
Abstract
While in the past much of our knowledge about memory representations in the brain has relied on loss-of-function studies in which whole brain regions were temporarily inactivated or permanently lesioned, the recent development of new methods has ushered in a new era of downright “engram excitement.” Animal research is now able to specifically label, track, and manipulate engram cells in the brain. While early studies have mostly focused on single brain regions like the hippocampus, recently more and more evidence for brain-wide distributed engram networks is emerging. Memory research in humans has also picked up pace, fueled by promising magnetic resonance imaging (MRI)-based methods like diffusion-weighted MRI (DW-MRI) and brain decoding. In this review, we will outline recent advancements in engram research, with a focus on human data and neocortical representations. We will illustrate the available noninvasive methods for the detection of engrams in different neocortical regions like the medial prefrontal cortex and the posterior parietal cortex and discuss evidence for systems consolidation and parallel memory encoding. Finally, we will explore how reactivation and prior knowledge can lead to and enhance engram formation in the neocortex.
Keywords
What Is an Engram?
The question of how memory is represented in our brain has fascinated scientists for centuries. The original definition of the term “engram” dates back to German psychologist Richard Semon, who outlined the engram as an “enduring though primarily latent modification in the irritable substance produced by a stimulus” (Semon 1921, p. 12). Based on his work, four essential features of a memory engram can be derived (Josselyn and others 2015): First, a memory engram is the result of experience and reflects the specific content of that experience. Second, a memory engram has to engender persistent, long-term stable changes, so that, third, later reactivation of the engram enables memory recall and thus adaptive behavior. Fourth, a memory engram does not only exist during encoding and retrieval while it is activated, but it also exists in a functionally inactive, “dormant” state in-between. Consequently, if detected changes in the brain conform to these criteria, they can be regarded as evidence for a memory engram.
Largely influenced by the cellular theory of the nervous system of Ramón y Cajal (1894) and by the concept of Hebbian learning (Hebb 1949), our current understanding is that information is stored in the connections between populations of neurons that were active during encoding and that it is the structural modifications to these connections that make it last (Dudai 2004). This definition already implies that engrams can be found at different levels of investigation. In animal models, enduring changes in the brain are most often described at a molecular level (Lisman 2017; Rossetti and others 2017), at the level of synaptic contacts (Hübener and Bonhoeffer 2010), or at the level of individual engram cells (Tonegawa and others 2015). However, information is represented in the brain in a hierarchical network of locations, each processing specific kinds of information (see Box 1). For example, in contextual fear conditioning, different regions carry different components of the engram, with contextual aspects relying on the hippocampus and emotional aspects on the amygdala (Zelikowsky and others 2014). The context engram in the hippocampus is stored in a number of distinct sub-ensembles of engram cells that are jointly activated. These sub-ensembles seem to represent distinct aspects of the total memory (Ghandour and others 2019). In an experiment on auditory fear conditioning it was shown that inducing long-term potentiation (LTP) by optogenetically stimulating auditory input pathways to the amygdala during foot shock presentation is sufficient to engender a conditioned memory, but inducing LTP in the amygdala alone is not (Nabavi and others 2014). Likewise, the complete artificial induction of odor conditioning by optogenetic stimulation of olfactory neurons and ventral tegmental neurons resulted in a memory engram in the amygdala (Vetere and others 2019). In sum, these results indicate that the full memory is represented by a complex circuit. The engram cannot be pinpointed to a single place, but rather is encoded in a pattern of distributed locations that synchronize to symbolize a certain mneme (Semon 1921). In order to fully understand how memories are stored in the brain, we therefore need to investigate the engram both on the microstructural level of molecules and cells and on the macrostructural level of regions, systems and connectivity (Asok and others 2019) (Fig. 1).
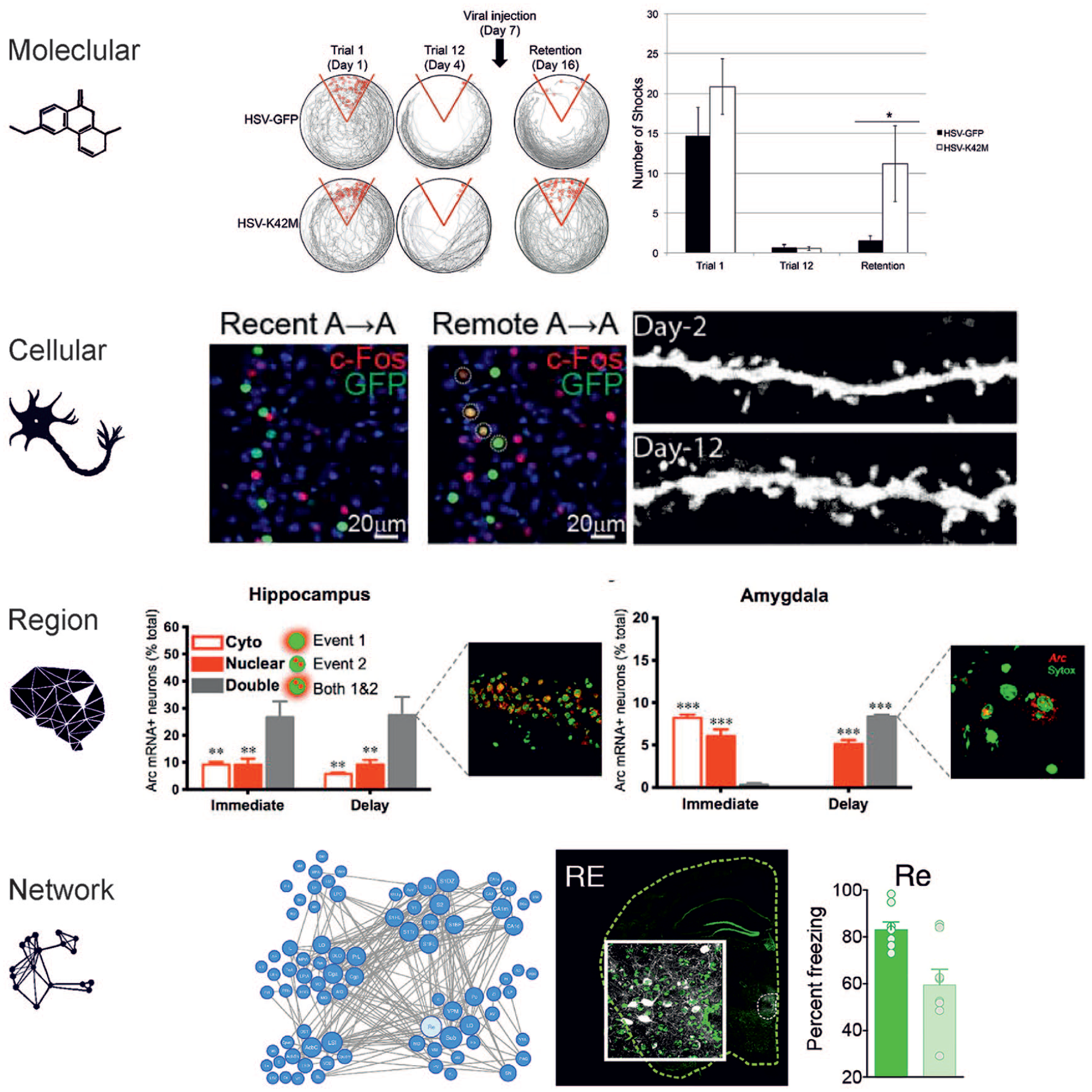
Levels of engram study. Engrams can be investigated on different scales ranging from the molecular level to the level of whole networks. Molecular level: The synaptic protein CamKII regulates memory storage. Memory retention of conditioned place avoidance was impaired when CamKII was inhibited after learning (HSV-K42M) compared with a control condition (HSV-GFP). From Rossetti and others (2017). Cellular level: Memory engram cells in prefrontal cortex become functionally mature over time. Prefrontal engram cells identified with activity-dependent labeling during contextual fear conditioning are preferentially reactivated (circled cells) by the conditioned context and show higher spine density on day 12 compared with on day 2 after conditioning. From Kitamura and others (2017). Brain region level: The hippocampus and amygdala track different components of fear memory. Investigating the time course of expression of the immediate-early gene Arc in rats that received footshocks in a specific context either directly (Immediate, no fear conditioning) or after exploration (Delay, fear conditioning) reveals distinct cell activation patterns per region. Hippocampal dorsal CA1 cells were activated both during footshock and at a later reexposure (Double) regardless of conditioning, indicating that the hippocampus preferentially tracks contextual information. In contrast, amygdala cells encode the emotional valence of a context, since cells were activated by both events together only in fear-conditioned but not in control animals. From Zelikowsky and others (2014). Network level: Highly connected nodes in a memory network have the largest impact on memory retention. Chemogenetic silencing of 21 different brain regions that are involved in fear memory after fear conditioning leads to different levels of memory impairment. In line with in silico simulations, silencing regions that are highly connected to other involved brain regions like the thalamic nucleus reuniens (Re) produces the largest performance deficits (light green). From Vetere and others (2017).
