
Abstract
Brain-on-a-chip is a microphysiologic platform that comprises cultured brain cells to understand brain disease pathogenesis and treatment. The blood-brain barrier (BBB) of the neurovascular unit serves as a highly selective molecular transport interface for brain homeostasis. BBB dysfunction promotes neuroinflammation, exacerbates disease progression, and contributes to neurodegenerative diseases. However, the mechanisms of BBB disruption underlying brain disorders remain poorly understood; thus, developing neurotherapeutics that can effectively cross the BBB remains a major challenge. Recent advances in microfluidic brain-on-a-chip platforms now enable the creation of BBB-on-a-chip systems that replicate key structural and functional aspects of the human BBB under dynamic flow conditions. Integration of microelectrode arrays into these microfluidic systems enhances their utility by enabling high-throughput drug screening and targeted delivery, allowing real-time monitoring of neuronal activity and network behavior. Although current brain organoid, brain-on-a-chip, and BBB-on-a-chip platforms remain in developmental stages, significant progress has been made using induced pluripotent stem cell–derived neurons, astrocytes, endothelial cells, pericytes, and microglia from healthy individuals and patients with neurodegenerative diseases. This review highlights recent advances in brain- and BBB-on-a-chip technologies and their potential applications in studying disease pathogenesis and preclinical drug screening for neurodegenerative disorders.
Keywords
Introduction
Neurodegenerative disorders, such as Alzheimer disease (AD) and Parkinson disease (PD), are characterized by neuroinflammatory processes and progressive neuronal loss in which the blood-brain barrier (BBB) plays a crucial role (Yoon et al 2021; Kempuraj et al 2024a, 2024b). BBB dysfunction, neuroinflammation, and immune cell infiltration into the brain parenchyma are hallmarks of many central nervous system (CNS) disorders. The BBB in the neurovascular unit (NVU) is a highly selective complex formed by astrocytes, pericytes, and microvascular endothelial cells, which contributes to the maintenance of CNS homeostasis by regulating the passage of molecules and immune cells between the blood and brain parenchyma (Figure 1; Kempuraj et al 2024b). The integrity of the BBB complex is maintained by astrocytes (end feet), pericytes, endothelial cells, neurons, tight junction (TJ) proteins, adherens junction complexes, and gap junction proteins. This regulation is crucial for normal brain function (Kempuraj et al 2020, 2024b). Chronic damage to the NVU and BBB complex leads to disruption, increased permeability, and neuroinflammation in neurodegenerative disorders (Kempuraj et al 2024b). Therefore, creating an efficient BBB model that recapitulates human barrier function in healthy and diseased states is urgently needed for a better understanding of CNS pathologies and for evaluating neurotherapeutics (Pediaditakis et al 2026).
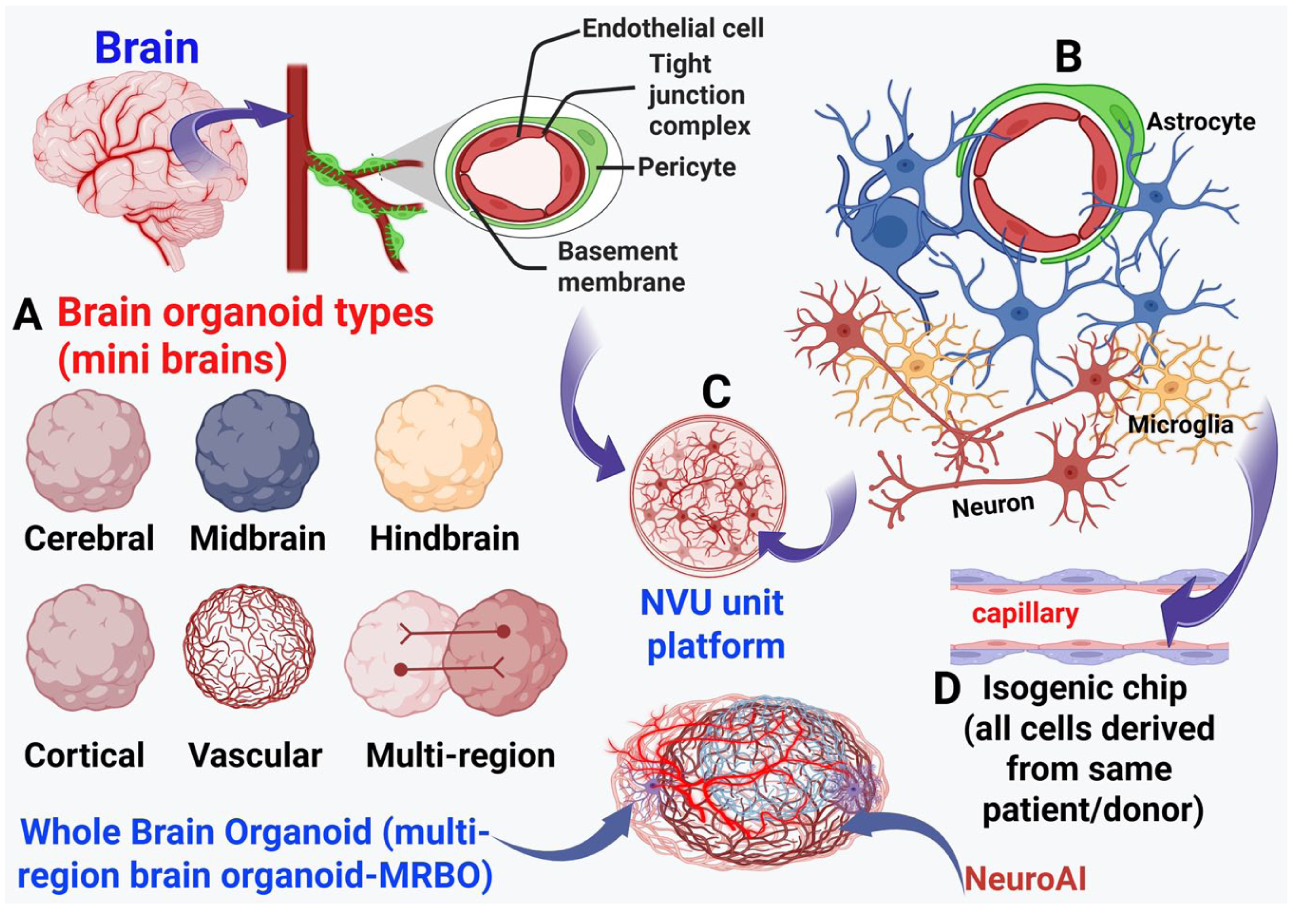
Schematic representation of the NVU/BBB-on-a-chip platform. The NVU includes the BBB with endothelial cells, astrocyte end feet, and pericytes. (A) Brain organoid types, such as cerebral, midbrain, hindbrain, cortical, vascularized, multiregion, and whole brain organoids. (B) NVU key connections/interactions with surrounding astrocytes, microglia, and neurons. Dysregulation of NVU/BBB function contributes to neurologic disorders, including neurodegenerative diseases such as Alzheimer disease and Parkinson disease. (C) Brain parenchyma includes NVU/BBB with astrocytes, microglia, neurons, and pericytes grown in vitro. (D) An isogenic NVU/BBB can be grown by using iPSC-derived cells from the same patient or healthy controls. NVU models may be established in standard culture dishes or on chip-based platforms using primary human cells, established cell lines, or iPSC-derived brain cells. Region-specific brain chips can be created by using region-specific brain cell types, such as cortical neurons, midbrain neurons, glutamatergic neurons, GABAergic neurons, or dopaminergic neurons, derived from patients or healthy controls for comparison. This can be integrated with NeuroAI. BBB, blood-brain barrier; iPSC, induced pluripotent stem cell; NeuroAI, Neuro Artificial Intelligence; NVU, neurovascular unit.
