
Abstract
Since the discovery of short, regulatory microRNAs (miRNA) 20 years ago, the understanding of their impact on gene regulation has dramatically increased. Differentiation of cells requires comprehensive changes in regulatory networks at all levels of gene expression. Posttranscriptional regulation by miRNA leads to rapid modifications in the protein level of large gene networks, and it is therefore not surprising that miRNAs have been found to influence the fate of differentiating cells. Several recent studies have shown that overexpression of a single miRNA in different cellular contexts results in forced differentiation of nerve cells. Loss of this miRNA constrains neurogenesis and promotes gliogenesis. This miRNA, miR-124, is probably the most well-documented example of a miRNA that controls nerve cell fate determination. In this review we summarize the recent findings on miR-124, potential molecular mechanisms used by miR-124 to drive neuronal differentiation, and outline future directions.
Introduction
Neurogenesis, the formation of new nerve cells from neural stem/progenitor cells (NSPCs), is tightly controlled at a molecular level. MicroRNAs (miRNAs) are small, 21 to 23 nucleotides long, single-stranded, endogenously expressed RNA that have been shown to play a central role in this process (Bartel 2009; Shi and others 2010). The mature miRNA is initially produced from a longer primary miRNA, which is transcribed from a miRNA encoding gene. While remaining in the nucleus, it is cleaved by the RNAse III enzyme Drosha, part of the microprocessor complex, forming a precursor miRNA. Once exported out from the nucleus, it is recognized by the RNAse III enzyme Dicer, which cleaves the miRNA to its mature form. The miRNA is incorporated into the RNA-induced silencing complex (RISC) and guides this complex to complementary 3′ untranslated regions (UTRs) of mRNAs where it acts through degradation or by repressing translation (Bartel 2009) (Fig. 1).
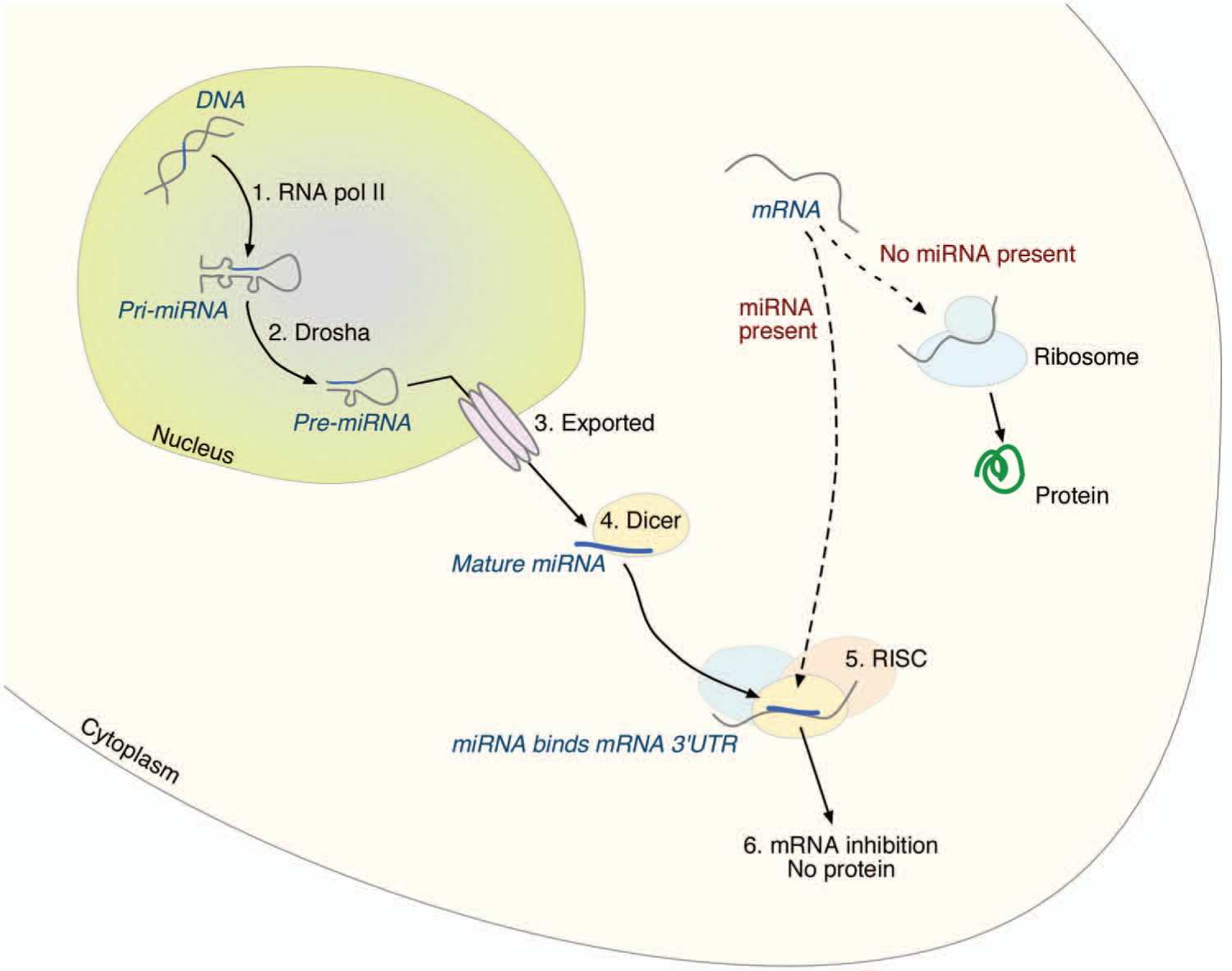
The miRNA biogenesis pathway. 1. The gene encoding the miRNA is transcribed by RNA polymerase II, producing a primary miRNA (pri-miRNA). 2. The pri-miRNA is further cleaved by Drosha, part of the microprocessor complex, to a precursor (pre-miRNA). 3. The pre-miRNA is exported out from the nucleus. 4. It is cleaved to its mature length by Dicer. 5. The mature miRNA is incorporated into RISC, where it binds to the 3′ UTR of its target mRNA. 6. The mRNA is inhibited from further translation into protein.