The Evolution of the Neocortical Engram
Local Plasticity
After the initial impulse to encode a new memory, a variety of processes is set in motion, which operate on different timescales, and which aim to establish an enduring physiological modification to the neuronal network that represents the new information. Largely, these can be grouped into synaptic processes and systems processes. The first set of mechanisms effects local changes at the synapses between those cells that have been involved in processing the encoded information. They are summarized under the term LTP and its molecular cascades have been described in detail (Lynch 2004). The general idea is that high-frequency stimulation induces short-lasting modifications of synaptic transmission for 30 to 60 minutes (early LTP), for example, by phosphorylation of receptors and receptor trafficking (Herring and Nicoll 2016). Subsequently, protein synthesis sets in (late LTP), which consolidates initial transient modifications in synaptic efficacy through morphological changes to the synaptic spines, representing a first sign of structural plasticity (Lynch 2004). Preexisting small spines can start growing within the first 5 minutes after stimulation (Matsuzaki and others 2004) and new spines can form within 30 minutes (Engert and Bonhoeffer 1999). For example, during optokinetic adaptation, the number of glutamatergic AMPA receptors and the corresponding increase in synapses in Purkinje cell connections of the cerebellum represent a short-term and a long-term engram, respectively (Wang and others 2014). Eventually, also axon collaterals undergo small- and large-scale plastic changes in response to learning (Stettler and others 2006; van Kerkoerle and others 2018). In non-invasive human experiments, synaptic plasticity can only be observed indirectly. Rapid adaptation of functional brain responses during learning within minutes and changes in microstructural tissue properties within an hour after learning can be seen as indications for early and late synaptic plasticity, respectively (Brodt and others 2016; Brodt and others 2018; Keller and Just 2016; Sagi and others 2012; Tavor and others 2020).
Another exciting set of changes has been discovered more recently. These occur at a later timepoint and entail a refinement of connectivity by changes in myelination. Learning has been shown to induce oligodendrocyte precursor cell proliferation and differentiation into oligodendrocytes, leading to new myelin formation within several weeks. Inhibiting proliferation impairs remote but not recent memory, speaking for a role of myelination in later stages of memory consolidation (Pan and others 2020; Steadman and others 2020). In humans, there is also evidence that microstructural changes in white matter occur after several weeks of motor skill training (Scholz and others 2009; Taubert and others 2010). The regulation of myelin influences axonal conduction velocity and might serve to optimize synchronization of information processing between neurons (Fields 2015). Besides changes in the myelin-forming oligodendrocytes, learning-induced activation of astrocytes might also constitute a mechanism for tailoring information exchange between different sites by promoting synchronized oscillatory states relevant for memory formation, such as slow and gamma band oscillations (Santello and others 2019).Together, these findings show that processes modulating neuronal efficacy occur everywhere in the brain at time scales between seconds and weeks after learning. They can not only be directly observed in animals but can also be inferred from studies using novel non-invasive human imaging methods.
Large Scale Reorganization
Apart from cellular and synaptic processes of memory consolidation, a second factor is crucial for the evolution of the neocortical engram. Since the investigations on patient H.M., it is clear that information can be stored in different locations in the brain, and that the contribution of these locations can change over time: whereas memories of more recent events were lost after removal of the hippocampus, more remote ones were kept intact (Scoville and Milner 1957). This observation led to the development of theories of systems consolidation. These theories assume in one form or another that the hippocampus is a fast learning system, which is able to rapidly encode and temporarily store large amounts of information in the form of an index that binds together all neocortical ensembles activated during encoding (Teyler and DiScenna 1986). On the other hand, the neocortex is supposed to be a slow learner, requiring many (hippocampus-driven) repetitions of the to-be-learned information, but eventually providing stable and permanent storage (Frankland and Bontempi 2005; McClelland and others 1995). It is a robust finding in lesion studies that the hippocampus is involved in the acquisition of declarative memories, which are the memories for facts and events (Spiers and others 2001; Tse and others 2007). Human neuroimaging studies also have confirmed a role of the hippocampus during encoding (Danker and others 2017; Himmer and others 2019; Kim 2011; Spaniol and others 2009). However, there is evidence that new declarative information can be learned to a certain degree even in the absence of an intact hippocampus. This was first observed in patient H.M., who used some words that entered language only after his hippocampal lesion and who was able to reproduce the layout of a house in which he was living after his lesion (Corkin 2002). Likewise, in an experimental study, four amnesic patients were able to acquire certain terms as descriptions for tangram figures, even at the same learning rate as healthy controls (Duff and others 2006). Rats have also been shown to acquire contextual fear in the absence of a hippocampus (Wiltgen and others 2006). As contextual fear can be abolished by post-learning lesions of the hippocampus, these findings indicate that the brain’s memory systems have a certain redundancy and that similar memories can be acquired by different memory systems depending on the initial conditions. However, the extent of similarity between representations of the same memory within different memory systems remains a highly debated topic and is one of the main distinctions between different theories of systems consolidation. The model of complementary learning systems in its original version assumed that identical information is transferred from the hippocampus to the neocortex over time (McClelland and others 1995) with the mPFC taking over the indexing function of the hippocampus (Frankland and Bontempi 2005). The current version of multiple trace theory, trace transformation theory (Winocur and others 2010), instead proposes that the reactivation of detailed, precise hippocampal traces results in an abstracted, semanticized version in the neocortex and that these two versions can coexist in parallel. It is even conceivable that completely parallel encoding and storage in neocortical and hippocampal systems occurs.
Recent studies on engram formation in the rodent brain have revealed that engram cells are established in the neocortex rapidly during initial training (Kitamura and others 2017) and that they carry enough information to enable recall. Specifically, optogenetic reactivation of retrosplenial or mPFC engram cells for contextual fear sufficed to activate the memory and induce freezing behavior even when the hippocampus was inactivated (Cowansage and others 2014; Kitamura and others 2017). However, memory recall under natural conditions required an intact hippocampus for several days after learning. Thus, neocortical engram cells must be tagged already shortly after encoding although they initially remain in a “silent” state. This silent neocortical engram requires the hippocampus to become “mature” and able to induce natural recall independently. Hippocampal engram cells, in contrast, become silent over time (Kitamura and others 2017). Together, these findings indicate that the interplay of different memory systems is much more flexible than previously thought. They fit very well with a model in which several engrams start to develop in parallel on encoding of new information (Tse and others 2011). Still, hippocampal and neocortical engrams will generally represent different aspects of a memory because of their differing basic computational properties. The hippocampus can store individual events sparsely by performing pattern separation, whereas the neocortex rather learns generalized patterns by extracting regularities from overlapping similar instances (Bakker and others 2008; Kumaran and others 2016). These substantially differing network properties imply that the strength of the respective engrams will benefit from different factors and that their development will diverge and become increasingly independent.
