
Abstract
Enhancers are cis-acting elements that control the transcription of target genes and are transcribed into a class of noncoding RNAs (ncRNAs) termed enhancer RNAs (eRNAs). eRNAs have shorter half-lives than mRNAs and long noncoding RNAs; however, the frequency of transcription of eRNAs is close to that of mRNAs. eRNA expression is associated with a high level of histone mark H3K27ac and a low level of H3K27me3. Although eRNAs only account for a small proportion of ncRNAs, their functions are important. eRNAs can not only increase enhancer activity by promoting the formation of enhancer-promoter loops but also regulate transcriptional activation. Increasing numbers of studies have found that eRNAs play an important role in the occurrence and development of brain diseases; however, further research into eRNAs is required. This review discusses the concept, characteristics, classification, function, and potential roles of eRNAs in brain diseases.
Introduction
Among noncoding DNA sequences, enhancers, as cis-acting elements, participate in the regulation of gene expression to ensure the normal development, differentiation, and apoptosis of cells (Ding and others 2018). Initial research found that under the effect of RNA polymerase II, enhancers are transcribed into noncoding RNAs termed enhancer RNAs (eRNAs). The distance between enhancers and their cognate promoters can vary from a few kilobases to hundreds of kilobases (Natoli and Andrau 2012). Previous reports also identified transcription sites outside of protein-coding genes that produce very low abundance transcripts during endotoxin activation in macrophages (De Santa and others 2010). The expression of eRNAs correlates with the presence of histone mark H3K27ac, but the absence of H3K27me (Rada-Iglesias and others 2011). In addition, enhancers are widely distributed in noncoding sequences of the genome, and thus are more difficult to determine and classify.
With the development of RNA sequencing, a large number of studies have found that eRNAs can regulate gene expression by activating enhancers (De Santa and others 2010). The regulation of enhancer activities indicates a potential target for disease treatment. Previously, enhancers and super-enhancers were research foci in cancer, whereas it was proposed recently that eRNAs affect the function of neurons, and mutations of enhancers might lead to nervous system diseases (Bhattarai and others 2021; Hauberg and others 2019; Le Gras and others 2017; Petrij and others 1995; Roelfsema and others 2005). This review focuses on the definition, characteristics, classification, and biological functions of eRNAs and discusses how eRNAs lead to neurodevelopmental diseases, neurodegenerative diseases, and other diseases related to the brain.
Overview of eRNAs
Properties of eRNAs
The properties of eRNAs are as follows:
eRNAs are transcribed from enhancers characterized by a high level of H3K4me1 and H3K4me2, relative to H3K4me3 (Kim and others 2010).
These enhancer regions are bound by lineage-determining transcription factors, associated mediator subunits, histone acetyltransferase p300/CBP, and cAMP response element binding protein (CEBP) (Lam and others 2013).
Expression of eRNAs is associated with a high level of H3K27ac and a low level of H3K27me3 (Rada-Iglesias and others 2011).
eRNAs regions are enriched with the transcriptional initiation complex and serine 5 phosphorylated RNA polymerase II. By contrast, enhancer regions are less enriched for serine 2 phosphorylated RNA polymerase II (Koch and others 2011).
eRNAs have shorter half-lives than mRNAs and long non-coding RNAs (lncRNAs); however, the frequency of transcription of eRNAs is close to that of mRNAs (Lam and others 2013).
Enhancer regions mostly show bidirectional transcription (2D-eRNAs), resulting in an eRNA exhibiting a 5′ cap without splicing or polyadenylation (De Santa and others 2010; Gil and Ulitsky 2018; Hah and others 2013; Natoli and Andrau 2012), while a small number of eRNAs are unidirectional (1D-eRNAs), polyadenylated, and spliced (Djebali and others 2012).
The Discovery of eRNAs
The identification and functional studies of individual eRNAs should first identify enhancers at the whole genome level. In 2010, Kim and colleagues used genome-wide sequencing methods to study stimulus-dependent enhancer function in neurons, which revealed widespread transcription at active enhancers (Kim and others 2010). Contemporaneous research conducted by De Santa and colleagues showed similar results (De Santa and others 2010). However, until recently, it was unknown whether eRNAs have biological functions (Kim and others 2010). In addition to these two studies, many studies have investigated eRNAs. A large number of approaches have been used to sequence eRNAs, including global run-on sequencing (GRO-seq; Fang and others 2021; Kim and others 2018; Rahnamoun and others 2017; Wang and others 2011), precision run-on nuclear sequencing (PRO-seq; Mahat and others 2016), cap analysis gene expression (CAGE; Arner and others 2015; Zhang and others 2020b), locked nucleic acids (LNA; Roux and others 2017), chromatin-immunoprecipitation sequencing (ChIP-seq; Kim and others 2020; Zhang and others 2020b; Zhao and others 2016), three-dimensional DNA selection and ligation (3D-DSL; Harismendy and others 2011), and deoxyribonuclease I (DNase I)-hypersensitive site sequencing (Dnase-seq; Boyle and others 2008). The development of these techniques has deepened our understanding of eRNAs. These studies revealed that approximately 40,000 to 65,000 eRNAs are expressed in human cells, accounting for a large proportion of initiation events (Arner and others 2015).