A single miRNA can have hundreds of target genes, and more than a thousand miRNAs have been identified in human cells. In addition, each target gene can have binding sites for several miRNAs, suggesting a broad and complex role of miRNAs on protein translation (Griffiths-Jones 2006). An increased understanding on how miRNAs control neural fate determination could in the end be exploited to enhance cell reprogramming and aid in the development of novel molecular targets in pathogenic states.
miRNAs in Brain Development
To understand the importance of miRNA in brain development, knockout mice of the Dicer enzyme have been generated to remove most mature miRNAs. Deletion of Dicer in mice results in death before axis formation (Bernstein and others 2003). Conditional Dicer ablation during brain development leads to major cellular, histological, and anatomical disturbances, suggesting a role for miRNAs in this process (Davis and others 2008; De Pietri Tonelli and others 2008; Kawase-Koga and others 2009). However, Dicer itself has other functions (Kaneko and others 2011), and the disruption of this gene could result in cellular consequences independent of miRNA, complicating the conclusion of the observed phenotypes. In addition, the long half-life of certain miRNAs must be taken into account when analyzing conditional knockouts (Konopka and others 2010), further complicating the interpretation after Dicer ablation. Therefore, additional studies where individual miRNAs are perturbed are necessary to understand their true function. Unfortunately, this is a complicated task because (a) one miRNA species can be encoded at multiple regions in the genome, (b) it can be located within genes, or (c) it is situated in clusters with other miRNAs (Griffiths-Jones 2006).
miR-9 and miR-124, two miRNAs known to be involved in neurogenesis, are transcribed from three different loci in mice and humans (miR-9-1, miR-9-2, miR-9-3 and miR-124-1, miR-124-2, miR-124-3, respectively), but their mature sequences are identical and conserved over species (Griffiths-Jones 2006). Sanuki and coworkers (2011) made a miR-124-1 knockout by depleting retinal noncoding RNA 3 (Rncr3), which is the dominant source of miR-124. Loss of miR-124-1 resulted in neural dysfunction and aberrant neuronal maturation (Sanuki and others 2011). For miR-9, a knockout mouse of miR-9-2 and miR-9-3 has been generated. These mice displayed severe implications in brain development (Shibata and others 2011). Although these studies demonstrate the importance of miR-9 and miR-124, it will be necessary to delete all three copies of these miRNAs to fully understand their role in the developing brain, because the remaining transcripts are likely to partially compensate for the deleted isoforms.
