
Abstract
The diagnosis, prognosis, and treatment of mild traumatic brain injuries (mTBIs), such as concussions, are significant unmet medical issues. The kinetic forces that occur in mTBI adversely affect the cerebral vasculature, making cerebrovascular injury (CVI) a pathophysiological hallmark of mTBI. Given the importance of a healthy cerebrovascular system in overall brain function, CVI is likely to contribute to neurological dysfunction after mTBI. As such, CVI and related pathomechanisms may provide objective biomarkers and therapeutic targets to improve the clinical management and outcomes of mTBI. Despite this potential, until recently, few studies have focused on the cerebral vasculature in this context. This article will begin by providing a brief overview of the cerebrovascular system followed by a review of the literature regarding how mTBI can affect the integrity and function of the cerebrovascular system, and how this may ultimately contribute to neurological dysfunction and neurodegenerative conditions. We then discuss promising avenues of research related to mTBI biomarkers and interventions that target CVI, and conclude that a clinical approach that takes CVI into account could result in substantial improvements in the care and outcomes of patients with mTBI.
Keywords
Introduction
Mild traumatic brain injury (mTBI) accounts for up to 90% of all TBI cases, with annual incidence estimated to be approximately 300/100,000 (Kay and Teasdale 2001; Nguyen and others 2016; Peeters and others 2015; Skandsen and others 2019). mTBIs are a result of mechanical linear and/or rotational forces to the brain that are common in falls, motor vehicle accidents, assaults, sport collisions, and war zone injuries (Armistead-Jehle and others 2017; James and others 2019; Theadom and others 2014). The majority of mTBI patients experience short-lived symptoms (Prince and Bruhns 2017), although there is growing evidence that >30% of patients with mTBI suffer persisting post-concussion symptoms (PPCS) (McMahon and others 2014). Studies have also linked mTBIs to a range of other long-term neurological abnormalities, including early cognitive decline and a number of neurodegenerative conditions (e.g., dementia, chronic traumatic encephalopathy [CTE], and Parkinson’s disease) (Blennow and others 2016; Gardner and others 2018; Gardner and Yaffe 2015).
Despite increasing awareness of the socioeconomic costs and poor long-term outcomes following mTBI, the clinical management of these injuries remains insufficient (Costello and others 2018; Te Ao and others 2014). mTBI diagnosis is based on evaluating the symptomatic status of patients using subjective and/or self-reported methods, and many mTBI cases go undiagnosed (Costello and others 2018). Recovery is determined to have occurred after symptoms have resolved, at which point the individual is medically cleared to return to preinjury activities (McCrory and others 2017). However, growing evidence indicates that a lack of symptoms does not equate to biological recovery (Maugans and others 2012; Talavage and others 2014; Wright and others 2016). Furthermore, although a substantial number of mTBI patients suffer from persisting neurological problems, there are currently no prognostic biomarkers that allow for the early identification of individuals at elevated risk for these issues (Symons and others 2020; Zetterberg and Blennow 2016). Even if reliable biomarkers are developed, treatment options are sparse and are not evidence-based, with the standard treatment for mTBI still being, rest until asymptomatic. Overall, the lack of mTBI biomarkers and treatments represent serious unmet knowledge gaps that prevent the personalized clinical care of mTBI that is necessary to achieve the best outcomes for patients.
To develop biomarkers and treatments for mTBI, it is imperative to understand the pathomechanisms (Shultz and others 2017). mTBI involves both primary and secondary injury processes (Kaur and Sharma 2018). Primary injury is the result of kinetic energy transferred to the brain at the moment of impact that causes structural damage to the various components of the brain tissue (e.g., axons, glia, and neurons). As such, initial mTBI biomarker studies focused largely on measures of neuronal, axonal, and glial injury with mixed results (Gan and others 2019; Kim and others 2018; Martinez and Stabenfeldt 2019). Secondary injury pathways are responses that are intended to limit the extent of the initial insult and restore homeostasis, but ultimately result in additional progressive brain damage. There are a number of known pathobiological processes that are active during the secondary injury phase, including inflammation, oxidative stress, mitochondrial and metabolic dysfunction, as well as proteinopathies (Kulbe and Hall 2017; O’Brien and others 2020; Shultz and others 2013; Tan and others 2016; Webster and others 2015; Wright and others 2016; Zetterberg and Blennow 2016). However, it should be noted that there is substantial heterogeneity in these mechanisms depending on different factors (e.g., injury mechanism, TBI severity, age, sex, comorbidities, and genetics).
Amongst the different structures/tissues that are affected by mTBI, the cerebral vasculature is especially prone to injury (Sandsmark and others 2019a). Cerebrovascular dysfunction has also been implicated in functional disturbances and neurodegenerative conditions linked to mTBI (Akoudad and others 2016; Korte and others 2020). Despite this, few studies have focused on the cerebrovascular system in the context of mTBI biomarkers and treatments. This article will begin with an overview of the cerebrovascular system, before reviewing the literature pertaining to how mTBI might affect cerebrovascular integrity and function, and how this can contribute to neurological abnormalities and neurodegeneration. We then discuss promising avenues of research aimed at developing biomarkers and interventions related to cerebrovascular injury (CVI) and conclude that these studies could one day improve the care and outcomes of patients with mTBI.
The Cerebrovascular System
The brain is an energy-expensive organ, and therefore requires sufficient perfusion to facilitate delivery of key metabolites such as glucose (Herculano-Houzel 2011). The blood-brain barrier (BBB) is a multicellular structure that is part of the neurovascular unit (NVU) and helps regulate the movement of molecules and cells between the brain parenchyma and systemic circulation (Liu and others 2012; Mastorakos and McGavern 2019). The NVU exists at all levels of the vascular network within the CNS and is composed of several components; endothelial cells (ECs), pericytes, basement membrane, astrocytes, neurons, and immune cells (Mastorakos and McGavern 2019). The function of the BBB largely results from the unique properties of ECs within the cerebrovascular system (Obermeier and others 2013). BBB ECs are surrounded by basement membrane, astrocytic foot processes, and pericytes, and have continuous intercellular tight junctions, lack fenestrations, and undergo low rates of transcytosis (Obermeier and others 2013). These properties help control paracellular and transcellular movement of molecules through the EC layer, and key molecules such as water and glucose must instead cross the BBB via specific transporters (Chandra and others 2017; Obermeier and others 2013). Additionally, ECs have low expression of leukocyte adhesion molecules and therefore immune cell infiltration into the healthy CNS is limited (Daneman and Prat 2015). However, the occurrence of an mTBI can adversely affect the function of the NVU, either via direct mechanical forces or secondary injury mechanisms, making an impaired NVU an important component of the pathogenic process of mTBI (Sandsmark and others 2019a).
CVI Following mTBI
Cerebral Blood Flow Changes
The neurovascular network maintains the cerebral perfusion that enables delivery of oxygen and essential micronutrition to the brain parenchyma (Sandsmark and others 2019a). Dysregulation of cerebral blood flow (CBF) after mTBI can be transient or lasting and is believed to be an important factor contributing to abnormal cerebral perfusion and symptom severity, including PPCS (Allen and Epps 2017).
