
Abstract
Oligodendrocytes generate myelin sheaths to promote rapid neurotransmission in the central nervous system (CNS). During brain development, oligodendrocyte precursor cells (OPCs) are generated in the medial ganglionic eminence, lateral ganglionic eminence, and dorsal pallium. OPCs proliferate and migrate throughout the CNS at the embryonic stage. After birth, OPCs differentiate into mature oligodendrocytes, which then insulate axons. Oligodendrocyte development is regulated by the extrinsic environment including neurons, astrocytes, and immune cells. During brain development, B lymphocytes are present in the meningeal space, and are involved in oligodendrocyte development by promoting OPC proliferation. T lymphocytes mediate oligodendrocyte development during the remyelination process. Moreover, a subset of microglia contributes to oligodendrocyte development during the neonatal periods. Therefore, the immune system, especially lymphocytes and microglia, contribute to oligodendrocyte development during brain development and remyelination.
Keywords
Introduction
Oligodendrocytes are myelinating glial cells in the central nervous system (CNS). They produce myelin sheath that insulates axons to promote neurotransmission by salutatory conduction. In addition, oligodendrocytes support axonal survival by secreting neurotrophic factors or transporting carbohydrates (Wilkins and others 2001; Wilkins and others 2003; Lee and others 2012). Thus, oligodendrocytes play crucial roles in the maintenance of CNS homeostasis.
Oligodendrocyte development is regulated by several extrinsic factors during brain development. For example, platelet-derived growth factor-A (PDGF-A) secreted from neurons promotes the proliferation of oligodendrocyte progenitor cells (OPCs) (Calver and others 1998; Fruttiger and others 1999; Yeh and others 1991). Fibroblast growth factor-2 (FGF-2)-deficient mice exhibit a reduction of oligodendrocytes (Murtie and others 2005). Moreover, it has been revealed that the immune system also contributes to oligodendrocyte development. Traditionally, the CNS is considered to be an immune-privileged organ because of the loss of lymphatic vessels and immune cells. However, a recent study has demonstrated that a lymphatic vessel is present in the brain, which lines the dural sinus (Louveau and others 2015). Furthermore, lymphocytes are localized in the meningeal space, choroid plexus, and perivascular spaces (Prinz and Priller 2017). These reports support the notion that immune cells are involved in brain development and function. Particularly, we recently found that lymphocytes are major contributor to oligodendrocyte development. In this review, we describe the function of lymphocytes and the immune system in oligodendrocyte development from recent findings.
Genes Expressed in Oligodendrocyte Lineage Cells and Their Functions
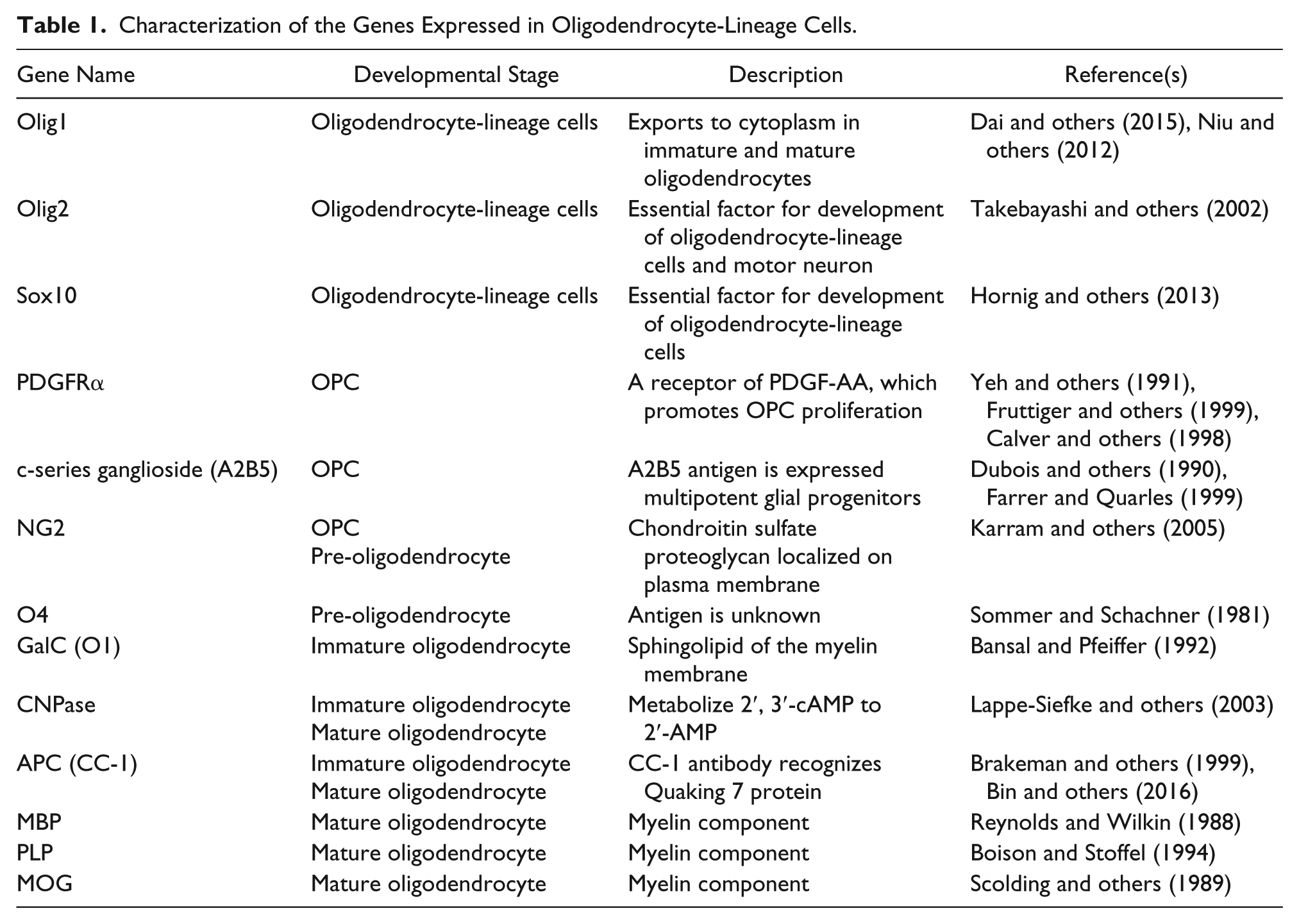
Oligodendrocytes develop through several stages during CNS development (Schumacher and others, 2012). Oligodendrocyte lineage cells are classified into OPCs, pre-oligodendrocytes, immature oligodendrocytes, and mature oligodendrocytes, and each type of cells expresses distinct genes. All oligodendrocyte lineage cells express Olig1, Olig2, and Sox10. These transcriptional factors are indispensable to the development of oligodendrocytes. Olig2-deficient mice lack oligodendrocytes and spinal motor neurons in the embryonic CNS (Takebayashi and others 2002). Notably, Olig2 is also expressed in embryonic spinal motor neurons (Lee and others 2005) and astrocytes of the postnatal subventricular zone (SVZ) (Marshall and others 2005). Thus, Olig2 itself is not a specific marker for oligodendrocyte lineage cells. Olig1 is also an important regulator for oligodendrocyte development. Olig1-deficient OPCs are more proliferative, but their development into mature oligodendrocytes is impaired, suggesting that Olig1 is an essential factor for oligodendrocyte differentiation (Dai and others, 2015). Although Olig1 is localized in the nucleus of OPCs and pre-oligodendrocyte, Olig1 is exported to the cytoplasm of immature and mature oligodendrocytes (Niu and others, 2012). CNS-specific Sox10-deficient mice exhibit a decrease of mature oligodendrocyte without any noticeable effect on the number of OPCs (Hornig and others 2013). Therefore, Olig1, Olig2, and Sox 10 are pivotal regulators of oligodendrocyte development.
