
Abstract
Chronic tinnitus, the perception of lifelong constant ringing in ear, is one capital cause of disability in modern society. It is often present with various comorbid factors that severely affect quality of life, including insomnia, deficits in attention, anxiety, and depression. Currently, there are limited therapeutic treatments for alleviation of tinnitus. Tinnitus can involve a shift in neuronal excitation/inhibition (E/I) balance, which is largely modulated by ion channels and receptors. Thus, ongoing research is geared toward pharmaceutical approaches that modulate the function of ion channels and receptors. Here, we propose a seesaw model that delineates how tinnitus-related ion channels and receptors are involved in homeostatic E/I balance of neurons. This review provides a thorough account of our current mechanistic understanding of tinnitus and insight into future direction of drug development.
Introduction
Tinnitus is the perception of sound without the presence of external acoustic stimuli. The sound perceived by sufferers is generally subjective and described as a ringing, whistling, clicking, buzzing, or roaring sound. The prevalence of tinnitus varies from country to country in the range between 4.6% and 30.3% (Gilles and others 2013; Jalessi and others 2013; Khedr and others 2010; McCormack and others 2014; Park and Moon 2014; Quaranta and others 1996; Sindhusake and others 2003). Tinnitus can be tolerated but, for about 2% of the patients, it is considered to be severely disabling (Henry and others 2003). Comorbid effects caused by tinnitus such as stress, insomnia, anxiety, depression, deficits in attention, and even suicide, are devastating in everyday life (Cima and others 2011; Crönlein and others 2007; Erlandsson and Hallberg 2000; Gomaa and others 2014; Mazurek and others 2015; Pajor and others 2013; Simoens and Hébert 2012). It has been suggested that tinnitus is usually associated with brain injuries, ageing, noise trauma, emotional and attentional instability (Langguth and others 2013; Martines and others 2015; Shore and others 2016). After being exposed to similar risk factors, some people do not develop tinnitus while others do develop it. This indicates that, other than the environmental risk factors, there is genetic susceptibility to tinnitus (Pawełczyk and others 2012; Sand and others 2011). It is notable that genetic factors are likely associated with chronic tinnitus but not acute tinnitus. Almost everyone develops tinnitus immediately after noise- or drug-induced tinnitus, but few develop chronic tinnitus (Gilles and others 2013; Salvi and others 2009; Widen and Erlandsson 2004). This suggests that chronic tinnitus might have a genetic basis. In line with this, some recent genome-wide association studies revealed a genetic basis for tinnitus (Gilles and others 2017; Vona and others 2017). Nonetheless, the causal association of genetic factors with locus remains to be elucidated.
Despite the various etiologies of tinnitus, it is highly associated with hearing loss. The cellular and neural mechanisms of tinnitus remain unclear. Hence, investigations of animal models where tinnitus is induced by hearing loss have yielded valuable information on the mechanisms of tinnitus (Eggermont and Roberts 2015; Lobarinas and others 2006; Yang and Bao 2013). It has been suggested that the induction of tinnitus is largely due to deafferentation of auditory nerve caused by damage of cochlear hair cells. Deafferentation generally leads to hyperexcitability or enhanced excitation/inhibition (E/I) ratio in central auditory neurons (Jastreboff 1990; Weisz and others 2006). For example, the deafferentation-induced tinnitus can lead to reduced inhibition (Rüttiger and others 2013), increased neural spontaneity (Roberts and others 2010), abnormal synchronous activity (Eggermont 2007), and altered homeostatic plasticity (Yang and others 2012; Yang and others 2011). The auditory areas such as the dorsal cochlear nucleus (DCN) (Koehler and Shore 2013; Shore and others 2008), auditory brainstem (Brozoski and others 2013), inferior colliculus (Mulders and Robertson 2009), auditory thalamus (Caspary and Llano 2017), and auditory cortex (Engineer and others 2011; Llano and others 2012; Yang and others 2011) are associated with tinnitus (Fig. 1). Besides the auditory pathways, other brain regions affected by tinnitus include the limbic system, prefrontal cortex, parietal cortex, cingulate cortex, cerebellum, and subcallosal region (Bauer and others 2013; Boyen and others 2013; Davies and others 2017; De Ridder and others 2011; Langguth and others 2012; Leaver and others 2011; Middleton and Tzounopoulos 2012; Mühlau and others 2006; Rauschecker and others 2010; Roberts and others 2013).
