
Abstract
Models advanced to explain hemispheric asymmetries in representation of emotions will be discussed following their historical progression. First, the clinical observations that have suggested a general dominance of the right hemisphere for all kinds of emotions will be reviewed. Then the experimental investigations that have led to proposal of a different hemispheric specialization for positive versus negative emotions (valence hypothesis) or, alternatively, for approach versus avoidance tendencies (motivational hypothesis) will be surveyed. The discussion of these general models will be followed by a review of recent studies which have documented laterality effects within specific brain structures, known to play a critical role in different components of emotions, namely the amygdata in the computation of emotionally laden stimuli, the ventromedial prefrontal cortex in the integration between cognition and emotion and in the control of impulsive reactions and the anterior insula in the conscious experience of emotion. Results of these recent investigations support and provide an updated integrated version of early models assuming a general right hemisphere dominance for all kinds of emotions.
Keywords
The First Descriptions and Interpretations of a Different Emotional Behavior in Patients with Right and Left Hemispheric Lesions
The hypothesis assuming that hemispheric asymmetries may exist in the representation of emotions is recent and controversial. It is recent, because a century elapsed between the first studies showing a left hemisphere (LH) dominance for language (Broca 1865; Dax 1865) and the first investigations showing laterality effects in the representation of emotions (Gainotti 1969, 1972; Rossi and Rosadini 1967; Terzian and Cecotto 1959). It was, and remains, controversial because different models of emotional lateralization were advanced, since these pioneering investigations, to clarify the meaning of the observed asymmetries. Terzian and Cecotto (1959) and Rossi and Rosadini (1967), who first described a different emotional behavior in patients submitted to pharmacological inactivation of the right and LH, relied on the model of the bipolar manic depressive disorders to explain their observations. They reported that injection of sodium amytal into the left carotid artery produces a “depressive-catastrophic reaction” (characterized by a sad attitude with bursts of tears), whereas inactivation of the right hemisphere (RH) is followed by a “euphoric-maniacal reaction” and attributed these different emotional reactions to disruption of neural mechanisms underpinning opposite aspects of mood. Depressive-catastrophic reactions were attributed to inactivation of a center for positive emotions located in the LH and euphoric-maniacal reactions to disruption of a center for negative emotions located in the RH. On the other hand, Gainotti (1969, 1972), who some years later made partly similar observations in right and left brain-damaged patients, describing “catastrophic reactions” in (aphasic) left, and “indifference reactions” in severe right brain–damaged patients, refuted the equivalence between catastrophic reactions and biological depression and between indifference reactions and euphoric-maniacal states. He considered catastrophic reactions as a dramatic, but psychologically appropriate form of response to a catastrophical event, and the indifference reaction of patients with severe RH lesions as an abnormal or inappropriate reaction to a dramatic event. To explain the contrast between the different emotional behavior of right brain– and left brain–damaged patients, Gainotti (1972) advanced the hypothesis that the RH may be dominant for emotion, just as the LH is dominant for language and that the emotional reaction may be inappropriate in right brain–damaged patients if the lesion disrupted structures crucially involved in emotional processing. Thus, the first clinical observations that have raised the problem of hemispheric asymmetries in emotion representation have also prompted the two models of emotional lateralization that are still considered as the most important ones in this area of inquiry and which can be labeled as “the right hemisphere” hypothesis and “the different hemispheric specialization” (or “the valence”) hypothesis. The first model supposes a general dominance of the RH for every kind of emotional response, regardless of affective valence, whereas the second model assumes an opposite dominance of the LH for positive and of the RH for negative emotions.
Models of Human Emotions that Have Oriented Investigations of Emotional Laterality in the Human Brain
The general model of emotions to which most subsequent investigations of emotional laterality in the human brain made reference was the Ekman’s (1977, 1984) model. This model, drawing on the Darwin’s (1872) claim that facial expression of emotions is part of man’s biological inheritance, maintained that the human expressive mimic reactions are automatically selected from a small number of innate operative patterns, corresponding to the basic emotions (happiness and sadness, fear and anger, surprise and disgust). This theoretical framework was greatly expanded by other authors (e.g., Frijda 1986; Izard 1977; LeDoux 1996; Panksepp 1989; Plutchik 1980) who considered emotion as a multicomponent adaptive system, whose functional architecture has developed across phylogeny, spanning from very primitive, hard-wired survival-related behavioral schemata to much more complex and learned social patterns, integrated with the cognitive system.
Relationships Between the Emotional and the Cognitive System
Oatley and Johnson-Laird (1987) maintained that the organism disposes of two operative systems to face a partially unpredictable environment: (1) the emotional system, considered as an emergency system, able to interrupt the ongoing activity to rapidly select a new operative scheme and (2) the cognitive system, viewed as a more complex and evolved system, but requiring much more time to accomplish its work. Both structural similarities and functional differences exist between emotional and cognitive system. The structural similarities stem from the fact that both systems base their activity on the integrated work of components that must (1) analyze the sensory data to compute their significance, (2) select the most suitable responses, and (3) put all this information into appropriate memory systems. The functional differences (which derive from the logic of the two systems and concern the manner in which each system deals with sensory information and selects specific action schemata) are reported in Box 1.