OPCs highly express platelet-derived growth factor receptor α (PDGFRα), A2B5, and neuron-glial antigen 2 (NG2). PDGFRα is a receptor for PDGF-A, which promotes the proliferation of OPC during brain development. A2B5 is a monoclonal antibody, which recognizes c-series ganglioside (Dubois and others 1990). A2B5 immunoreactivity is high in multipotent glial progenitors and decreases when the progenitors mature into glial cells (Farrer and Quarles, 1999). NG2 is a chondroitin sulfate proteoglycan that localizes to the plasma membrane (Karram and others, 2005). As NG2 is also expressed in CNS pericytes, it is difficult to distinguish the vascular OPCs and pericytes by immunostaining with an anti-NG2 antibody. NG2 expression in OPCs persists to the pre-oligodendrocytes phase. The markers for pre-oligodendrocytes are NG2 and O4. O4 is a monoclonal antibody, which recognizes the antigen localized on the surface of pre-oligodendrocytes (Sommer and Schachner, 1981). However, the antigen recognized by the O4 antibody has not been identified so far. Immature oligodendrocytes express 2′,3′-cyclic-nucleotide 3’-phosphodiesterase (CNPase) and galactosylceramide (GalC). CNPase is an enzyme that metabolizes 2′,3′-cAMP to 2′-AMP in immature and mature oligodendrocytes. Disruption of CNPase expression results in the impairment of myelination (Lappe-Siefke and others, 2003). GalC is the sphingolipid of the myelin membrane, which is recognized by the O1 monoclonal antibody (Bansal and Pfeiffer 1992). Moreover, a monoclonal antibody against adenomatous polyposis coli (APC) (CC-1 clone) is used to detect immature oligodendrocytes and mature oligodendrocytes. Previous studies have shown that the CC-1 antibody does not bind to APC but recognizes Quaking 7 (QKI-7), which is an RNA binding protein expressed in immature and mature oligodendrocytes (Bin and others, 2016; Brakeman and others, 1999). Myelinating mature oligodendrocytes express myelin oligodendrocyte basic protein (MBP), myelin oligodendrocyte glycoprotein (MOG), and proteolipid protein (PLP) (Boison and Stoffel, 1994; Reynolds and Wilkin 1988; Scolding and others, 1989). These proteins are the components of myelin in the CNS and are produced by mature oligodendrocytes. The list of oligodendrocyte-related genes and their roles is shown in Table 1.
Characterization of the Genes Expressed in Oligodendrocyte-Lineage Cells.
Oligodendrocyte Development during Embryonic and Neonatal Periods
OPCs in the forebrain are derived from the NKX2.1+ area, which is indicative of the neuroepithelium and includes the medial ganglionic eminence (MGE) and anterior entopeduncular area (AEP). PDGFRα+ OPCs are initially seen in the NKX2.1+ MGE area at embryonic day 12.5 (E12.5) in mice (Tekki-Kessaris and others 2001), and tangentially and dorsally migrate throughout the brain (Kessaris and others 2006; Ono and others 2001) (Fig. 1A). OPC production from the MGE is mediated by sonic hedgehog (SHH) signaling (Nery and others 2001). SHH is secreted from MGE and promotes the differentiation of neural stem cells to OPCs by driving the expression of NKX6.1 and NKX6.2, which regulate the transcriptions of Olig1 and Olig2. Moreover, FGF-1 and FGF-2 are also involved in OPC production, as mice deficient in FGF-1 and FGF-2 exhibit a reduction of OPCs in the forebrain at E12.5 and E14.5 (Furusho and others 2011). Furthermore, FGF-2 treatment promoted OPC differentiation from neural stem cells isolated from the dorsal area of the neuroepithelium in vitro (Chandran and others 2003). These results indicate that SHH and FGF signaling are necessary for OPC production in the embryonic forebrain. At E15.5, OPCs are newly generated from the GSH2+ area in the lateral ganglionic eminence (LGE), and then migrate to the cerebral cortex (Fig. 1A) (Chapman and others 2013; Kessaris and others 2006). GSH2 deficiency increased the number of OPCs in the SVZ of the LGE, suggesting that GSH2 works as a developmental clock by regulating the timing of OPC differentiation in the LGE. At postnatal day 0 (P0), OPCs are generated from an EMX1+ area, which is indicative of the dorsal pallium (DP), and are displaced by the MGE-derived OPCs by P10 in the cerebral cortex and corpus callosum (Fig. 1A) (Kessaris and others 2006). Thus, OPC production phases consist of three waves during brain development.