The Classification of eRNAs
To date, the major types of eRNAs discovered are non-polyadenylated eRNAs (polyA− eRNAs, 2D-eRNAs) and polyadenylated eRNAs (polyA+ eRNAs, 1D-eRNAs; Djebali and others 2012). PolyA− eRNAs are shorter than polyA+ eRNAs, and are bidirectionally transcribed, while polyA+ eRNAs are unidirectionally transcribed (De Santa and others 2010; Gil and Ulitsky 2018; Natoli and Andrau 2012; Sartorelli and Lauberth 2020; Table 1, Fig. 1). In addition, polyA− eRNAs tend to be capped, which was demonstrated by 5′ GRO-seq and CAGE (Core and others 2014; Lam and others 2013).
Classification and Characteristics of Enhancer RNAs (eRNAs).
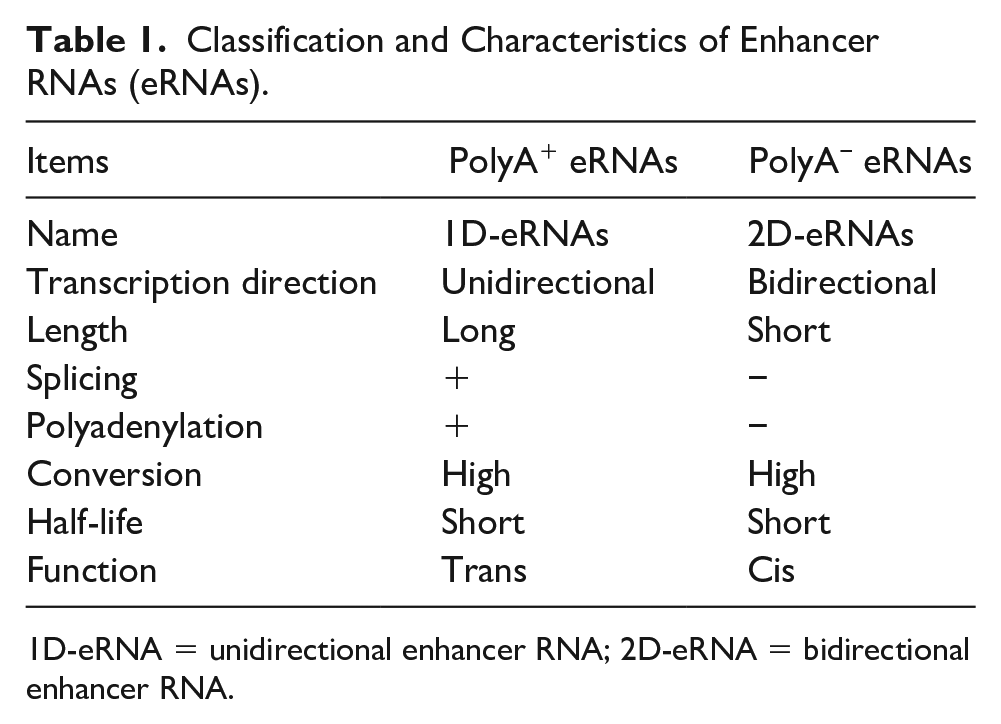
1D-eRNA = unidirectional enhancer RNA; 2D-eRNA = bidirectional enhancer RNA.