In clinical studies, symptomatic mTBI patients have been found to have a reduction in CBF in the absence of detectable structural brain injury (Bonne and others 2003). CBF change after mTBI is likely to be attributed to disruption in cerebrovascular reactivity and dysregulation in neurovascular coupling (Allen and Epps 2017). A Doppler ultrasonography study showed that cerebrovascular reactivity was not impaired at rest in patients following an mTBI but was impaired in response to respiratory stress (Len and others 2011; Len and others 2013). In concussed athletes, Doppler ultrasound revealed acute changes in neurovascular coupling response dynamics that persisted beyond 2 weeks but retuned to baseline by 1 month (Wright and others 2017). Using advanced arterial spin labeling (ASL) magnetic resonance imaging (MRI), Wang and colleagues found that adolescent footballers have significantly lower CBF in a variety of brain regions compared to controls, at 24 hours and 8 days after concussion (Wang and others 2016). Interestingly, while CBF in controls remained unchanged between the two timepoints, the concussion group had decreased CBF in the frontal and temporal lobes at 8 days compared with 24 hours (Wang and others 2016). Similarly, in another study of sports-related concussion in young adults, ASL found decreased regional CBF at 1-day and 1-week post-concussion. In this study, the CBF abnormalities were typically resolved at 1 month; however, concussed athletes with worse outcomes had lower CBF in the dorsal midinsular cortex compared to those with better outcomes (Meier and others 2015). Lower regional CBF was also reported as long as 3 to 12 months after concussion in adolescents, though it was unclear whether previous concussions in some subjects could have influenced this outcome (Wang and others 2015). Although decreased CBF is most commonly reported after mTBI, it is important to note that there have been some contradictory findings. An ASL study evaluated CBF changes in adolescent athletes 2 and 6 weeks after sport-related concussion and found higher CBF at 6 weeks to be associated with symptoms (Stephens and others 2018). Another ASL study found decreased regional CBF during the second week after mTBI; however, worse symptoms were correlated with higher CBF in these hypoperfused areas (Lin and others 2016).
CBF abnormalities have also been reported in rodent mTBI studies. For example, Ichkova and colleagues reported early cerebrovascular hypo-oxygenation in the impacted brain area hours after mTBI in juvenile mice, which was restored at 3 days (Ichkova and others 2020). Furthermore, these were accompanied by modified cortical cerebrovascular reactivity (Ichkova and others 2020). A laser Doppler imaging study of mice given repetitive mTBI found reduced CBF in the occipital cortex (Ojo and others 2016). In another mouse study of mTBI, subtle regional changes of CBF were observed at 3 days, including hypoperfusion in the hippocampus and cortex proximal to the injury site, and hyperperfusion in the surrounding cortical area (Lyons and others 2018). The temporal profile of CBF changes, as well as other pathomechanisms, biomarkers, and treatments discussed below that are relevant to mTBI, is shown in Figure 1.
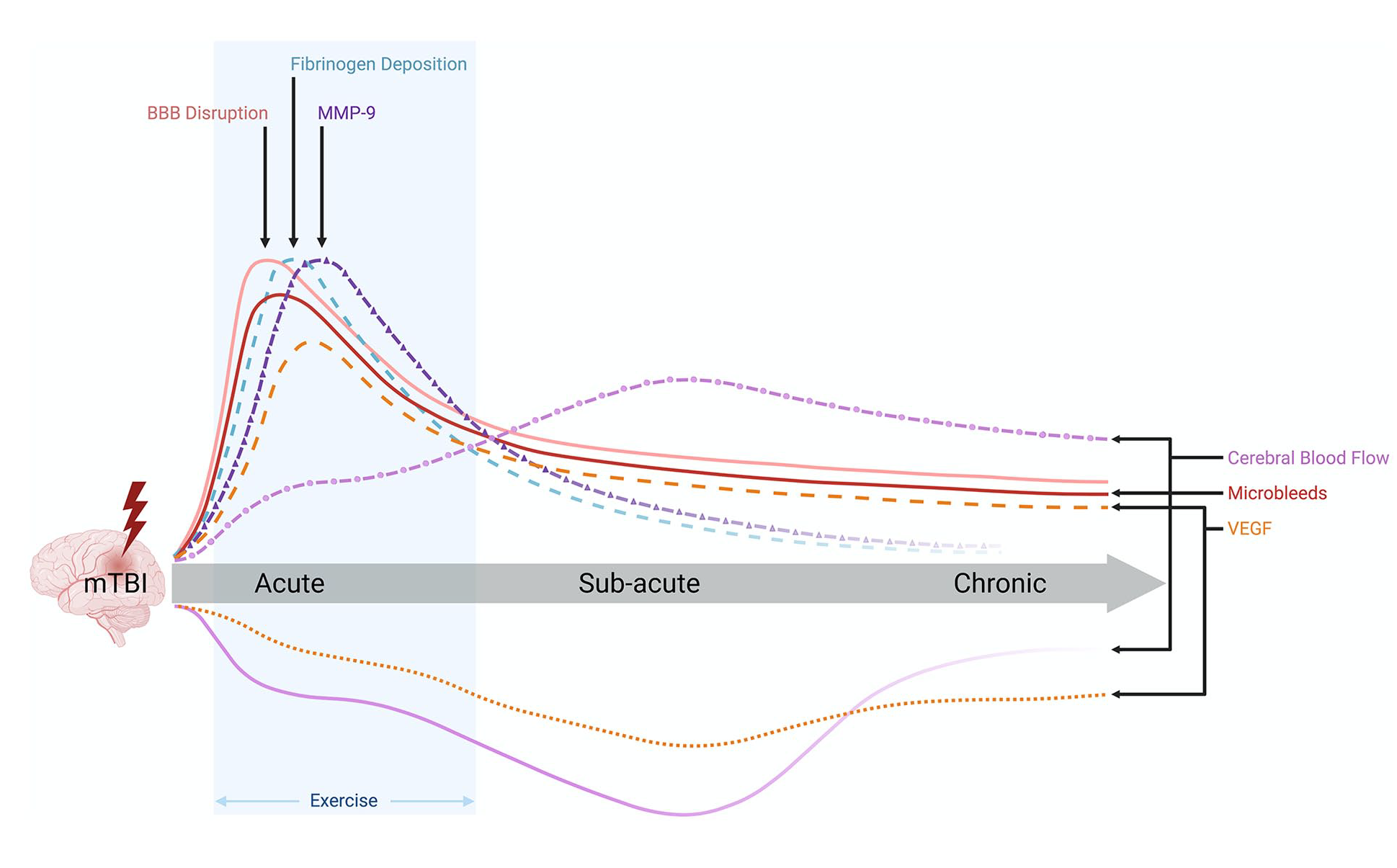
Temporal profile of pathophysiology, biomarkers, and interventions related to cerebrovascular injury (CVI) after mild traumatic brain injury (mTBI). The biomechanical forces at the time of mTBI result in an acute increase in microbleeds, blood-brain barrier (BBB) dysfunction (e.g., damage to endothelial cells, destabilization of tight junctions, and elevated levels of circulating occludin and claudins), fibrinogen deposition, and matrix metalloproteinase–9 (MMP-9). Some studies suggest that these may persist into the subacute and chronic stages, either due to a lack of recovery or secondary injury mechanisms (e.g., oxidative stress, inflammation). Most studies indicate that cerebral blood flow (CBF) is decreased after mTBI, although others have indicated an increase in CBF after mTBI. The majority of studies investigating vascular endothelial growth factor (VEGF) indicate that it is elevated in response to mTBI; however, some studies have found an acute decrease that persists chronically. There is growing evidence that subsymptom threshold aerobic exercise early after mTBI can facilitate recovery. Solid line = strong evidence; dashed line = moderate evidence; dotted line = preliminary evidence.