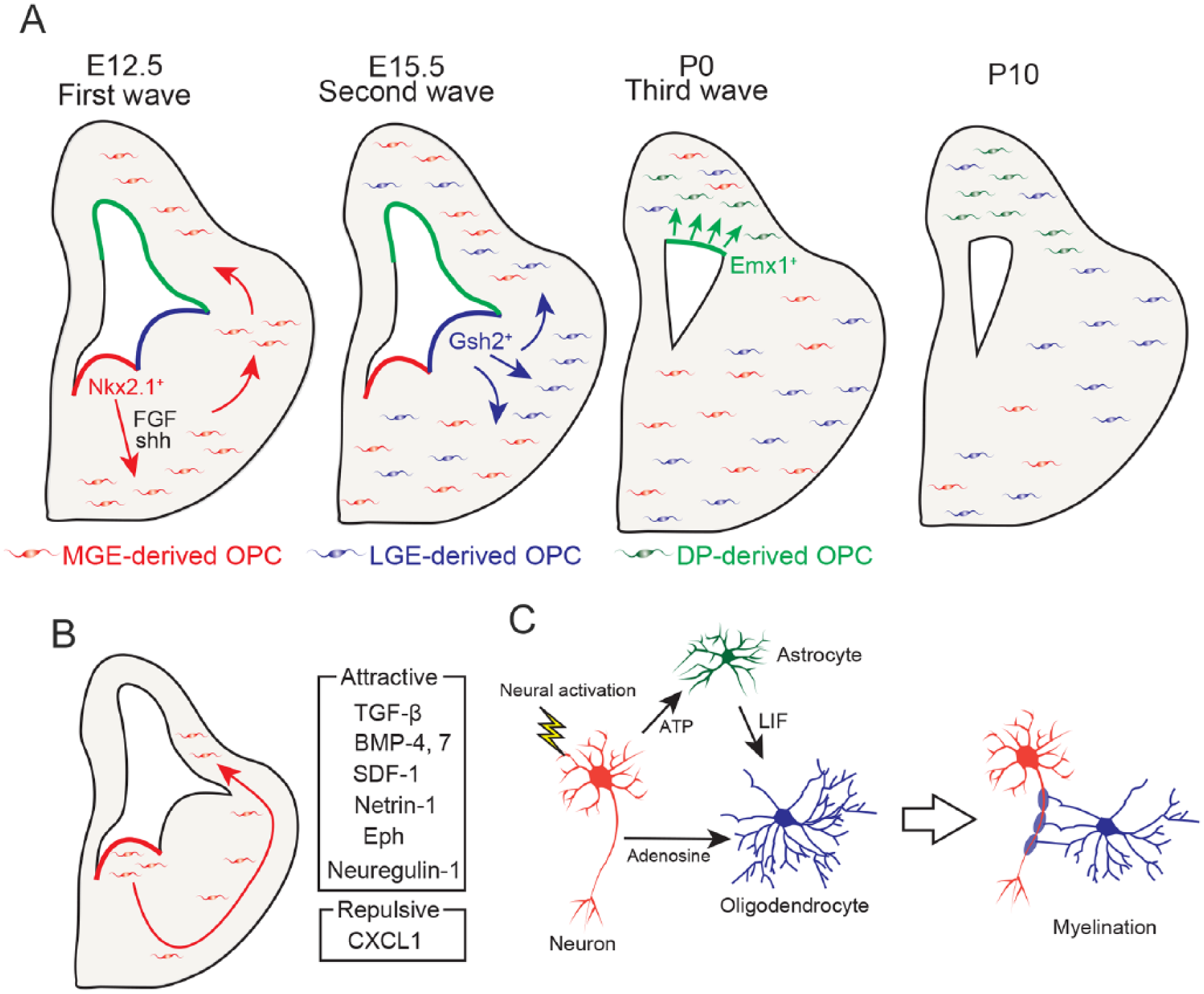
Oligodendrocyte developmental processes in the developing brain. (A) Oligodendrocyte precursor cell (OPC) development consists of three waves (Kessaris and others 2006). At E12.5, OPCs are generated from the NKX2.1+ medial ganglionic eminence (MGE) area in an SHH- and FGF-dependent manner and migrate throughout the brain (first wave). At E15.5, OPCs are generated from the GSH2+ lateral ganglionic eminence (LGE) area (second wave) and migrate throughout the brain. At P0, OPCs are generated from the EMX1+ dorsal pallium (DP) area and migrate to the cerebral cortex (third wave). At P10, MGE-derived OPCs in the cerebral cortex and corpus collosum are displaced by LGE-derived and DP-derived OPCs. (B) MGE-derived OPCs migrate throughout the brain during the embryonic period. Attractive and repulsive molecules related to OPC migration are shown on the right side. (C) Activated neurons promote myelination by secreting ATP and adenosine. ATP induces the secretion of leukemia inhibitory factor (LIF) by astrocytes. Adenosine and LIF promote myelination by mature oligodendrocytes.
After OPC development, OPCs migrate throughout the CNS. Migration of OPC is modulated by a number of extrinsic environmental factors including neuronal axons, the extracellular matrix, and the vasculature (Fig. 1B). OPC migration is regulated by Eph/Ephrin and neuregulin-1/EbrB4 signaling in the optic nerve (Ortega and others 2012; Prestoz and others 2004). Netrin-1 is required for OPC distribution in the spinal cord (Tsai and others 2006). OPC migration is also regulated by chemokines and cytokines. C-X-C motif chemokine ligand 1 (CXCL1) is the chemokine family member that attracts leukocytes to target sites by interacting with C-X-C motif chemokine receptor 2 (CXCR2). Ex vivo and in vitro studies have shown that CXCL1 inhibited OPC migration in the spinal cord by interacting with CXCR2, suggesting that CXCL1 regulates the proper migration of OPCs in the spinal cord (Tsai and others 2002). OPCs express various chemokine receptors, including CXCR4, which binds to stromal cell-derived factor 1 (SDF-1). CXCR4 antagonist and anti-SDF-1 antibody treatment impaired OPC migration in the embryonic cerebral cortex (Dziembowska and others 2005). SDF-1 is secreted from vascular vessels and is involved in OPC migration in the embryonic brain (Tsai and others 2016). OPCs migrate along with vascular vessels in the embryonic brain as shown by time lapse imaging studies, and CXCR4 signaling is required for OPC migration mediated by the vascular system. Indeed, OPC migration is impaired in G protein-coupled receptor 124-deficient mice, which have disrupted vascular system development, in the embryonic cerebral cortex. Thus, OPC migration is regulated by the vascular system in a SDF1-CXCR4 signaling-dependent manner. OPC migration to the cerebral cortex is regulated by bone morphogenetic protein (BMP)-4, BMP-7, and transforming growth factor (TGF)-β (Choe and others, 2014). During embryonic development, OPCs are generated from the MGE and migrate from the ventral forebrain to the dorsal cerebral cortex. In situ hybridization studies have shown that BMP4 and BMP7 are expressed in the ventral forebrain in facial mesenchymal cells and meninges. Inhibition of BMP signaling resulted in the disruption of OPC distribution in the cerebral cortex. TGF-β is expressed in pericytes of the cerebral cortex, and mice with a pericyte-specific deletion of TGF-β had reduced a number of OPCs in the cerebral cortex. Thus, TGF-β expressed in pericytes promote the migration of ventral OPCs to the cerebral cortex. Collectively, OPC migration is spatially and temporally regulated by a number of factors.
OPCs differentiate into mature oligodendrocytes during neonatal development. Although MBP-positive mature oligodendrocytes are not visible at P1 in the white matter of mice, they are sparsely seen at P5 in the corpus callosum (Fig. 2A). From P7 to P14, mature oligodendrocytes increase with age, and begin to myelinate the axons (Fig. 2A and B). At early neonatal periods, inhibitory signals such as Notch-Jagged or Wnt/β-catenin inhibit oligodendrocyte differentiation (Fancy and others 2009; Wang and others 1998). Maturation of OPCs to oligodendrocytes is rigorously regulated by a number of mechanisms including neural activity, interaction with astrocytes, and the immune system (Fig. 1C). Adenosine secreted from neurons in response to neuronal activity promotes oligodendrocyte differentiation and myelination (Stevens and others 2002). Adenosine triphosphate (ATP) produced by activating axons induces the secretion of leukemia inhibitory factor (LIF) from astrocytes, which promotes myelination (Ishibashi and others, 2006). In addition, some lymphocytes and immune-related molecules affect oligodendrocyte development.