Although the idea of systems memory consolidation has been elaborated in greatest detail for declarative memory, there is some evidence that other types of memories make use of similar principles. In the domain of motor learning, the contributions of different neocortical areas, for example, the primary motor cortex, motor association areas and the posterior parietal cortex, vary considerably between the early, fast and the later, slow learning phases (Dayan and Cohen 2011). Also, some forms of motor sequence learning can use either hippocampus or striatum, and the interaction between these regions can evolve during a consolidation period (Albouy and others 2008). How this substantial reorganization is effected however is still not well understood.
How to Detect Engrams in the Human Brain
With the development of new cell labeling techniques and optogenetics, animal research is now able to specifically label engram cells that are active during memory acquisition, track them over extended time periods, and precisely manipulate them on the millisecond scale. By conducting invasive loss-of-function studies that disrupt memory by temporarily inactivating or permanently lesioning engram cell ensembles, it was possible to show that the underlying neuronal circuits are necessary for engram formation and expression. On the other hand, gain-of-function studies can induce artificial engrams and thus indicate processes and structures that are sufficient for engram formation (Josselyn and Tonegawa 2020; Martin and others 2000). In contrast, most human research relies on observational studies that detect the co-occurrence of memory with changes in neuronal activity and structure that fulfill the engram criteria specified above.
With methods that allow us to observe task-related functional activity, like functional magnetic resonance imaging (fMRI), electroencephalography (EEG), or magnetoencephalography (MEG), we can detect brain areas that are engaged during memory encoding or retrieval. Learning-induced changes in functional responses can pinpoint areas that undergo short- or long-term modifications. For example, in an fMRI study on spatial maze learning, increasing activation was induced in the posterior neocortex over 60 minutes of learning, but not in a similar control condition that precluded memory formation by constantly changing the maze layout outside the participants’ field-of-view. This learning-induced increase in activity during navigation was stable after 12 hours and correlated with memory performance, thus fulfilling the first three criteria for an engram by Semon’s definition (Brodt and others 2016). Other studies, using resting-state fMRI, have shown changes in functional connectivity after learning that were associated with later performance, providing an additional means to investigate consolidation processes (Guidotti and others 2015; Stevens and others 2010; Wagner and others 2019).
Recently, multivariate methods were introduced that allow decoding the information processed by the brain from activity patterns. They enable us to assess whether a brain region not only performs processing that supports a memory task but also actually holds the memory content itself (Kunz and others 2018). In a correlational approach, learning-related changes in the similarity of stimulus-specific activity patterns can be linked to the gradual formation of engrams (Schlichting and others 2015; Tompary and Davachi 2017). Such analyses have, for example, shown that the representational similarity of events in the anterior hippocampus corresponds to their spatial and temporal proximity (Nielson and others 2015). In another approach, a machine learning classifier is trained to distinguish two or more classes of stimuli on the basis of their evoked functional activity patterns (Kuhl and Chun 2014; Polyn and others 2005). If the classifier is able to perform this task in a specific brain region, this region must process stimulus-associated information. In addition to using functional patterns to show online memory processing during learning and retrieval, they can also be used to show off-task memory processing, for example, during spontaneous reactivation in sleep or resting wakefulness (Deuker and others 2013; Schönauer and others 2017).
Is it enough to show that learning leads to an enduring, stimulus-specific, recall-enabling change in the functional brain response to also show local memory storage? Typically, functional methods measure the online response to a given stimulus. In studies of memory this represents the activity during encoding or recall. We can assume that memory storage occurs in the same neural circuits in which stimuli are processed (see Box 1). Therefore, functional activity can indicate the activation of a memory engram. However, we have to exclude the possibility that it represents other, auxiliary processes. Attention-regulating regions might show functional responses to stimuli without actually processing them. To detect an engram, we must therefore show that a functionally active region is indeed processing memory-related information, which we can do with the information-based multivariate methods mentioned above. In addition, engram encoding must manifest in a change in the functional response over the course of memory acquisition, which is an indication of an underlying change in the neuronal substrate. This change must be stable over a period during which no active processing occurs to exclude active rehearsal, which represents (working) memory without an engram, and fatigue, which represents a transient neuronal change. It has to be kept in mind, however, that fMRI activity reflects the input into a cortical region rather than its output (Logothetis and Wandell 2004). Especially in the case of long-range communication between brain areas, the interpretation of the actual location of the engram based on functional activity alone is therefore sometimes not obvious. Thus, while activity indicates the presence of an engram, the location of that engram can only be determined by finding the related structural changes.
The Organization of Memory in the Brain.
Over the decades, different ideas of how widely a memory is distributed across the brain were put forward. Memory was assumed to be stored in highly localized, sparsely distributed, or extensively dispersed, interconnected systems, depending on the theoretical approach. The extreme form of localized storage is represented by the idea of the “grandmother cell,” a cell responding if and only if my grandmother crosses my thoughts. This concept has also been discussed in terms of “pontifical cells,” individual cells which have the final authority of defining a specific concept (Barlow 1972). Cells with such properties were reported to exist recently (Quiroga and others 2005). These cells responded with high specificity to individual concepts (e.g., that of the actress Halle Berry) no matter how they were presented (e.g., as different images or as a string of letters). Also, the idea of functional brain mapping relies on the proposition that individual aspects of cognition can be precisely pinpointed. The opposite end of the spectrum from local to distributed storage has been advocated by Karl Lashley (1950), who by lesioning rat brains came to the conclusions that all neocortical regions are functionally equivalent (“equipotentiality”) and only the amount of lesioned tissue determines the extent of functional impairment (“mass action”). Within this framework, memories are stored in the brain similar to information in a hologram. Cutting a hologram in two pieces does not split the scene in two, but gives two full representations, albeit with less sharpness or a restricted viewing angle.
Our current view reconciles both the localist and the connectionist standpoints. We know that there is a functional specialization of regions in the brain, beginning from retinotopic, somatotopic or tonotopic representations of the environment in the primary sensory cortices and leading up to specialized areas for complex concepts such as faces, objects, or places in higher-order association regions (Martin 2007). However, information is never stored in a single location alone, regardless of the level of inspection. Even low-level sensory stimuli are perceived by a number of neurons with varying tuning curves (Butts and Goldman 2006). Complex concepts activate large, distributed networks across the brain, comprising all lobes of the neocortex (Wang and others 2010). To integrate both points of view, it seems reasonable to assume that concepts are stored in the brain as a hierarchy of localized, functionally distinct nodes, each consisting in turn of a number of other, more basic concepts, possibly descending down to the level of simple perceptual features. For a full understanding of engrams, it is therefore vital to combine the primarily cellular and the primarily network perspectives taken by animal and human research, respectively.