Microhemorrhage/Microbleeds and Other BBB Abnormalities
As shown in Figures 1 and 2, kinetic energy transferred by the mTBI impact to brain tissue can result in microhemorrhages/microbleeds, as well as other damage to the NVU and BBB. Extravasation of blood and other molecules into the brain parenchyma then occurs, which can be cytotoxic and initiate secondary injury mechanisms that further compromise the integrity of the cerebrovascular system (Logsdon and others 2015). Even in the absence of microbleeds, the primary mechanical insult can induce other BBB abnormalities and initiate secondary injury processes that contribute to BBB dysfunction as described below (Fig. 2).
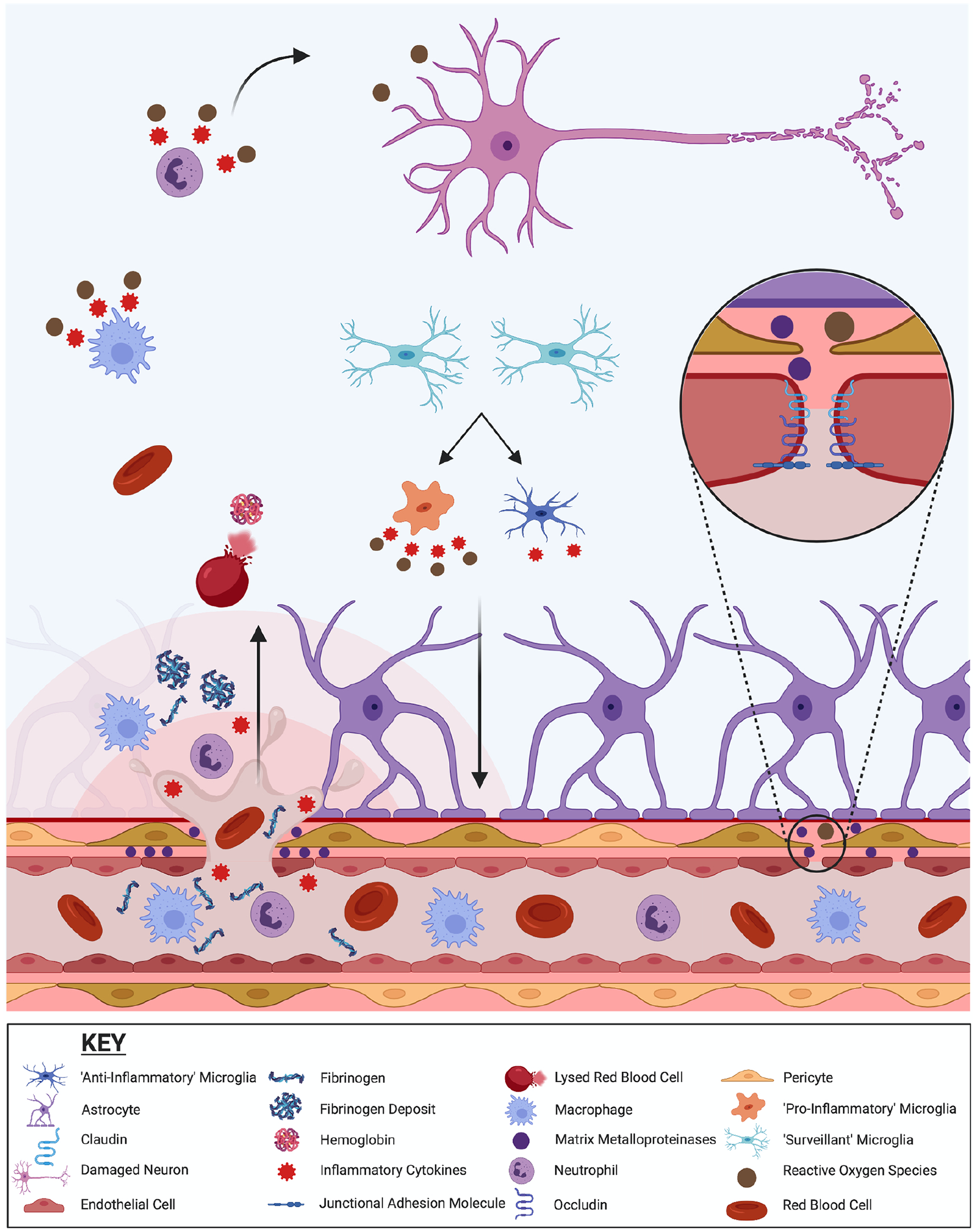
Potential pathomechanisms related to cerebrovascular injury after mild traumatic brain injury (mTBI). Kinetic energy due to the mTBI impact can result in microhemorrhages/microbleeds, as well as other damage to the neurovascular unit (NVU) and blood-brain barrier (BBB). Red blood cells and peripheral immune cells, such as macrophages and neutrophils, can infiltrate the brain parenchyma. Red blood cells may lyse, releasing hemoglobin and iron that increase oxidative stress and the consequent damage to neurons. Infiltrating peripheral immune cells also release reactive oxygen species and inflammatory cytokines that can be cytotoxic and initiate/exacerbate the activation of microglia and further neuroinflammation. Fibrinogen deposits and activation of matrix metalloproteinases (MMPs) may also occur, and further promote neuroinflammation and cerebrovascular injury. Even in the absence of microbleeds directly due to mechanical injury, mTBI can induce other abnormalities to the cerebrovascular system. The kinetic forces can dissociate endothelial tight junction proteins (e.g., occludin, claudin-5) resulting in the opening of the BBB. Oxidative stress, MMPs, and neuroinflammation in and around brain capillaries can also contribute to BBB dysfunction and damage.
Clinical studies have primarily relied on MRI methods to investigate the presence of microbleeds after mTBI. Griffin and colleagues acquired MRI images of 365 mTBI patients within 48 hours of injury and found that 97 had microbleeds as identified by T2* gradient recalled echo images and confirmed by susceptibility weighted imaging (SWI) (Griffin and others 2019). Notably, these microbleeds were also predictive of worse outcome (Griffin and others 2019). In another SWI study, Wang and colleagues found that 32 of 165 mTBI patients had evidence of microbleeds within 3 days of the injury, and that microbleeds in the frontal, parietal, and temporal lobes were likely predictive of post-traumatic depression (Wang and others 2014). There is also clinical evidence that microbleeds can persist long after mTBI. For example, Huang and colleagues reported microbleeds detected via SWI in mTBI patients at 8 to 55 days post-injury that were positively correlated with short-term memory deficits (Huang and others 2015). Another recent study of a small mTBI cohort found that 28% of the mTBI patients had microbleeds within 48 hours of the injury that were detectable for as long as 5 years post-injury (Rizk and others 2020). Using dynamic contrast enhanced (DCE)-MRI in high-risk mTBI populations, such as professional fighters and rugby players, disruption of BBB post-fight/-season/-match was found, dependent on the level of exposure (O’Keeffe and others 2020). Initial preclinical studies are largely consistent with clinical findings in demonstrating that microbleeds and BBB disruption are common in mTBI. A study in mice found that cerebral microbleeds were detectable at 48 hours after a single mTBI (Bouley and others 2019). In a mouse model of repetitive mTBI, mice had microbleeds in the cortex, corpus callosum, hippocampus, and brain stem 3 days after the final injury (Chen and others 2019). In another study of mouse repetitive mTBI, focal cortical microhemorrhages were found at 1 week post-injury (Robinson and others 2017).