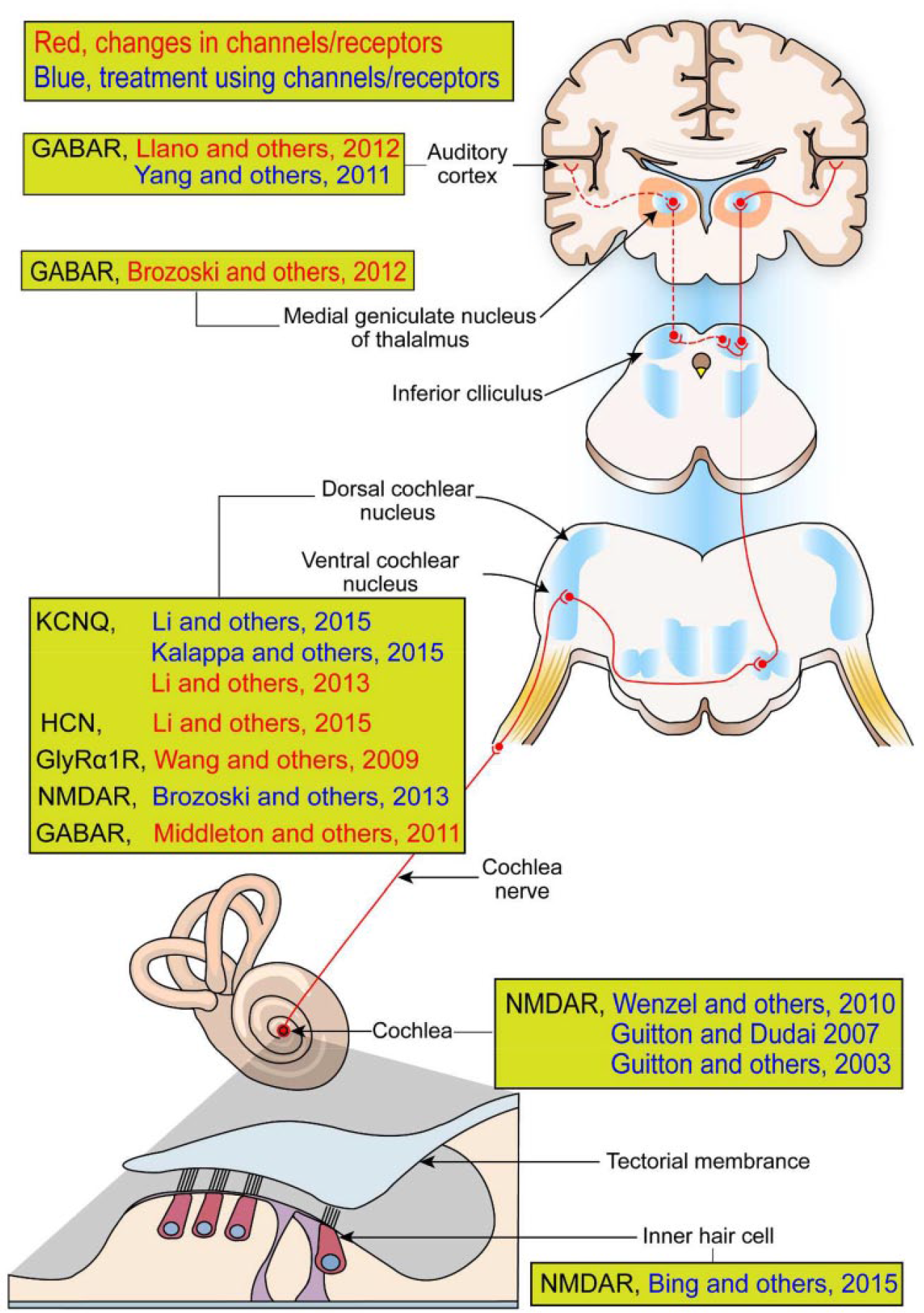
Tinnitus-related ion channels and receptors studied in both the peripheral and central auditory pathway. Red and blue are cases about changes in ion channels and receptors and about treatment using their agonists or antagonists, respectively. It is assumed that effects of ion channels and receptors performed for animals can also be applied to humans, as shown with the human brain. Abbreviations: NMDAR, N-methyl-
Many attempts have been made to treat tinnitus. Therapeutic drugs alleviating tinnitus have been widely utilized with other treatments such as physical therapy, psychological therapy, acupuncture, and acoustic therapy (Joos and others 2015; Langguth and others 2009; Michiels and others 2016; Park and others 2000; Salvi and others 2009; Scott and others 1985). There are lines of evidence that drug therapies are associated with the function of ion channels and/or neurotransmitter receptors (Elgoyhen and Langguth 2010; Langguth and others 2009; Salvi and others 2009). To pinpoint suitable druggable targets, there is a need to understand the roles of ion channels and receptors in tinnitus. Successful drug therapies require long-term efficacy of drug administration with a lack of adverse side effects, including addiction and biocompatibility. We believe that this can be achieved in the near future by focusing on the contribution of ion channels and receptors to tinnitus. Here we provide a broad overview and summary (Table 1) of the involvement of ion channels and receptors and propose that, by modulating these targets and related plasticity in the auditory system, we can develop new treatments for this debilitating disorder.
Summary of Ion Channels and Receptors Studied in Tinnitus.