Common aspects and distinguishing features of the emotional and cognitive systems.
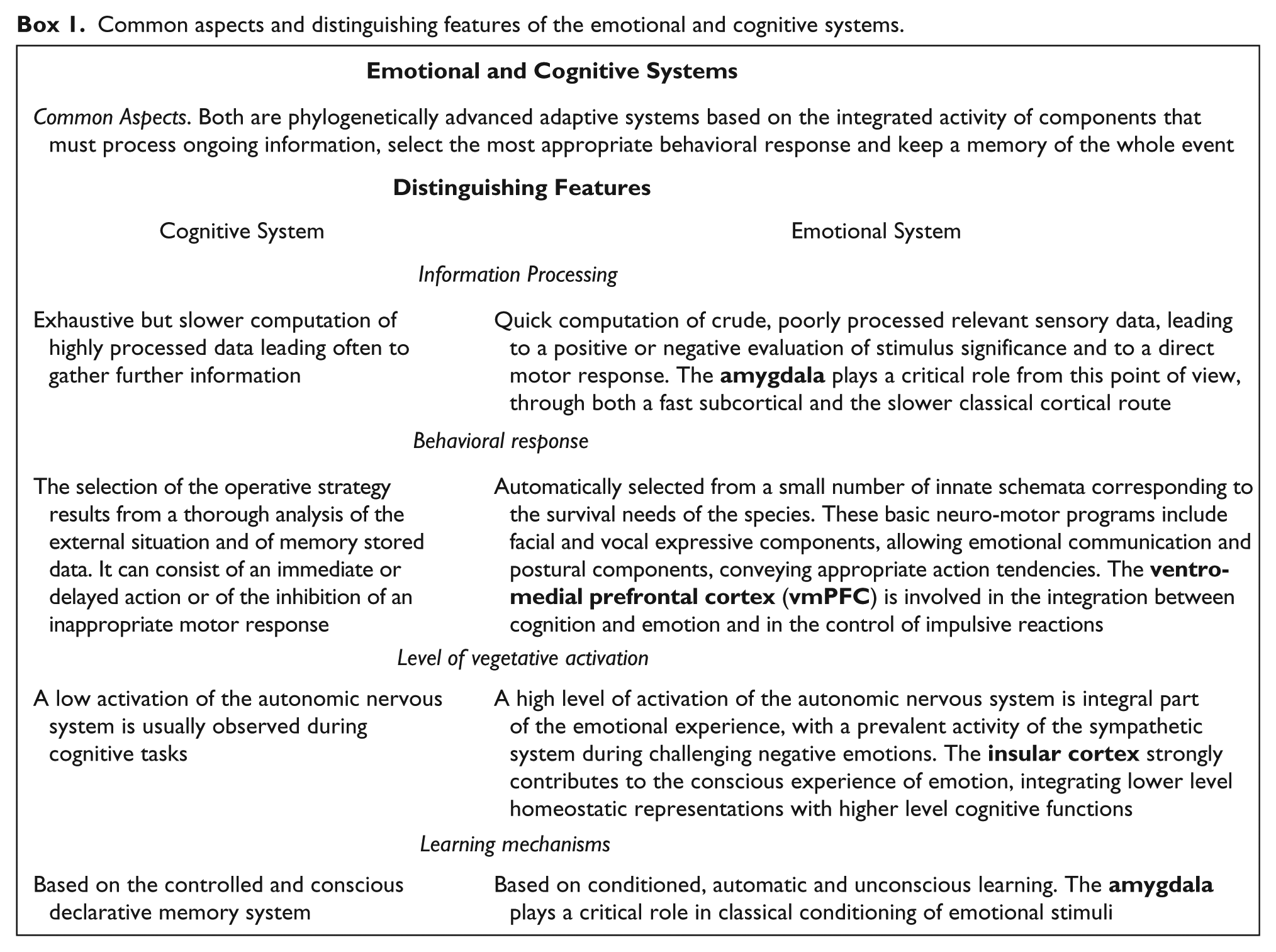
In addition to describing the typical features of the emotional system in these different functions, in Box 1 are also reported the brain structures that play a critical role in each of them, namely, the amygdala in the quick and raw computation of sensory data, the ventro-medial prefrontal cortex (vmPFC) in the integration between cognition and emotion and the control of inappropriate emotional reactions, and the insular cortex in providing a substrate to the conscious experience of emotion. In later sections of the present review, laterality effects within each of these brain structures will be taken separately into account, to evaluate their contribution to hemispheric asymmetries in emotion processing and representation.