In addition to microbleeds, mTBI can disrupt other aspects of BBB integrity. In a rat model of focal mTBI, increased BBB permeability was found in the superficial cortical layers as early as 1 hour post-injury (Li and others 2016). This continued to evolve and peaked at 3 days, then decreased toward normal at 7 days (Li and others 2016). In a mouse study, mTBI resulted in acute BBB dysfunction, as well as dysmorphic capillaries, abnormal endothelial cells, and abnormal perivascular astrocyte end-feet at 2 weeks post-injury (Tagge and others 2018). Using a swine model of concussion, Johnson and colleagues found multifocal BBB disruption 6 to 72 hours post-injury, as evidenced by extravasation of blood-borne serum products (e.g., fibrinogen), in the absence of hemorrhage or other focal pathology (Johnson and others 2018). Fibrinogen is a potent inflammatory promoter, and thus could promote CVI via neuroinflammation and the recruitment of circulating immune cells into the brain parenchyma (Muradashvili and Lominadze 2013). Postmortem analysis of more severe TBI in human brain tissue has demonstrated parenchymal fibrinogen deposition, especially during the acute stages of injury, which was associated with microglial/macrophage activation (Jenkins and others 2018). Whether this also occurs in mTBI remains to be determined. There is also evidence that fibrinogen can compromise vascular integrity by altering endothelial cell tight junction proteins (Patibandla and others 2009), and through the activation of MMPs (Muradashvili and others 2012). MMPs are extracellular enzymes that have been relatively understudied in the context of mTBI but have been implicated in BBB breakdown after more severe TBI and related conditions (Guilfoyle and others 2015). In one of the few mTBI studies, it was found that MMP-9 knock-out mice had less fibrinogen deposition and cerebrovascular permeability (Muradashvili and others 2015). In patients with contusion TBI, brain microdialysate analysis revealed that MMP-9 concentrations were acutely elevated (Guilfoyle and others 2015). TBI in mice has been found to result in activation of MMP-9 within 24 hours, and this remained elevated for 7 days (Hadass and others 2013). However, it should be noted that MMP-9 has also been found to promote angiogenesis and neovascularization, for example, during the delayed but not the acute phase of stroke (Cai and others 2017; Zhao and others 2006). Animal model studies have also found that mTBI can disrupt the BBB via oxidative stress mediated vascular inflammation (Abdul-Muneer and others 2013; Mishra and others 2016; Shetty and others 2014). Taken together, BBB disruption can be induced by both primary and secondary injury mechanisms involved in mTBI.
The Role of CVI in Neurological Dysfunction and Neurodegeneration after mTBI
As alluded to above, there is initial clinical evidence suggesting that cerebrovascular abnormalities may contribute to the symptomology that occurs following mTBI, including PPCS, depression, and cognitive deficits (Bonne and others 2003; Huang and others 2015; Lin and others 2016; Meier and others 2015; Stephens and others 2018; Wang and others 2014). Animal model studies provide additional evidence linking cerebrovascular disturbances to behavioral dysfunction after mTBI. For example, in a rat model of mTBI, hemodynamic disturbances in CBF and cerebral vascular reactivity were observed in the first 14 days post-injury and occurred in the presence of sensorimotor deficits (Long and others 2015). Another rat study found that males had worse cerebrovascular and neuromotor function post-injury than their female counterparts (Wright and others 2020). In a mouse model of repeated mTBI, CBF was significantly impaired at 7 months post-injury and associated with impaired acquisition and consolidation of spatial memory in the Barnes maze (Lynch and others 2016).
Repeated mTBI is now recognized as a risk factor for consequent neurodegenerative disease, including CTE and different forms of dementia (Blennow and others 2016; Gardner and Yaffe 2015). There is also growing evidence supporting an important role for CVI in the pathophysiology of neurodegeneration (Sandsmark and others 2019a). As discussed earlier (Fig. 2), mTBI can result in a range of cerebrovascular abnormalities and related pathomechanisms, including microbleeds, BBB disruption, neuroinflammation, oxidative stress, changes in CBF, increased levels of fibrinogen, and activation of MMPs. Importantly, each of these factors have been implicated in neurodegeneration. For example, the presence of brain microbleeds has been associated with an increased risk for dementia in the general population (Akoudad and others 2016). An essential pathological feature of CTE, a neurodegenerative condition postulated to be induced by repetitive mTBIs, is the accumulation of perivascular phosphorylated tau (McKee and others 2016). In a CTE study involving both human and experimental mTBI, BBB breakdown and perivascular neuroinflammation were evident, with the presence of tauopathy (Tagge and others 2018). In a CTE case study, BBB disruption was found in areas with dense perivascular phosphorylated tau (Doherty and others 2016). Fibrinogen (Jenkins and others 2018), MMPs (Hadass and others 2013), neuroinflammation (Sun and others 2017; Webster and others 2015), and oxidative stress (Webster and others 2015) have all been found to contribute to neurodegeneration after TBI (Sulimai and Lominadze 2020). There is also evidence suggesting that decreased CBF after mTBI can be long-lasting and may contribute to the neurodegenerative process (Meier and others 2015; Sullivan 2019). Of potential relevance to the cumulative effects of repeated mTBIs, it has been postulated that inadequate blood supply post-mTBI may leave the brain in a state of increased cerebral vulnerability (ICV) (Giza and Hovda 2014; Prins and others 2013). Sustaining a successive mTBI during a period of ICV has been postulated to contribute to worse outcomes and neurodegeneration (Povlishock 2013; Prins and others 2013). Although further research is ultimately required to investigate the relationship between mTBI, cerebrovascular abnormalities, and consequent neurodegeneration, the available evidence indicates that such endeavors are warranted.
Blood and Neuroimaging Biomarkers of CVI
Blood Biomarkers
Current clinical management of mTBI (e.g., diagnosis, recovery/return to pre-injury activities) is guided by the presence or absence of symptoms (Costello and others 2018; McCrory and others 2017), but there is increasing evidence that symptom-based approaches are neither specific nor reliable indicators of mTBI (Iverson and Lange 2003; McDonald and others 2021; Wang and others 2006). Importantly, they do not accurately indicate when the brain has physiologically recovered (i.e., clinical recovery precedes physiological recovery) (Maugans and others 2012; Talavage and others 2014; Wright and others 2016). Conversely, blood-based molecular biomarkers have the potential to provide reliable diagnostic, prognostic, and predictive information in mTBI (Wright and others 2016). Much research to date has focused on proteins that may be released into circulation from damaged neurons, axons, and glia after mTBI. Although some markers associated with neuronal, axonal, and glial injury have shown promise as diagnostic indicators of mTBI, other data indicate that mTBI may not reliably increase circulating levels of markers associated with these processes (Zetterberg and Blennow 2016). As such, it is possible that blood-borne biomarkers associated with damage or repair of the cerebral vasculature may also have utility in the context of mTBI, particularly in cases in which neuronal, axonal, or glial damage may be negligible. Moreover, given the aforementioned contribution that CVI may play in the acute and chronic neurological consequences of mTBI, there is potential that biomarkers of CVI may have utility in the context of prognostics or determining recovery.
The most commonly investigated fluid indicator of BBB damage is the ratio of albumin found in the cerebrospinal fluid (CSF) compared with the serum, with increases in this ratio reflective of abnormal leakage of albumin into the brain (Blyth and others 2009). Increased CSF:serum albumin ratio (QA) occurs following moderated-severe TBI (Blyth and others 2009), indicative of BBB disruption; however, studies to date have revealed that this ratio may not be altered in mTBI (Blennow and others 2011; Zetterberg and others 2006). It is possible BBB may remain intact in some cases of mTBI, or that opening may be transient and therefore providing a narrow window for QA utility, or that this measure may lack the sensitivity required to detect subtle changes in mTBI. Moreover, the requirement for CSF collection limits the clinical appeal of QA assessment in the context of mTBI. As such, alternative indicators of BBB damage may have more promise, particularly if able to be measured in readily accessible fluids such as serum/plasma or saliva.
