Characteristics of Non-Small Cell Lung Carcinoma Cohort.
Molecular Profiling
All NSCLC tumor specimens underwent targeted Next Generation Sequencing (Oncomine Assay) at our Tumor Profiling Laboratory. 30 Briefly, the NSCLC samples were enriched for malignant cells by microdissection of formalin-fixed, paraffin-embedded tissue. Germline control DNA from the sample patient was obtained either from formalin-fixed, paraffin-embedded non-tumor tissue, or from patient's blood or a buccal swab. Genomic DNA was extracted from both tumor and nontumor samples using the Gentra PureGene extraction kit (Qiagen, Valencia, CA) and was quantitated on a Qubit 2.0 fluorimeter. The isolated DNA was amplified using a modification of the On AmpliSeq Oncomine Comprehensive Assay Version 3 multiplex PCR primer sets. The primers amplify 3781 amplicons within 161 genes. Amplicon sequencing was performed on an Ion S5tm XL next generation sequencer (ThermoFisher Scientific, Inc.). The raw data generated was preprocessed within Torrent Suite with the alignment, variant calling and copy number variant detection. Chromosomal positions of variants refer to the human reference sequence assembly released by the Genome Reference Consortium in February 2009 (GRCH37). The analytic sensitivity (lower limit of detection) of the test is estimated to be 5%.
Immunohistochemical Staining
Cell blocks of cytology specimens were prepared by CellientTM method. 31 Immunohistochemistry was performed on representative tumor resection, biopsy cores or cytology cell block slides according to our histology laboratory protocol. Briefly, after deparaffinization and rehydration, heat-induced antigen retrieval was performed using the DAKO high pH antigen retrieval solution in a steamer for 20 min at 97 °C followed by incubation in warm buffer. All sections were incubated with a 1:50 dilution of a mouse monoclonal primary anti-BAP1 antibody (clone C-4, Santa Cruz Biotechnology, Dallas, Texas USA). Detection of the immunostaining was carried out utilizing the Envision FLEX/HRP detection system (DAKO A/S, Glostrup, Denmark) with the 3- diaminobenzidine (DAKO) as chromogen.
After counterstaining with hematoxylin, the slides were independently assessed by two pathologists (TS and MZ) for detection of loss of nuclear staining of BAP1. Negative staining was defined as complete loss of nuclear staining, irrespective of cytoplasmic staining, in tumor cells with retention of staining in non-tumor tissue (normal lung tissue and inflammatory cells) from the same section.
Statistical Analysis
Multivariant regression analysis and other statistical analyzes were performed using Prism 8 GraphPad Prism 8.0 software (San Diego, CA).
Results
Non-Small Cell Lung Carcinoma Cohort Characteristics
A total 356 NSCLC cases were recruited in the current study (Table 1). The diagnosis of NSCLC was confirmed in tissue samples and histologic subtypes of tumors were identified using the World Health Organization classification. 32 Among the cohort, 37% of patients are female (N = 133) and 63% are male (N = 323), with a median age of 63 years-of-age (range, 44 to 89 years). More than half of the samples were obtained through small bore biopsy (199, 56%) or fine needle aspiration (FNA) cytology specimens (59, 17%), while 98 samples were selected from primary resection specimens (27%). Most of the cases (259, 73%) were from primary lung tumor, whereas 97 cases (27%) were from lymph node or distant organ metastases. None of the NSCLC cases received neoadjuvant or adjuvant chemotherapy or radiation therapy before tissue sampling. Among them, 214 (60%) were adenocarcinoma, 89 (25%) were squamous cell carcinoma, and 53 (15%) were diagnosed as “non-small cell lung carcinoma” without specified subtype.
BAP1 Gene Mutation
At least one missense or nonsense mutation in BAP1 gene was detected in 8 tumors with an overall incidence of 2.2% in all screened NSCLC tumors. The incidence of BAP1 gene mutation was slightly higher than, but still comparable to what reported in lung cancer cohort of The Cancer Genome Atlas Program (TCGA database), which was 1.4% (8 of 566) in lung adenocarcinoma and 0.8% (5 of 584) in lung squamous cell carcinoma.