The representational hierarchy of memory is also reflected in the idea of temporally receptive windows (Lerner and others 2011). Analogous to spatially receptive fields in the visual system, it has been shown that along the cortical hierarchy, areas are able to accumulate and retain information on increasingly longer timescales, from several tens of milliseconds in primary sensory areas up to several minutes in higher-order association areas like the medial posterior parietal cortex (mPPC) and the medial prefrontal cortex (mPFC; Hasson and others 2015). In humans, this has been elegantly shown with functional neuroimaging of participants that were either listening to a narrated story or viewing a movie (Baldassano and others 2017; Lerner and others 2011). Importantly, event segmentation according to the intrinsic timescale of higher-order association areas re-occurs during recall and even shows anticipatory reinstatement when reexposed to the same narrative again, indicating that temporally receptive windows might constitute an inherent principle not only of stimulus processing, but also memory storage (Baldassano and others 2017).
Thus, it becomes clear that the brain is—unlike a computer–not divided into separate processing and storage units. Instead, information is stored in the same neural circuits that participate in its initial processing, and also at the same level of conceptual abstraction (Fuster 1997; Martin 2007). In fact, it is this processing of stimuli that induces neuronal changes that represent memories, for example, in the form of LTP. The same idea is also reflected in the observation that—again unlike a computer—the brain does not seem to have a location-independent, general-purpose code for representing information. If, accordingly, meaning and location cannot be separated, information cannot be “transferred” between brain regions without being processed, transformed, abstracted, or integrated. Thus, it is reasonable to assume that a memory with a specific content must be stored where this content is processed during online stimulation. Functional brain activity can therefore be conductive in locating the memory engram.
Changes in functional activity provide only indirect evidence of a memory engram. To reveal the actual structural change induced by new memory encoding, other methods are needed. Using structural MRI, studies in humans in the last two decades have focused on long-term brain changes induced by extensive training. Usage of a brain area over a long period, for example, during the acquisition of new knowledge or a new skill, can increase the macroscopic extent of brain regions in a way that is detectable using voxel-based morphometry. Thus, it has been shown that the hippocampal gray matter volume of taxi drivers increases during learning of spatial information (Woollett and Maguire 2011). Also, training a complex motor skill like juggling increases gray matter density in the posterior neocortex (Draganski and others 2004; Scholz and others 2009).
Another approach allows mapping of more fine-grained, microstructural changes in the brain using diffusion-weighted MRI (DW-MRI). DW-MRI measures the random Brownian motion of water molecules along different gradient directions, which is sensitive to tissue microstructure. Using DW-MRI, it has been shown that several weeks of training lead to noticeable changes in white matter tracts (Scholz and others 2009; Taubert and others 2010). Only recently, it has been shown that DW-MRI is able to also detect rapid, experience-driven structural plasticity in human gray matter as early as 90 minutes after learning (Brodt and others 2018; Keller and Just 2016; Sagi and others 2012; Tavor and others 2020). These structural changes were assessed via a decrease in mean diffusivity (MD), which is a non-directional measure of the average amount of diffusion within a voxel. A decrease in MD over time indicates a higher substance density within that voxel. Histological findings from animal studies show that sites of learning-induced MD decrease also display an increase in synapse number, expression of brain-derived neurotrophic factor, as well as astrocyte activation (Sagi and others 2012). Critically, all of these potential underlying mechanisms are tightly linked to learning-induced microstructural plasticity. Besides neuronal processes like expansion of post-synaptic densities and dendritic spine formation (Matsuzaki and others 2004), astrocytes also play a key role in learning and memory, as astrocytic signaling supports BDNF-dependent LTP and induces synapse formation (Santello and others 2019; Vignoli and others 2016). Therefore, MD is a highly useful marker of memory engrams: A learning-induced change in MD that remains stable over an extended period and that relates to successful memory recall indicates the presence of an engram. Generally, while functional methods have their strengths in uncovering the spatiotemporal patterns of mnemonic processing and thus enable us to assess stimulus specificity and relation to recall performance on a trial-by-trial basis, the advantage of microstructural imaging is that it provides evidence for structural changes and a possibility to observe the engram during offline phases. Therefore, both methods combined provide a comprehensive set of tools to detect and study engrams in both their active and their dormant states (Fig. 2).
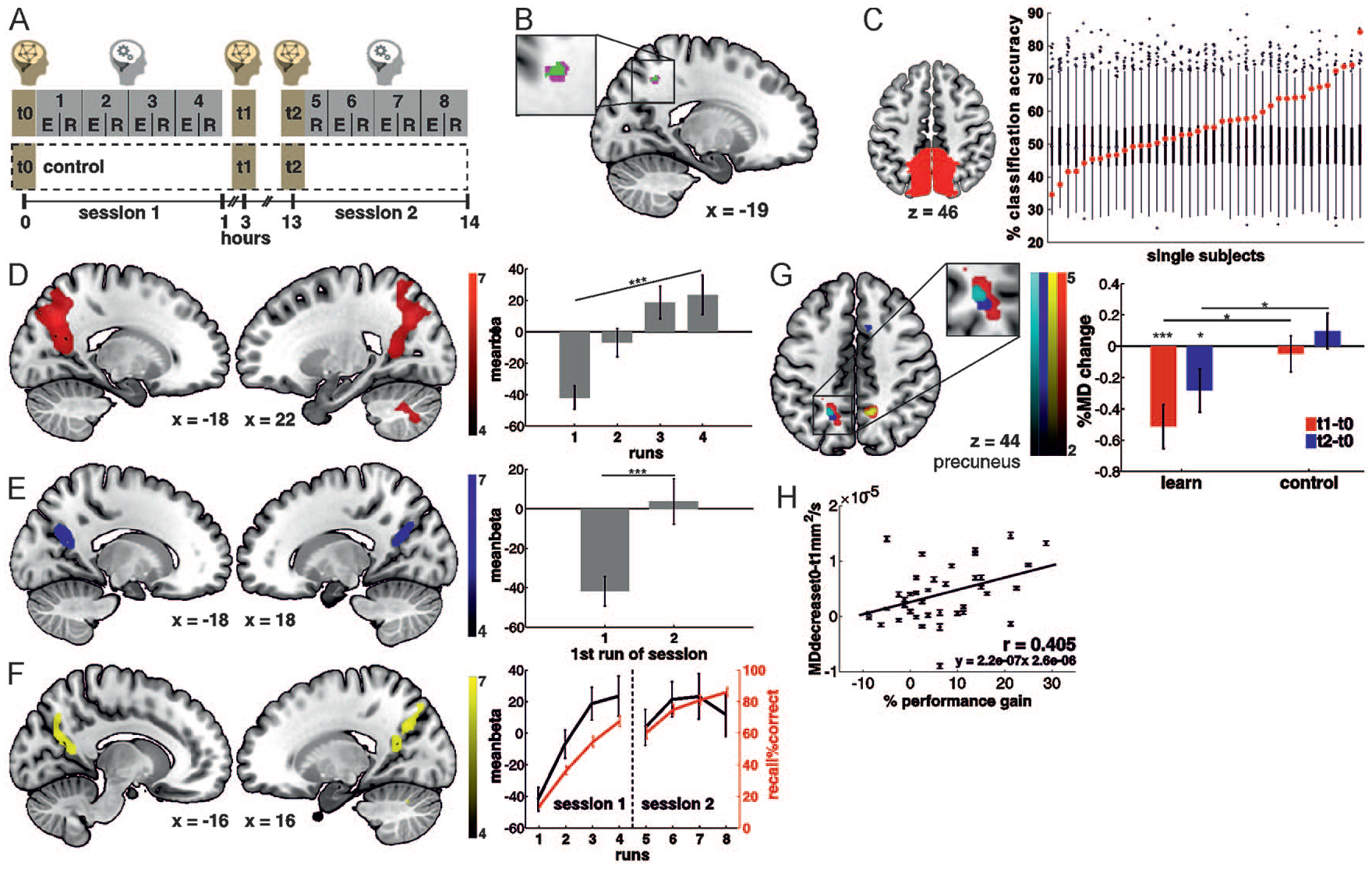
Detecting human engrams with combined functional magnetic resonance imaging (fMRI) and diffusion-weighted MRI (DW-MRI). (A) Participants learned object-place associations in several encoding-recall repetitions over the course of 13 hours. Task-related activity was recorded with fMRI and tissue microstructure was assessed at three time points, at baseline immediately before learning, 90 minutes after learning and again after an offline interval of 10 hours. The control condition followed the same protocol without learning. (B) A conjunction analysis of the minimum statistic across functional and microstructural data shows that the human precuneus fulfills all criteria for a memory engram. (C-F) fMRI analyses. (C) In an anatomical precuneus region of interest (ROI), the category of the presented stimuli could be decoded from functional activity during memory encoding, indicating that this region processes the learned content. (D) Increases in precuneus activity over the encoding-recall repetitions were (E) stable over an offline period of 12 hours and (F) related to successful memory recall. (G-H) DW-MRI analyses. (G) A decrease in mean diffusivity 90 minutes after learning was experience-dependent as it was specific to the learning condition, persisted over an offline interval of 10 hours and correlated with memory retention. From Brodt and others (2018).