Peripheral blood concentrations of astrocytic protein S100 calcium-binding protein B (S100B) have been widely investigated in the context of TBI and may be reflective of BBB disruption (Thelin and others 2017). A calcium-binding protein predominately expressed by perivascular astrocytes, serum S100B levels have been shown to rise acutely after TBI and may be predictive of computed tomography (CT) abnormalities in mTBI patients (Thelin and others 2017). Several studies have interpretated raised serum S100B levels as reflective of CVI, either due to the integral role of S100B-containing astrocytes in the BBB, or as a result of the relatively high brain S100B concentrations facilitating concentration gradient-induced efflux on BBB opening. Nonetheless, although S100B appears to have utility in the context of mTBI, its use as a specific indicator of BBB damage is uncertain, with some studies in more severe forms of brain injury reporting a correlation between serum S100B levels and QA after TBI (Blyth and others 2009), and others no such correlation (Kleindienst and others 2010). Indeed, whether these findings translate to milder forms of TBI remains unknown.
Proteins involved in the stabilization of tight junctions between endothelial cells, such as occludin and the claudins, have recently shown potential as circulating indicators of CVI after mTBI. Plasma occludin levels were significantly elevated acutely after mTBI in humans when compared with uninjured controls; however, levels were also elevated in orthopedic injury controls (Shan and others 2016). Acutely elevated plasma occludin was also recently observed after mTBI in mice (Robinson and others 2020). To the best of our knowledge, no studies have assessed how blood occludin levels align with the extent of BBB damage after mTBI, although a recent study on ischemic stroke in rats revealed a strong correlation between acute serum occludin levels and BBB integrity as indicated by Evans blue extravasation (Pan and others 2017). Increased claudin-5 has been found in CSF acutely after mild-moderate blast-induced TBI in swine (Ahmed and others 2012) and also in plasma at 48 hours after mild blast-induced TBI in rats (Kamnaksh and others 2014). The findings indicate that occludin and claudin-5 may have significant potential as fluid biomarkers of mTBI-induced CVI.
The pathophysiological response to CVI and endothelial stress involves a cascade of molecular changes that may have utility as biomarkers of mTBI. For example, angiogenesis is a critical component of repair after CVI, and is reliant on increased expression of molecules such as vascular endothelial growth factor (VEGF) (Salehi and others 2017). Clinical and pre-clinical studies have investigated peripheral VEGF levels in both the acute and chronic stages after mTBI, with findings to date largely mixed. In the acute stages after mTBI, plasma VEGF levels were significantly elevated in individuals that were deemed to be either computed tomography (CT) positive or MRI positive (CT negative) when compared with controls that were both CT and MRI negative (Edwards and others 2020). Chronic investigations have revealed no differences in both plasma VEGF and plasma exosome-derived VEGF concentrations between military personnel with and without a history of mTBI (Guedes and others 2020). However, a recent investigation of Australian football players (i.e., Australia’s most participated collision sport) found that serum VEGF levels were increased in male players when compared to control athletes (Major and others 2020). Increased VEGF levels in CSF and plasma have been reported in animal models of blast-induced mTBI (Ahmed and others 2012; Ahmed and others 2015; Kamnaksh and others 2014), whereas a prolonged decrease in plasma VEGF was found in rats after single and repeated mild TBI induced by fluid percussion injury (Wright and others 2016; Wright and others 2019), potentially indicating the that mechanisms of injury may be an important factor. Of interest, the abovementioned preclinical blast mTBI study also found elevated circulating levels of von Willebrand factor (vWF) (Ahmed and others 2013), which is a protein that is upregulated and secreted by endothelial cells in response to stress/damage and has an important role in thrombus formation (Lenting and others 2012). Clinically, one study found elevated plasma levels of vWF in TBI patients with intracranial hemorrhage (Sandsmark and others 2019b), and others have found that elevated vWF levels are associated with worse outcomes after severe TBI (De Oliveira and others 2007; Yokota and others 2002). While these initial findings pertaining to VEGF and vWF are promising, further research is required to characterize the changes in these potential biomarkers in the acute and chronic stages after mTBI.
Circulating extracellular vesicles (EVs), such as exosomes, have shown much promise as biomarkers of injury and disease due to their protein and RNA content reflecting that of the cell of origin. The concept of isolating “brain-specific” EVs is particularly enticing; however, isolating EVs originating exclusively from cells specific to the cerebral vasculature is likely to be challenging (Ramirez and others 2018). Finally, recent studies have indicated that mTBI can result in alterations in circulating microRNAs (Polito and others 2020; Qin and others 2018); future research is required to determine if microRNAs associated with CVI have potential as blood biomarkers of mTBI.
Neuroimaging Biomarkers
MRI techniques such as functional MRI (fMRI), ASL, SWI, quantitative susceptibility mapping (QSM), and contrast-enhanced imaging have all been previously employed to investigate changes in the brain’s functional connectivity, CBF, microhemorrhages, and structural integrity of the BBB following mTBI (see Fig. 3).