Hierarchical Organization of the Emotional System
The acknowledgment that emotional and cognitive systems become more and more interconnected during the development prompted the construction of hierarchically organized developmental models of emotions, that aimed to explain how complex emotions can be formed starting from the simplest ones and how the highest components keep under control the lowest parts of the emotional system. In particular, Leventhal (1974, 1987) proposed a developmental model which assumed that human emotions may be based on three functional levels: (1) the sensorimotor, (2) the schematic, and (3) the conceptual level. The sensorimotor level consists of a set of innate, universal expressive-motor programs, which are triggered automatically by a certain number of stimuli and include components of motor and autonomic activation, as well as the corresponding subjective emotional feelings. The schematic level results by the association, during the individual development, of these innate programs with situations of individual experience. This leads to the construction of “emotional schemata,” which are automatically elicited, correspond to spontaneous emotions and are accompanied by subjective feelings, which are the hallmarks of a true emotion. The conceptual level, which is the last stage of this model, is based on mechanisms of conscious declarative memory and does not store instances of concrete emotional experiences, but abstract notions about emotions and about the social rules allowing their expression.
Early Experimental Investigations that Have Evaluated Comprehension or Expression of Positive and Negative Emotions at the Facial or Vocal Level
In the years following the first descriptions of a different emotional behavior in patients with right and left hemispheric lesions, several experimental investigations were conducted in healthy subjects and in patients with unilateral brain injury, to clarify the meaning of the different emotional reactions observed in unilateral brain-damaged patients. Since the easiest manner of studying positive and negative emotions consists in contrasting their expression at the facial or vocal level, these investigations used methodologies that evaluated comprehension or production of positive and negative emotions through facial expression or vocal emotional prosody. In investigations conducted on normal subjects, the monohemispheric treatment of emotional information was studied both in the visual modality (lateralized tachistoscopic presentation) and in the auditory modality (dichotic listening or lateralized auditory presentation), whereas hemispheric asymmetries on the expressive side were studied primarily by comparing the emotional expressiveness of the two hemifaces. Box 2 reports some data about methodology and results obtained in these studies.
Methodology and results of investigations that have studied comprehension and expression of emotions in normal subjects through the affective tone of voice or facial emotional expressions.

Data reported in Box 2 show that comprehension and expression of emotions through face and voice are lateralized to a great extent in the RH, regardless of valence. Consistent with data obtained in normal subjects were results of investigations which studied comprehension and expression of positive and negative emotions in patients with unilateral brain lesions, comparing the ability of right brain– and left brain–damaged patients to recognize or express emotions both with voice (emotional prosody) and with facial expression. Some authors did, indeed, show that right brain–damaged patients can understand the meaning of a sentence, but are unable to recognise the emotion expressed by the tone of voice of the speaker (see Ross 1981, 1984; Gainotti 2000, 2001; and Ross and Monnot 2007 for reviews). Other authors showed that patients with RH damage are seriously impaired in recognizing facial emotional expressions (see Mandal and others 1993; Gainotti 2001, 2005; and Borod and others 2002 for reviewes). Similar results were obtained studying the capacity of right brain– and left brain–damaged patients to express emotions through the tone of voice (see Ross 1981, 1984; Gainotti 2000, 2001; Ross and Monnot 2007 for reviews) and through facial emotional expression (see Borod and others 1986; Mandal and Ambady 2004; and Gainotti, 2001, 2005 for reviews). Altough most data obtained in normal subjects and in unilateral brain-damaged patients strongly support the hypothesis of a general RH dominance for emotional comprehension and expression, results of some investigations conducted in normal subjects with different experimental procedures (and also reported in Box 2) could support the hypothesis of a different specialization of the RH for negative emotions and of the LH for positive emotions. Thus, Reuter-Lorenz and Davidson (1981), Reuter-Lorenz and others (1983), Natale and others (1983), and Rodway and others (2003) documented a RH involvement in comprehension of negative emotion and a LH involvement in comprehension of positive emotions, whereas Sackeim and Gur (1978), Heller and Levy (1981), Sackeim and Grega (1987), Schiff and Lamon (1989), and Mandal and Singh (1990) found a greater left-sided intensity in facial expression of emotions only for negative, but not for positive emotions.
Models of Emotional Laterality Prompted by the Aforementioned Studies of Comprehension and Expression of Positive and Negative Emotions
Three main models of emotional laterality came from the aforementioned studies of comprehension and expression of positive and negative emotions at the facial or vocal level. The first model stressed the RH superiority in comprehension and expression of emotions, assuming that the RH dominance for emotions may basically concern their communicative aspects. The second model emphasized the greater hemiface asymmetry for negative with regard to positive emotions. The third model assumed that hemispheric asymmetries observed at the level of the frontal lobes during positive and negative emotions, may not be related to the valence of the emotional stimuli but rather to the motivational (approach vs. avoidance) systems engaged by these stimuli. Each of these models will be separately taken into account in the present review.