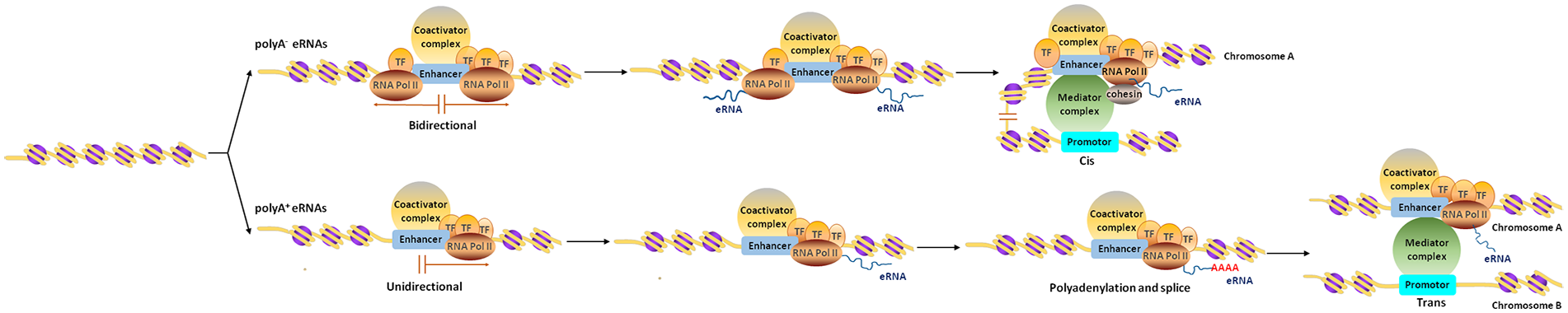
Schematic diagram of the synthesis and function of eRNAs. eRNAs, classified as polyA− and polyA+ eRNAs, are transcribed from enhancers. The polyA− eRNAs are bidirectional and function in cis without polyadenylation and splicing. The polyA+ eRNAs are unidirectional and function in trans after polyadenylation and splicing.
eRNAs can also be categorized into two subgroups: long noncoding enhancer RNAs (lnc-eRNAs) and the other subtype of eRNAs. Compared with the other subtype of eRNAs, lnc-eRNAs are more stable and share more features with lncRNAs. Specifically, lnc-eRNAs are annotated transcripts whose initiation sites overlap enhancer regions with particular histone modification patterns, such as H3K4me1 and H3K27ac, and are currently recorded in lncRNA databases, whereas the other subtype of eRNAs comprise annotated transcripts from enhancer-like regions and are not currently recorded in lncRNA databases (Li and others 2016).
Currently, it is believed that eRNAs probably occur within ~150 nucleotides (nt) of the enhancer transcription start site, which was predicted by PRO-seq and Start-seq datasets (Austenaa and others 2015; Henriques and others 2018). The length of eRNAs provides a way of classifying them, marking eRNAs as a class of RNA molecules whose transcripts are more than 200 nt in length (Ding and others 2018; Guttman and Rinn 2012; Ulitsky and Bartel 2013). However, the classification of lncRNAs and eRNAs cannot completely depend on their length, because the relationship between lncRNAs and eRNAs is not totally exclusive. Some studies demonstrated that in certain cases, lncRNAs might have evolved from eRNAs (Alvarez-Dominguez and others 2017; Gil and Ulitsky 2018; Kopp and Mendell 2018; Paralkar and others 2016). Further studies are needed to clarify the relationship between eRNAs and lncRNAs, and the classification of eRNAs.
Functions of eRNAs
Although eRNAs have been recognized as the most significant markers of activated enhancers, scholars have different views on the function of eRNAs. One of the controversies is whether eRNAs are markers of activated enhancers (Carullo and others 2020), and have corresponding functions in the link between gene expression and transcription. Some scholars (Li and others 2016) believe that eRNAs are transcriptional noise, and have no specific function in increasing the activity of enhancers. They suggest that most eRNAs originate from the random initiation mechanism of RNAPII and are recruited to nearby enhancers only as part of the transcription mechanism. However, using transcription inhibitors and gene knockdown techniques, it has been found that eRNAs can activate enhancers, thereby regulating local gene expression (Heward and others 2015; Kaikkonen and others 2013; Natoli and Andrau 2012).
Public Databases Related to eRNAs
Some public databases are available for the analysis of eRNAs, including HeRA (Zhang and others 2021), TiED (Xiong and others 2018), eRic (Zhang and others 2019), and TCeA (Chen and Liang 2020; Table 2). These databases provide information related to eRNAs, which are useful to predict tissue-specific eRNAs, annotate connections between eRNAs and human complex diseases, and explore eRNA regulation.
Annotation of Public Databases Related to Enhancer RNAs (eRNAs).

Enhancers Are Activated by eRNAs via Regulating Chromatin Looping
The transcription of eRNAs is regulated negatively by Rev-Erbs (retinoic acid receptor-related orphan receptors) in macrophages, targeting them for degradation, and the expression of nearby mRNA was reduced (Lam and others 2013). It was speculated that the inhibitory function of Rev-Erb nuclear receptors at a distance might be caused by repressing eRNA transcription, indicating that eRNAs have a function in stimulating enhancer activity (De Santa and others 2010; Heward and others 2015; Kaikkonen and others 2013; Lam and others 2013). In terms of improving enhancer activity, eRNAs have been found to increase enhancer activity by promoting the formation of enhancer-promoter loops (Lin and others 2012). Knockdown of eRNAs might damage these loops (Hsieh and others 2014; Lai and others 2013; Li and others 2013; Pnueli and others 2015). Moreover, enhancers looped to the promoter of protein-coding genes have higher eRNA expression (Sanyal and others 2012).