Recently, alternative approaches to permanently inhibit miRNA have been developed, such as the use of sponge viral vectors (Gentner and others 2009). Sponges are transgenic transcripts complimentary to the mature miRNA, except for a region of approximately four base pairs that forms a bulge when paired. As a result, the targeted miRNA is sequestered or degraded and cannot inhibit its natural mRNA targets (Ebert and others 2007). The use of viral vectors to stably express the sponge sequence has been demonstrated to efficiently inhibit miRNA activity and is a promising approach for miRNA loss-of-function studies (Akerblom and others 2012; Bhalala and others 2012; Gentner and others 2009; Luikart and others 2011; Mei and others 2011; Pathania and others 2012). The use of sponge expressing vectors is a feasible advance in the permanent knockdown of an individual miRNA during brain development, circumventing the disadvantages with knockouts of miRNA or Dicer. Possibly, generation of sponge-expressing transgenic mice could stably inhibit the desired miRNA family. Another approach could be the use of in utero electroporation to introduce sponges at an early, specific time point during development. This method has the advantage of targeting specified brain areas. Thus, there are possibilities using novel technologies to study individual miRNA loss of function during brain development.
miR-124 as a Neuronal Fate Determinant
miR-124 was first reported as a brain-specific miRNA and has more recently been described as neuron specific. In 2002, Lagos-Quintana and coworkers estimated that miR-124 accounts for 25% to 50% of total adult brain miRNA, and its high expression has been confirmed in many studies (Reviewed in Gao 2010). Its expression is initiated some time during the differentiation process from neural stem cell (NSC) to neural precursor cell, and the levels accumulate on neural maturation (Fig. 2A) (Akerblom and others 2012; Cheng and others 2009; Krichevsky and others 2006; Maiorano and Mallamaci 2009), suggesting that it plays a central role in regulating transcripts, hence protein products, in nerve cells. It is not expressed in other cell types in the CNS such as ependymal cells, astrocytes, microglia, oligodendrocytes, or NSCs (Akerblom and others 2012).
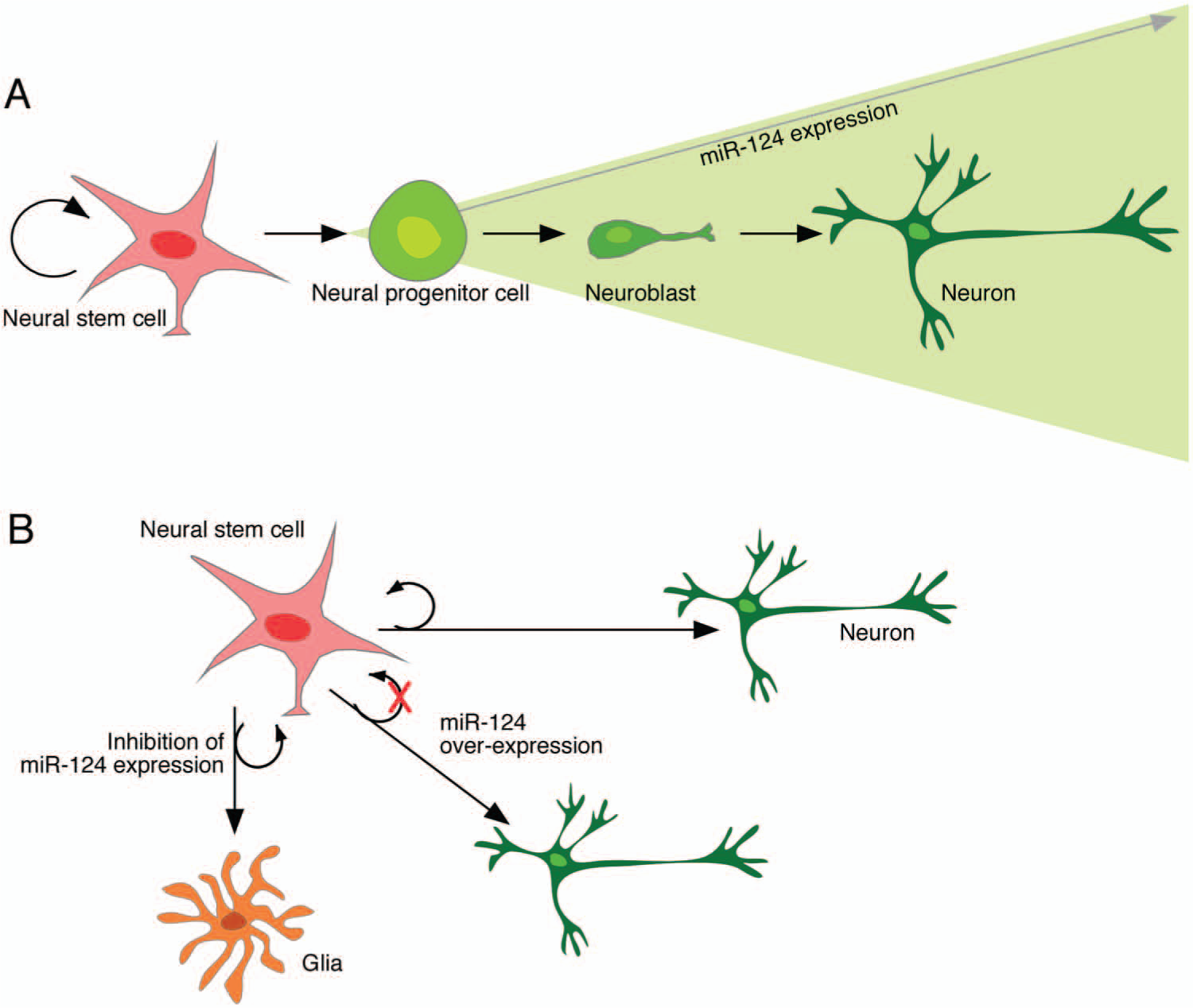
Expression and function of miR-124 in the adult subventricular zone. (A) miR-124 is not expressed in neural stem cells (NSCs); its expression is initiated at the neural progenitor stage, and expression increases with neural maturation. (B) When miR-124 is knocked down in NSCs, neurogenesis is blocked and gliogenesis is up-regulated. On the contrary, overexpressing miR-124 in NSCs leads to precocious neural maturation and loss of NSC self-maintenance.