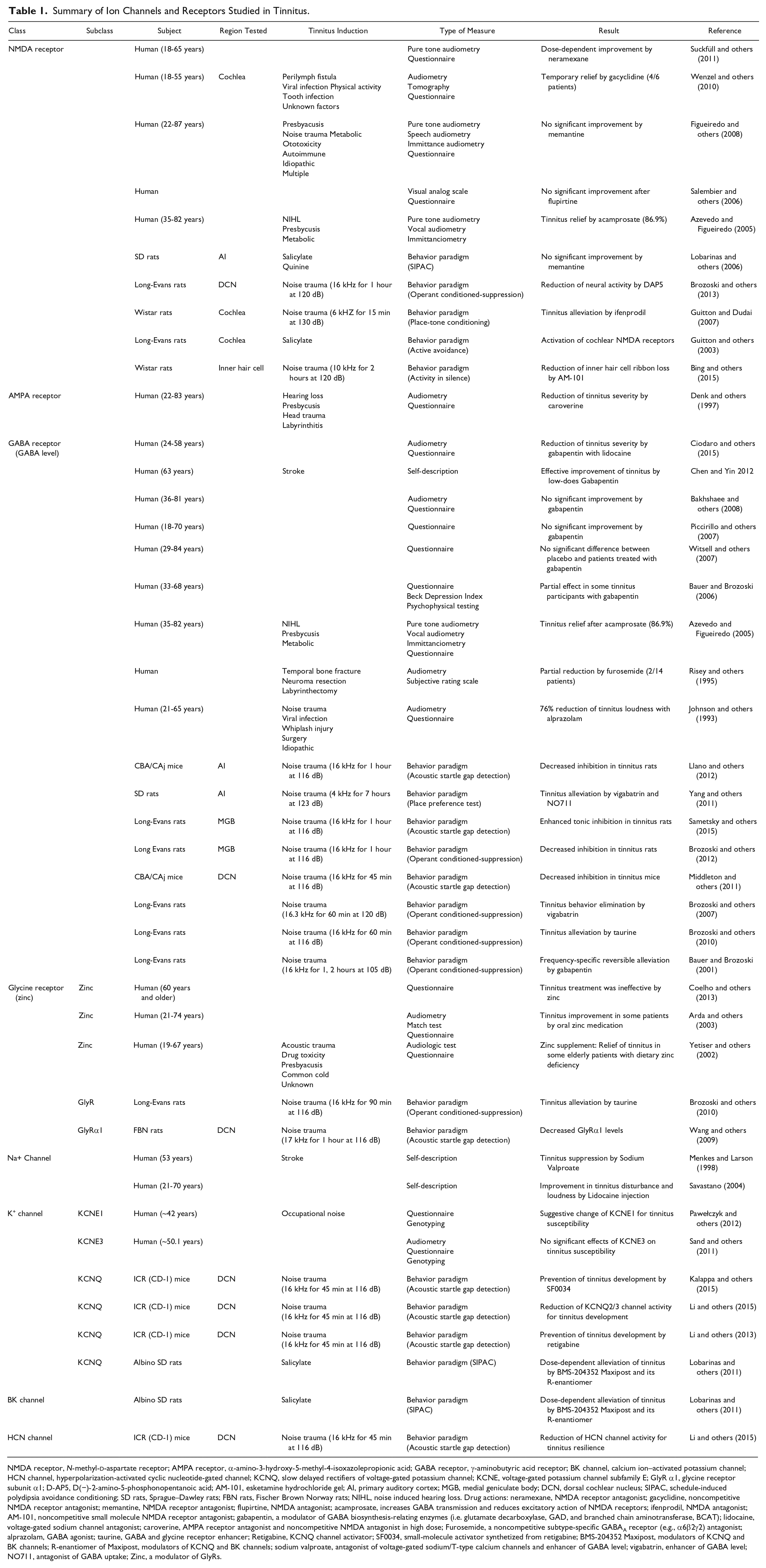
NMDA receptor, N-methyl-
Ion Channels and Receptors in Tinnitus
Ion channels and ionotropic neurotransmitter receptors have aqueous pores located at the plasma membrane of cells and enable ions to move in or out of cells, thereby altering cellular excitability (Hille 2001; Moody and Bosma 2005). There are various types of ion channels/receptors, and they localize to different subcellular compartments such as dendrites, axons, and somata. They can largely be identified by how they are activated and what types of ions move through them. Ion channels/receptors can be gated by several factors such as membrane voltage change, ligand, second messenger, phosphorylation, pH change, temperature and mechanical distortion. It is well-known that dysregulation of ion channels/receptors results in neural dysfunction by altering neuronal synaptic inputs, excitability, or release properties (Weilinger and others 2013). The following is a list of ion channels/receptors shown to be involved in tinnitus (highlighted in Fig. 1).
Glutamate Receptors
Glutamate receptors are the main class of receptors that mediate excitatory transmission in the nervous system (Plested and Mayer 2009). Two major glutamate receptors—AMPA (2-amino-3-(5-methyl-3-oxo-1,2-oxazol-4-yl) propanoic acid) and NMDA (N-methyl-
Glutamate receptor–mediated excitability has been deeply involved in the pathophysiology of tinnitus animal models. In a noise-induced tinnitus animal model, widespread changes in glutamate levels were found along the auditory pathway but in a region-specific manner: Increment in bilateral DCN, bilateral auditory cortex and ipsilateral auditory thalamus; decrement in contralateral auditory thalamus (Brozoski and others, 2012). The application of ifenprodil, an NMDA receptor antagonist, into the cochlea alleviated animal tinnitus behavior of place-tone conditioning (Guitton and Dudai 2007). The cochlear round-window application of another NMDA receptor antagonist, AM-101, reduced the noise-induced loss of ribbon synapses of inner hair cells of AM-101 responders, therein suggesting the reduction of the risk of tinnitus development (Bing and others 2015). Consistent with these findings, tinnitus was attenuated when an NMDAR antagonist, D(−)-2-amino-5-phosphonopentanoic acid (D-AP5) was employed in the rats’ DCN (Brozoski and others 2013). Also, the use of noncompetitive NMDA receptor antagonists, MK-801, 7-chlorokynurenate (7-CK), and gacyclidine, decreased a salicylate-induced tinnitus behavior in rats (Guitton and others 2003; Ruel and others 2008). These studies implicate the significance of NMDA receptors on tinnitus development.