Traditional animal models and static in vitro systems have provided insights into BBB biology but often fall short in capturing the complexity and dynamic nature of the human BBB, thereby limiting the translation of laboratory findings to clinical applications (Palma-Florez et al 2023). These limitations hinder the translation of preclinical findings to clinical use. However, recent advances in organ-on-a-chip technologies have enabled the development of BBB-on-a-chip platforms that more closely mimic human BBB pathophysiology (Vetter et al 2025). These systems incorporate human astrocytes, pericytes, and endothelial cells in controlled in vitro conditions, which allow for improved real-time analysis of barrier function, cell migration, and drug permeability (Yoon et al 2021; Nair et al 2023). Fabrication of in vitro BBB-on-a-chip platforms utilizes a microfluidic chip, matrix materials such as hydrogels, brain cells such as brain microvascular endothelial cells, electrodes, permeable membrane with specific thickness, channels, creation of flow, shear stress, and specific cell culture medium (Vetter et al 2025). Therefore, BBB-on-a-chip represents a promising tool for investigating BBB dysfunction–related disease mechanisms and accelerating neurotherapeutic development. While the current brain-on-a-chip technologies have advanced significantly for modeling neurologic diseases, these models are still in an early development stage, and the existing neurospheroids (an aggregation of induced pluripotent stem cell [iPSC]–derived cells, such as neurons and astrocytes in the culture) do not yet closely recapitulate human brain tissue. Therefore, the development of more advanced, clinically important bioengineered models of human brain-on-a-chip for drug efficacy evaluation are needed (Staicu et al 2021; Cui and Cho 2022).
BBB and the NVU
The brain microvascular endothelial cells that form the BBB are sealed by TJ proteins such as occludin, claudin 5, and junctional adhesion molecules (Figure 2). In addition to limiting paracellular permeability, these TJ proteins support the brain’s ionic equilibrium (Kadry et al 2020; Lochhead et al 2020). Adherens junction proteins, such as vascular endothelial–cadherin, provide additional structural integrity to the endothelial barrier (Lochhead et al 2020). In addition, pericytes embedded in the basement membrane surrounding the small blood vessels control immune cell entry, vesicular transport, angiogenesis, and barrier tightening. Nearly the entire vascular surface is encircled by astrocytic end feet, which secrete trophic factors that maintain BBB phenotype and transporter expression, such as vascular endothelial growth factor, transforming growth factor β, glial cell line–derived neurotrophic factor, and angiopoietin 1 (Kadry et al 2020). Beyond endothelial cells, pericytes, and astrocytes, the NVU comprises neurons, microglia, and extracellular matrix components and operates under shear stress generated by blood flow (Kugler et al 2021). These components closely regulate metabolism, cytokine signaling, and dynamic variations in BBB permeability (Zlokovic 2008; Smith et al 2022).
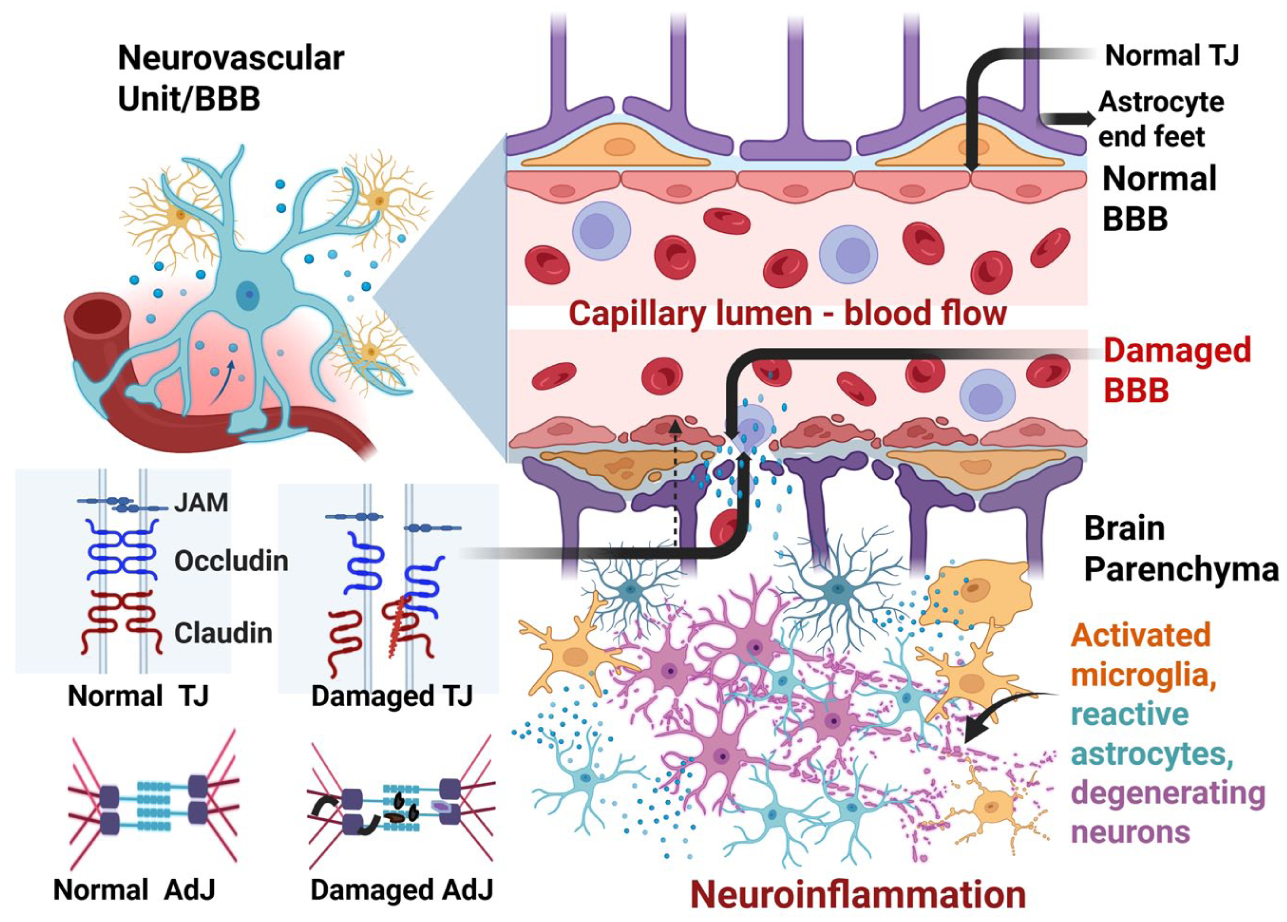
The BBB maintains brain homeostasis by regulating exchanges between the brain and blood, including nutrient transport. Disruption or damage to the BBB endothelial cells, pericytes, or astrocytic end feet increases BBB permeability and allows the entry of immune cells and pathogens from blood to brain parenchyma. Persistent BBB dysfunction can promote neuroinflammation and neurodegeneration, contributing to the onset and progression of neurological disease. BBB disruption may activate resident microglia and astrocytes, leading to release of various proinflammatory and neurotoxic mediators that exacerbate the neurodegeneration process. AdJ, adherens junction; BBB, blood-brain barrier; JAM, junctional adhesion molecule; TJ, tight junction.