There are several studies demonstrating that overexpression of miR-124 in neural progenitors, embryonic stem cells, and glioma cells results in forced neuronal differentiation (Krichevsky and others 2006; Lang and others 2012; Lim and others 2005; Silber and others 2008; Smirnova and others 2005; Xia and others 2012). Overexpression of miR-124 results in up-regulated expression of neuronal markers as well as morphological changes including enhanced neurite outgrowth and complexity (Sanuki and others 2011; Yoo and others 2011). In vivo studies have shown that miR-124 can promote neurogenesis during brain development as well as in the adult subventricular zone (SVZ) stem cell niche (Akerblom and others 2012; Cheng and others 2009). The effect of miR-124 appears to be, at least to some extent, independent of cellular context. One classical example is the delivery of miR-124 duplexes to HeLa-cells that induces a neuronal gene profile (Lim and others 2005). More recently, it has been shown that the expression of miR-124 (when overexpressed together with miR-9*) in fibroblasts induces the conversion to neurons (Yoo and others 2011). Also, repression of PTBP in fibroblasts, which is suppressed by miR-124 during normal brain development (see below), is sufficient to transdifferentiate fibroblasts into neurons (Xue and others 2013). These studies suggest that miR-124 has an instructive role in promoting neuronal differentiation.
Loss of function studies of miR-124 in NSPCs has yielded more inconsistent results, probably due to technical issues. However, we have recently used lentiviral sponge vectors to permanently suppress miR-124 in the adult SVZ-stem cell niche. This experiment demonstrated that neurogenesis was down-regulated and replaced with gliogenesis (Fig. 2B), with the result of astrocytes migrating to the olfactory bulb (Akerblom and others 2012). Taken together, these data suggest that miR-124 acts as a neuronal fate determinant in NSPCs.
Molecular Targets of miR-124
As mentioned above, overexpression of miR-124 results in forced differentiation of progenitors to neurons. As miRNAs act as repressors, this suggest that miR-124 block pathways that maintain the stem/progenitor states and also block pathways driving alternative cell fates such as towards glia. Thus, miR-124 is likely to mediate its effects in a complex manner on several levels. In line with this, computational predictions as well as experimental approaches suggest that miR-124 has the potential to regulate hundreds of target genes. Several individual genes as well as regulatory networks have been suggested and experimentally verified as the functional downstream targets for miR-124 (Fig. 3).