The Model Assuming that the Right Hemisphere Dominance for Emotions May Only Concern the Communicative Aspects of Emotions
This model was proposed by Ross (1981, 1984), who maintained that disorders of emotional communication are the primary defect resulting from RH damage and that other emotional disturbances of right brain–damaged patients are only a consequence of this basic inability to comprehend and express emotions. This viewpoint was, however, at variance with results obtained by Wittling (1990) in healthy subjects and by Heilman and others (1978), Morrow and others (1981), and Zoccolotti and others (1982, 1986) in brain-damaged patients. Wittling (1990) studied blood pressure changes during lateralized presentation of an emotional film and showed that RH film presentation caused a significantly higher increase in systolic and diastolic pressure than LH viewing of the same film. In a similar vein, Heilman and others (1978), Morrow and others (1981), and Zoccolotti and others (1982, 1986) showed that right brain–damaged patients present not only disorders of emotional communication but also a flattened galvanic skin response (GSR) to painful or distressing emotional stimuli. Taken together, these data show that right brain damage disrupts not only the communicative aspects but also the autonomic components of emotions.
The Greater Hemiface Asymmetry for Negative Emotions and the Social Use of Smile and Other Positive Facial Expressions
Etcoff (1986) noted that smiling differs from other emotional facial expressions not only because of its positive emotional polarity, but also because it represents the most easy “emotional” facial expression to reproduce voluntarily and the most currently used for approach and social communication. She, therefore, claimed that, in the case of smiling, a LH dominance for the intentional control of the facial expressive apparatus could counterbalance the greater left half-face expressiveness, resulting from the general RH prevalence for the spontaneous expression of emotions. A partly similar interpretation was advanced by Gainotti and others (1993), drawing on the distinction proposed by Leventhal (1974, 1987) between “schematic” and “conceptual” levels of emotions. These authors suggested that the RH might be involved preferentially in the spontaneous expression of emotions, whereas the left frontal lobe might be mainly involved in the intentional control of the emotional expressive apparatus. Results consistent with this line of thought were recently obtained by Ross and Pulusu (2013) contrasting spontaneous with posed facial emotional expressions, because these authors showed that spontaneous (schematic) expressions are usually initiated on the left side of the face, whereas posed expressions (linked to social communication and associated display rules) are initiated on the right side of the face.
The Davidson Hypothesis Assuming that Frontal Lobe Asymmetries May Not Be Related to Emotional Valence but to the Motivational Systems Engaged by the Stimuli
Drawing on electroencephalography (EEG) asymmetries observed by Davidson and others (1979) and by Davidson and Fox (1982) at the level of the frontal lobes during positive and negative emotions, Davidson (1983, 1992, 1998) claimed that these asymmetries were not related to the valence of the emotional stimuli but to the motivational systems engaged by these stimuli. This author proposed that the left prefrontal cortex (PFC) may be involved in a system facilitating approach to appetitive stimuli, whereas the right PFC may be involved in a system facilitating withdrawal from aversive stimuli. According to Davidson (1992), the arousal of approach-related emotions should be associated with activation of the left, and arousal of withdrawal-related emotions with activation of the right frontal region. Even if after the 1990 the measure of the frontal EEG asymmetries and the motivational approach-avoidance model became extremely popular, pruducing hundreds of research articles, both the relation between PFC and approach/avoidance and the use of frontal EEG asymmetries to assess emotion or motivation have been the object of documented critical reviews (e.g., Spielberg and others 2008 for the first issue and Allen and others 2018 and Reznik and Allen, 2018 for the second methodological problem). Furthermore, some authors have challenged the equivalence approach—positive and avoidance—negative emotions, stressing the fact that anger, which is a negative emotion (Russell and Barrett 1999), often evokes an approach motivation and usually coincides with activation of the left hemisphere (Carver and Harmon-Jones 2009). More in general, it must be acknowledged that, even if this model has been very influential, it can hardly explain results of the experimental and clinical studies reported in Box 2 of the present review for at least two reasons. The first is that the difference between positive and negative emotions studied at the expressive level in the aforementioned investigations is only in part captured by the distintion between approach and withdrawal tendencies proposed by Davidson (1983, 1992, 1998). The second reason is that the aforementioned studies have shown that the RH dominance concerns not only the expression of emotions, subserved by the anterior frontal regions, but also the recognition of emotionally laden information, subserved by posterior cerebral cortices. Instead, the approach/ withdrawal model only concerns the experience and expression of emotions at the level of the PFC. A general dominance of the RH in the processing of emotional information has, indeed, been acknowledged by Davidson (1983), who suggested an interaction between the right/left and the anterior/posterior dichotomy in the regulation of human emotions. This interactive account assumed an overall dominance of the right parietal region for the treatment of emotional information and a diverging specialization of the right and left frontal lobes for the expression of negative and positive emotions. However, if this model was correct, we should predict that the interaction between valence and laterality should be greater at the expressive than at the receptive level, whereas data reported in Box 2 show a mild trend in the opposite direction, istead of supporting this prediction.