The BBB protects the brain’s chemical environment through highly selective transport mechanisms. Nutrients travel through the BBB via solute carriers such as glucose transporter 1 (Kempuraj et al 2024b). Receptor-mediated transcytosis pathways, including low-density lipoprotein receptor–related protein 1 and the transferrin receptor, provide additional yet highly selective routes for transport (Alahmari 2021). Aquaporin in the BBB also plays a crucial role in regulating water and solute balance, preventing edema and other harmful conditions. Together, these mechanisms limit excessive activation of peripheral immunologic signals within the CNS, restrict the entry of pathogens and toxins, and maintain steady neurotransmitter levels (Wu et al 2023). Additionally, endothelial cells in vivo are constantly subjected to shear stress generated by blood flow (typically a few dynes per square centimeter), which promotes the development of TJs and overall barrier integrity (Konig et al 2025). Many recent microfluidic BBB-on-a-chip technologies now incorporate this, enabling them to more accurately recapitulate in vivo BBB behavior and providing a foundation for downstream disease modeling and therapeutic evaluation (da Silva et al 2025).
Molecular Mechanisms of BBB Disruption in Neurodegeneration
Neurodegenerative diseases are increasingly recognized to involve significant dysfunction of the BBB, driven in part by chronic neuroinflammatory processes (Figure 3). In AD and PD, pathologic protein aggregates, activated glial cells, and circulating immune mediators converge on the cerebral endothelium, leading to progressive barrier impairment (Hussain et al 2021; Yuan et al 2023). In PD, α-synuclein oligomers stimulate leukocyte recruitment and paracellular leakage across the BBB by activating endothelial cells and upregulating adhesion molecules, including intercellular adhesion molecule 1 and vascular cell adhesion molecule 1 (Figure 3; Bogale et al 2021; Lau et al 2024). Persistent microglial activation exacerbates endothelial dysfunction and disrupts TJ integrity through the release of inflammatory mediators such as interleukin 1β (IL-1β), tumor necrosis factor α, IL-6, and matrix metalloproteinases (Hussain et al 2021; Yuan et al 2023). Such neurovascular changes also play a crucial role in the neurodegeneration in PD (Al-Bachari et al 2017). Together, these mechanisms affect synaptic stability, energy metabolism, and brain perfusion, underscoring the need for human-relevant BBB models to analyze how alterations in barrier permeability contribute to neurodegenerative diseases. Similar mechanisms operate in AD, where beta-amyloid (Aβ) accumulation, oxidative stress, and chronic cytokine exposure impair endothelial transport function and downregulate TJ and adherens junction proteins. BBB disruption in AD is associated with reduced clearance of neurotoxic peptides, altered cerebral perfusion, and increased infiltration of peripheral immune cells, amplifying neuroinflammation and neuronal injury (Hussain et al 2021; Yuan et al 2023). Nitric oxide released from glial cells can cause neuroinflammation, neurodegeneration, increased BBB permeability, and edema (Thiel and Audus 2001; Doherty 2011). Beyond inflammatory signaling, additional contributors to BBB dysfunction in neurodegeneration include mitochondrial dysfunction within endothelial cells, altered shear stress responses, dysregulated vesicular transport, and age-associated changes in pericyte coverage of small blood vessels (Hussain et al 2021; Yuan et al 2023). These multifactorial mechanisms highlight the need for experimental platforms capable of dissecting dynamic, cell-specific contributions to BBB breakdown in humans.