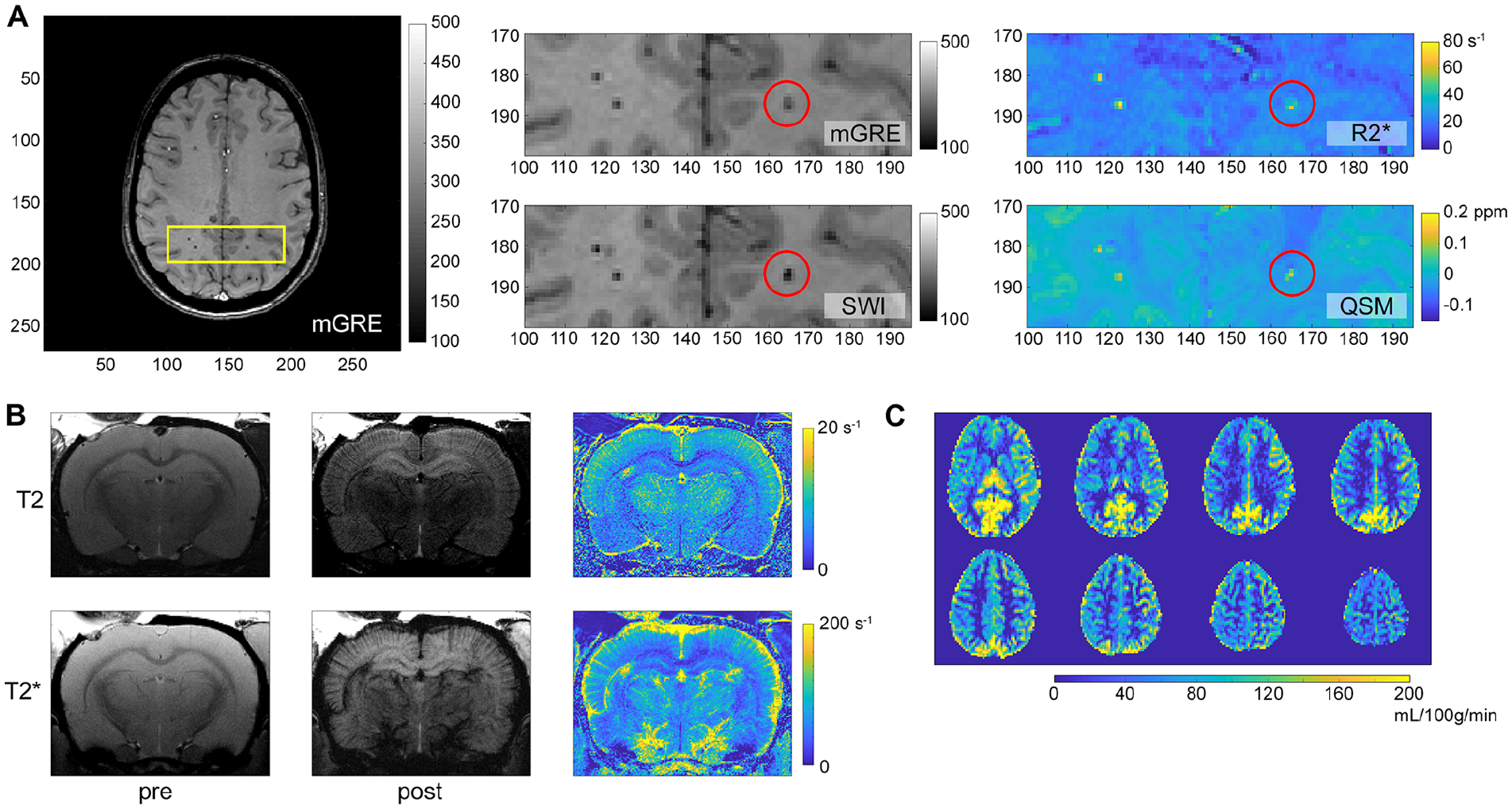
Magnetic resonance imaging (MRI) can reveal insights related to cerebrovascular injury (CVI) following mild traumatic brain injury (mTBI). (A) Multiple gradient echo (mGRE) weighted image permits the reconstruction of multiple image contrasts. Susceptibility weighted imaging (SWI) exploits phase information in the received signal to increase image contrast, particularly in microhemorrhages (e.g., red circle). These microhemorrhages appear hyperintense on the R2* map and on the quantitative susceptibility mapping (QSM) image, reflecting the increased iron (paramagnetic) in deoxygenated blood. Images were acquired from a participant within 48 hours of mTBI. (B) T2-weighted and T2*-weighted images acquired pre and post injection of an iron-based contrast agent, in this case ferumoxytol, can be used to interrogate cerebral blood volume in smaller (top row) and across a broader range (bottom row) of blood vessels. (C) Arterial spin labeling (ASL) can also be used to quantify cerebral blood flow (CBF), as shown here in a subject less than 48 hours after mTBI.
In functional MRI (fMRI), blood oxygenation level dependent (BOLD) activity is used as a surrogate for neuronal activity (Ogawa and others 1990) given the relationship between neural activation, local increases in blood flow (due to neurovascular coupling), and consequent decreases in deoxyhemoglobin concentration. Resting state fMRI (rs-fMRI) can be used to examine spatially separate brain regions that are functionally related within the whole brain. Observable changes in brain network connectivity and CBF associated with a history of mTBI have been described in several studies (Churchill and others 2017; Vergara and others 2017; Vergara and others 2018). Consequently, rs-fMRI may be a valuable measure to identify mTBI patients. However, findings across the rs-fMRI literature are highly varied. Altered functional connectivity has been observed across visual, motor, and executive functioning networks with no consistent direction of change (Borich and others 2015; Czerniak and others 2015; Meier and others 2017; Slobounov and others 2011). The default mode network has been extensively explored following mTBI, with both increases and decreases in connectivity reported (Borich and others 2015; Johnson and others 2012; Militana and others 2016; Zhu and others 2015). Similarly, fMRI studies exploring task-related activation within a network such as dorsolateral prefrontal cortex have reported both decreased (Chen and others 2004; Chen and others 2007; Chen and others 2008; Keightley and others 2014) and increased (Dettwiler and others 2014; Zhang and others 2010) activation following mTBI. Moreover, altered activity outside of the specific task-related regions has also been found, with some suggesting this may be compensatory in nature (Chen and others 2004; Chen and others 2007; Chen and others 2008; Johnson and others 2015; Slobounov and others 2010). The heterogeneity across injury mechanism, recovery time, and severity may be modifying factors contributing to the variability in fMRI findings, as well as the methodological difference across studies (Kamins and others 2017; McCrea and others 2017).
ASL is a quantitative method of assessing CBF using MRI (Petcharunpaisan and others 2010). The ASL sequence utilizes unique radiofrequency pulses that excite/tag water protons in the blood as they pass through the neck arteries. The magnetization is trailed for a period of time whilst the blood travels to the brain, wherein the label is transferred to the brain parenchyma via capillary exchange (Andre 2015). ASL provides measures of relative CBF (rCBF), which outputs a value for a given region of interest (ROI) relative to the perfusion within the rest of the brain. Alternatively, absolute CBF (aCBF), which quantifies the perfusion of a given ROI, can be calculated independently of other brain regions. Over a dozen studies have now employed ASL in the context of mTBI. These studies have reported inconsistent patterns which may be somewhat influenced by the timing of analyses relative to injury. Acutely after mTBI, there appears to be a reduction in CBF (Lin and others 2016; Meier and others 2015; Peng and others 2016; Wang and others 2015). In the subacute stages, findings become contradictory, with some reporting a decrease in CBF (Lin and others 2016; Wang and others 2015), and others reporting an increase (Stephens and others 2018). Taken together, ASL is a valuable marker of alterations in CBF following mTBI; however, additional studies involving larger sample sizes and longitudinal scans across acute, subacute, and chronic recovery times are required.
SWI is a technique that exploits the magnetic susceptibility differences of different compounds (e.g., iron) that are present in microhemorrhages/microbleeds (Halefoglu and Yousem 2018; Huang and others 2015; Kenney and others 2016). Consequently, several mTBI studies have employed SWI to explore the presence of microhemorrhages with varied results. In the three most extensive SWI studies to date, increased numbers of microhemorrhages have been reported following mTBI when compared with non-injured controls (Huang and others 2015; Trifan and others 2017; Wang and others 2014). For example, Huang and colleagues reported microbleeds in 23.4% mTBI patients at 8-55 days post-injury that appeared to region specific (i.e., predominately in the cortex/subcortical region) in comparison to the fewer microbleeds detected in controls (Huang and others 2015). However, smaller mTBI studies applying SWI have failed to find an increase in microhemorrhages after mTBI. Illustrating this, a study found no differences in the number of microhemorrhages in concussed ice hockey players compared to healthy controls (Jarrett and others 2016). Furthermore, mTBI studies in pediatric populations have also reported normal SWI results post-mTBI (Maugans and others 2012; Sigmund and others 2007; Yang and others 2012). QSM has also been used to identify cerebrovascular abnormalities in mTBI (Liu and others 2019). In a study of military patients with mTBI, venous volume above the lateral ventricle was assessed using QSM images, and increased segmented volumes were found compared with the controls, suggesting potential venous stasis, damaged veins, extravasated blood, and microthrombosis (Liu and others 2019). In addition, QSM has recently been used to quantify the levels of cerebral venous oxygen saturation (SvO2), which may potentially serve as a marker of CVI after mTBI. Chai and colleagues found that QSM identified increased regional cerebral SvO2 in patients with mTBI 12 to 360 hours post-injury (Chai and others 2017).