Perplexities about Existence and Nature of a Lateralization of Emotional Functions Drawn from Functional Neuroimaging Studies of the Neural Correlates of Emotions
Data relevant to the existence and nature of a lateralization of emotional functions can be drawn from the study, using positron emission tomography (PET) and functional magnetic resonance imaging (fMRI), of the activation produced in various brain structures of healthy subjects by experimental paradigms assessing different facets of emotions. Several authors, therefore, explored the neuroanatomical substrates of emotions using different emotional categories, experimental paradigms and kinds of stimuli. To give an overall survey of these studies, Wager and others (2003) performed a quantitative meta-analysis on 65 neuroimaging studies of emotion focusing on the effects of emotional valence on regional brain activations, with particular emphasis on hypotheses concerning lateralization of brain function in emotion. These authors found no support for the hypothesis of overall right-lateralization of emotional function, and limited support for valence-specific lateralization of emotional activity in frontal cortex. Similar results were obtained by Fusar-Poli and others (2009) in a voxel-based meta-analysis of studies employing emotional faces paradigms in healthy subjects, because this study too failed to support both the “right hemisphere” and the “valence” hypothesis. These rather disappointing results must be considered with caution, because they can be due to several factors concerning both methodological and experimental reasons. For instance, constraints and limits imposed by functional imaging methodologies constitute a far from ideal environment for studying full-blown emotional processes. Thus, Levenson and others (2014) noticed that, even if it were possible to induce powerful emotions in the scanner, the attendant muscular activity in body and face would produce huge signal artifacts. As a result, a great deal of emotional research conducted in the scanner falls into the domain of emotional cognition (where the focus is on how we think about emotions and make judgments about them), rather than the actual processes as they unfold in real time. More interesting than results obtained on heterogeneous whole brain activation studies must, therefore, be considered data obtained in anatomo-clinical or activation studies of brain structures, such as the amygdala, the vmPFC and the anterior insula, that, as shown previously and in Box 1 of this review, play very relevant roles in specific components of emotional functions. A diagram showing the functional interactions between these components of the emotional system is reported in Figure 1.
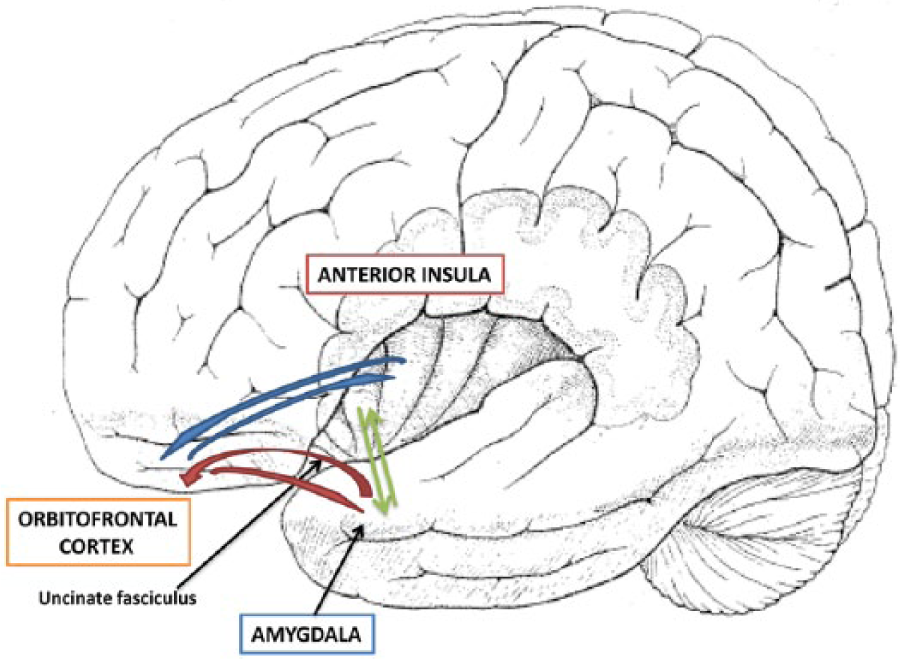
Schematic representation of the functional interactions existing between the main components of the emotional system. Bilateral connections exist between the amygdala (where emotional information is computed), the orbitofrontal cortex (where they are integrated with social rules allowing emotional expression), and the anterior insular cortex (which contributes to the conscious experience of emotions). The uncinate fasciculus connects the anterior portions of the temporal lobe (temporal pole and amygdala) with the inferior frontal gyrus, passing inward of the insular cortex and allowing a “topdown” modulation of intense emotional responses.