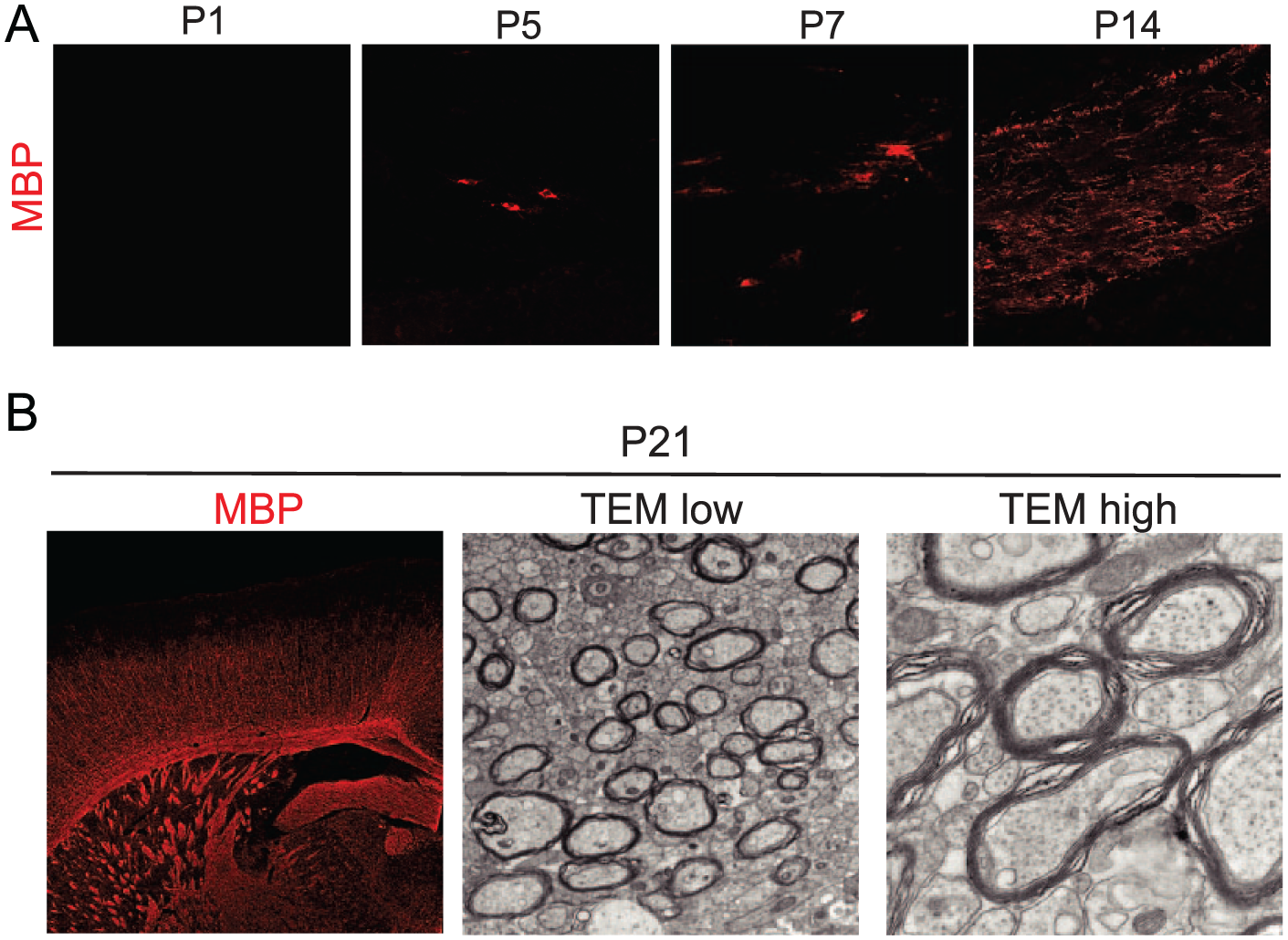
Oligodendrocyte development during brain development. (A) Representative images show the immunohistochemistry for myelin oligodendrocyte basic protein (MBP) in the corpus callosum of mice brain at P1, P5, P7, and P14. MBP+ mature oligodendrocytes are initially seen in the developing brain at P5 and increased with age. (B) Representative images show the immunohistochemical staining for MBP (left), transmission electron microscopy (TEM) at low magnification (center), and TEM at high magnification (right) in the corpus callosum of mouse brain at P21. A number of myelinated axons could be observed at P21.
Contribution of B Lymphocyte to Oligodendrocyte Development
Lymphocytes are one type of leukocytes that participate in the adaptive immune system. B lymphocytes, T lymphocytes, and natural killer cells are involved in humoral immunity, cell-mediated immunity, and cytotoxic innate immunity, respectively. Traditionally, lymphocytes were considered to be unable to infiltrate into the CNS due to the presence of the blood-brain barrier (BBB), even in the developing CNS. The BBB develops between embryonic days 11 to 13 in rodents, and after 12 weeks of gestation in humans (Bauer and others 1993; Grontoft 1954). Although lymphocytes are not present in the CNS parenchyma, they are present in the meningeal space and are involved in brain functions. For example, interleukin (IL)-4-secreting T lymphocytes regulate cognitive function, and interferon-γ-secreting T lymphocytes mediate neural connectivity (Derecki and others 2010; Filiano and others 2016). These reports indicate that lymphocytes are pivotal players in regulating brain functions.
We reported that B lymphocytes contribute to oligodendrocyte development (Tanabe and Yamashita 2018). Flow cytometry analysis showed that B lymphocytes are abundant in the neonatal brain. B lymphocytes are antibody-producing lymphocytes and could be classified into several subtypes such as B-2 lymphocyte, regulatory B lymphocytes, B-1a lymphocyte, and B-1b lymphocytes (Baumgarth 2011). B-2 lymphocytes produce antigen-specific antibodies in response to the stimulation from helper T cells. Regulatory B lymphocytes produce anti-inflammatory cytokines such as IL-10 and TGF-β. B-1a and B-1b lymphocytes participate in the innate immune system by secreting natural IgM antibodies, which bind to pathogens, without the stimulation of helper T cells. Furthermore, flow cytometric analysis demonstrated that the B lymphocytes in the neonatal brain are CD19+ CD45R+ CD5+ CD21low CD23– CD93– CD1dmid CD43+ IgMhi B-1a lymphocytes (Fig. 3A). B-1a lymphocytes are located in the meningeal space and choroid plexus of the neonatal brain. On the other hand, the B lymphocytes in neonatal blood are CD19+ CD45R+ CD93+ immature B lymphocytes, suggesting that the subtype of B lymphocytes is altered after infiltrating into the brain. We investigated the molecular mechanism of B-1a lymphocyte migration into the neonatal brain. It has been reported that B-1 lymphocytes migrate to the body cavity via CXCL13 and CXCR5 signaling (Ansel and others 2002). Thus, we examined whether B-1a lymphocytes also migrate to the developing brain in a CXCL13-CXCR5-dependent manner. Expression analyses showed that B lymphocytes in the neonatal blood and brain express CXCR5, and CXCL13 is expressed in the choroid plexus of the neonatal brain. Administration of an anti-CXCL13 antibody inhibited the migration of B lymphocytes to the neonatal brain. These results indicated that B lymphocytes are attracted by the choroid plexus in a CXCL13-CXCR5-dependent manner (Fig. 3B). The molecular mechanism that induces the upregulation of CXCL13 in the choroid plexus in the neonatal brain remains to be determined.