The α-estrogen receptor (ERα) in MCF-7 breast cancer cells can promote the production of eRNAs in an estrogen-dependent manner. Furthermore, eRNAs can stabilize the enhancer-promoter loop by interacting with binding loop forming factors (adhesin and the mediator complex; Li and others 2014). A similar mechanism of action also applies to androgen-induced eRNAs (Hsieh and others 2014). Specifically, the androgen receptor (AR) in prostate cancer cells promotes the production of eRNAs by binding to enhancer regions in an androgen-dependent manner. The eRNA transcribed from the upstream enhancer of KLK3 (encoding kallikrein-related peptidase 3) can promote the interaction between the KLK3 enhancer and the KLK2 promoter, thereby increasing long-distance KLK2 transcriptional activation (Hsieh and others 2014). Estrogen-induced eRNA is a necessary condition to promote the enhancer-promoter loop and plays an important role in estrogen-upregulated coding genes through an interaction with cohesion, which ultimately increases the stability of the enhancer-promoter loop (Hah and others 2013).
To date, many studies have proved the reliability of the eRNA-dependent chromosome circularization model, indicating that eRNAs play an important role in the formation and stabilization of enhancer-promoter loops (Lai and others 2013; Lai and others 2015; Li and others 2013). However, there are still disagreements regarding their mechanism of action in gene transcription. Schaukowitch and colleagues found that chromatin cyclization was not affected by eRNA knockdown, and eRNAs could act as a bait for the negative elongation factor (NELF) complex by inducing immediate early genes (IEG) in neurons, thereby promoting the transition from suspended RNA polymerase II to production elongation (Schaukowitch and others 2014). Moreover, Hah and colleagues found that eRNAs promoted the release of Pol II by inducing the NELF complex (Hah and others 2013). RNA polymerase II was used as an extension inhibitor to successfully reduce the number of eRNAs synthesized at two sites and the transcription of the coding gene, without significantly affecting the formation of enhancer-promoter loops at the P2RY2 or GREB1 loci. This phenomenon might be attributed to interference by small interfering RNA (siRNA) with the expression of eRNAs and the inhibition of RNA polymerase II extension, thereby affecting the expression of eRNAs. A recent study also confirmed the above view, that is, not all eRNAs have enhancer-promoter loops, and they might only exist in certain enhancers with stimulus-induced eRNAs (Hah and others 2013).
Regulation of Gene Expression by eRNAs
The expression level of eRNAs is usually related to the expression of nearby genes, which indicates their potential role in gene transcription (De Santa and others 2010; Kaikkonen and others 2013; Lai and Shiekhattar 2014; Li and others 2013). By observing the transcription of eRNAs of hypersensitive site 2 (HS2) in the β-globin locus control region (LCR), Gurumurthy and colleagues reported that depletion of HS2 3′ eRNA led to a reduction in β-globin gene transcription and the recruitment of RNA polymerase II (Gurumurthy and others 2021). As a hypersensitive site in the β-globin LCR, inserting the lac operon/R repressor complex downstream of the HS2 enhancer terminated HS2-mediated transcription and reduced the promoter activity of the target gene (Ling and others 2004). Similarly, inserting the transcription termination sequence TerF between the promoters of the β-globin LCR and human growth hormone (hGH) genes resulted in lower expression of hGH in transgenic mice, confirming the potential impact of eRNAs on gene transcription (Ho and others 2006). Before transcriptional induction of the β-globin locus with chemical treatment, levels of RNA polymerase II were very low in the LCR HS loci. After induction, however, higher levels of RNA polymerase II were observed (Kim and others 2015). These findings indicated that chromatin circularization and eRNA transcription precede gene transcription and activation, suggesting that there is a correlation between enhancer transcription and promoter and gene expression (Alvarez-Dominguez and others 2014; Kim and others 2015).
Recently, new evidence has indicated that eRNAs are directly involved in transcriptional activation and play corresponding functional roles (Alvarez-Dominguez and others 2014; Hsieh and others 2014; Lai and Shiekhattar 2014; Li and others 2013; Melo and others 2013; Mousavi and others 2013; Pnueli and others 2015). Small interfering RNAs, short hairpin RNAs, and locked nucleic acid oligonucleotides have been adopted by different studies to knockdown eRNAs, and it was observed that the transcription of the coding genes homologous to the eRNAs was reduced, thus proving the important function of eRNAs in the transcription process. Using the “bundling” strategy to fuse RNA to eRNAs, chimeric RNAs were synthesized successfully. Such eRNAs can bind to the recombinant adaptor protein, and then combine with the specific sequence of the reporter plasmid vector (Li and others 2013; Melo and others 2013). These findings revealed that the transcriptional activity of the combined gene increased, suggesting that eRNAs can activate the transcription of downstream genes.