Investigations that Have Documented a Different Role of Right and Left Amygdala in the Evaluation of Emotional Stimuli
A very influential role, within the anatomo-clinical and activation studies that have proposed a different role of the right and left amygdala in emotional functions, was played by Morris and others (1998), who showed that stimuli processed below the level of awareness mainly activate the right amygdala, whereas consciously processed emotional stimuli preferentially activate the left amygdala. Trying to elucidate the neural mechanism that could subsume this dissociation, Morris and others (1999) documented a different functional connectivity of right and left colliculo-pulvinar pathways during processing of seen and unseen (masked) fear-conditioned faces. Increased connectivity between right amygdala, pulvinar, and superior colliculus was found when masked fear-conditioned faces were presented, whereas the left amygdala showed no masking-dependent changes in connectivity with superior colliculus or pulvinar. In line with this model, Wright and others (2003) and Gläscher and Adolphs (2003) proposed that the right amygdala might be involved in the rapid detection of emotional stimuli, while the left amygdala could play a role in the more elaborate stimulus evaluation. A selective right amygdala activation during unconscious processing of emotional stimuli was also reported by Nomura and others (2004) during cognitive evaluation of facial expressions primed by masked angry faces, because activity in the right amygdala was greater with subliminal presentation of the angry prime compared with subliminal presentation of a neutral face or white-blank stimuli. Williams and others (2006) also used fMRI in healthy human subjects to study the functional connectivity between amygdala and related cortical and subcortical structures during conscious and non-conscious perception of fearful facial expressions. Conscious perception was associated with negative connectivity but non-conscious perception with positive connectivity between superior colliculus, pulvinar and right amygdala. Hung and others (2010), studying with magnetoencephalography (MEG), the effect on event-related neural responses of unattended emotional faces, located in the left or right visual fields confirmed that that the amygdala processes threat-related information through a right fast subcortical route and a left slower cortical feedback mechanism (see Gainotti 2012a for a more detailed description of the aforementioned investigations).
Activation and Lesion Studies Documenting Asymmetries of Emotional Functions in the Ventro-medial Prefrontal Cortex
The right vmPFC may have a “general” role in emotional processing (see Drevets and Raichle 1998 for review), in the integration between cognition and emotion and in the control of impulsive reactions. As for the first point, Shamay-Tsoory and others (2003) claimed that the right vmPFC regulates the interaction of emotion and cognition in the production of empathic responses and the same authors (Shamay-Tsoory and others 2005) maintained that impaired “affective” theory of mind (TOM) is associated with right vmPFC damage. As for the second issue, Tranel and others (2002) demonstrated that patients with lesions of the right vmPFC show considerably more deficits in social, emotional, and decision-making domains than those with left-sided lesions. In a similar vein, Zald and Andreotti (2010) showed that damage to the vmPFC (especially in the right hemisphere) is connected with deficits in detecting irony, sarcasm, and deception, whereas Boes and others (2009) found a significant correlation between low impulse control and decreased right vmPFC volume in impulsive boys. These data are consistent with the the positions of Aron and others (2004 and 2014), who reviewed evidence suggesting that a sector of the right inferior frontal cortex (rIFC) implements a general inhibitory control via a wider prefrontal-basal ganglia network. All these data suggest a right lateralization of functions subsumed by the the ventral PFC, because this structure plays a critical role in the integration between cognition and emotion and in functions of “topdown” modulation of intense emotional responses, thanks to its connections with the amygdala through the uncinated fasciculus (UF), as is shown in Figure 1. Consistent with these views are data showing that connections between anterior temporal lobe (ATL) and orbito-frontal areas (through the UF) are stronger in the RH, while connections between ATL and inferior frontal gyrus (through the arcuate fasciculus), which have a key role in phonological processing are greater in the LH (Papinutto and others 2016). The relationships between right UF and and emotional functions were also documented by Coad and others (2017), who showed that in healthy subjects’ interindividual variability in the ability to decode facial emotion expressions is linked to the right hemisphere UF microstructure.
The Contribution of Right and Left Insular Cortex to Conscious Experience of Emotion
Since the insula is known to contain the primary gustatory cortex, earlier studies had focused on its special role in disgust (an emotion closely associated with the sensation of bad taste). In recent years, however, more emphasis has been placed on the insular contribution to conscious experience of emotion, because this experience depends on the perception of bodily reactions to emotion-provoking objects and the cognitive interpretation of these interoceptive informations. The insula receives interoceptive inputs from the whole body, and has bidirectional connections with the frontal, parietal, and temporal lobes, the anterior cingulate cortex and the amygdala. This connectivity pattern allows to integrate lower level homeostatic representations with higher level cognitive functions. Craig (2009, 2010, 2011) therefore proposed a posterior-to-anterior gradient in the insular cortex, in which physical features of interoception are processed in the posterior insula and the integration of interoception with cognitive and motivational information is made in the anterior insular cortex (AIC). A schematic representation of the integration of homeostatic and cognitive aspects of the conscious experience of emotions in the right AIC is reported in Figure 2.