The involvement of NMDA receptors in tinnitus has been also studied with human subjects. When gacyclidine, a noncompetitive NMDAR antagonist, was administered to cochlea of tinnitus patients, 67% of patients experienced temporary recovery from tinnitus (Wenzel and others 2010). In another patient study, oral administration of neramexane, an NMDA receptor antagonist, significantly alleviated tinnitus annoyance and tinnitus impact of life in a dose-dependent manner (Suckfüll and others 2011). Also, caroverine, an AMPA and NMDA receptor antagonist, was able to reduce the severity of tinnitus (Denk and others, 1997). Acamprosate, which is known to reduce the excitatory effect of NMDA receptors, was used to successfully relieve tinnitus symptoms of about 86.9% of subjects (Azevedo and Figueiredo 2005).
It is important to note, however, that treatments using NMDA receptor antagonists have not always been successful in alleviating tinnitus symptoms (Figueiredo and others 2008; Yang and others 2011). Memantine, an open-channel NMDA receptor antagonist, failed to alleviate tinnitus of human participants (Figueiredo and others 2008). Also, memantine failed to alleviate salicylate-induced tinnitus in rats (Lobarinas and others 2006). The oral intake of flupirtine, a drug that antagonizes NMDA receptors by increasing Mg2+ blockage, could not treat tinnitus of participants (Salembier and others 2006). Moreover, patients suffered from severe side effects such as amnesia and attention disorders; it is probably because of the global targeting of glutamate receptors in brain regions that are important for learning and memory such as hippocampus and prefrontal cortex. Nonetheless, many studies with humans and animals showed that antagonism of auditory NMDA receptors could be, in general, a strategy for treating tinnitus symptoms.
γ-Aminobutyric Acid (GABA) Receptors
Two types of major inhibitory receptors, ionotropic GABAA and metabotropic GABAB receptor, play a critical role in decreasing neuronal activity and maintaining homeostatic E/I balance in the brain (Wenner 2011). It is well known that neurotransmitter GABA plays an integral role in signal processing in auditory centers such as the cochlear nucleus, superior olivary complex, inferior colliculus, medial geniculate body (MGB), and auditory cortex (Brozoski and others 2012; Vater and others 1992). As such, altered inhibition has been implicated in hearing diseases including tinnitus.
Animals with noise-induced tinnitus have shown decreased GABAergic inhibition in the auditory DCN, thalamus, and cortex, suggesting the involvement of GABA metabolism in tinnitus (Brozoski and others 2012; Middleton and others 2011). In line with this, taurine, which is known as an enhancer of both GABAA and GABAB receptors, has been reported to ameliorate rat tinnitus behaviors following acoustic trauma (Brozoski and others 2010). Complementary to these findings, the reduction of GABAergic synaptic transmission in rat inferior colliculus was observed in salicylate-induced tinnitus models (Jastreboff 1995; Wang and others 2006). Furthermore, noise-induced tinnitus animals have showed reduction in GABAA-mediated phasic and tonic inhibition along the auditory pathway (Brozoski and others 2012; Sametsky and others 2015; Yang and others 2011). Vigabatrin, a chemical that increases GABA levels, was used to successfully eliminate tinnitus behavior in noise-induced tinnitus rats (Brozoski and others 2007; Yang and others 2011). Gabapentin, a modulator of GABA biosynthesis-related enzymes (i.e., glutamic acid decarboxylase [GAD], and branched chain aminotransferase [BCAT]), has been used as tinnitus treatment in animals and patients (Bauer and Brozoski 2001; Chen and Yin 2012; Russell and Baloh 2009). However, one report revealed that thalamic GABAergic function was not altered in noise-induced tinnitus animals (Sametsky and others 2015).
As for clinical cases, alprazolam, a GABAA receptor agonist, was able to decrease the loudness of tinnitus in 76% of participants compared with 5% of placebo group (Johnson and others, 1993). Intravenous administration of furosemide, a noncompetitive subtype-specific GABAA receptor (e.g., α6β2γ2) antagonist, is reported to have reduced tinnitus severity in about 14% of tinnitus patients (Korpi and Luddens 1997; Risey and others 1995). Acamprosate, which is known to antagonize NMDA receptor-induced excitability, also acts on increasing GABA transmission. Its application to patients successfully relieved their tinnitus symptoms (Azevedo and Figueiredo 2005). In other clinical cases related with GABA transmission, it was found that gabapentin is largely ineffective in treating tinnitus (Bakhshaee and others 2008; Bauer and Brozoski 2006; Piccirillo and others 2007; Witsell and others 2007). When combined with intradermal administration of lidocaine, a sodium channel antagonist, however, gabapentin alleviated tinnitus symptoms up to about 6 months, compared with those that received gabapentin alone or placebo, suggesting that targeting more than one ion channel can be effective in tinnitus treatments (Ciodaro and others 2015). So far, it is not clear how gabapentin can therapeutically help tinnitus due to its multivariate and unknown actions in neurons. Nonetheless, there is general consensus that augmenting GABA receptor–mediated inhibition is crucial to decrease hyperexcitability involved in tinnitus (Fig. 2).