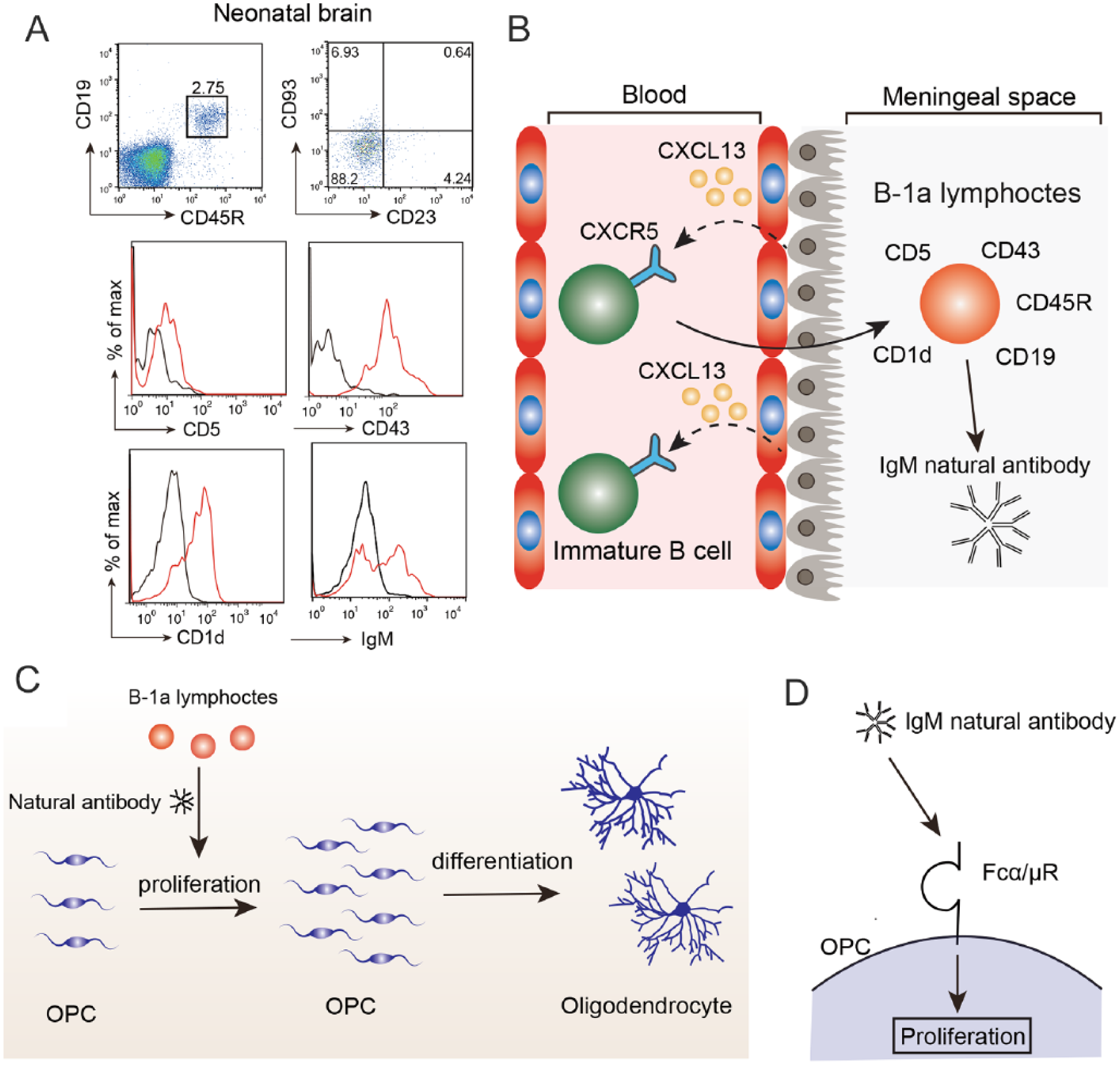
Molecular mechanism of oligodendrocyte development by B-1a lymphocytes. (A) Flow cytometry data of B lymphocytes in the neonatal brain. CD19+ CD45R+ B lymphocytes are CD93– CD23– mature B lymphocytes. Histogram data represent the expression levels of CD5, CD43, CD1d, and IgM. Black, isotype IgG; red, specific antibody. (B) During neonatal periods, B lymphocytes in the blood are immature, express CXCR5, and are attracted to the meningeal space in a CXCL13-CXCR5-dependent manner. After migrating to the meningeal space, immature B lymphocytes differentiate into B-1a lymphocytes, which express CD19, CD45R, CD5, CD43, CD1d, and natural IgM antibodies. (C) B-1a lymphocytes secrete natural IgM antibodies. The natural IgM antibodies promote oligodendrocyte precursor cell (OPC) proliferation and increases the number of mature oligodendrocytes. (D) Natural IgM antibodies bind to the Fcα/μR receptor expressed in OPCs, leading to the promotion of proliferation.
In vitro experiments using a co-culture of B-1a lymphocytes with neural stem cells demonstrated that B-1a lymphocytes increased the number of oligodendrocytes and promoted the proliferation of OPCs. Therefore, B-1a lymphocytes have the capacity to promote the proliferation of OPCs in vitro. For in vivo experiments, we depleted B lymphocytes by injecting a depletion antibody into the developing brain. Consequently, mature oligodendrocytes were reduced in the corpus callosum, and OPC proliferation was inhibited by the depletion of B lymphocytes. The administration of B-1a lymphocytes rescued the proliferation of OPCs in the developing B cell-depleted brain. Thus, B-1a lymphocytes play a pivotal role in oligodendrocyte development in vivo (Fig. 3C). B-1a lymphocytes secrete natural IgM antibodies (Hayakawa and others 1983). As oligodendrocyte development is regulated by the Fc receptor for IgG (Nakahara and others, 2003), it is possible that B-1a lymphocytes promote the proliferation of OPCs by the signaling of the Fc receptor and IgM natural antibody. Indeed, OPCs express the Fc receptor for IgM (Fcα/μR), and injection of natural IgM into the neonatal brain promoted OPC proliferation (Fig. 3D). Fcα/μR is a single transmembrane protein with an Ig-like domain, and a blocking antibody against Fcα/μR was developed (Shibuya and Honda 2006). We administered the blocking antibody against Fcα/μR and found that the anti-Fcα/μR treatment reduced the OPC proliferation and number of mature oligodendrocytes. Moreover, anti-Fcα/μR treatment reduced the number of myelinated axons without affecting the myelin thickness at P21. These results suggested that B-1a lymphocytes at the neonatal stage contribute to oligodendrocyte development and myelination by secreting natural IgM antibodies via Fcα/μR (Fig. 3C and D).