The Location of the Neocortical Engram
Studies on memory engrams in rodents have mostly investigated contextual fear conditioning (Josselyn and Tonegawa 2020). Although many of these studies have focused on the hippocampus and the amygdala, engram cells have also been found distributed throughout the neocortex (de Sousa and others 2019; Tonegawa and others 2018; Vetere and others 2017). In humans, studies of the engram can be divided into those investigating declarative memory and those studying non-declarative memory. Whereas studies on non-declarative memory mostly investigate the engram in modality-specific association areas, studies on declarative memory find evidence for an important role of a number of hub regions, which belong to a network of higher-order association cortices often subsumed as the default mode network (Buckner and DiNicola 2019; Fig. 3). Apart from the hippocampus, especially two neocortical hubs along the cerebral midline have been identified as central to this network: the mPFC and the mPPC, which encompasses the precuneus, posterior cingulate cortex and retrosplenial cortex.
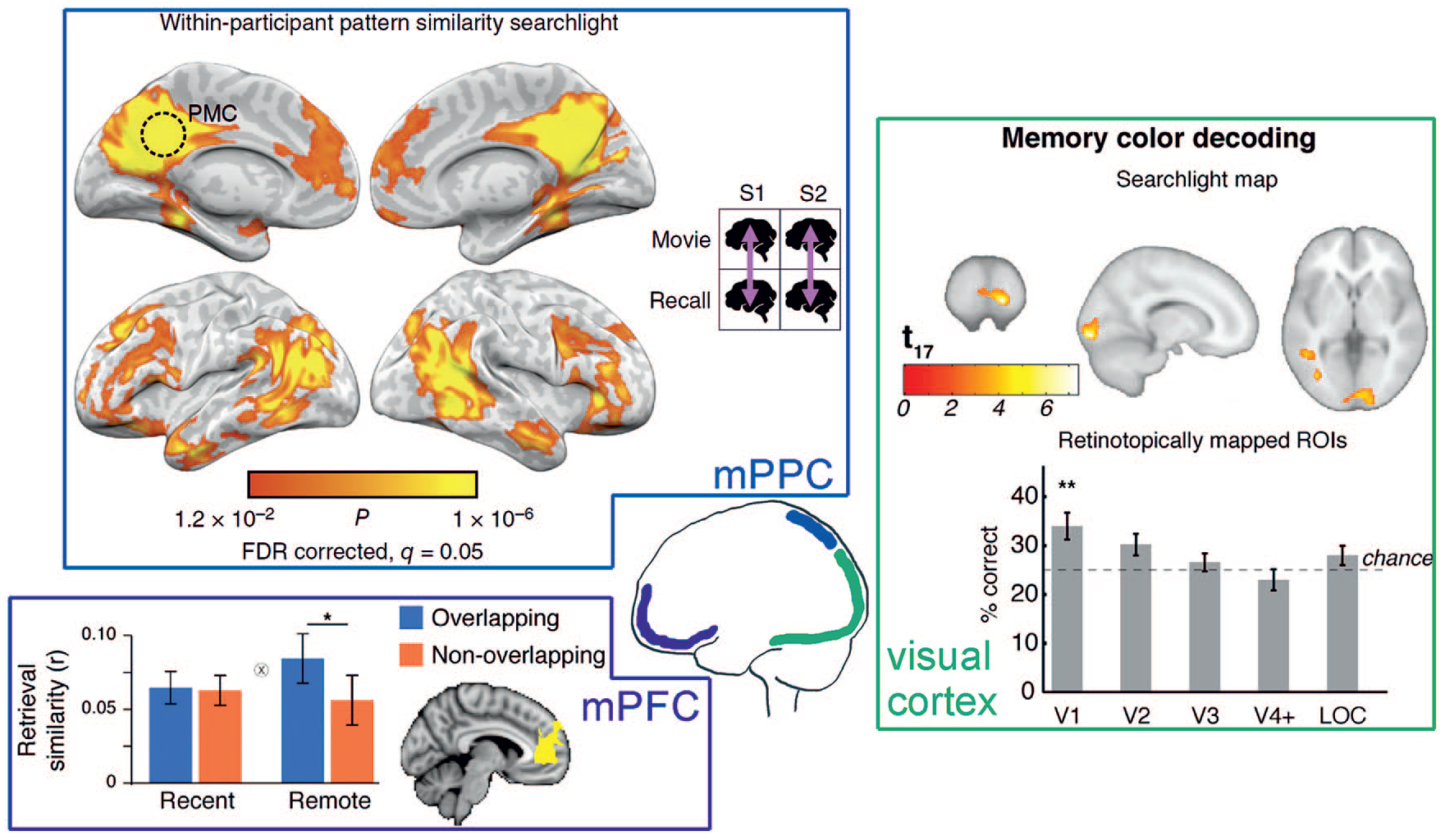
Potential locations of human memory engrams in the neocortex. Medial posterior parietal cortex (mPPC): When comparing functional activity recorded during watching movie scenes and during later verbal recall within the same subject, the mPPC was one of the regions displaying the highest pattern similarity. A similar pattern also emerged for between subject analyses (not shown), indicating that the mPPC stores modality-free abstract information that is similarly structured over individuals. From Chen and others (2017). Medial prefrontal cortex (mPFC): The mPFC exhibits greater pattern similarity of objects that were encoded on separate trials but within the same scene (overlapping) than of objects that were encoded in different scenes specifically during memory retrieval after 1 week (remote), but not during immediate retrieval (recent). Thus, memory representations in the mPFC seem to change with consolidation toward stronger representation of commonalities between stimuli. From Tompary and Davachi (2017). Visual cortex: It is possible to decode the color from functional brain activity while seeing a grayscale image of a well-known colorful object from V1, like, for example, the color “yellow” from a gray banana. These results show that memory recall involves the reactivation of sensory features in early sensory areas and might hint at a memory engram in the visual cortex. From Bannert and Bartels (2013).