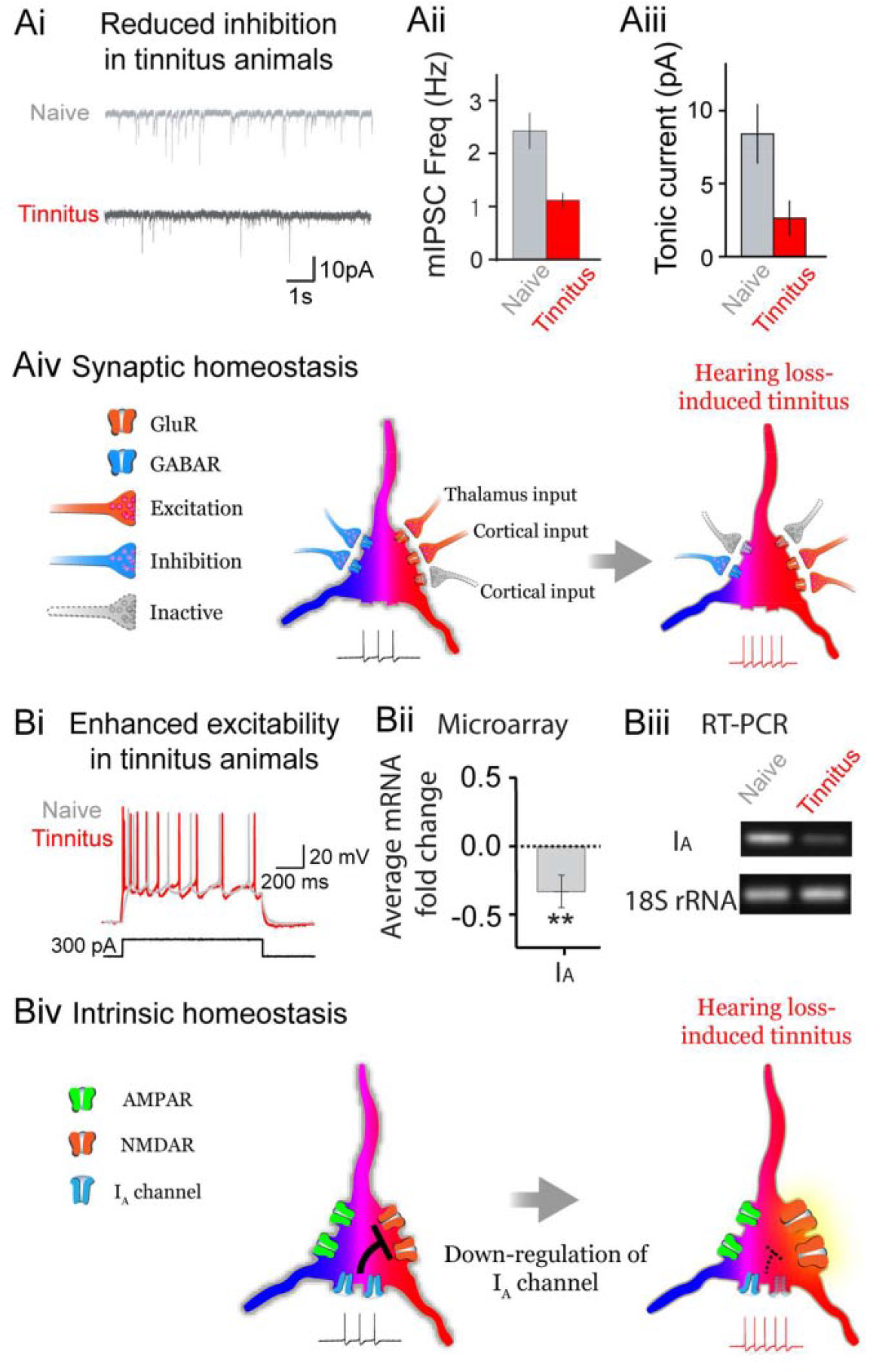
Homeostatic regulation of ion channels and receptors in hearing loss-induced tinnitus. (A) Hearing loss reduces γ-aminobutyric acid receptor (GABAR)–mediated phasic (Ai and Aii) and tonic (Aiii) inhibition, which is tightly related to tinnitus manifestation, as measured in miniature inhibitory postsynaptic currents and charge transfer, respectively. This figure is adapted and modified from Yang and others 2011. A schematic of synaptic homeostasis shows reduced inhibition, but not excitation which is presumably compensated from intercortical inputs. Note enlarged reddish color of a neuron indicates hyperexcitation in hearing loss-induced tinnitus. (B) Hearing loss increases intrinsic excitability by down-regulating IA channels (Bii and Biii). In our preliminary data, Kv1.4 (a A-type potassium channel) gene expression of the auditory cortex of tinnitus animals is down-regulated in both microarray and reverse transcription–polymerase chain reaction (rt-PCR) data. A schematic of intrinsic homeostasis shows exaggerated N-methyl-
Glycine Receptors (GlyRs)
GlyRs are known to gate chloride ions and mediate fast inhibitory neurotransmission mostly in the spinal cord, brainstem, and midbrain (Avila and others 2013; Betz 1992; Nicoll and others 1990; Probst and others 1986). However, the activity of GlyRs in the brain is also implicated in tinnitus.