Many studies have explored the mechanism by which eRNAs affect gene activation. Among them, some studies have proposed a mechanism called “transcription factor trapping,” pointing out that there are many types of eRNAs that have the ability to bind to transcription factors (TFs), resulting in, for example, enhanced binding of TF YY1 to its enhancer DNA (Sigova and others 2015). Some studies reported that the state of chromatin might be related to eRNAs (Liang and others 2016; Pnueli and others 2015; Yang and others 2016). Short hairpin RNA (shRNA) knockdown of the MYC-428 and -525 ESE eRNAs caused growth arrest, reduced cell growth of lymphoblastoid cell (Sigova and others 2015), and increased histone H3 occupancy at the enhancer region, resulting in a loss of H3K4m3 at the promoter region (Liang and others 2016). Furthermore, Mousavi and colleagues reported that eRNAs transcribed from the distal regulatory region and core enhancer are involved in a cell-type-specific transcriptional circuit by regulating the accessibility of chromatin (Mousavi and others 2013).
ERNAs and Brain Diseases
eRNAs have been identified to act as epigenetic regulators that control the chromatin state at genomic loci, and are involved in the development of a series of human diseases, such as brain injuries, cancer, and immuno-metabolic diseases (Bhattarai and others 2021; Chen and Liang 2020; Fang and others 2020; Guo and others 2021; Huang and others 2021; Kim and others 2010; Mousavi and others 2013; Zhang and others 2020a). For example, lnc-eRNA SEELA, localized on chromatin, mediates cis-activated transcription of SERINC2 (serine incorporate 2) via directly interacting with histone H4 to strengthen histone recognition and enhancer transcription. Furthermore, the SEELA-SERINC2 axis drives the initiation and progression of leukemia in vivo and in vitro by influencing sphingolipid metabolism (Fang and others 2020). Several reviews of eRNAs in cancer (Adhikary and others 2021; Lee and others 2020) have been published; therefore, in the present study, we focused on the role of eRNAs in nervous system diseases.
ERNAs in Neurodevelopmental and Neurodegenerative Diseases
Autism spectrum disorder (ASD) is a type of neurodevelopmental disorder with a large number of genetic variants. Genome-wide association study (GWAS) of ASD has shown that the risk variants are located in intronic and intergenic regions (Anney and others 2010; Anney and others 2012; Weiss and others 2009). Notably, ASD-related gene variants are enriched in enhancer regions expressing eRNAs in specific brain regions, including the frontal, temporal, and occipital regions, and the cerebellar cortex (Yao and others 2015). These findings suggested that genetic variants in noncoding sequences might play a role in ASD susceptibility by regulating the expression of eRNAs and/or their target genes.
Fragile X syndrome (FXS) is the main genetic cause of autism, which is characterized by the deficiency of the fragile X mental retardation protein (FMRP). The phosphoinositol-3-kinase (PI3K) enhancer (PIKE) is the target of this protein and is upregulated in Fmr1 knock-out (KO) mice (Romero-Zerbo and others 2009). The increased expression of the enhancer PIKE plays an important role in mediating the activation of group 1 metabotropic glutamate receptors (mGlu1/5) signaling, giving rise to neuronal defects in FXS in Centg1 KO mice (Gross and others 2015). The topologically associating domain (TAD) boundary around the Fmr1 gene, which is disrupted in patients with FXS, correlates with FMR1 silencing. The possible mechanism might be that the absence of CCCTC binding factor (CTCF) occupancy affects chromatin looping to cause aberrant gene expression (Sun and others 2018; Zhang and others 2020a).
Rubinstein-Taybi syndrome (RSTS) is a rare neurodevelopmental disorder characterized by intellectual disability, postnatal growth retardation, and skeletal abnormalities. Both the CREB binding protein (CBP) and p300 are transcriptional co-activators that regulate gene expression and enhancer activities. Mutations of the genes encoding CBP or p300, with the loss of histone acetyltransferase activity, are main causes of RSTS (Petrij and others 1995; Roelfsema and others 2005), which might result in loss of function or gain of function of eRNAs.
Huntington’s disease (HD) is a neurodegenerative disease characterized by neuropathy of the spine neurons in the striatum, and the pathogenesis of HD is mainly related to neuronal death caused by the downregulation of transcription in neurons in the striatum (Hodges and others 2006). The expression of downregulated genes is regulated by super-enhancers, which are marked by H3K27 acetylation (H3K27ac) and RNAPII (Achour and others 2015). Compared with wild-type mice, eRNAs levels in the striatum were decreased in HD mice. The reduced eRNAs were highly enriched in biological processes, such as synaptic plasticity and neuronal transmission and learning, and correlated with genes involved in striatal neuron identity and loss of RNAPII binding sites (Le Gras and others 2017). Therefore, the targeted improvement of eRNA levels might provide inspiration for new treatments of HD.