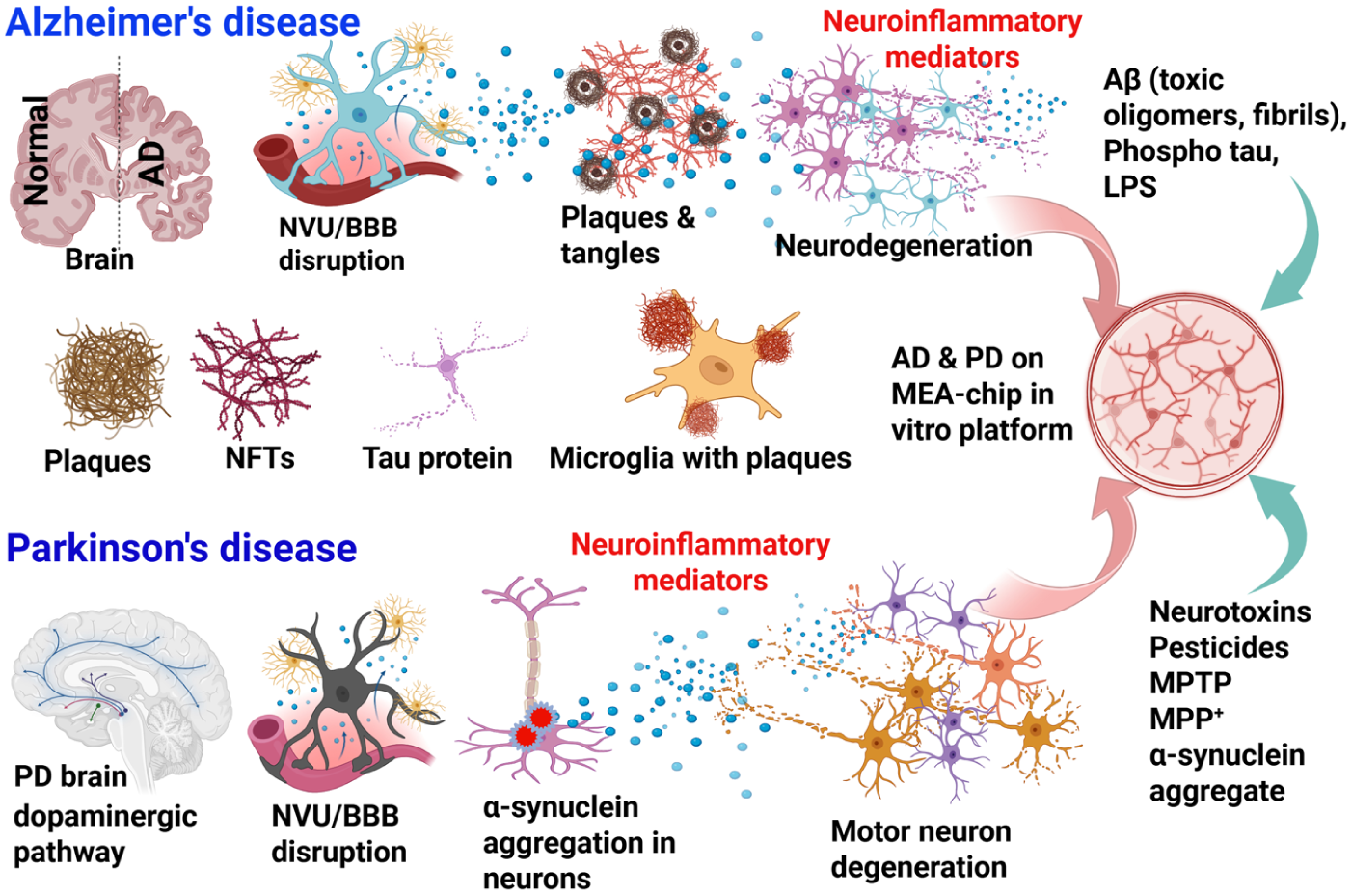
Brain-on-a-chip in vitro platform modeling for AD and PD. This in vitro platform may incorporate neurons, astrocytes, pericytes, endothelial cells, microglia, T cells, growth factors, cytokines, chemokines, inflammatory mediators, and neurotoxic mediators specific to AD or PD. AD models can be created using iPSC-derived neurons, endothelial cells, pericytes, and microglia from patients with AD and grown on standard tissue culture plates, microfluidic MEA plates, or compartmentalized MEA chips. The AD chips may be incubated with AD-relevant stimuli such as Aβ peptide, phosphorylated tau or LPS to induce AD-relevant responses in the brain-on-a-chip. Similarly, a PD-specific brain-on-a-chip can be created from iPSC-derived brain cells, including midbrain dopaminergic neurons from patients with PD. PD-relevant stimuli, including MPP+, α-synuclein, pesticides, and other neurotoxicants, can be used to incubate the cells to study the pathogenesis of PD in vitro. Aβ, beta-amyloid; AD, Alzheimer disease; iPSC, induced pluripotent stem cell; LPS, lipopolysaccharide; MEA, microelectrode array/multielectrode array; MPP+, 1-methyl-4-phenylpyridinum; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; NFTs, neurofibrillary tangles; PD, Parkinson disease.
Brain-on-a-Chip and BBB-on-a-Chip
Brain-on-a-chip platforms are microphysiologic systems composed of cultured brain cell types in microfabricated devices under regulated perfusion, shear stress, and biochemical gradients. These systems may incorporate neurons, astrocytes, pericytes, endothelial cells, microglia, T cells, growth factors, cytokines, chemokines, inflammatory mediators, and neurotoxic mediators (Figures 1 and 3). By supporting 2- and 3-dimensional (3D) configurations and enabling spatial organization of multiple cell types, brain-on-a-chip systems support long-term, physiologically relevant cocultures and model specific brain regions by modifying channel geometry, matrix composition, and flow profiles (Akcay and Luttge 2023). Different hydrogels are used as a matrix material or as a thin coating on channels or membranes for cell adherence in BBB-on-a-chip platforms on the brain side and the blood side (Vetter et al 2025). Hydrogels such as Matrigel, gelatin, collagen, fibrin, and hyaluronic acid are commonly used in BBB-on-a-chip platforms (Motallebnejad et al 2019; Vetter et al 2025). BBB-on-a-chip platforms can be designed into sandwich, parallel, 3D, or self-assembly with vasculogenesis models based on the structural design (Vetter et al 2025). BBB models are also created using transwell inserts with various brain cell types (Stone et al 2019). Although the transwell system is simple to set up, it has some disadvantages, including a lack of sufficient cell-cell interactions, shear stress, and low barrier integrity. Controlled deposition of various brain cells in different plating chambers is reported to provide a structural platform for studying human neurodegenerative diseases (Maisonneuve et al 2022).