Decreased GlyR α1 protein levels were observed in DCN of noise-induced tinnitus rats (Wang and others 2009). It is particularly of interest that a chemical, zinc ion, known to modulate GlyRs, could inhibit spontaneous firing of fusiform cells in mouse DCN (Perez-Rosello and others 2015). It should be noted that some studies suggested that zinc is ineffective except in some patients with zinc deficiency (Arda and others 2003; Coelho and others 2013; Person and others 2016; Yetiser and others 2002). Taurine, which activates both glycine and GABA receptors, was used to palliate tinnitus behavior in rats; currently, it is not clear that the action of taurine on tinnitus is through GABA, glycine receptors or both (Brozoski and others 2010). Nonetheless, these results substantiate the notion that inhibition decrement is correlated with tinnitus. GlyRs, though being expressed mostly in lower brain areas, are believed to one key component for tinnitus development.
Voltage-Gated Potassium Ion (K+) Channel
Voltage-gated K+ channels play a key role in dampening neuronal excitability and returning membrane potential to resting condition (Hille 2001). They have been reported to regulate neuronal excitability while their malfunction is involved in many types of brain diseases such as epilepsy, long-QT syndromes, ataxia/myokymia, Bartter’s syndrome, neuromuscular disorders, hearing disorder, and vestibular diseases (Bernard and others, 2004; Shieh and others, 2000).
Mutation in KCNE1 (a voltage-gated potassium channel, Isk-related family member 1) was suggested to predispose humans to tinnitus development; however, such observation was not concrete because the finding could not be replicated in different patient populations (Pawełczyk and others 2012). Similarly, altered expression level of KCNE3 in tinnitus patients could not reliably predict the risk of developing tinnitus due to small population size (Sand and others 2011). In noise-induced animal models, there is a decrease in KCNQ2/3 (a slow delayed rectifier of voltage-gated potassium channel) activity and accordingly an increase in neuronal activity of the DCN fusiform cells, both of which lead to increased susceptibility to tinnitus development (Li and others 2013). Consistent with this finding, retigabine, a small molecule that activates KCNQ channels and its variant (SF0034) prevented tinnitus development by suppressing noise-induced hyperexcitability in DCN neurons (Kalappa and others 2015). Application of retigabine reduced the membrane potential of DCN cells to more resting state, which conferred protection from developing tinnitus in acoustic trauma-induced mice (Li and others 2013). Also, salicylate-induced tinnitus behavior in rats was successfully silenced by application of BMS-204352 Maxipost and its R-enantiomer (R-Maxipost), both positive modulators of KCNQ ion channels (Lobarinas and others 2011). Collectively, these studies converge on to the notion that regulators of potassium ion channels are effective therapeutic candidates for tinnitus. These studies draw attention to the DCN as a possible site for tinnitus induction.
Calcium Ion–Activated Potassium (BK) Channels
BK channels, which are activated by both voltage change and Ca2+ ions, put a check on neuronal excitability and neurotransmitter release (Zheng and Trudeau 2015). They mostly localize to the dendrites and axons of neurons including cochlear hair cells (Hafidi and others 2005; Lee and Cui 2010).
Maxipost and R-Maxipost, activators of BK as well as KCNQ channels, were used to effectively diminish salicylate-induced tinnitus behavior in rats; nonetheless, it is unclear whether the tinnitus alleviation was from the activation of KCNQ, BK channels or both (Hewawasam and others 2002; Lobarinas and others 2011). It is possible that BK channels contribute to restoration of depolarized neurons back to the resting state, thereby alleviating tinnitus. For now, further clinical validation is required.
Hyperpolarization-Activated Cyclic Nucleotide-Gated Channels (HCN Channels)
HCN channels (or Ih) permit the passage of non-specific cations of Na+, Ca2+, and K+ and are abundantly expressed in most neurons (Benarroch 2013). Activated by hyperpolarization, HCN channels re-depolarize neurons, thereby bringing them close to threshold for spiking. They contribute to regulating neuronal excitability and synaptic transmission along the auditory pathway (Benarroch 2013; Kase and Imoto 2012; Kim and Holt 2013). It is also well-known that Ih enhances time-locking ability by shortening the cell’s time constant and reducing temporal width of spikes, eventually improving temporal precision of signals (Bal and Oertel 2000; Koch and Grothe 2003; Yang and Feng 2007; Yang and others 2009). Malfunction of these channels is involved in several brain diseases such as epilepsy, Parkinson’s disease, and neuropathic pain (Adams and others 2009; DiFrancesco and DiFrancesco 2015; Jung and others 2007; Yang and Cox 2008).
The involvement of HCN channels in tinnitus was observed in only one recent animal study (Li and others 2015). A reduction in HCN channel activity was found in the DCN of noise-induced tinnitus mouse. It is of interest that such reduction conferred resistance to tinnitus development. The result draws attention to the fact that HCN channels are involved in homeostatic compensation to hyperexcitability that would occur only after acoustic trauma. Further research into HCN channels is required to pinpoint their precise roles in processing intrinsic and extrinsic auditory signals.