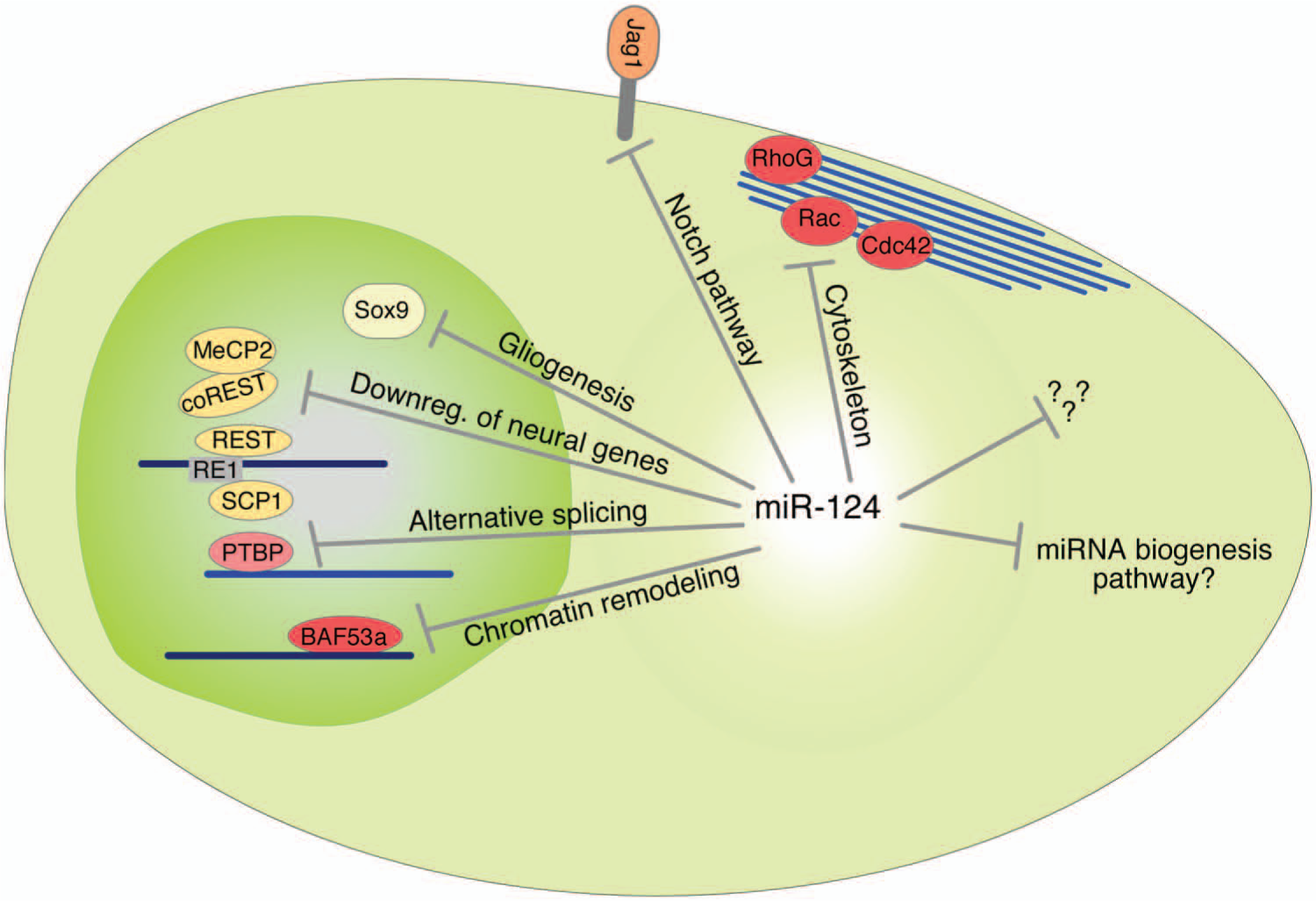
miR-124 target genes and pathways. Summary of miR-124 target genes and pathways important in neural fate determination.