ERNAs and Psychiatric Diseases
Anxiety is a neuropsychiatric illness that can cause hippocampal atrophy and disruption of the amygdala-prefrontal cortex circuit (Mah and others 2016; Sun and others 2020; Wyse and others 2020). In a rat anxiety model induced by adolescent intermittent ethanol (AIE) exposure, histone H3 deacetylation was induced, which led to the downregulation of synaptic plasticity-associated genes in the amygdala, such as brain-derived neurotrophic factor (Bdnf) and activity-regulated cytoskeleton-associated (Arc) (Pandey and others 2015). An evolutionarily conserved intronic enhancer region in the Bdnf gene can initiate bidirectional transcription of eRNAs and regulate both basal and stimulus-dependent expression of Bdnf exon I-, IIc-, and III-containing transcripts in mouse and rat neurons. The neuron-specific activity of the intronic enhancer region is modified by a functional E-box element in the enhancer (Tuvikene and others 2021).
The Arc enhancer, located 7 kb upstream of the transcription start site, is necessary for Arc eRNA synthesis (Kim and others 2010; Schaukowitch and others 2014). In response to neuronal activity, the Arc enhancer was brought into the proximal promoter, leading to a rapid increase in Arc eRNA levels, which facilitated the release of the NELF complex from immediate early genes in neurons (Schaukowitch and others 2014). By contrast, after AIE exposure, the expression levels of Arc mRNA and eRNA decreased in the amygdala of adult rats, accompanied by a reduced number of synapses in the central nucleus of the amygdala and the medial nucleus of the amygdala, in addition to increased anxiety-like behaviors. The suppression of Arc eRNA might be attributed partly to epigenetic reprogramming, because the abundance of H3K27me3 increased and lysine demethylase 6B (KDM6B), CBP, and H3K27ac signals decreased, triggering condensed chromatin and the binding of NELF to the promoter of the Arc gene (Kyzar and others 2019; Fig. 2).
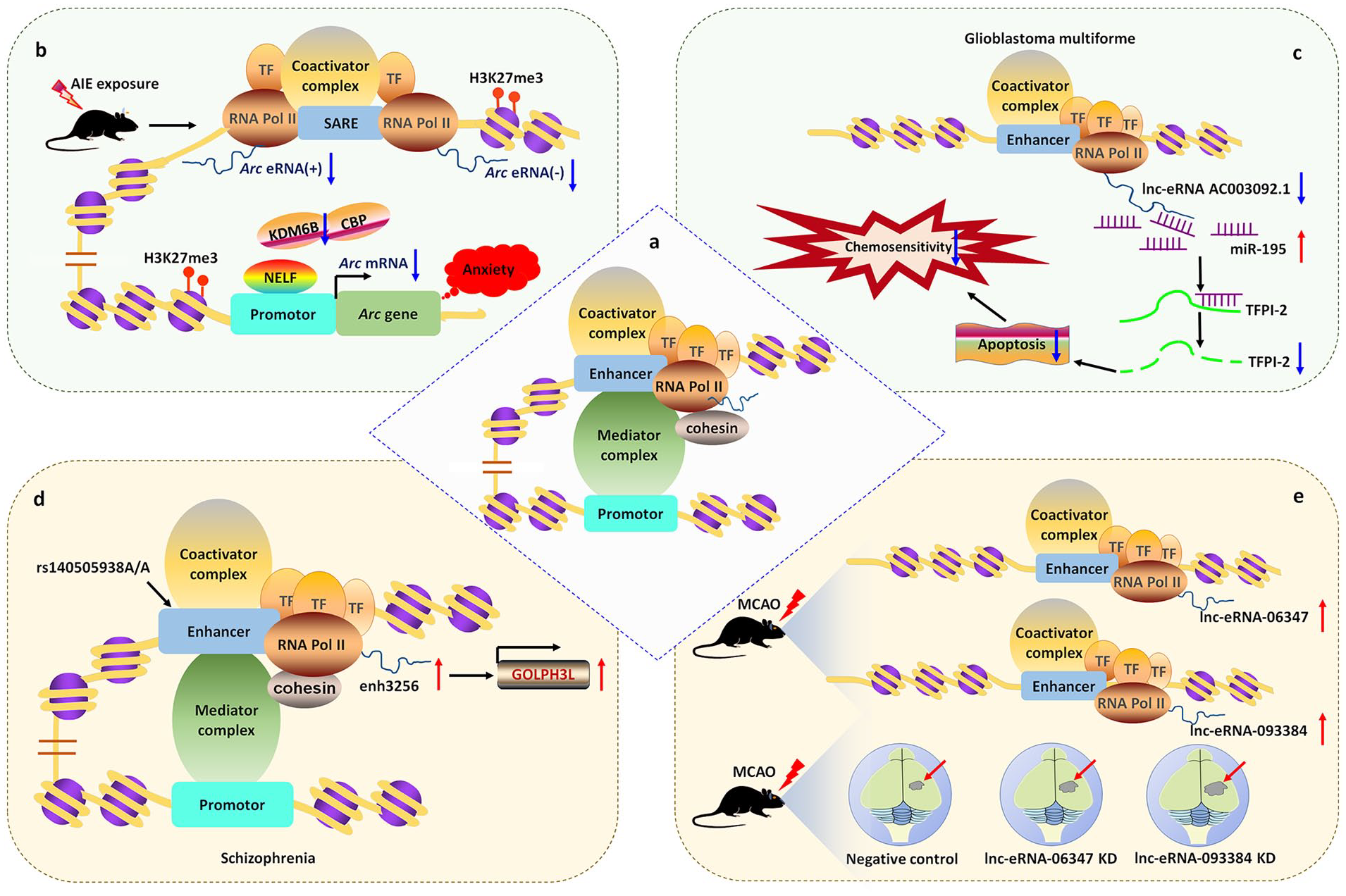
Mechanisms of eRNAs in brain diseases. (a) Schematic diagram of eRNA transcripts and their potential function. (b) Adolescent intermittent ethanol (AIE) exposure increases H3K27me3, suppresses the expression of lysine demethylase 6B (KDM6B) and CREB binding protein (CBP) in the Arc promoter region, which is associated with low levels of Arc mRNA and eRNA in the amygdala of adult rats. This triggers negative elongation factor (NELF) binding to the promoter of Arc gene, and finally confers anxiety susceptibility. (c) In glioblastoma multiforme, lnc-eRNA AC003092.1 was downregulated. LncRNA AC003092.1 functions as a miR-195 sponge in tissue factor pathway inhibitor-2 (TFPI-2)-mediated chemosensitivity. (d) In