Voltage-Gated Calcium Ion (Ca2+) Channels
Voltage-gated Ca2+ channels are localized to different compartments in a neuron and play different roles in regulating synaptic and firing activity. T- and L-type calcium channels are localized in dendrite and soma, while N- and P/Q-type calcium channels are at synaptic terminals (Dolphin 2012). It is also known that the T- and L-type calcium channels are responsible for subthreshold depolarization (or oscillations) and gene transcription whereas N- and P/Q-type calcium channels are critical for neurotransmitter release (Dolphin 2012; Hille 2001). Accordingly, dysfunction in these channels leads to various neurological disorders such as pain, epilepsy, ataxia, and psychiatric disorders (Frank 2014; Simms and Zamponi 2014).
Nimodipine, an L-type calcium channel antagonist, partially alleviated tinnitus of patients; improvement was observed in some patients, but worsening was in others after nimodipine treatment (Davies and others 1994; Elgoyhen and Langguth 2010). As described earlier (see GABA receptors), gabapentin. which is known to regulate GABA biosynthesis, had some positive effects on tinnitus alleviation. It is notable that the Gabapentin may also act via modulating calcium channels as it has been reported to bind to voltage-gated calcium ion channels and NMDA receptors, which are also calcium-permeable. It would be important to further investigate how different types of calcium channels regulate neuronal excitability and gene regulation of sound processing and tinnitus.
Voltage-Gated Sodium Ion (Na+) Channel
Voltage-gated Na+ channels are responsible for generation and propagation of action potentials in neurons (Catterall 2012). Because of this, they have been usually a major target of pharmacological therapy (Dib-Hajj and others 2015; Istvan and others 2007; Persson and others 2016; Zheng and Trudeau 2015).
Sodium valproate, a drug which blocks both voltage-gated sodium and T-type calcium channels, alleviated tinnitus symptoms in humans (Menkes and Larson 1998). However, another study reported that this drug triggered tinnitus during the treatment period (Hori and others 2003; Reeves and others 2000). Lidocaine, an antagonist of voltage-gated Na+ channels, has been also used for tinnitus research (Onizuka and others 2004). When administered intradermally to tinnitus patients, it caused substantial improvement in the loudness and level of tinnitus (Savastano 2004). Intradermal drug administration of lidocaine had less adverse effects than intravenous (Israel and others, 1982; Staffen and others 1999) and transtympanical drug (Coles and others 1992) administration. However, the use of Na+ channel blockers for tinnitus is problematic because it disturbs normal cells’ excitability broadly throughout the brain. Further research on the dose-dependency of these drugs should be considered to avoid such a problem.
Perspective
Most drugs currently used for tinnitus treatment directly modulate ion channels and receptors (Table 1). However, the fact that those drugs act on a global scale over whole brain areas leads to discontinuation of pharmaceutical treatment due to nonspecific side effects (Langguth and others 2009; Salvi and others 2009). For example, drugs that serve as antagonists of NMDA receptors and voltage sodium channels for alleviating tinnitus can cause serious disruption of normal physiological functions along both auditory and nonauditory areas. Also, drugs that activate GABA receptors have emotionally sedating and even suicidal effects in tinnitus patients, leading to negative consequences on quality of life. One way to circumvent these problems is to consider methods of administration of therapeutic drugs locally. Local drug delivery to suppress tinnitus could lead to better clinical outcomes, avoiding aberrant brain activity in different regions. Additionally, the location and mechanisms of tinnitus could be readily identified if local drug delivery is available. A second consideration is to identify optimized drug concentration. An optimized concentration can be effective only in the affected auditory areas even when administered globally in the brain. Last, it would be beneficial to apply a drug that indirectly modulates ion channels/receptors which are essential for normal excitability of neurons (e.g., Na+ channels, NMDA and GABA receptors). Our group previously found that noise-induced tinnitus animals exhibit intrinsic hyperexcitability (Yang and others 2012). If tinnitus is related to exaggerated activity of NMDA receptors, it would be beneficial for tinnitus alleviation to employ an enhancer of A-type potassium channels (or IA). It is because IA is known to be activated largely in a hyperexcitable condition of NMDA receptors, both of which have been known to play a counterbalancing role in regulating neuronal excitability (Yang and others 2014; Yang and others 2015). Furthermore, IA was, in our preliminary data, downregulated in tinnitus animals and accordingly its activation can potentially alleviate tinnitus by suppressing intrinsic hyperexcitability, but does not severely affect normal signal transmission and membrane excitability (Fig. 2). Thus, the indirect modulators of physiologically essential channels/receptors should be considered to design therapeutic target of tinnitus. As mentioned above (Glutamate Receptors section), drugs that can indirectly modulate the target receptors should also be considered as viable treatments of tinnitus. Those strategies might not interfere with the normal physiological function of individuals, therefore having long-term clinical benefits.