Natural monoclonal IgM antibodies facilitate the therapeutic effects to demyelinating diseases of mouse model (Wright and others 2009). They recognized myelin antigens in the slice culture system and promoted remyelination in experimental autoimmune encephalomyelitis (EAE), which is an inflammatory demyelinating animal model, and Theiler’s murine encephalomyelitis (TMEV)-induced demyelinating mouse models (Lemus and others 2017; Warrington and others 2000). It is considered that these antibodies recognize membrane glycolipids on oligodendrocytes. However, we showed that Fcα/μR was expressed in OPCs, but not mature oligodendrocytes. Thus, therapeutic effects of monoclonal IgM antibodies to demyelination are not mediated by Fcα/μR. It is expected that administration of several types of IgM may be more effective to demyelinating diseases through multiple mechanisms.
The Role of T Lymphocytes in Oligodendrogenesis and Demyelinating Diseases
It has been reported that T lymphocytes also contribute to oligodendrocyte development in the demyelinating rodent models. Lysophosphatidylcholine (LPC), a bioactive lysophospholipid, is used to induce a focal demyelination in the CNS. LPC treatment disrupts the BBB and induces the infiltration of immune cells such as CD4+ T lymphocytes, CD8+ T lymphocytes, macrophages, and neutrophils (Kuroda and others 2017; Ousman and David 2000). Moreover, recombination activating gene-1 (RAG-1)-deficient mice (T and B lymphocyte-deficient mice) showed a reduction of spontaneous remyelination after LPC treatment (Bieber and others 2003). Depletion of CD4+ T lymphocytes or CD8+ T lymphocytes also reduced remyelination, suggesting that T lymphocytes contribute to remyelination as well as demyelination. In contrast, nude mice (T lymphocyte-deficient mice) exhibit a reduction of the demyelination area after LPC treatment (Ghasemlou and others 2007). The difference in the phenotype may be explained by the difference of mouse strain. The mouse strain used in the RAG-1-deficient mouse study is C57BL/6J, but BALB/c mice were used in the nude mouse study. The proportion of the T lymphocyte subtypes differs between C57BL/6J (Th1 dominant) and BALB/c mice (Th2 dominant) (Mills and others 2000). Indeed, BALB/c mice are resistant to demyelination induced by cuprizone feeding (Skripuletz and others 2008). Therefore, the role of T lymphocytes in oligodendrocyte development and remyelination might be dependent on the proportion of the T lymphocyte subtype.
CD4+ helper T lymphocytes are classified into several subtypes such as Th1, Th2, Th17, and regulatory T (Treg) cells. Th1 cells are involved in cellular immunity by secreting interferon-γ and IL-12 and are considered to mediate inflammation in autoimmune diseases. Th2 cells are associated with humoral immunity by secreting IL-4, IL-5, and IL-13. Th17 cells play critical roles in the pathogenesis of autoimmune diseases by secreting proinflammatory cytokines such as IL-17, IL-21, and IL-22. Treg cells suppress immune responses by secreting anti-inflammatory cytokines such as IL-10 and TGF-β. Conditioned medium from Th1 cells inhibits human oligodendrocytes differentiation in vitro (Moore and others 2015). Th2 cells treated in vitro with glatiramer acetate, which is used as an immunomodulating treatment for patients with multiple sclerosis, increased the number of oligodendrocyte progenitors by producing insulin-like growth factor-2 (IGF-2) (Zhang and others 2010). However, Th2 conditioned medium, not treated with glatiramer acetate, did not affect the number of oligodendrocytes in vitro (Moore and others, 2015). These reports indicated that glatiramer acetate was necessary to facilitate oligodendrogenesis by Th2 cells. A recent study showed that ablation of Treg cells inhibited remyelination after LPC injection (Dombrowski and others, 2017). Treg cell administration increased the number of myelinating oligodendrocytes after LPC injection. Ex vivo and in vitro experiments demonstrated that Treg cells directly promoted oligodendrocyte differentiation and remyelination by secreting CCN3, a growth regulatory protein. These results suggested that Treg cells have the capacity to promote oligodendrocyte differentiation and remyelination. On the other hand, IL-17, which is a pro-inflammatory cytokine secreted from Th17 cells, inhibited oligodendrocyte maturation and reduced the survival of oligodendrocytes (Kang and others 2013). Ablation of IL-17-mediated signals in oligodendrocyte lineage cells by using the Cre-loxP system attenuated EAE severity by reducing demyelination and neuroinflammation. However, ablation of IL-17-mediated signals in mature oligodendrocytes did not attenuate EAE severity, suggesting that IL-17 signals in OPCs or immature oligodendrocytes are indispensable for demyelination and neuroinflammation. IL-17 induces apoptosis of OPCs and inhibited OPC differentiation into oligodendrocytes by promoting the expression of NOTCH1 (Wang and others 1998). Moreover, IL-17 enhances inflammatory gene expression via NOTCH1 signaling in OPCs (Wang and others 2017). These reports suggest that Th17 cells promote the neuroinflammation and demyelination by interacting with OPCs in EAE pathology. Collectively, the contribution of T lymphocytes to oligodendrocyte differentiation and remyelination depends on the subtype of T lymphocytes (Fig. 4, Table 2). To our knowledge, it is still unknown whether T lymphocytes mediate oligodendrocyte maturation and myelination during brain development. Thus, further studies are required to elucidate the temporal-spatial effects of T lymphocytes on oligodendrocyte maturation during brain development.