schizophrenia, an index polymorphism rs140505938, located within the enhancer sequence activates the enhancer activity of schizophrenia-associated eRNA enh3256, which promotes the expression of its target gene GOLPH3L (encoding Golgi phosphoprotein 3-like). (e) In a mouse model of middle cerebral artery occlusion (MCAO), certain lnc-eRNAs are differentially expressed. Among them, the levels of lnc-eRNA_06347 and lnc-eRNA_093384 were significantly higher after injury. Knockdown (KD) of the lnc-eRNAs resulted in a larger post-stroke infarct volume.
Schizophrenia is a chronic brain disorder that is caused by the complex interplay between heredity and environmental factors. Interestingly, some genetic variants of schizophrenia are located in noncoding regions with enhancer-like activity (Dong and others 2018; Schizophrenia Working Group of the Psychiatric Genomics Consortium 2014). The variant rs4420550, located within an enhancer region affecting chromatin accessibility and cell proliferation, regulates the transcription of genes on chromosome 16p11.2 by interacting with the promoter region, ultimately leading to the onset of schizophrenia (Chang and others 2021). By performing RNA sequencing of the dorsolateral prefrontal cortex from patients with schizophrenia (n = 258) and control subjects (n = 279), a gene expression resource and its genetic regulation were created. After analyzing the data using state-of-the-art methods, enrichment of cis-expression quantitative trait loci (eQTLs) within enhancer sequences was observed in human brain tissues. Altered gene expression induced by schizophrenia risk loci changed neurodevelopment and neural migration (Fromer and others 2016). Based on the regulatory nature of schizophrenia risk variants, it is plausible that schizophrenia-linked eRNAs might affect downstream changes in gene expression. Using an assay for transposase-accessible chromatin followed by sequencing, 118 eRNAs that were differentially transcribed in schizophrenia were detected and schizophrenia-associated gene/eRNA co-expression modules were identified. The differentially expressed transcripts were enriched in certain common signaling pathways, such as the Roundabout receptors pathway, which is involved in axon repulsion and neurogenesis (Blockus and Chedotal 2016). Data from brain gene eQTLs and enhancer eQTLs demonstrated that genetic polymorphisms affected the expression of 927 enhancers. Among them, an index polymorphism, rs140505938, located within the enhancer sequence, altered the enhancer activity of the schizophrenia-associated eRNA enh3256, which regulated the expression of its target gene GOLPH3L (encoding Golgi phosphoprotein 3-like; Hauberg and others 2019; Fig. 2).