RE1 silencing transcription factor (REST) inhibits neural genes in nonneural cells by recruitment of histone deacetylases. Timely down-regulation of REST leads to neural differentiation (Qureshi and others 2010). In neurons, miR-124 inhibits REST, allowing neuronal genes to be expressed. However, in nonneural cells miR-124 is kept silent by REST. Thus, miR-124 and REST act in a reciprocal manner (Conaco and others 2006; Visvanathan and others 2007; Yoo and others 2009) to establish the neuronal fate. Moreover, small c-terminal domain phosphatse 1 (SCP1) is another important nonneural factor whose down-regulation is required to promote neural differentiation. SCP1 is recruited to neural genes containing RE1, linking it to the REST regulatory pathway. miR-124 suppresses SCP1 in the developing spinal cord, which in turn promotes neural differentiation (Visvanathan and others 2007). In addition, computational approaches found miR-124 binding sites in the 3′ UTRs of MeCP2 and coREST (Wu and Xie 2006). Thus, miR-124 has several targets in the REST pathway and thereby confers neural identity.
The Notch signaling pathway controls NSPC maintenance (Louvi and Artavanis-Tsakonas 2006). The Notch transmembrane receptor is expressed on NSCs, and by binding to its transmembrane ligand Jagged1 (Jag1), neural stem cell self-renewal is promoted and differentiation is inhibited (Imayoshi and Kageyama 2011). miR-124 suppresses Jag1 and thereby allows exit from the nondifferentiated state (Cheng and others 2009; Liu and others 2011). The Sox9 transcription factor that regulates glial fate specification during development is another miR-124 target in the SVZ (Cheng and others 2009). The suppression of Sox9 is likely to participate in miR-124’s role in blocking gliogenesis.
Furthermore, miR-124 has been reported to promote neural differentiation by triggering brain-specific alternative pre-mRNA splicing (Makeyev and others 2007). PTBP1 is a global repressor of alternative splicing in nonneural cells. PTBP1 targets the pre-RNA of PTBP2 leaving out an important exon (Spellman and Smith 2006). During neural differentiation, miR-124 down-regulates PTBP1, leading to accumulation of correctly spliced PTBP2 protein and increasing brain-specific alternative splicing patterns (Makeyev and others 2007). In addition, PTBP has the potential to regulate the whole miRNA machinery by masking miRNA binding sites on mRNA (Xue and others 2013). As miR-124 suppresses PTBP, an interesting hypothesis may be formed that miR-124 controls the entire miRNA pathway and in that way governs cellular fate.
An important step in the formation of new nerve cells is the mitotic exit, when the NSPC lose its self-renewal capacity and start to differentiate and form stable connections. During development, the transition is associated with a switch in the ATP-dependent chromatin remodeling mechanism. In the neural-progenitor-specific BAF complex (npBAF), the subunits are replaced to form a neuron-specific BAF (nBAF); expression of BAF53a in npBAF allows BAF53b to be expressed in nBAF, leading to postmitotic neural development and dendritic morphogenesis. If the switch of subunits fails, mice have defects in neural tube closure. miR-124, together with miR-9*, is responsible for the repression of BAF53a, allowing BAF53b to be expressed. This expression is necessary for loss of proliferation and dendritic outgrowth (Yoo and others 2009).
Taken together, these studies confirm that miR-124 targets a large network of genes and pathways in NSPCs (Fig. 3). Still there is no complete understanding of the molecular events that underlie the capacity of miR-124 to promote neuronal differentiation. Future studies should aim to provide experimental data on the complete set of miR-124 targets in NSPCs. Such studies are necessary to appreciate exactly how this miRNA can mediate its proneuronal effects.
miR-124 in Other Species
The mature sequence of miR-124 and its brain-enriched expression are well conserved over species. However, the aforementioned studies are primarily preformed in mice, and it is becoming clear that the functional role of miR-124 differs in invertebrates. In Aplysia californica, miR-124 is expressed in sensory neurons and inhibits serotonin-induced transmission through the regulation of CREB (Rajasethupathy and others 2009), a transcriptional activator required for long-term potentiation. Deletion of miR-124 in Caenorhabditis elegans did not result in apparent defects in sensory neuron differentiation (Clark and others 2010). In Drosophila melanogaster, miR-124 is expressed in proliferating neuronal precursors and differentiating postmitotic neurons (Weng and Cohen 2012). The complete knockout of miR-124 in Drosophila resulted in almost normal behavior of the flies and they were fully viable with a shorter life span (Sun and others 2012; Weng and Cohen 2012). In contrast to mice, miR-124 is required to support neuroblast proliferation and not to promote differentiation (Weng and Cohen 2012). Also, miR-124 target genes are different between vertebrates and invertebrates. These studies point out a divergence in the functional role of miR-124 between invertebrates and vertebrates such as mice.