By combining human iPSC (hiPSC)–generated neurons and glia with microvascular endothelial cells and pericytes, disease-focused brain-on-a-chip models extend these capabilities, recapitulate important components of the NVU (Pediaditakis et al 2022), and allow exposure to cues relevant to the disease, such as Aβ, α-synuclein, inflammatory cytokines, or hypoxic stress (Figure 3; Amartumur et al 2024; Rodrigues et al 2024). To monitor network activity, barrier dynamics, and released biomarkers in real time during medication schedules or simulated disease progression, many of these chips have embedded microelectrodes, clear optical access, and biochemical sensors (Zhao et al 2023; Spitz et al 2024). The brain-on-a-chip models can be extended across multiple organs to study the role of “axes” such as the microglia-gut-brain axis in neurodegenerative diseases, including PD (Raimondi et al 2019). These features transform brain-on-a-chip platforms into human-based, mechanism-driven models that bridge simplified in vitro systems and animal models (Zhao et al 2023; Amartumur et al 2024).
One study demonstrated an improved microfluidic device that supports electrical modulation of neural circuits and their physiologic property measurement (Han et al 2023). This is valuable for understanding synaptic dysfunction, neural circuits, and their role in several neurologic diseases, such as autism spectrum disorder and Huntington disease (Han et al 2023). The BBB prevents the delivery of drugs into the brain, which can limit their therapeutic potential in the treatment of neurologic disorders. Focused ultrasound has been reported to disrupt the BBB temporarily and allow the entry of therapeutic agents into the brain. Microbubbles, when combined with focused ultrasound, can pass through the BBB into the brain. Therefore, a combination of focused ultrasound and drug-encapsulated microbubbles can be used for drug delivery for neurodegenerative diseases, including AD (Rouhi et al 2024).
Whole-Brain Organoid/Multiregion Brain Organoid and Vascularized Organoids
Brain organoids are in vitro 3D culture models that are self-organizing, 3D, and a miniaturized in vitro platform generated from hiPSCs (Kistemaker et al 2025; Figures 1 and 4). There are several types of brain organoids, such as cerebral, midbrain, hindbrain, cortical, vascularized, multiregion, and whole brain organoids/multiregion brain organoid (MRBO; Kshirsagar et al 2025). MRBOs show the early stages of human brain development, are not conscious, are smaller than the actual human brain, and contain about 6 to 7 million neurons as compared with approximately 100 billion neurons in the adult human brain (Goriely 2025; Kshirsagar et al 2025). These MRBOs are the next-generation brain organoids and exhibit neuronal activity and features of BBB activity (Kshirsagar et al 2025). This is one of the first MRBOs to contain cells from various brain regions, connected and acting together, with characteristics of fetal brain tissue (Kshirsagar et al 2025). Most brain organoids replicate brain regions but not the whole brain. The whole brain organoids mimic neurologic diseases such as AD, schizophrenia, and autism spectrum disorder, which affect the whole brain, not just a part of the brain. The brain organoids are used to investigate the pathogenesis of neuroimmune and neurologic diseases such as AD, PD, and autism spectrum disorder and drug discovery (Ji et al 2025; Kim et al 2025; Li and Knoblich 2025). Due to the complex nature of neurodegenerative disorders, animal models and single-cell cultures have proven inadequate to study the pathogenesis. This void has recently been filled by the use of organoids from hiPSCs derived from healthy controls and patients (Hong et al 2023). Although vascularized brain organoids are already available to some extent, they lack vascular structures, blood flow, and a functional BBB complex (Kistemaker et al 2025; Song et al 2026). Vascularized brain organoids using normal hiPSCs and hiPSCs overexpressing the endothelial transcription factor ETS variant 2 were recently developed to study the effect of HIV and neurotropic pathogens in the brain (Selvam et al 2026). The study concluded that future research is needed to further characterize lumen formation, perfusibility, and dynamic interactions with neural and glial components. In another study, a human pluripotent stem cell–derived vascularized neuroimmune organoid model containing neurons, microglia, astrocytes, and blood vessels was generated, and organoid neuronal activities were analyzed by the microelectrode array system (AXION Biosystems). This study demonstrated that brain extracts from patients with AD can induce AD-like pathologies in organoids, such as plaque-like Aβ aggregates, tau tangle–like aggregates, and neuroinflammation (Ji et al 2025). Recent developments have focused on improving brain organoids by enhancing their regional specificity and reproducibility. Region-specific brain organoids can produce highly uniform neurons and progenitor cells through the regulation of developmental signaling pathways. Human microphysiological systems, such as organ chips, are also emerging rapidly. Region-specific brain spheroids have the advantage of creating and modeling individual brain regions for therapeutic studies, since different brain regions can respond differentially to therapeutics in neurologic diseases (Strong et al 2023). Mast cells are implicated in various neuroimmune and neurodegenerative diseases (Theoharides et al 2016; Kempuraj et al 2018; Theoharides et al 2023). The addition of iPSC-derived mast cells to the brain organoid can be useful to investigate neuroimmune and neuroinflammatory disorders, as shown previously (Igarashi et al 2018; Saglam-Metiner et al 2024).