Clinical, pathological and molecular characteristics of these 8 patients, including other detected genetic alterations, are summarized in Table 2. Among these 8, 4 specimens were from primary lung tumors, and the other 4 were from metastatic tumors (50%), including adrenal, lymph node and liver metastases. Five tumors were histologically diagnosed as adenocarcinoma, 2 were classified as squamous cell carcinoma, whereas 1 was called NSCLC without further subclassification. The majority patients (7 of 8 cases) with BAP1 mutation were stage IV disease at diagnosis. Only one patient was diagnosed at stage IIIA. During a median time of 42-month clinical follow-up, four patients died of disease, resulting in a median 25-months overall survival time. More than 60% of patients had smoking history (N = 6, range 8 to 80 packyears). Smoking related lung cancer driver genes, such as KRAS and PTEN, were identified in NSCLC of smokers.
Clinical, Pathological and Molecular Characteristics of Lung Non-Small Cell Carcinoma with BAP1 Mutation.
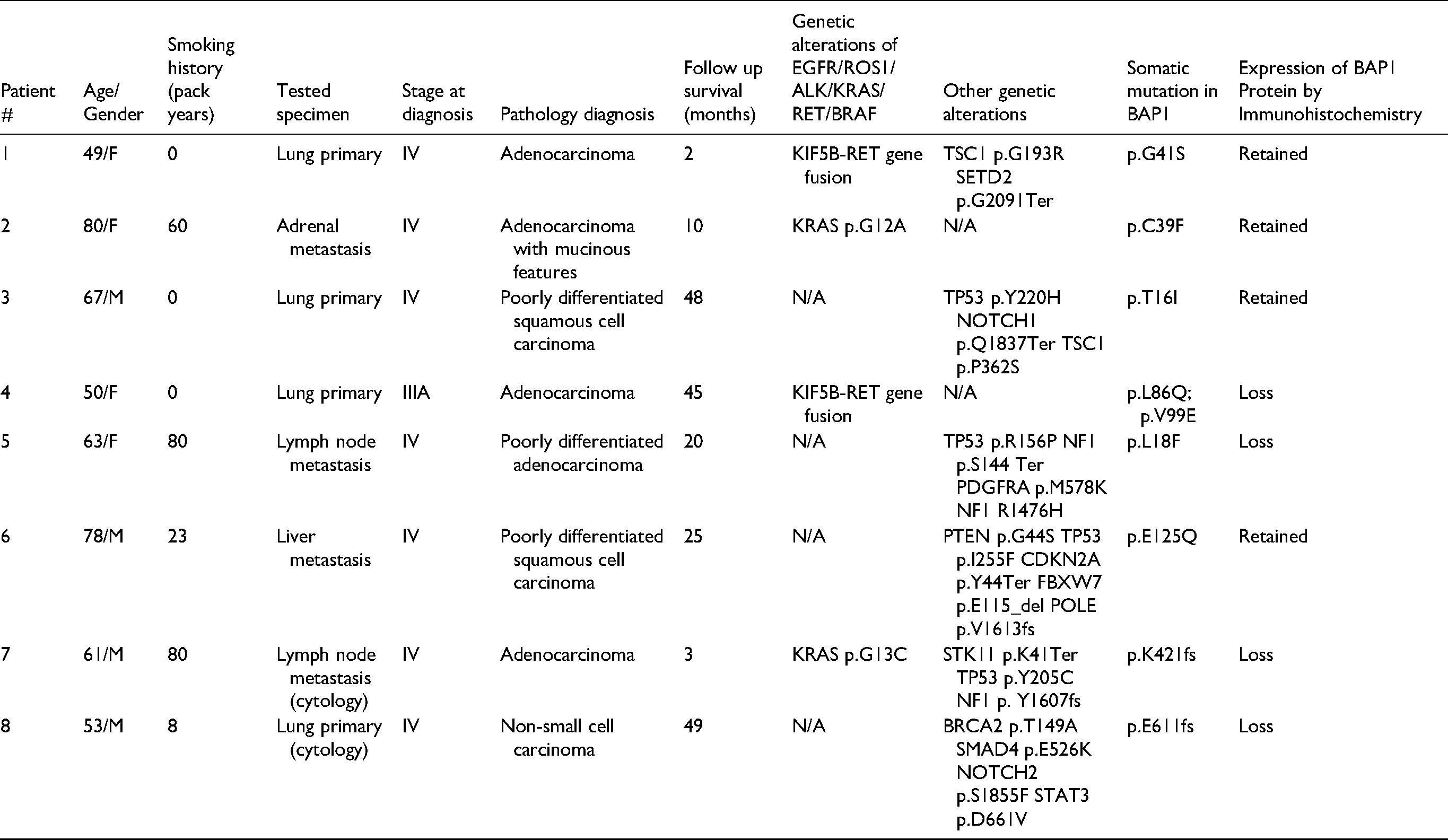
Six of 8 tumors harbored missense BAP1 somatic mutations, while the other 2 had reading frameshifting mutations. One tumor had 2 different loci missense mutation of BAP1 simultaneously. Half of these BAP1 genetic alterations have been reported by Catalog of Somatic Mutations in Cancer (COSMIC) database. All tumors were also found harboring at least one pathologic genetic mutation or other alterations affecting TP53 (n = 4) and NF1 (n = 2), and NOTCH1, NOTCH2, TSC1, PDGFRA, PTEN, CDKN2A, POLE, STK11, BRCA2, SMAD4 (one each). One-half were found to have positive molecular testing results for CAP/IASLC/AMP suggesting efficacy for targeted tyrosine kinase inhibitor therapy, 33 with 2 harboring RET gene fusion and 2 harboring KRAS mutations.
BAP1 Protein Expression