Past studies have also utilized both steady state contrast-enhanced (SSCE) and DCE-MRI to provide insight into neurovascular integrity (Sweeney and others 2019). Wright and colleagues applied SSCE MRI to a rat model of TBI and found detectable decreased vascular density in the injured cortex and hippocampus at 48 hours after the TBI (Wright and others 2020). DCE-MRI studies typically evaluate rapid volume transfer (Ktrans) of contrast agent delivered to the extracellular space, with accumulation reflecting damage of normal tight junctions between neurovascular endothelial cells. Yoen and colleagues computed Ktrans values for 42 mTBI patients and compared them with 29 healthy controls, reporting significantly higher mean Ktrans values in the bilateral cerebral cortex as well as lower fractional plasma volume in cerebellar white matter in the mTBI patients (Yoen and others 2021). Recently, a modified DCE-MRI approach has been developed to detect subtle BBB dysfunction (O’Keeffe and others 2020; Veksler and others 2020; Weissberg and others 2014). Using this modified DCE-MRI protocol, amateur American football players with repeated mTBI were found to have abnormal normalized permeability index (NPI) values (Veksler and others 2020). Specifically, the NPI identified several affected brain regions, including long white matter tracts, brainstem structures and left temporal cortex. This study provides evidence of focal brain microvascular pathology persisting for up to 10 months post-injury. Additionally, a subset of football players was scanned longitudinally (on average 8 months after the initial scan). Over 50% of this subset had increased NPI in the second scan, suggesting progressive BBB disruptions in a subgroup of individuals (Veksler and others 2020). Other DCE-MRI studies in amateur rugby (O’Keeffe and others 2020) and American football (Weissberg and others 2014) players have also reported BBB abnormalities in these athletes.
Approaches to Enhance Cerebrovascular Recovery after mTBI
Endogenous Cerebrovascular Repair Mechanisms
Before discussing interventions to promote cerebrovascular recovery, it is important to acknowledge the endogenous repair mechanisms that occur. Although the time course and underlying mechanisms of endogenous CVI repair after mTBI are still being elucidated, initial studies have provided some insight. For example, a recent study investigated meningeal vascular injury and recovery after mTBI in both humans and mice (Russo and others 2018). In humans with mTBI, the meningeal vascular injury resolved in the majority of patients within 2 to 3 weeks, although the injury persisted for months in some patients (Russo and others 2018). To investigate potential mechanisms the authors next employed a mouse model of mTBI. Angiogenesis in the injured meningeal vasculature was found to initiate within days, and this was mediated by infiltrating myeloid cells. The authors also found that a subsequent mTBI at 1 day (i.e., the acute inflammatory phase) after the initial mTBI restricted this recovery process, whereas a subsequent mTBI at 4 days after the initial mTBI (i.e., the wound-healing phase) did not (Russo and others 2018).
A study in rats found that an initial decrease in cerebral microvascular density after TBI normalized by 2 weeks post-injury (Park and others 2009). The authors suggested that this repair was associated with increased VEGF-A expression in the injured brain, which was regulated by a hypoxia-mediated signaling pathway (Park and others 2009). This is consistent with the general belief that cerebrovascular damage and the resultant hypoxic environment can prompt vascular repair including angiogenesis via hypoxia-inducible factor and prolyl-hydroxylases (Salehi and others 2017). Of relevance to this pathway, mTBI patients with CT abnormalities had elevated circulating VEGF-A levels within 24 hours of the injury (Edwards and others 2020). Furthermore, a study in male athletes with a history of mTBI had increased circulating levels of VEGF-A compared to controls (Major and others 2020).
Aerobic Exercise to Promote Cerebrovascular Recovery
Regular physical exercise has been shown to promote neurogenesis, angiogenesis, and synaptic plasticity (Farmer and others 2004; Isaacs and others 1992), reduce neuroinflammation (Liu and others 2020; Sung and others 2012), and alter CBF (Alfini and others 2019). Within the context of mTBI, a pre-clinical study found that initiation of exercise within 1-3 days of mTBI, which is known to upregulate VEGF and other growth factors (e.g., brain-derived neurotrophic factor (Sleiman and others 2016)) in the brain (Morland and others 2017), improved cognitive and motor outcomes in rats (Mychasiuk and others 2016). With regards to human studies, a recent clinical trial found that an early subsymptom threshold exercise intervention reduced recovery times and the number of symptoms in concussed adolescents (Leddy and others 2019). Another study found that exercise may promote recovery in patients with mTBI suffering from persisting symptoms for >3 weeks, however this remains inconclusive due to the low subject number within the control group (Baker and others 2012). These initial studies support exercise as a potential intervention to facilitate acute and subacute recovery after mTBI, though future large-scale studies that include other mTBI populations (e.g., adult mTBI patients recruited from emergency departments) and determine therapeutic window are required.
While the application of exercise therapy for mTBI has so far focused on the acute and subacute treatment window, there is also evidence to suggest that exercise could be protective against the chronic neurodegenerative processes linked to mTBI. Exercise promotes a number of beneficial factors involved in CBF regulation, including the regulation of brain metabolics and neuronal activity, arterial generated partial pressure of carbon dioxide, blood pressure, and cardiac output (Ogoh and Ainslie 2009). It appears that extended periods of aerobic exercise promote endothelial integrity and slow aging processes within the NVU (Latimer and others 2011). Pre-clinical studies suggest that angiogenic capacity undergoes heterogeneous changes throughout the brain during normal aging, with exercise providing therapeutic benefit at the level of the NVU, but only within certain regions (Murugesan and others 2012). For example, while aging greatly altered expression of many angiogenic-associated genes across all brain regions, physical exercise selectively increased capillary branching and density only within the hippocampus, having no effect in the cortex and corpus callosum (Murugesan and others 2012). In addition, through the stabilization of astrocytic APOE levels, exercise prevents aging related neurovascular deterioration of cells within the basement membrane, astrocytic dysfunction, and pericyte loss in aged mice (Soto and others 2015). Taken together, exercise therapy could also mitigate the chronic consequences of mTBI although further studies are required to evaluate this.
Potential Pharmaceutical Interventions
As already discussed, it is likely that VEGF, a potent promoter of angiogenesis and other important aspects of vascular health, plays an important role in neovascularization after TBI. Consequently, VEGF could be a promising therapeutic target to rescue CVI and restore blood supply to injured brain tissue (Lee and Agoston 2010; Reeson and others 2015; Tado and others 2014) (see Figs. 1, 2, and 4). While the therapeutic effects of modulating VEGF after mTBI remains unknown, intracerebroventricular administration of VEGF after experimental moderate TBI was neuroprotective (Lee and Agoston 2010). On the other hand, administration of the VEGF neutralizing antibody, bevacizumab, led to exacerbated contusion necrosis and neurological deficits after moderate TBI in rats (Tado and others 2014). This suggests a role of VEGF in promoting angiogenesis and microcirculation surrounding the contusion core (Tado and others 2014). Furthermore, a study found that both early and delayed upregulation of VEGF after moderate TBI resulted in improvements in microvascular recovery in rats (Siddiq and others 2012). Another study found that the implantation of synthetic self-assembling peptide-based hydrogel into the injured brain after a moderate blunt TBI in rats led to the activation of VEGF-receptor 2 and significant blood vessel formation. It is however notable that the exogenous administration of VEGF to promote neovascularization after TBI remains controversial, as there is conflicting evidence with regard to VEGF promoting BBB permeability (Croll and others 2004; Koyama and others 2007; Tado and others 2014), as well as its association with cerebral edema after TBI (Gao and others 2015; Koyama and others 2007; Tado and others 2014). For example, while Tado and colleagues found that neutralizing VEGF after experimental TBI had no effect on cerebrovascular permeability and cerebral edema (Tado and others 2014), Gao and colleagues reported that anti-VEGF treatment in severe TBI mice lowered BBB permeability and reduced brain contusion (Gao and others 2015). Taken together, the therapeutic potential of VEGF in mTBI holds promise but requires further clarification.