miRNA Expression in NSCs and Their Importance in Neurogenesis
As mentioned previously, miR-124 is not expressed in NSPCs and is not initiated until the neural differentiation process has begun. Therefore, it is possible that other miRNAs, which are present in the actual progenitor population, may contribute to neuronal fate determination. For example, Let-7 is the highest expressed miRNA-family in NSPCs, and it constitutes more than 50% of all miRNAs in this cell type, suggesting that the Let-7 family has a large impact on neurogenesis and as neural fate determinants (Marson and others 2008; Wulczyn and others 2007; Zhao and others 2010). Other miRNAs that are highly expressed in NSPCs are miR-103, miR-181a/b, miR17-20, miR-130/301, miR-21, miR-15/16, miR-9, and miR-26, as identified by a small RNA sequencing (Marson and others 2008). In addition, miR-125b and miR-132 have been linked to neurogenesis (Franke and others 2012; Le and others 2009; Luikart and others 2011; Magill and others 2010; Pathania and others 2012; Yoo and others 2009). The functional role of these miRNAs in neurogenesis remains largely unexplored, and it will be very interesting to follow future studies investigating the role of the miRNome in NSPCs.
Conclusion
It is tempting to argue that miR-124 has a powerful and instructive role in governing neurogenesis and neuronal differentiation. This is strongly suggested by (a) its expression and activity increase on the neuronal differentiation process, (b) the presence of dysfunctional neurons in a knockout of two of three miR-124 genes, and (c) the switch of cell fate toward gliogenesis after knockdown of miR-124 in cells of the SVZ. Overexpression of miR-124 in different cell types is potent and promotes a shift to neuronal outcome, probably independent of cellular context. Overexpression of miR-124 in neurons or NSPCs results in precocious differentiation, promotion of neurogenesis, up-regulated expression of neuronal markers, as well as morphological changes. An accurate understanding of the functional role of miR-124 in the brain and in neurogenesis would be greatly aided by the generation of conditional knockout mice capable of deleting all three copies of the miR-124 gene.
Several miR-124-target genes have been validated, and even though they all may be important, the picture is far from complete. miR-124 most likely acts by targeting hundreds of mRNAs implicated in different gene networks. The possibility that miR-124 may influence the whole miRNA regulatory pathway (by regulating PTB) makes it a potentially very broadly acting miRNA. The use of techniques such as HITS-CLIP (Chi and others 2009) to experimentally identify miR-124 targets in NSPCs on a large scale would greatly aid in the clarification of how miR-124 acts on a molecular level.
Not all miRNAs are as powerful influencers on gene regulation as miR-124. Many studies confirm a modest role of individual miRNAs in regulating gene expression (Ebert and Sharp 2012). However, there is emerging evidence that different miRNAs have combinatorial and synergistic roles. This is exemplified by Yoo and others (2009), that demonstrated that miR-9* and miR-124 were both required to reprogram fibroblasts into neurons. Moreover, there is growing number of other noncoding RNA, such as long noncoding RNA (lncRNA), piwi interacting RNA (piRNA), small nucleolar RNA (snoRNA), and small interfering RNA (siRNA) that have been linked to important roles in gene regulation (Qureshi and Mehler 2012). In addition to this, endogenously expressed circular sponges, regulating miRNA expression, demonstrate yet another level of RNA regulation (Hansen and others 2013; Memczak and others 2013). Hence, with miRNA, we might only have begun to understand the impact of noncoding RNAs in fate determination.
Because of the fact that miR-124 has a powerful and instructive role in directing differentiation, it is appealing to speculate that the proneuronal effects could be used for therapeutic purposes. Hypothetically, miR-124 can be exploited to enhance the formation of new nerve cells used in therapy for neurodegenerative disorders such as Parkinson’s disease. It could also be used to differentiate glioma stem cells toward a neuronal fate, thereby restraining the oncogenic characteristic of these cells. In conclusion, increased understanding of the miRNA regulatory mechanisms in fate determination would facilitate finding novel molecular targets for brain disorders.
Footnotes
Acknowledgements
We are grateful to Josephine Malmevik for excellent comments on the article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Our research is supported by the Swedish Research Council (Grant No. 2009-2483), the Swedish Cancer Foundation (Grant No. 2010/400), and the Swedish Brain Foundation.