One of the main functions traditionally ascribed to the prefrontal cortex is executive control (Miller and Cohen 2001). It plays a central role in top-down modulation and behavioral inhibition (Munakata and others 2011), which allows it to support goal maintenance and attention (Miller and Cohen 2001). The latter two functions have also been suggested to be crucial for the role of the prefrontal cortex in working memory (Gazzaley and Nobre 2012), in which the mPFC collaborates with the hippocampus for maintenance intervals that are longer than a few seconds (Lee and Kesner 2003). As to the role of the mPFC in long-term memory, most current theories of systems consolidation describe it as a region that takes over the function of the hippocampus in binding cortical information modules into a coherent representation of a memory after consolidation (Frankland and Bontempi 2005). Consequently, there have been many studies investigating the role of the mPFC for remote memory. Engram cells for remote contextual fear memory were found in the prefrontal cortex of rats (Kitamura and others 2017) and human studies have shown increased mPFC activation during retrieval of remote versus recent memories (Gais and others 2007; Takashima and others 2006). Some of these human imaging studies might be interpreted in light of an increased need of executive strategy during later memory recall instead of direct engram activation. However, the delayed involvement of the mPFC might also be explained by the fact that this region seems specialized for processing abstract, schema-like knowledge (Gilboa and Marlatte 2017; van Kesteren and others 2010). During categorization tasks, the mPFC preferentially represents prototypes of learned categories instead of single exemplars (Bowman and Zeithamova 2018); and online memory representations of experimentally linked objects in mPFC become more similar over time (Tompary and Davachi 2017). Also, sudden, insight-like encoding of semantic memories has been shown to depend on the mPFC in addition to parietal areas (Kizilirmak and others 2019). In any case, mPFC activity is often observed in synchrony with the hippocampus. For example, during overlearning of word-color associations, both hippocampus and mPFC became less active while posterior parietal areas increased activity (Himmer and others 2019). This idea is also supported by the strong connections between both regions, the bilateral information flow, and their electrophysiological oscillatory synchrony in the theta band (Eichenbaum 2017). Thus, it is possible that the mPFC is a region that is processing and storing new information as more abstract knowledge like concepts, schemata, or task rules (Gilboa and Marlatte 2017; van Kesteren and others 2010). Initial encoding might already start in parallel to the hippocampus and full engrams develop through reactivation afterward (Euston and others 2012). It must be kept in mind, however, that the mPFC is a heterogeneous region that might serve a number of different functions in different subregions (Ciaramelli and others 2019).
In contrast to the mPFC, the posterior parietal cortex received attention in the context of memory only recently, and evidence for a genuine memory function not only of the mPPC but also of the inferior and superior parietal lobules, has only begun to accumulate (Gilmore and others 2015). Functional neuroimaging studies in humans have shown the involvement of this region during encoding and retrieval of mnemonic information, in particular denoted by an increase in its functional response over learning (Brodt and others 2016; Brodt and others 2018; Himmer and others 2019; Schott and others 2019). Patterns of encoding activity in mPPC are reinstated during retrieval and the strength of reinstatement correlates with behavioral performance (Bird and others 2015; Chen and others 2017). Several recent fMRI studies using different material have demonstrated that changes in memory-related activity in mPPC are experience-dependent, long-term stable and relevant for behavior, thereby pointing to an online memory representation (Brodt and others 2016; Brodt and others 2018; Himmer and others 2019). Additionally, using DW-MRI, rapid microstructural plasticity in the same location as these experience-induced functional changes in the precuneus has been found (Brodt and others 2018). These findings provide evidence for a memory engram in the mPPC and show its role in neocortical memory storage. Additionally, a recent study combining lesion sites of 53 amnesic patients with human connectome data has identified the retrosplenial cortex as the center of a human memory circuit (Ferguson and others 2019), further corroborating the idea that the mPPC constitutes an important neocortical memory hub. Concerning the nature of memory representations in the mPPC, multivariate pattern analyses have confirmed the existence of exemplar-level as well as category-level memory representations within this region (Lee and others 2019). In monkeys, the mPPC has also been shown to simultaneously hold the memory of an abstract task rule as well as spatial sensory information (Gail and Andersen 2006). Both findings fit well with the idea that it plays a role in memory integration, linking episodic detail with pre-existing knowledge (Bird and others 2015; Chen and others 2017). Similar to the mPFC, the mPPC represents a large area of cortex that serves many different functions. We are only beginning to understand which subregions supports which function for what type of memory (Sestieri and others 2017).
In accordance with the idea that the neocortex is best suited to store generalized knowledge, neocortical areas have mostly been associated with the storage of semantic information, in particular those regions (posterior parietal cortex, anterior temporal lobe) at the top of the multisensory processing hierarchies (Binder and others 2009; Gilboa and Marlatte 2017). But, evidence for memory engrams has also been found further upstream in the sensory neocortex, downstream in the motor cortex, and in several other neocortical areas. In the primary visual cortex of adult monkeys, structural changes can be found related to learning a novel contour detection task. Axons in the cortical region representing the trained visual field undergo sprouting and pruning after learning (van Kerkoerle and others 2018). In humans, during recall of common items, the visual cortex holds the information about their color; for example, during the presentation of a gray “banana,” the color “yellow” can be decoded from the primary visual cortex (V1) (Bannert and Bartels 2013). As there is no bottom-up color stimulation, the color information must be coming via top-down feed-back connections. It can be argued that the concept of a “yellow banana” is located upstream of the primary visual cortex, for example, in V4, which has strong similarity in representational structure to V1 (Bannert and Bartels 2013), and V1 is activated as a result of memory retrieval in V4. However, it can also be argued that the concept is located in the whole network of neurons and that activation of V1 is integral to its retrieval. There are a number of other studies that relate retrieval of visual features from memory and V1 activity, not only showing participation of these early areas in the engram but also indicating that processing and storage happen in the same neuronal circuits (Harris and others 2001). Even adaptation-induced plasticity of tuning-curves found in the visual cortex of rats can be seen as a (very basic) engram of visual properties (Dragoi and others 2000). On the other hand, V1 has also been shown to be able to learn complex information like visually presented sequences in humans (Rosenthal and others 2016). Complex visual stimuli like imagined spatial landmarks can be decoded with fMRI from neocortical regions higher up in the ventral processing stream (Boccia and others 2017).
For other modalities, most evidence comes from animal studies. The auditory cortex, analogous to the visual cortex, also seems able to hold specific engrams; for example, fear conditioning of complex tones requires auditory cortex plasticity in rats (Dalmay and others 2019). For olfactory fear conditioning, chemogenetic inactivation and activation shows that olfactory cortex activity is both necessary and sufficient to elicit a conditioned response, thus it holds the engram for this type of memory (Meissner-Bernard and others 2019). In human cross-modal episodic memory with visual and olfactory stimulation, a visual recall cue elicits fMRI activity in the olfactory cortex (Gottfried and others 2004). Interestingly, the modality-specific sensory cortices seem to be required especially for late-stage memory of fear conditioning in rats, indicating that systems consolidation does not only manifest in an interaction between the hippocampus and the hub regions mentioned above but can also involve other neocortical regions (Sacco and Sacchetti 2010). Regarding motor engrams, intracranial recordings in monkeys show that the primary motor cortex is able to learn a stable movement mapping, which can later be used to decode imagined movements and control a neuroprosthetic device, thus fulfilling all criteria for a motor memory engram (Ganguly and Carmena 2009). Targeting and optogenetically shrinking an estimated 400,000 synapses in the rat motor cortex that were previously potentiated during motor skill learning can erase a corresponding engram (Hayashi-Takagi and others 2015). Motor engrams have also been found in the cerebellum (Wang and others 2014), and even corticospinal neurons undergo plasticity after learning, showing a very low-level neuronal involvement in the motor engram (Peters and others 2017). It remains to be seen whether the motor output system has a similar hierarchical structure of motor concepts as the semantic system has for sensory concepts.