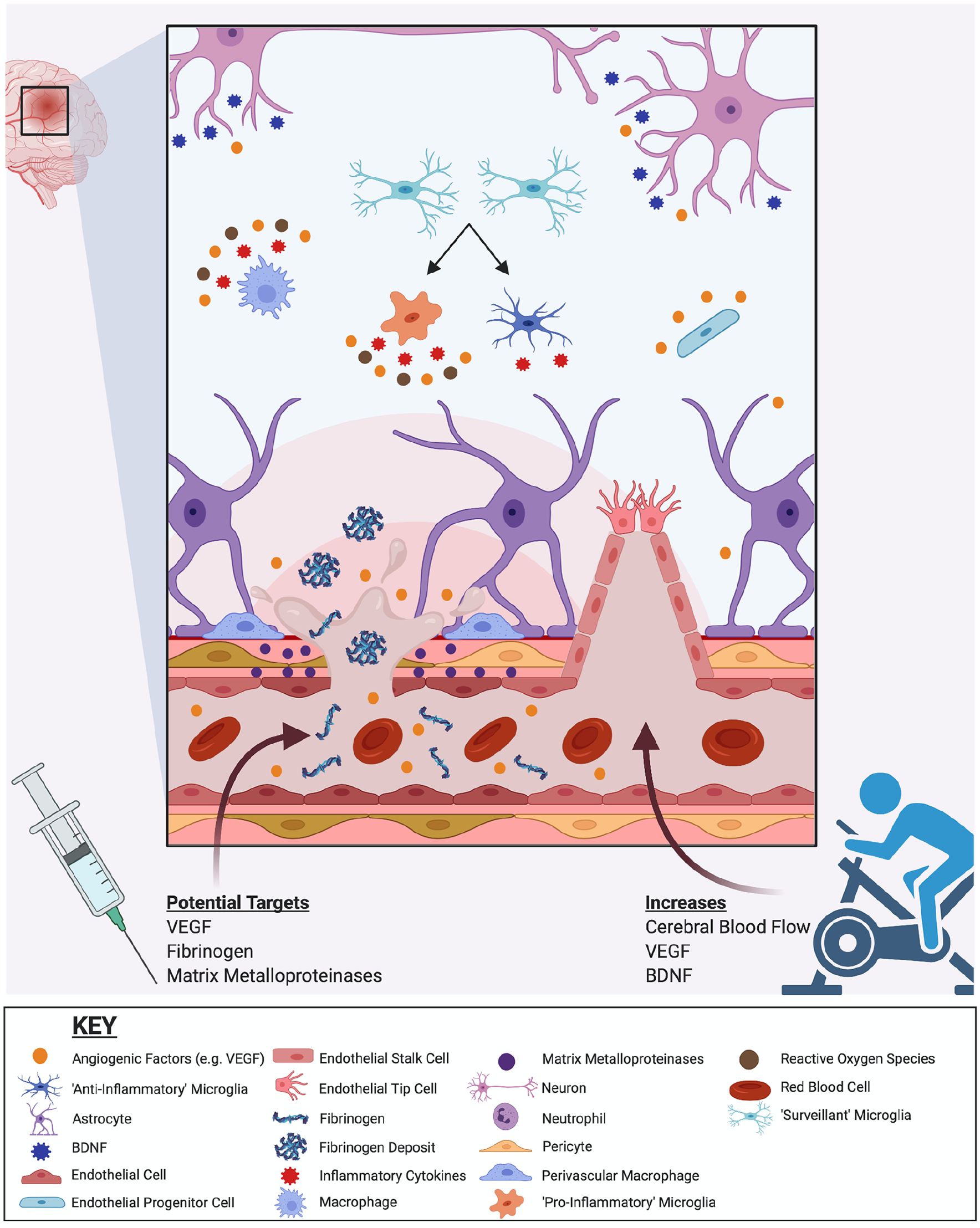
Endogenous and exogenous processes that may facilitate cerebrovascular recovery after mild traumatic brain injury (mTBI). Endogenous cerebrovascular repair after mTBI includes angiogenesis that is supported by a range of cells (e.g., endothelial cells, immune cells, glia, neurons) and mediated by angiogenic signaling pathways (e.g., vascular endothelial growth factor [VEGF], hypoxia-inducible factor, and prolyl-hydroxylases). Endothelial tip and stalk cells are involved in guiding and extending the sprout toward the source of angiogenic factors. mTBI-induced changes in cerebral blood flow evolve and recover endogenously over time. Recovery of the cerebrovascular system may also be facilitated by exogenous interventions. Exercise can increase cerebral blood flow, as well as VEGF and BDNF levels, and may improve mTBI recovery. Although not yet rigorously studied, pharmaceutical interventions that target VEGF, matrix metalloproteinase, fibrinogen, and other related pathways (e.g., neuroinflammation, oxidative stress) pathways may also be beneficial and warrant further investigation.
As detailed above (see Figs. 1, 2, and 4), fibrinogen and MMPs can also contribute to CVI and represent potential therapeutic targets to reduce BBB breakdown after mTBI. Promisingly, acute inhibition of MMP-9 has been shown to attenuate BBB hyperpermeability following mTBI in mice (Alluri and others 2016). Using in vitro models of BBB, inhibition of MMP-9 was found to attenuate BBB permeability induced by IL-1β (Alluri and others 2016). A study in mice found that mTBI-induced fibrinogen deposition was mitigated in MMP-9 gene knockout mice, and this was accompanied by improved memory outcomes (Muradashvili and others 2015). Taken together, these initial findings suggest that targeting fibrinogen and MMP-9 could be a promising treatment approach in mTBI, although further investigation is needed. These strategies could also decrease downstream neuroinflammation and oxidative stress, further improving mTBI outcomes.
Conclusions
There is growing evidence that CVI, including microbleeds, CBF dysregulation, and BBB dysfunction, are common pathophysiological features of mTBI that can contribute to poor outcomes. However, to date this remains an understudied topic and future studies are necessary to provide insight into a number of important questions. For example, studies are required to comprehensively characterize the temporal and spatial profile of CVI and recovery, and how this relates to promising blood and imaging biomarkers. Animal models of mTBI could be particularly useful in this context as they would allow for the validation of these biomarkers against direct measures of CVI in brain tissue. Another important area of future study is how CVI contributes to vulnerability to PPCS, repeated mTBIs, and neurodegeneration. The information gained from such studies would provide a foundation to rigorously examine the utility of promising interventions that may promote CVI recovery. Another important factor that should be incorporated into future studies is the role of biological sex, particularly considering that initial evidence suggests sex differences in CVI after mTBI (Bai and others 2019; Hamer and others 2020; Major and others 2020; Thibeault and others 2019). In addition, as aging has an influential effect on the deterioration of the cerebrovasculature (Takechi and others 2013; Yang and others 2020), future studies should include both aged and non-aged subjects to investigate CVI after mTBI (Sun and others 2020). Considering the current lack of effective biomarkers and interventions for mTBI, these future CVI studies are warranted and could help transform the clinical management and treatment of these common injuries.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: SRS, RM, and SJM are supported by funding from the Australian National Health and Medical Research Council.