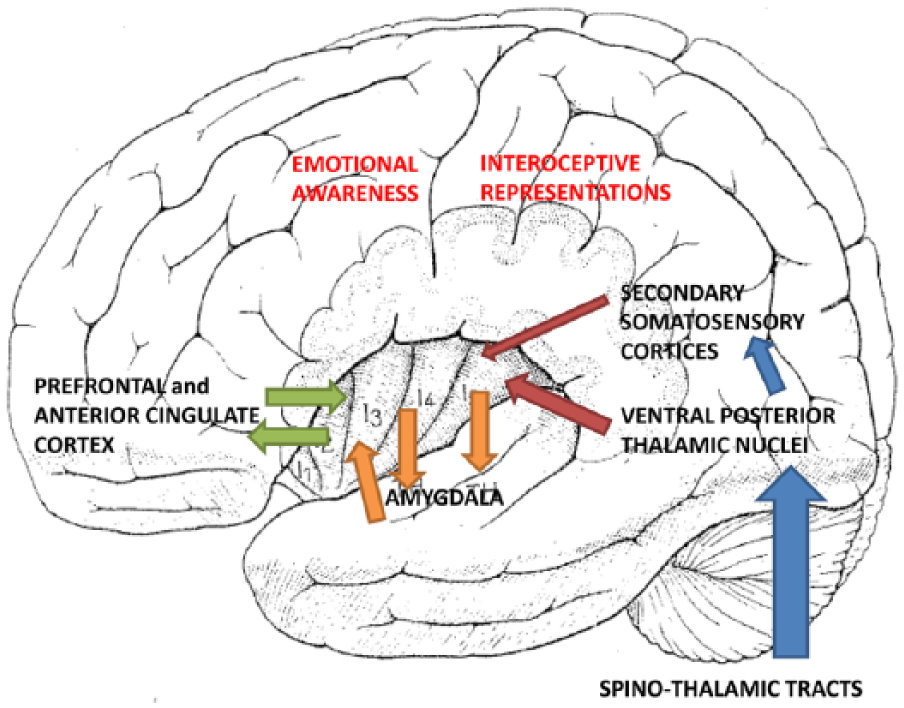
The figure shows in a schematic manner how interoceptive inputs coming from the whole body, through the spino-thalamic tracts, the ventral posterior thalamic nuclei and the secondary somatosensory cortices, reach the posterior insula providing first-order homeostatic representations. The figure also shows how the anterior insular cortex (and in particular the right anterior insula) thanks to its connections with the amygdala, the frontal cortices, and the anterior cingulate cortex, integrates lower level homeostatic representations with higher level cognitive functions and becomes the substrate of the conscious experience of emotions.
Craig (2005) also proposed a major contribution of the right AIC to the emotional experience and assumed that this hemispheric asymmetry may be based on an unequal representation of homeostatic activities, resulting from asymmetries in the peripheral autonomic nervous system. Results consistent with a greater role of the right insula in homeostatic representations were obtained by Critchley and others (2004), who measured regional brain activity by fMRI during an interoceptive task wherein subjects judged the timing of their own heartbeats. These authors observed that neural activity in right AIC predicted subjects’ accuracy in the heartbeat detection task and that greater AIC volume correlated with increased accuracy in this subjective sense of the inner body, and with the corresponding negative emotional experience. Their interpretation of these results was that the right AIC supports a representation of visceral responses accessible to awareness and provides a substrate for subjective feeling states. Results consistent with these views were obtained by by Gray and others (2007), who scanned participants while judging emotional face stimuli, during both exercise and non-exercise conditions in the context of true and false auditory feedback of tonic heart rate and by Xue and others (2010), who showed that the experience of taking (and especially winning) a gamble was associated with significantly stronger activation in the right AIC. From the neuroanatomical point of view, the greater role of the right AIC in emotional functions, cound be due to prevalence on this side of the von Economo neurons (VENs), which project from the frontoinsular (FI) cortex to frontal pole, septum, and amygdala, relaying to these structures information related to autonomic control or awareness of homeostatic representations. According to Allman and others (2010), these neurons are about 30% more numerous in the right than in the left FI cortex.