Taken together, these findings support the idea that neocortical memory traces are distributed broadly throughout task-relevant regions (Eichenbaum 2016) and that engram ensembles in different regions are specialized to store different aspects of an engram (Josselyn and Tonegawa 2020; Zelikowsky and others 2014). They also fit very well with accounts proposing the same distributed neural circuits for processing and storage of information (Fuster 1997; Hasson and others 2015; Martin 2007) (see Box 1). Certainly, one of the most interesting open issues in this context is the question how memory is implemented on the network level. Both the mPFC and the mPPC constitute hubs which are located at the top of the cortical processing hierarchy (Binder and others 2009; Gilboa and Marlatte 2017; Margulies and others 2016). Their dense connections to cortical and subcortical circuits make them most suited on the one hand to communicate with other memory systems and on the other hand to orchestrate engram ensembles in downstream cortical areas. For example, for schema-related information, the (ventral) mPFC is assumed to activate relevant neocortically stored information and to inhibit hippocampal encoding while enabling the establishment of intracortical connections for direct integration of the new schema memory into the neocortical network (Ciaramelli and others 2019; van Kesteren and others 2012). Likewise, for spatial information, it has been shown that the mPPC displays increased functional connectivity with occipitotemporal cortices during memory encoding (Cohen and others 2019; Schott and others 2019) as well as modified connectivity patterns with the hippocampus across memory formation (Brodt and others 2016).
Factors Supporting Neocortical Memory Formation
Reactivation
During systems consolidation, the contribution of individual memory systems to an engram undergoes a change. The main proposed mechanism underlying this gradual shift is an off-line reactivation of the neuronal ensembles that were activated during initial encoding (Frankland and Bontempi 2005), which allows separate memory systems to interact (Helfrich and others 2019; Ji and Wilson 2007). Spontaneous reactivation of previous experience has been shown in many neocortical regions, from higher association cortices down to sensory and motor regions (Klinzing and others 2019). As predicted from consolidation theories, neocortical reactivation is often, but not exclusively observed in coordination with the hippocampus (Girardeau and others 2017; Ji and Wilson 2007; O’Neill and others 2017; Rothschild and others 2017). Artificially enhancing hippocampal-neocortical coupling promotes neocortical storage and subsequent memory performance (Maingret and others 2016). However, the original idea that there is a unidirectional flow of information from hippocampal to neocortical memory stores has recently been called into question (Helfrich and others 2019; Rothschild and others 2017).
Many studies have shown that there is spontaneous reactivation during sleep (Buzsaki 2015). Indeed, sleep is known to promote cortical plasticity and benefit subsequent behavior (Aton and others 2013; Kuhn and others 2016; Niethard and others 2018; Yang and others 2014). In animals, it has been shown that sleep stabilizes cortical immediate-early gene upregulation and leads to subsequent dominance of neocortical memory traces (Genzel and others 2017). In humans, sleep leads to a reorganization of cortical activity in declarative and motor memory systems (Gais and others 2007; Sterpenich and others 2009; Vahdat and others 2017). Sleep has also been associated with a decrease in retrieval-related hippocampal activation (Takashima and others 2006) and the stabilization of a repetition-induced hippocampal deactivation (Himmer and others 2019). All of these findings indicate less reliance on the hippocampus and an increased dependence of memory retrieval on neocortical networks after sleep. On a behavioral level, sleep-dependent reorganization leads to enhanced category learning, rule abstraction and memory integration, features typically linked to neocortical representations (Ashton and others 2018; Durrant and others 2011; Nieuwenhuis and others 2013). Apart from during sleep, a growing amount of literature also reports spontaneous reactivation in the neocortex during resting wakefulness, with similar beneficial consequences on subsequent memory performance (Tambini and Davachi 2019). For example, in a recent human fMRI study it has been shown that after participants engaged in a decision-making task, the hippocampus replays sequences of task states during awake rest. This replay correlated with the fidelity of task representations in the neocortex, specifically the orbitofrontal cortex, which in turn was positively related to task performance (Schuck and Niv 2019).
Recent work supports the idea that just like spontaneous endogenous reactivation, also stimulated reactivation in the form of repeated learning or repeated retrieval, might provide a benefit particularly for neocortical memory (Antony and others 2017; Karpicke and Roediger 2008). Accordingly, animal studies show that increased training intensity renders fear memories independent of the hippocampus within a much shorter timeframe than commonly observed (Lehmann and others 2009; Pedraza and others 2016), speaking for accelerated neocortical memory formation. In line with these findings, human neuroimaging studies observe a decrease in hippocampal activity and hippocampal-neocortical connectivity over repeated encounters with previously learned information (Brodt and others 2016; Himmer and others 2019). At the same time, repetitions enhance neocortical activity, particularly in posterior parietal areas, and this activity correlates with subsequent memory performance (Brodt and others 2016; Brodt and others 2018; Himmer and others 2019; Kremers and others 2014; Vannini and others 2013). Compared with simple passive restudy, especially repeated active retrieval of learned content has been linked to neocortical memory formation. Repeated retrieval sharpens cortical memory representations (Karlsson Wirebring and others 2015), benefits associated memories potentially via co-activation in parietal cortex (Jonker and others 2018) and leads to adaptive forgetting via cortical pattern suppression of competing traces (Wimber and others 2015). At the same time, retrieval also promotes neocortical memory integration (Ferreira and others 2019; Lee and others 2019).
Thus, repeated memory retrieval as well as spontaneous reactivation leads to restructuring of the engram and benefits memory performance. Both seem to be able to provide the repetitions the neocortex requires to encode new information. Systems consolidation therefore seems to be a process depending not on passage of time, but on reactivation and repetition. A particular advantage that both retrieval and endogenous reactivation might have over repeated encoding is that they both represent a top-down flow of information and they might thus close a loop with the bottom-up pathway established during learning. Together, these findings argue that the neocortex should rather be considered a “multi-trial” instead of a “slow” learner.
Prior Knowledge
Integration of new information into prior knowledge is a powerful tool to accelerate learning of related information and to promote memory retention in experimental settings as well as in real life. The method of loci for example is a mnemonic device that enables rapid encoding of large amounts of novel information by mentally linking each item to a specific location along a well-known route. Animal studies have shown that rats can integrate location information into an existing spatial map in a single trial whereas they need training over several sessions to learn the same location without a pre-existing spatial schema. Moreover, the rat neocortex supports this novel schema-related information independent of the hippocampus within 48 hours, which can take months without prior knowledge (Tse and others 2007). Because this type of fast learning elicits upregulation of immediate early genes in neocortical structures within 80 minutes and because an intact neocortex at the time of encoding is necessary for long-term memory retention, this finding speaks for a direct parallel encoding into neocortical networks (Tse and others 2011). In humans, it was found that the existence of a schema increases connectivity between the hippocampus and neocortical regions during encoding of related information (van Kesteren and others 2010). Furthermore, as prior knowledge increases, retention of related information depends less and less on hippocampal activity at encoding, indicating increased reliance on neocortical networks (Brod and others 2016). Along the same lines, brain activity during retrieval of schema-related novel information shifts much more rapidly from the hippocampus to the neocortex than for schema-unrelated information (Sommer 2017). Finally, schema learning has been shown to induce neocortical structural changes in the form of myelin formation in mPFC (Hasan and others 2019).