ERNAs and Brain Tumors
Using RNA-sequencing data from the Cancer Genome Atlas database, 70 lnc-eRNAs were identified to be prognosis-related transcripts of glioblastoma multiforme (GBM). Among them, AC003092.1 was demonstrated as an immune-related lnc-eRNA, with low AC003092.1 expression predicting a better outcome. CIBERSORTx and single-sample gene-set enrichment analysis showed that high AC003092.1 expression occurred mainly in monocytes, neutrophils, immune cells, and stromal cells, while low AC003092.1 expression occurred mainly in naive B cell, T cells, follicular helper cells, M1 macrophages, and GBM tumor cells (Guo and others 2021). The low levels of lnc-eRNA AC003092.1 in GBM correlated with temozolomide resistance, higher risk of relapse, and poor prognosis. Overexpression of lnc-eRNA AC003092.1 promoted temozolomide sensitivity, stimulated cell apoptosis, and suppressed cell proliferation through targeting tissue factor pathway inhibitor-2 (TFPI-2). Mechanistically, lnc-eRNA AC003092.1 functions as a competing endogenous RNA to regulate TFPI-2 expression by sponging miR-195 (Xu and others 2018; Fig. 2).
ERNAs and Brain Injuries
Previous studies have shown that lncRNA expression levels were altered significantly in patients and animal models of ischemia, involving various functions, such as development, apoptosis, and neuroinflammation (Bhattarai and others 2017; Dykstra-Aiello and others 2016). Using genome-wide RNA-sequencing, Bhattarai and colleagues discovered differentially expressed lncRNAs in the mouse cortex following transient focal ischemia. Most of the lncRNAs were derived from non-genic loci, possibly included regulatory elements, such as enhancers (Bhattarai and others 2017). Therefore, it is possible that some of the lncRNAs might be transcribed from enhancers activated in response to ischemic injury and could have downstream regulatory effects on other ischemic response genes. To confirm this hypothesis, Bhattarai and colleagues identified stroke-induced lnc-eRNAs in the cerebral cortex of male mice during ischemia using genome-wide H3K27AC ChIP-sequencing and genome-wide RNA-sequencing. The authors found that the expression of 77 lnc-eRNAs changed significantly during the occurrence of cerebral ischemia, of which 55 were exclusively expressed in cerebral ischemia. When two lnc-eRNAs (lnc-eRNA_06347 and lnc-eRNA_093384) were knocked down, the poststroke infarct volumes increased by 13.63% and 19.51%, respectively, indicating that the lnc-eRNAs might protect against poststroke cortical brain damage. Interestingly, the expression of stroke-induced lnc-eRNAs revealed sex-dependent patterns, with lnc-eRNA_160141, lnc-eRNA_156140, and lnc-eRNA_04759 showing significantly lower expression levels in the poststroke female cortex compared with those in the poststroke male cortex (Bhattarai and others 2021; Fig. 2).
Conclusion
As cis-acting elements that regulate the transcription of target genes, the regulatory functions of enhancers are affected by eRNAs. With the development of eRNA sequencing research, our understanding of the biological function of eRNAs is increasing rapidly. eRNAs could improve enhancer activities by promoting the formation and stabilization of enhancer-promoter loops to regulate gene expression. Moreover, eRNAs could be directly involved in transcriptional activation of nearby genes via various mechanisms, such as TF trapping. Studies have revealed the respective mechanisms by which eRNAs regulate target gene expression in different enhancer regions and in different research systems. In this review, discussion of the functions of enhancers without transcription is an exception. Nevertheless, several questions remain: How do eRNAs without enhancer-promoter loops activate enhancers? How do eRNAs regulate remote gene expression? In addition, we still lack a widely accepted hypothesis concerning how eRNAs regulate gene expression. Moreover, the study of eRNAs still faces some difficulties. For example, enhancers are widely distributed in noncoding sequences of the genome, making their identification and classification difficult. After gaining a deeper understanding of the regulatory role of eRNAs in gene expression, we found that the mutation of enhancers is associated significantly with the development of neurodevelopmental diseases, neurodegenerative diseases, and other brain-related diseases, thus providing novel therapeutic targets. However, as mentioned above, the specific mechanisms by which eRNAs contribute to dysfunction of enhancers related to the increased risk of brain diseases remain unclear. Furthermore, how value of eRNAs in the treatment and prevention of disease is also unknown. Therefore, further studies are required to explore the utility of eRNAs in the treatment of human diseases.
Footnotes
Author Contributions
LG conceived the project and revised the manuscript. YS, ZH, RY, and YC performed the literature search and drafted the manuscript. QW helped with the preparation of figures.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (No. 32071182) and the Fundamental Research Funds for the Central Universities (No. 2018SCUH0053).