Four of 8 NSCLC tumors harboring BAP1 somatic mutations (50%) showed complete loss of BAP1 protein expression (Table 2, Figures 1 and 2) from tumor resection or biopsy specimens. Intriguingly, except 2 expected tumors which have reading frameshifting mutations, one harboring 2 missense mutations and another with only one missense mutation, also showed abolished expression. All BAP1 protein-deficient tumors have other genetic alterations, including RET gene translocation and other common cancer-related gene mutations.

Representative lung adenocarcinoma resection specimens with immunohistochemical stain of BRCA1-associated protein 1 (BAP1) and corresponding H&E sections. (A) and (B) Tumor #2. A lung adenocarcinoma with BAP1 mutation but retained BAP1 expression. (C) and (D) Tumor #4. A lung adenocarcinoma with BAP1 mutation and RET1 fusion, showing papillary growth pattern and loss of BAP1 expression. (E) and (F) Tumor #5. A lung adenocarcinoma tumor with BAP1 mutation and other multiple oncogenic mutations, showing a poorly differentiation morphology. Note normal respiratory epithelium and stromal and inflammatory cells showing positive nuclear BAP1 expression, serving as positive internal controls.

Cell blocks from two cytology specimens with lung adenocarcinoma. (A) and (B) H&E and BAP1 immunohistochemical stains in a NSCLC metastatic to mediastinal lymph node fine needle aspiration with metastatic lung adenocarcinoma (Tumor #7). (C) and (D) H&E and BAP1 immunohistochemical stains in a lung nodule FNA specimen with poorly differentiated carcinoma (Tumor #8). Note background lymphocytes and normal respiratory cells are serving as positive internal controls.
Discussion
NSCLC has been reported in patients with germline mutation in BAP1 as a rare BAP1 tumor predisposition syndrome manifestation.1,7 For common sporadic NSCLC cases, genetic alterations in BAP1 gene and consequent expression loss have not been fully explored.28,29 In the current study, we comprehensively sequenced BAP1 gene and assessed the mutations which leading to expression loss in a large NSCLC cohort. Our findings confirmed that sporadic mutation of BAP1 is a rare event in NSCLC but can lead to loss of expression.