Tinnitus, such as the condition caused by hearing loss, induces widespread changes in expression and function of ion channels and receptors along the auditory pathway. These changes can be the primary effect of the hearing loss, or a secondary effect caused by homeostatic compensation executed by the various neurons in the system. Gene expression of ion channels is a potent way to modify the membrane composition of ion channels and receptors in a given neuron, and hence its excitability and E/I balance (Fig. 3). Many molecules are involved in the genetic regulation of ion channels and receptors: transcription factors of the immediate-early gene Arc/Arg3.1, secreted factors such as retinoids, TGF-β, BDNF, and glia cell-derived TNF-α, and signaling molecules such as CaMKIIα and β (Aoto and others 2008; Pozo and Goda 2010; Shepherd and others 2006; Stellwagen and Malenka 2006; Sweeney and Davis 2002; Thiagarajan and others 2002). Temporary alteration of E/I balance by drugs can be quickly nullified due to neuron’s capability to continuously compensate for these changes (i.e., homeostatic regulation of neural circuits), when drugs are washed away or drug effect is desensitized. This hypothesis explains why most drugs that either modulate excitatory or inhibitory level alone have not been successful (Langguth and others 2009; Salvi and others 2009). Thus, blocking or activating molecules related to homeostatic plasticity can be a plausible approach to fundamentally cure brain diseases. Research that incorporates modern genetic analysis tools such as gene knockout technology or CRISPR/Cas9 gene modification is required to delineate the precise contributions of genes to tinnitus development. Understanding how modulators of ion channels and receptors induce plasticity and alter sound processing will allow us to better select for drug treatments for this debilitating disorder.
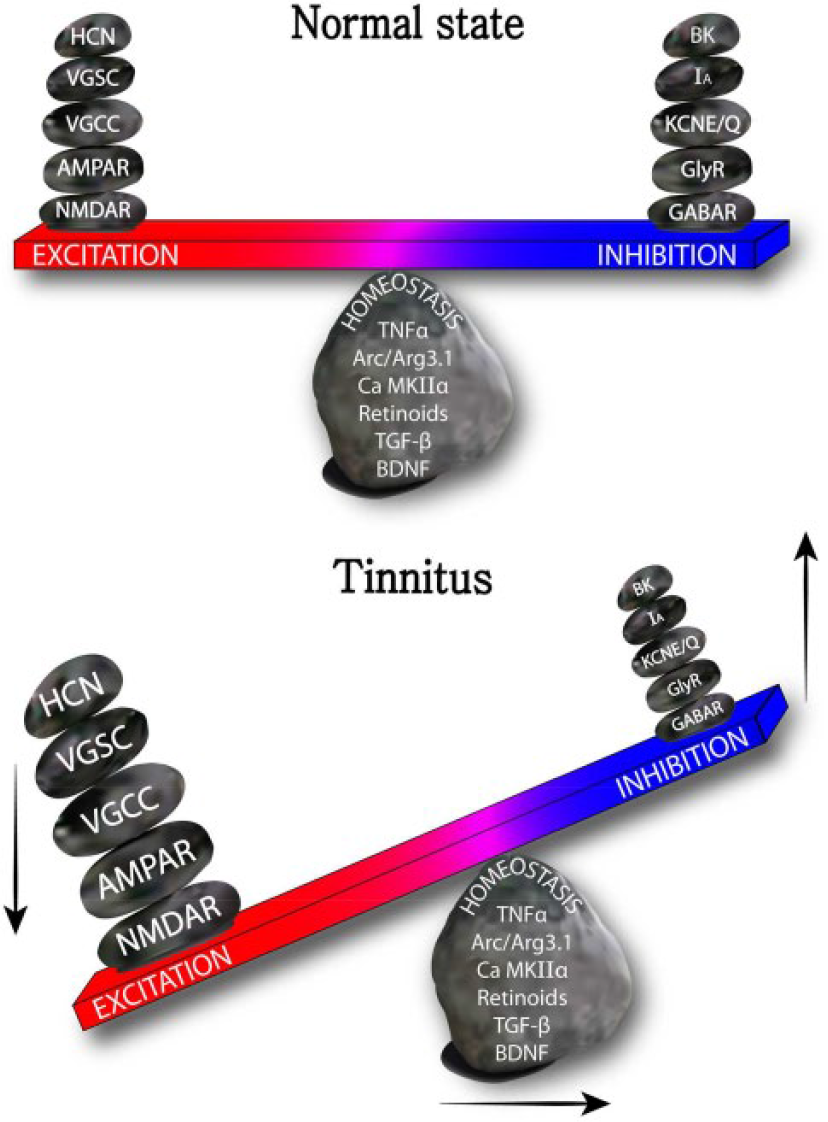
A seesaw model of tinnitus channelopathy. Environmental (e.g., auditory trauma) and genetic (e.g., overexpression of genes susceptible to tinnitus) changes cause action of homeostatic molecules (retinoids, Arc/Arg3.1, CaMKIIα and β, TGF-β, TNF-α, and BDNF) to be shifted into increased E/I balance. NMDAR, N-methyl-
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2016R1D1A1B04930938) and an Incheon National University (International Cooperative) Research Grant for Dr. Sunggu Yang, an internal startup grant from the City University of Hong Kong and the Centre for Biosystems, Neuroscience, and Nanotechnology for C. Geoffrey Lau, and Food and Health Bureau of Hong Kong Special Administrative Region Government (9211132), GRF grants from the Research Grant Council of the Hong Kong Special Administrative Region Government (9048067) for Sungchil Yang.