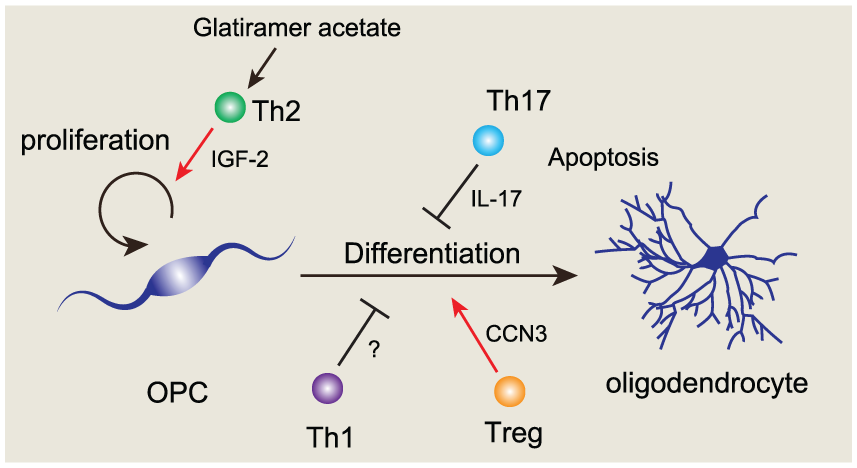
Regulation of oligodendrocyte development by T lymphocytes. Glatiramer acetate stimulated-Th2 cells promote the oligodendrocyte precursor cell (OPC) proliferation by secreting IGF-2 (Zhang and others, 2010). Th17 cells inhibit oligodendrocyte differentiation by secreting IL-17 (Kang and others 2013). In addition, Th17 cells induce the apoptosis in oligodendrocytes, and promote neuroinflammation. Th1 cells inhibit oligodendrocyte differentiation (Moore and others 2015). Treg cells promote oligodendrocyte differentiation by producing CCN3 (Dombrowski and others 2017).
Role of T Cells in Oligodendrogenesis.
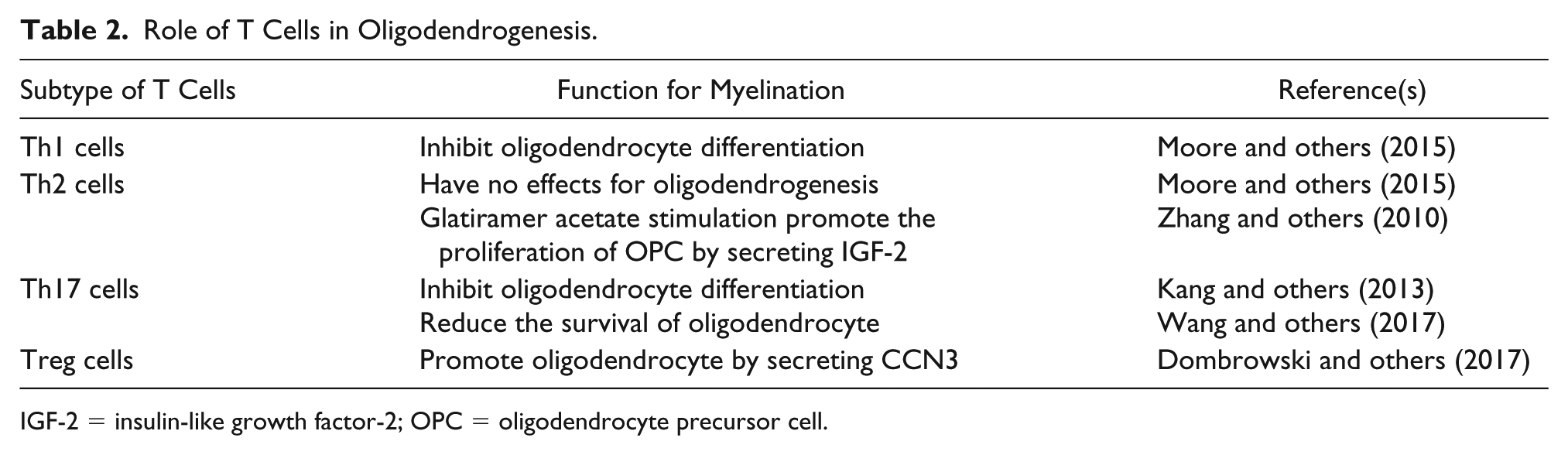
IGF-2 = insulin-like growth factor-2; OPC = oligodendrocyte precursor cell.
Contribution of Microglia to Oligodendrocyte Development
Microglia are tissue-resident myeloid cells in the CNS and are characterized by the expression of CD11b, CX3C chemokine receptor 1 (CX3CR1), and ionized calcium-binding adapter molecule 1 (Iba-1). Microglia originate from primitive c-kit+ erythromyeloid precursors from the yolk sac and migrate to the CNS during early embryonic periods (Ginhoux and others 2010; Kierdorf and others 2013). Traditionally, the function of microglia has been studied extensively under the inflammatory conditions of neurological diseases. However, recent studies have shown that microglia are also involved in brain development by regulating the neural development and synapse elimination (Bilimoria and Stevens 2015).
At early neonatal periods, microglia are activated and accumulate around the SVZ. Minocycline treatment, which suppresses the microglia activation, decreased the number of O1+ immature oligodendrocytes and MBP+ mature oligodendrocytes in the SVZ, suggesting that activated microglia contribute to oligodendrocyte development (Shigemoto-Mogami and others, 2014). Microglia are classified into several subtypes. CD68+ Arginase-1+ microglia and macrophages promote oligodendrocyte differentiation by secreting activin-A, which belongs to the transforming growth factor superfamily (Miron and others 2013). Arginase-1+ microglia are found in the neonatal brain (Lenz and others 2013). These reports imply Arginase-1+ microglia are involved in oligodendrocyte development in the developing brain. A recent study identified a unique microglia subtype that contributes to myelination during brain development (Wlodarczyk and others 2017). They showed that CD11c+ microglia were abundant in the corpus callosum and cerebellar white matter of the neonatal brain (Fig. 5A) and are a major source of insulin like-growth factor-1 (IGF-1). IGF-1 is a neurotrophic factor associated with cell survival, proliferation, and differentiation (Joseph D’Ercole and Ye 2008). IGF-1 deficiency from CD11c+ microglia resulted in the reduction of myelin-related proteins and abnormal myelinogenesis, suggesting that CD11c+ microglia promote myelination during brain development by secreting IGF-1. Indeed, other studies showed that IGF-1 receptor signaling is required for oligodendrocyte development and proper myelination (Zeger and others 2007). An in vitro study demonstrated that IGF-1 promoted the proliferation of OPCs by PI3K/Akt, MEK/ERK, and Src-like tyrosine kinase signaling (Cui and Almazan 2007).