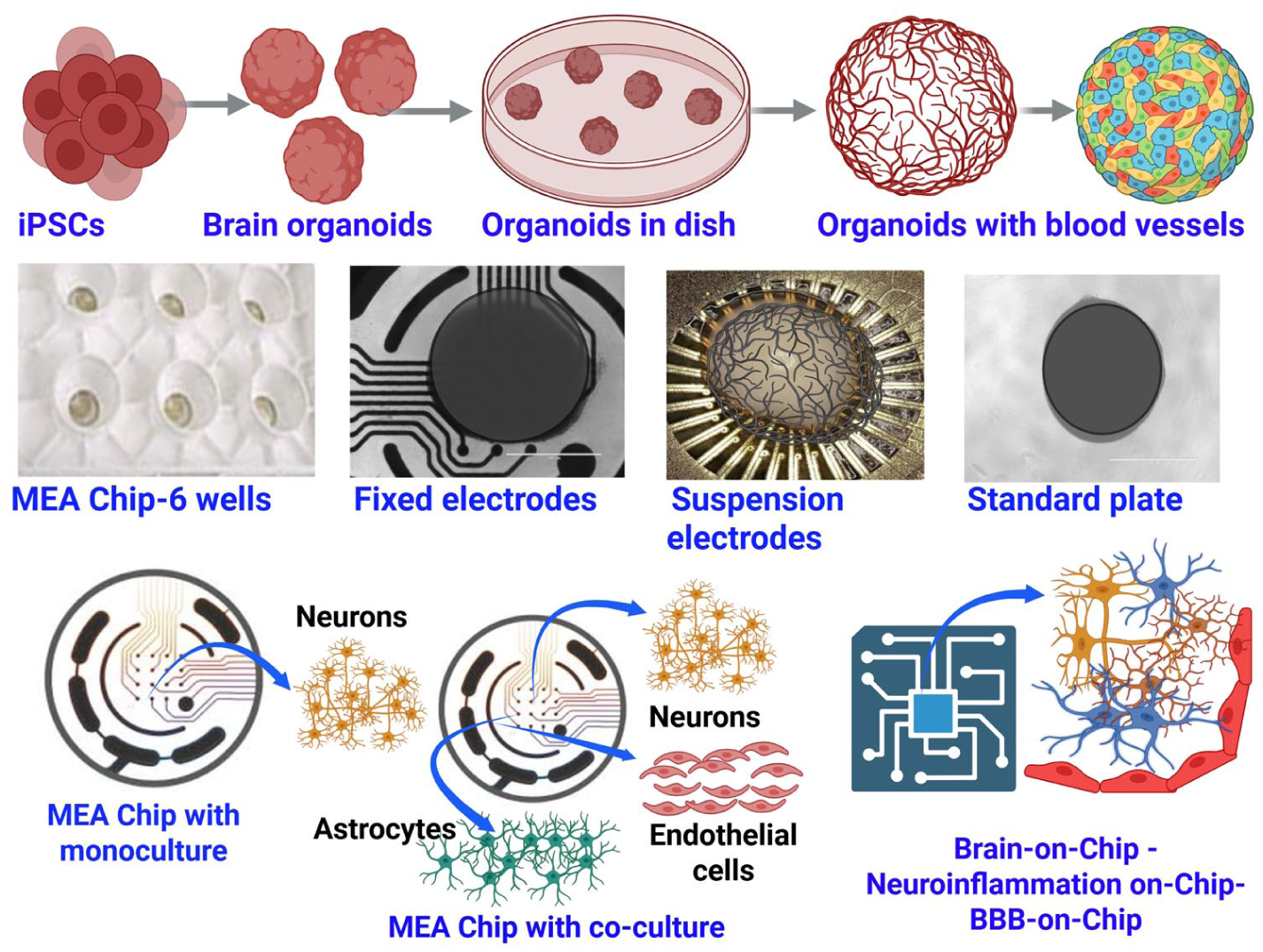
Schematic diagram illustrating a brain-on-a-chip model on the MEA platform. Various brain organoids/brain-on-a-chip models can be grown using iPSC-derived brain cells, such as neurons, astrocytes, and pericytes. These organoids can be brain region specific, such as cortical and midbrain organoids, with different cell compositions. Vascular-like structures can also be incorporated in brain organoids using brain-specific endothelial-like cells. These 3-dimensional brain organoids/brain-on-a-chip models can be grown on standard plates or MEA plates with either fixed or suspension electrodes. Suspension electrodes are a newer approach and may offer several advantages over fixed electrodes. Neuronal signals are picked up by the electrodes and read by special software to quantify neuronal activity. In addition, disease- or region-specific brain cells (2- or 3-dimensional) can be grown in coculture configurations to assess cell type–specific contributions to neuroinflammation and neurodegeneration. BBB, blood-brain barrier; iPSC, induced pluripotent stem cell; MEA, microelectrode array/multielectrode array.
One study developed a model by utilizing human primary astrocytes and pericytes, as well as iPSC-derived cortical neurons, microglia, and brain microvascular endothelial-like cells, to mimic human cortical parenchyma and BBB (Chim et al 2024). In addition, the development of the assembloid technique (a combination of cell types, organoids, and spheroids) has allowed for the study of interactions among these brain regions. Engineering technologies such as bioreactors and bioprinting have aided in increasing the reproducibility of brain organoids by mitigating variability and improving culture techniques (Sun and Pan 2025). Human brain organoids offer a more human-relevant platform for neurotoxicity assessment and can effectively recapitulate chronic, low-dose, and mixture environmental risks; moreover, coupling brain organ-on-a-chip with artificial intelligence (AI) could enhance next-generation risk assessment strategies (J Liu et al 2026). AI has recently been increasingly useful for disease diagnosis and report generation (Ramaswamy 2026). NeuroAI integrates neuroscience and AI tools to analyze neural data and understand brain function under disease conditions (Cohen et al 2022). Integration of NeuroAI with BBB-on-a-chip, brain-on-a-chip, and brain organoids modeling will advance neuroscience discovery (Sadeh and Clopath 2025).
The technological method of 3D bioprinting is a novel approach for creating brain organoids, which more accurately recapitulate in vivo processes (Juraski et al 2023; Fang et al 2026). However, limitations such as a lack of high-fidelity cell types, insufficient maturation, and significant heterogeneity reduce the reliability and applications (Fang et al 2026). Genetic engineering through CRISPR (clustered regularly interspaced short palindromic repeats) has allowed brain organoids to be utilized in investigating neurologic diseases under an environment that resembles human tissue (Fang et al 2026). Brain organoids are also being explored for their potential to mitigate the need for extensive clinical trials during drug screening (Aili et al 2024). Regenerative medicine is another field that is predicted to greatly benefit from advances in brain organoids (Hongxi et al 2025). Visual function has been demonstrated to be restored in rats through transplantation of human forebrain organoids (Jgamadze et al 2023). Last, researchers are exploring the creation of synthetic biological intelligence through the integration of AI with brain organoids/organoid intelligence/MRBO intelligence (Smirnova et al 2023). While these approaches may advance our understanding of human cognition, they raise some ethical considerations, such as defining where sentience begins and the relationship between stem cell donors and organoid intelligence systems.
Brain-on-a-chip platforms broadly model neural tissue organization and function, but accurate modeling of the BBB within these systems presents additional challenges due to the need for specialized vascular architecture and highly regulated transport properties (Hajal et al 2022). Traditional in vitro BBB models, including endothelial monolayers cultured on plastic substrates, transwell coculture systems, and 3D brain organoids, have been widely used to study basic barrier function and molecular permeability. However, these approaches fail to reproduce critical in vivo features of the BBB, such as cylindrical microvessel geometry, physiologic shear stress, direct interactions with pericytes and astrocytic end feet, and mature junctional protein complex organization. As a result, their predictive power for human BBB integrity and drug transport is limited, although they remain useful for preliminary screening. To address these limitations, microfluidic BBB-on-a-chip platforms have been developed that more closely recapitulate NVU structure and function. These systems organize human primary cells or iPSC-derived brain endothelial cells into perfusable small vessels and position astrocytes and pericytes abluminally in adjacent 3D matrix compartments. Since these systems generate physiologic shear stress under continuous flow and enable direct measurements of permeability and transendothelial electrical resistance (TEER), they offer a more quantitative and physiologically relevant assessment of barrier integrity and transporter function than static models (Hajal et al 2022; Kawakita et al 2022).