General Discussion
The aim of this discussion will consist in trying to summarize results obtained in various sections of this review with respect to the general problem of models advanced to explain the lateralization of emotional functions in the human brain and to see if results obtained in more recent studies can clarify those of more ancient investigations. The first clinical observations made on patients with focal brain damage by Gainotti (1972) and the first clinical and experimental studies which have investigated comprehension and expression of emotions at the facial or vocal level have generally suggested a greater involvement of the RH in emotional functions, irrespectively of their positive or negative valence (see Borod and others 1997, 2001; Gainotti 2001, 2005; and Mandal and Ambady 2004 for critical reviews). The hypothesis of a different lateralization of positive and negative emotions was supported only by a small number of experimental studies conducted in healthy subjects but became very influential when it was reframed in terms of approach versus withdrawal motivational tendencies. This hypothesis was advanced by Davidson (1983, 1992, 1998) relying on EEG asymmetries observed at the level of the frontal lobes during positive and negative emotions, but in recent years Allen and others (2018) and Reznik and Allen (2018) have raised strong methodological objections to the use of frontal EEG asymmetries to assess emotion or motivation. Furthermore, a careful inspection of data repported in in Box 2 shows that investigations which had studied in healthy subjects the expression of positive and negative emotions had not documented a greater involvement of the RH and LH in the production of negative and, respectively, of positive emotions, but only a greater engagement of the RH in the generation of negative, in comparison with positive, emotions. The observation of a greater expressivity of the left hemiface for negative emotions than for smiling or other positive emotions had, therefore, been interpreted by Gainotti and others (1993) and by Ross and Pulusu (2013) as related more to a greater involvement of the RH in the spontaneous expression of emotions and of the left frontal lobe in functions of emotional control than to the distinction between negative and positive emotions. Recent studies that have documented asymmetries of emotional functions in the ventro-medial PFC (Aron and others 2004, 2014; Boes and others 2009; Tranel and others 2002) are, however, at variance with this model, because they suggest a right lateralization of all emotional functions subsumed by these structures, including the “topdown” modulation of impulsive emotional reactions.
Other investigations that have tried to clarify the mechanisms underlying the processing of emotional information at the level of the right and left amygdala and the experience of emotions in the anterior insular cortex are also consistent with the model assuming a general dominance of the RH for various aspects of emotional functioning. Several functional neuroimaging experiments have, indeed, been pioneered by the influential articles of Morris and others (1998 and 1999) on conscious and unconscious forms of emotional learning in the human amygdala and by results of fMRI experiments conducted by Critchley and others (2004) and Gray and others (2007) to evaluate the contribution of the anterior insula to the conscious experience of emotions. These experiments have consistently shown: (1) that stimuli processed below the level of awareness mainly activate the right amygdala, whereas consciously processed emotional stimuli preferentially activate the left amygdala; (2) that the right amygdala activation during unconscious treatment of emotional stimuli is due to an increased connectivity between right amygdala, pulvinar, and superior colliculus whereas an involvement of this subcortical pathway is not observed when emotional stimuli are consciously processed; and (3) that the right AIC supports a representation of visceral responses accessible to awareness and provides a substrate for subjective emotional feelings. A model that according to Gainotti (2005, 2007a, 2007b) could explain the relationships between unconscious emotional processing and right amygdala activation through a subcortical visual pathway draws on the general account that many authors (e.g., Ekman 1984; Frijda 1986; LeDoux 1996; Panksepp 1989) have given of emotion, considered as a phylogenetically older, automatic and poorly controlled emergency adaptive system. The Gainotti’s (2005 and 2007a) model assumes that the non-verbal functional organization of the RH may be considered as more primitive than the verbal functional organization of the LH and may be caracterized by a higher degree of reliance on sensorimotor functions, emotional processing, unawareness, and automaticity, whereas the LH’s functional organization might be characterized by a prevalence of verbal cognitive processing, consciousness and intentionality. This model could explain the different format of conceptual and familiar people representations, which are stored in a sensory non-verbal format in the right anterior temporal lobe ATL and in a verbal modality in the left ATL (Gainotti 2007b, 2012b, 2015). It could also explain some automatic functions typical of the RH, such as the automatic generation of familiarity feelings, involved in the recognition of facial identity (Gainotti 2007a) and the disruption of the automatic components of the spatial orienting of attention that charaterizes the RH syndrome of unilateral spatial neglect (see Gainotti 1996 for a critical review). As for the properly emotional functions, this model could explain both the persistence in the RH of a more primitive treatment of emotions through the subcortical pathway (LeDoux 1996; Tamietto and de Gelder 2010) and the greater involvement of the RH in comprehension and expression of negative emotions, because the definition of emotions as a phylogenetically old emergency system could apply more to danger-related negative emotions than to positive and social types of emotions. Consistent with the aforementioned model are also results of activation studies, conducted by Critchley and others (2004) and Gray and others (2007), which have shown that even the subjective emotional feelings, that are the distinctive feature of spontaneous emotions, are mainly subsumed by RH structures, and, in particular by the right AIC, which represents emotional experience, because it receives and integrates interoceptive inputs from the whole body, guiding second-order “cognitive” representations of bodily arousal state. Taken together, all these investigations, which have shown that the right amydgala plays a critical role in the early assessment of emotional stimuli, the right anterior insula provides the substrate for subjective feeling states and the right vmPFC is involved in the control and “topdown” modulation of intense emotional responses support and provide an updated, detailed and congruent version of early models assuming a general RH dominance for emotions.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.