Several different alterations in the BAP1 gene have been described, including large deletions of exons, frameshift mutations, splice site mutations and the base substitution leading to nonsense and missense mutations.2,17,18 Frameshift mutations, missense and nonsense substitutions are the most common sequence alterations. In our study, we also found missense and frameshift mutations are the most common BAP1 alterations in NSCLC cases and have the potential to result in expression loss of BAP1. Intriguingly, identified missense mutations which did not lead to BAP1 loss were not reported by COSMIC, indicating those were probably not truly pathogenic.
Deletion of the 3p21 region, the chromosomal location of BAP1, is a frequent and early event in the formation of a multitude of different tumors.2,3,17,18 In addition, epigenetic inactivation of genes located in the region is a common and important event in the carcinogenesis.2,7 Given the importance of BAP1 in tumorigenesis, loss of heterozygosity (LOH) or deletion in the 3p21 region could play a role in NSCLC. In our study, two patients with frameshift mutation and one with double missense mutation showed loss of BAP1 expression. In addition, one NSCLC tumor with only one detected missense mutation also showed complete loss of BAP1 protein expression, which may be explained by an underlying undetected cause of LOH.
Loss of BAP1 protein expression has been identified as an adverse prognostic indicator in several different cancers, potentially related to its role in tumor suppression.7,10,19 In studied cases of malignant mesothelioma, loss of BAP1 detected by immunohistochemistry has shown contradictory prognostic implication, with some studies reporting improved patient outcomes with loss of BAP1.13,14,16 Our study data showed that NSCLC with BAP1 mutations and consequent BAP1 expression loss are with at least one additional common lung carcinogenesis mutation, indicating that BAP1 loss might be a late event.
Our study has limitations. Loss or inactivation of BAP1 may occur as a result of chromosomal deletion involving the BAP1 gene locus (3p21.1), or due to sequence variation in the BAP1 gene.2,7,17,18,21 Two types of molecular genetic testing have been used to detect BAP1 mutations. 34 Sequence analysis can detect variants that are benign, of uncertain significance and pathologic, with a reported pathogenic variant detection rate of more than 87.5%. Other gene-targeted deletion/duplication analysis methods including PCR, multiplex ligation-dependent probe amplification (MLPA) and gene target microarray can detect intragenic deletion or duplications, which can identify the parts of the remaining 12.5% of the pathogenic variants. 34 In the current study, we utilized sequencing-based method. Though sequencing-based methodology can detect most pathogenic mutations in BAP1 in NSCLC, a potential risk of missing less frequent deletional alteration in the BAP1 gene in NSCLC exists. Furthermore, BAP1 loss has been reported in other cancer could be due to epigenetic alteration. 35 In current study, we did not examine BAP1 expression status in tumors without detected BAP1 mutations and might miss potential BAP1 loss caused by other mechanisms.
Given the high incidence of BAP1 double-hit inactivation in mesothelioma, loss of expression of BAP1 has been considered as a reliable adjunct and strongly bolsters the diagnosis of mesothelioma in effusion cytology cell block preparation.24–27 Though loss of immunohistochemical staining for BAP1 is an attractive ancillary marker for mesothelioma exhibiting high specificity, our data from cytology specimens suggest that BAP1 immunohistochemistry should not be used alone without other lineage markers such as calretinin or WT1 for diagnosing mesothelioma. 22 Two NSCLC cytology specimens in our study with frame shifting mutations showed complete loss of BAP1 expression. Cytology specimen often has limited cellularity. Without lineage marker support, the utility of BAP1 as diagnostic marker alone should be interpreted with extra caution, since NSCLC is the most important differential diagnosis of epithelioid mesothelioma. Our results demonstrated a potential diagnostic pitfall for negative staining for BAP1 in cytology specimens.
In conclusion, BAP1 inactivating mutation rarely occurs in NSCLC, which is often accompanied with other complex genetic alteration, and is frequently associated with advanced stage disease.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This study is supported by grants from Department of Pathology, Yale School of Medicine.
Ethical Approval
All specimens from the patients in this study were obtained with appropriate consents and approval of the institutional review board of Yale School of Medicine (IRB no. 9505008219). All samples were made anonymous and waived the need for written informed consent.
Informed Consent
Not applicable, because this article does not contain any studies with human or animal subjects.
Trial Registration
Not applicable, because this article does not contain any clinical trials.