Disease-focused BBB-on-a-chip models extend these platforms by using chemokine gradients or proinflammatory cytokines to induce endothelial activation, TJ disruption, and T-cell migration across the barrier (Nair et al 2023; Ohbuchi et al 2024). In a subset of advanced brain- and BBB-on-a-chip platforms, functional sensing modalities have been incorporated to assess downstream neural consequences of barrier disruption. Notably, the integration of microelectrode arrays enables real-time monitoring of neuronal and network-level electrical activity in response to changes in BBB integrity, inflammatory signaling, or therapeutic compound transport (Hajal et al 2022; Kawakita et al 2022). Collectively, these features establish BBB-on-a-chip platforms as powerful experimental tools for studying BBB integrity, immune interactions, and neurovascular dysfunction in humans (Figures 4 and 5).
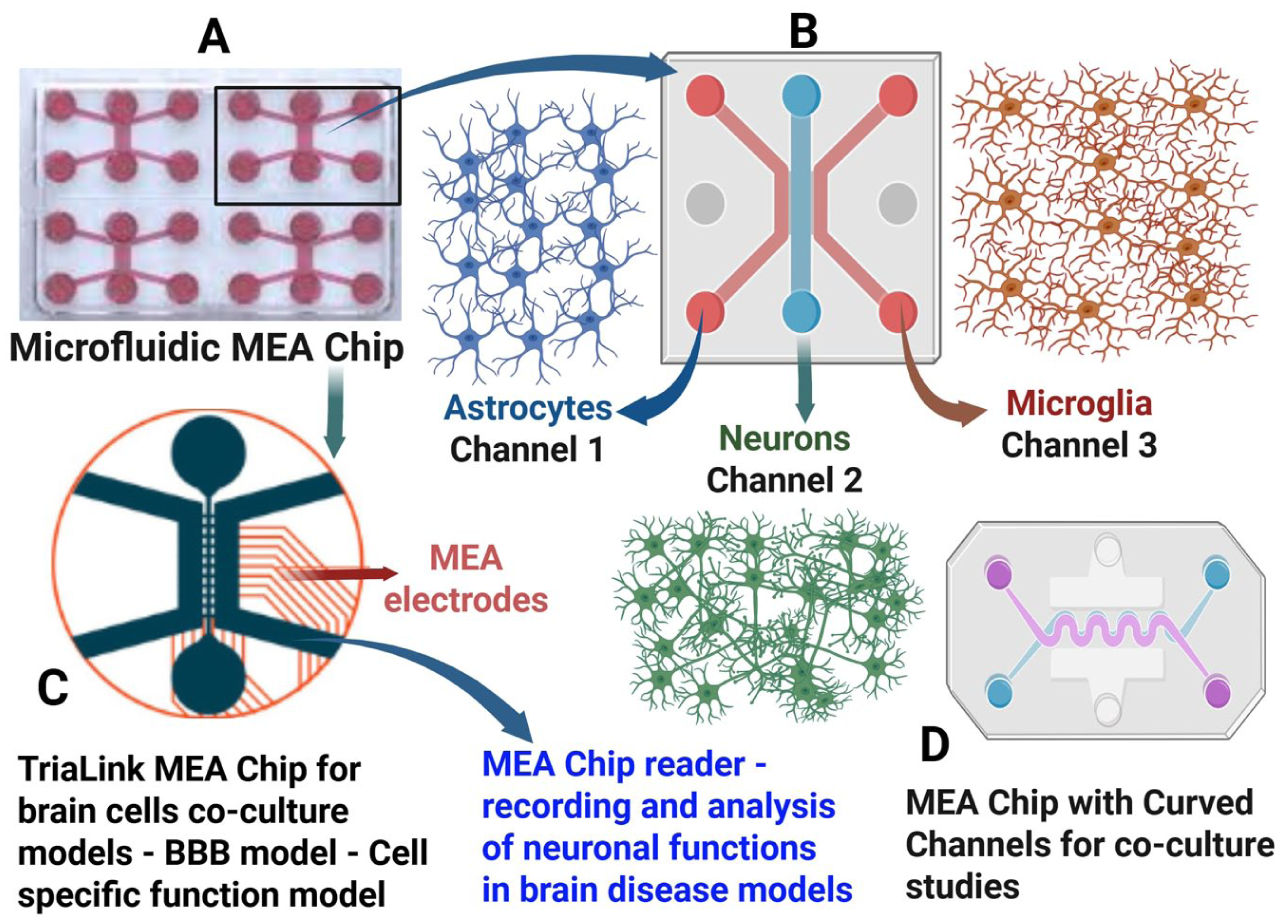
Schematic diagram illustrating the brain-on-a-chip model on the compartmentalized MEA platform. This platform utilizes a high-throughput technology with multiple compartments (channels) to culture distinct cell types with fluidic isolation and controlled connectivity via microchannels. The platform can be used to study Alzheimer disease, Parkinson disease, and other neurologic diseases, including disease pathogenesis and treatment evaluation, and it supports marker-based and functional characterization of the brain-on-a-chip models. (A) TriaLink MEA with 3 channels in each chip, in which the middle channel is connected to 2 side channels. (B) Sample triculture with astrocytes (channel 1), neurons (channel 2), and microglia (channel 3). (C) TriaLink MEA for brain cell coculture with surface fixed electrodes. (D) Curved channels for compartmentalized culture. BBB, blood-brain barrier; MEA, microelectrode array/multielectrode array.
A recent study using an organ-on-a-chip model of BBB evaluated the effects of clinical radiation on BBB permeability and inflammatory cytokine release (Yock et al 2026). This BBB model was created with human endothelial cells, astrocytes, and pericytes and then irradiated with 0, 2, or 20 Gy with a 6-MV x-ray beam from a medical linear accelerator. At 0, 6, 24, and 48 h postirradiation, barrier permeability was evaluated. The study reported a dose- and time-dependent increase in BBB permeability with increased levels of vascular endothelial growth factor and IL-1β.