Another line of evidence on how prior knowledge influences memory storage in humans comes from studies employing the paradigm of fast mapping. Here, the name of an unknown item can be inferred through elimination because it is presented together with a well-known item. Subsequent memory for information that is encoded with fast mapping can be best predicted from activation patterns in the anterior temporal lobe, a neocortical region associated with semantic knowledge, whereas the hippocampus is the best predictor if the same material has been explicitly learned (Atir-Sharon and others 2015). Similarly, retrieval of fast mapped compared with explicitly encoded information preferentially engages the anterior temporal lobe (Merhav and others 2015) and patients with damage to this area are impaired at fast mapping whereas patients with hippocampal lesions are impaired at explicit learning (Sharon and others 2011). The pattern of associated behavioral performance suggests that memory is successfully integrated into existing memory networks within 10 minutes after fast mapping, whereas it takes more than a day after explicit learning (Coutanche and Thompson-Schill 2014). Together, prior knowledge research suggests that novel information can rapidly become stored in neocortical memory networks when linked to an existing neocortical anchor (Kumaran and others 2016).
In sum, reactivation and pre-existing neocortical representations benefit neocortical memory formation (Fig. 4). Additionally, several findings indicate that these two factors interact. For one, post encoding reactivation in neocortex contributes to the emergence of schematic representations and facilitates subsequent encoding of related information (Lewis and Durrant 2011; Tompary and Davachi 2017). Prior knowledge, in turn, not only modulates hippocampal-neocortical interaction during post-encoding rest (van Kesteren and others 2010) but also reorganizes the content of reactivation (Liu and others 2019).
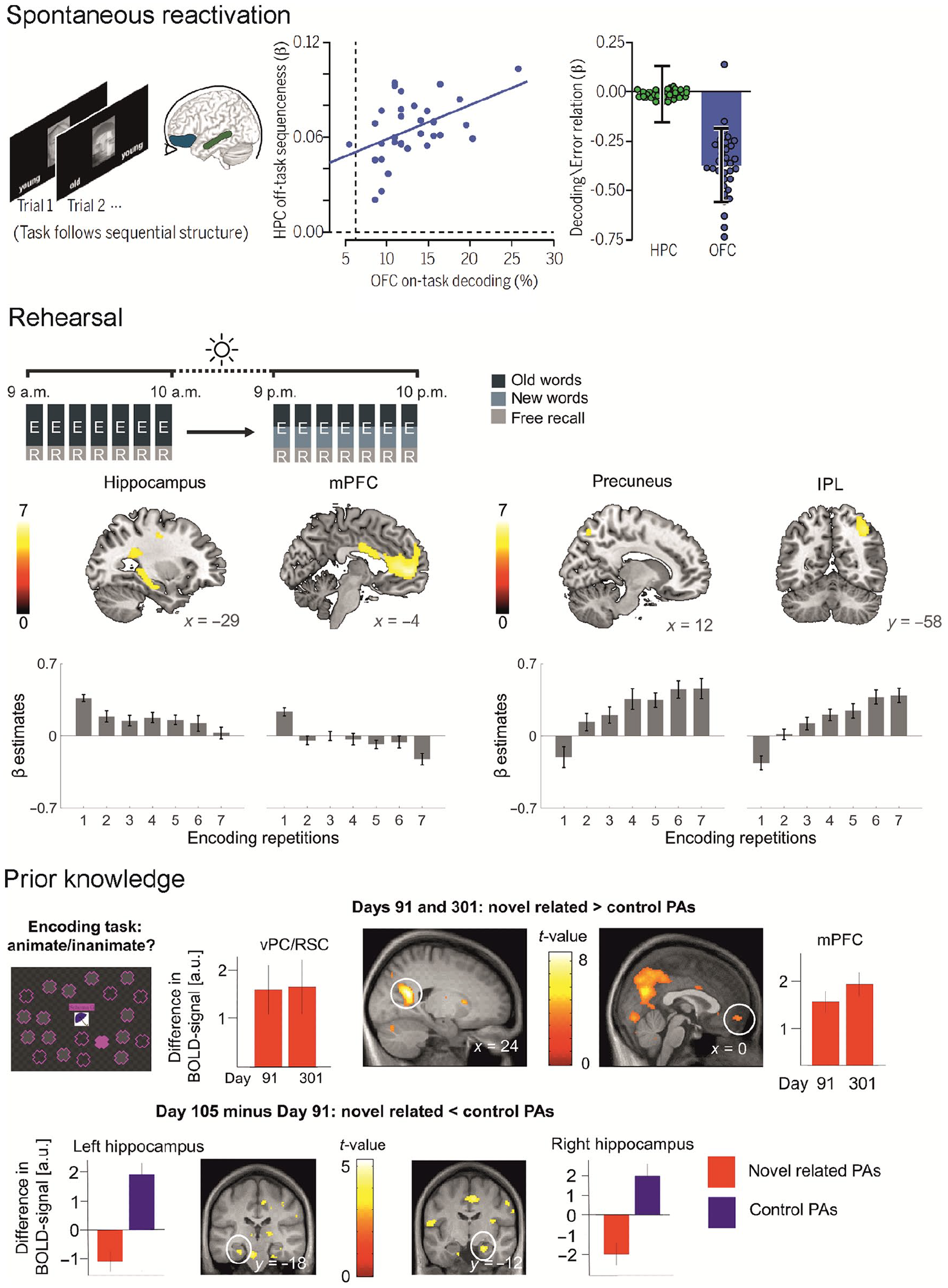
Beneficial factors for neocortical engram formation. Spontaneous reactivation: After participants were engaged in a decision-making task, sequential replay of the task structure could be observed with functional magnetic resonance imaging (fMRI) in the hippocampus during awake rest (left panel). Strong sequential offline replay in the hippocampus (HC) was related to higher decodability of task states in the orbitofrontal cortex (OFC, middle panel). Higher decodability in OFC, but not in HC, was associated with lower error rates (right panel). This pattern indicates a role of hippocampal replay for neocortical task representations that benefit behavior. From Schuck and Niv (2019). Rehearsal: Over multiple encoding-recall repetitions of a word list, hippocampal and mPFC activity decreases, whereas activity in the precuneus and the inferior parietal lobe increases indicating stable learning-dependent changes in these areas. From Himmer and others (2019). Prior knowledge: In an fMRI study on object-location schema learning with multiple sessions over 1 year, different activity patterns for novel but schema-related paired associates (novel PA) and novel unrelated stimuli (control PA) could be observed. Specifically, novel related PAs elicited higher activity than control PAs in neocortical regions like the ventral precuneus (vPC) and retrosplenial cortex (RSC) during encoding (not shown) as well as immediate recall (upper right panel) at later timepoints when the schemata were well established. In contrast, hippocampal activity during successful retrieval decreased over time for novel related PAs and increased for control PAs (lower panel). From Sommer (2017).
Conclusion and Future Directions
Thanks to the methodological advancements of the last years, our knowledge of how our brain forms and retains memories has advanced substantially. With the possibility to detect engrams, we have learned that they are widely distributed throughout the brain and inherently dynamic. Different sets of engram ensembles can switch between silent and active states, supporting memory at different time points and under different circumstances. With this knowledge, our view on how and when the neocortex can support declarative memories has also changed and calls for an update of existing models. We now know that rehearsal through repeated learning or endogenous reactivation supports the rapid establishment of neocortical memory engrams. What remains open is under which circumstances they also allow neocortical memory formation independent of the hippocampus. However, there is no doubt that in humans the search for the engram and the associated detection of neuronal plasticity is still in its infancy. In the future it will be important to further develop the existing detection methods, increasing spatial and temporal resolution, and extending scanning modalities to include different aspects of neuronal and astrocytic plasticity. With the currently available methods, it is already possible to detect a human memory engram if an experimental paradigm induces changes in functional as well as structural markers of plasticity, which fulfill the defining criteria of an engram. However, it is important to remember that to fully understand how memories are stored in the brain, all levels of study from individual synapses to macrostructural activity patterns have to be considered, thus pointing to the necessity of integrating findings in animals and humans.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Deutsche Forschungsgemeinschaft (DFG GA730/6-1).