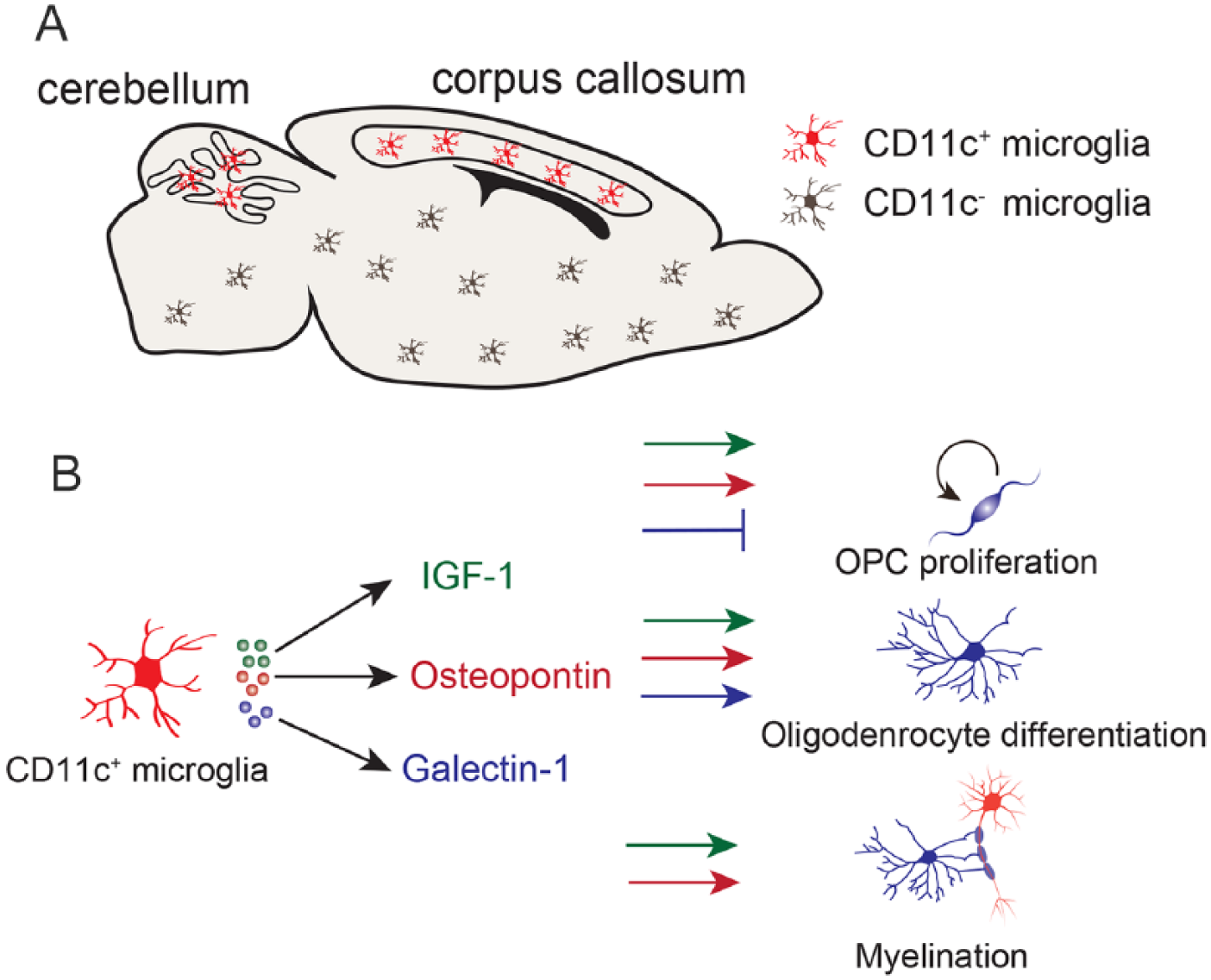
The role of CD11c+ microglia in oligodendrocyte development. (A) CD11c+ microglia are mainly located in the corpus callosum and white matter of the cerebellum in the neonatal brain (Wlodarczyk and others 2017). Red, CD11c+ microglia; brown, CD11c– microglia. (B) CD11c+ microglia secrete IGF-1 (green), osteopontin (red), and Galectin-1 (blue). Both IGF-1 and osteopontin promote oligodendrocyte precursor cell (OPC) proliferation, oligodendrocyte differentiation, and myelination. Galectin-1 inhibits OPC proliferation but promotes oligodendrocyte differentiation. Galectin-1 does not have any effects on the myelination process.
CD11c+ microglia also express osteopontin (also known as Spp1) and Galectin 1 (also known as Lgals1) (Wlodarczyk and others 2017). Osteopontin is an extracellular matrix protein that plays a role in inflammation, cell remodeling, and differentiation. Microglia around the SVZ of the developing brain express osteopontin, which promotes the proliferation of OPCs and myelin formation (Choi and others 2004; Selvaraju and others 2004). These studies indicate that osteopontin from microglia also modulates the oligodendrocyte development in the developing brain. Indeed, osteopontin contained in the milk promotes the expression of oligodendrocyte-related genes (Jiang and others 2018). Thus, osteopontin from microglia and milk plays crucial roles in myelin development in the developing brain. Galectin-1, a family of animal lectins, binds to β-galactoside and is involved in inflammation, infection, and cell proliferation. Galectin-1 treatment promoted remyelination in the corpus callosum after LPC treatment (Rinaldi and others 2016). Galectin-1 treatment inhibits OPC proliferation but promotes oligodendrocyte development in vitro. However, galectin-1 treatment did not affect myelin thickness after remyelination in vivo. These results suggest that Galectin-1 regulates remyelination by promoting oligodendrocyte differentiation. It remains unclear whether Galectin-1 affects oligodendrocyte development in the developing brain. Collectively, microglia regulate oligodendrocyte development by secreting factors such as IGF-1, osteopontin, and Galectin-1 (Fig. 5B).
Conclusion
In this review, we discuss the relationship between oligodendrocyte development and the immune system during brain development. Oligodendrocyte development is modulated by the extracellular environmental factors such as neuronal activity, developmental clock, and the immune system. In this review, we introduced that B lymphocytes, T lymphocytes, and microglia contribute to oligodendrocyte development in the developing brain. Importantly, lymphocytes affect myelin formation by producing secreting factors.
We demonstrated that B lymphocytes promote the proliferation of OPCs via natural IgM antibodies. T lymphocytes are involved in oligodendrocyte differentiation in the demyelination animal model. However, it remains unclear whether B lymphocytes or T lymphocytes also mediate the oligodendrocyte migration or differentiation in the developing brain. It has been reported that myelin abnormalities are associated with several psychiatric and neurodevelopmental diseases. For example, myelin abnormalities were detected in the autopsied brain of patients with schizophrenia and autism (Deoni and others 2015; Flynn and others 2003). Moreover, a genome-wide association study demonstrated that the schizophrenia and neurodevelopmental disorders were associated with immune molecules including CD19 and CD20, which are B lymphocyte-related molecules (Schizophrenia Working Group of the Psychiatric Genomics Consortium 2014; Voineagu and others, 2011). These reports imply that the immune system mediates the pathology of schizophrenia and neurodevelopmental disorders. Although it remains unclear whether the myelin abnormalities in patients with schizophrenia and neurodevelopmental disorders are caused by immune dysfunction, it is worth studying whether immune dysfunction is involved in the pathogenesis of schizophrenia and neurodevelopmental disorders by impairing the myelinogenesis during brain development.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a Grant-in-Aid for Scientific Research (S) from the Japan Society for the Promotion of Sciences (17H06178) to T.Y.