Patients with long COVID develop chronic neuropsychiatric symptoms, but the precise pathogenesis is not clearly known. Long COVID causes inflammation and neuroinflammation, affects the NVU/BBB, and contributes to neurologic symptoms (Paidas et al 2022; Kempuraj et al 2024a). Microfluidic BBB-on-a-chip, brain-on-a-chip, and brain organoid models can be developed to understand the disease pathogenesis and for evaluation of therapeutic agents for SARS-CoV-2 infection/long COVID in association with neurologic diseases (Erickson et al 2023).
BBB-on-a-Chip Reveals Barrier Disruption, Endothelial Inflammation, and Immune Cell Trafficking
Neuroinflammation-driven BBB dysfunction is one of the most extensively modeled pathologic processes using BBB-on-a-chip platforms, owing to its central role in immune cell trafficking and early neurodegenerative disease progression. A representative membrane-free BBB-on-a-chip model demonstrates how inflammatory cues directly alter endothelial barrier integrity and promote immune cell infiltration. The OrganoPlate 3 lane culture model includes 40 microfluidic chips, each composed of a top, middle, and bottom lane (Nair et al 2023). This model supports dynamic culture conditions as it can generate shear stress that mimics BBB physiology and functions under bidirectional perfusion. The model was also measured with TEER, sodium fluorescein assays, and immunocytochemistry assays to confirm the presence of TJ integrity before the induction of disease-mimicking conditions. To stimulate neuroinflammation, tumor necrosis factor α and IL-1β were used, resulting in reduced barrier integrity and disruption of TJ organization. This inflammatory stimulation also upregulated intercellular adhesion molecule 1 and vascular cell adhesion molecule 1. Treatment with natalizumab reduced T-cell adhesion and showed decreased transmigration, highlighting the utility of the BBB-on-a-chip for evaluating neuroinflammatory mechanisms and testing antineuroinflammatory therapeutics. While the system offers a real-time and 3D model for studying BBB inflammation and immune trafficking, its use of endothelial cells alone and lack of patient-specific or stem cell–derived components currently limit its ability to capture the full cellular complexity of the human NVU (Roheger et al 2018).
BBB-on-a-Chip Models of AD- and PD-Associated Barrier Dysfunction
BBB dysfunction is an early and mechanistically important feature of AD, making it a key target for BBB-on-a-chip modeling of disease-associated vascular pathology (Takeuchi et al 2025). To study the role of the cerebral vasculature in AD, a physiologically relevant 3D human neural cell culture microfluidic model was developed incorporating a brain endothelial cell monolayer with a BBB-like phenotype (Shin et al 2019). This model recapitulated several key aspects of BBB dysfunction observed in patients with AD, including increased BBB permeability and altered junctional protein expression. Specifically, the model demonstrated reduced expression of TJ and adherens junction such as claudin 5 and vascular endothelial–cadherin, as well as increased expression of matrix metalloproteinase 2 and reactive oxygen species and deposition of Aβ peptides at the vascular endothelium. Thus, it provides a well-controlled platform for investigating BBB functions and for therapeutic strategies aimed at preserving BBB integrity. Brain-on-a-chip for AD can be created by using iPSC-derived neurons, endothelial cells, pericytes, and microglia from patients with AD and grown on a normal tissue culture plate, microfluidic microelectrode array plate, or compartmentalized microelectrode array chip (Figures 3-5). Similarly, brain-on-a-chip can be developed with iPSC-derived dopaminergic neurons, astrocytes, and pericytes and incubated with PD-associated neurotoxins (Figures 3-5).
The dopaminergic neuronal loss in the substantia nigra is a major hallmark of PD (Cramb et al 2023; Thangavel et al 2024). Monoclonal antibodies have shown rather disappointing results in targeting α-synuclein aggregates in PD (Messak et al 2026). Abnormal immune activation, BBB disruption, and astrocyte dysfunction have become of particular interest understanding the pathogenesis of PD. In one study, investigators utilized an iPSC-derived BBB-chip model consisting of brain microvascular endothelial cells, pericytes, and astrocytes to provide further evidence for the altered astrocyte role in BBB impairment (de Rus Jacquet et al 2023). This reinforces previous studies detailing astrocyte dysfunction in PD and suggests the BBB-on-a-chip’s potential as a vital tool in the study of PD pathogenesis and vascular changes in the substantia nigra. Another study utilized a specific model based on substantia nigra on a chip, which constructed the NVU with brain endothelial cells lining the vascular channel, introducing α-synuclein fibrils to demonstrate PD and BBB disruption (Pediaditakis et al 2021).
NVU-on-a-Chip Models of Ischemic Stroke and BBB Disruption
In ischemic stroke, impaired cerebral blood flow leads to hypoxia and energy failure, which disrupts BBB integrity, and the resulting barrier breakdown exacerbates neuronal injury by allowing neurotoxic substances and immune cells to bypass the barrier (Wevers et al 2021). To better understand these mechanisms, researchers developed a human NVU-on-a-chip model that consists of a parallel culture of 40 chips, each containing a perfused vessel of primary human brain endothelial cells cultured with iPSC-derived astrocytes and neurons in a 3D culture under flow. Under stroke-mimicking conditions, including hypoxia, hypoglycemia, and halted perfusion, the model exhibited pronounced BBB disruption, reduced mitochondrial membrane potential, and decreased ATP levels, closely mimicking in vivo ischemic pathology. Collectively, these findings highlight the utility of NVU-on-a-chip platforms for dissecting mechanisms of acute BBB failure and neurovascular dysfunction in stroke. However, limitations remain, such as the absence of pericytes and patient-derived endothelial cells used in the model, suggesting that future directions could focus on further increasing cellular completeness and personalization to more fully capture human stroke heterogeneity.
A study reported a microfluidic multisize spheroid array capable of culturing 3D multicellular spheroids with high reproducibility in medium- to high-throughput formats using a range of tissue types (Kawakita et al 2022). Through this platform, a BBB model was created with a triple-culture spheroid system consisting of human brain endothelial cells, pericytes, and astrocytes, thereby allowing cell-to-cell interactions that enhance BBB integrity. Barrier integrity was shown to depend on spheroid size and cell composition, indicating the utility of this BBB model for studying BBB integrity, compound penetration, and drug screening (Eilenberger et al 2021; Kawakita et al 2022).
Another study developed a microengineered physiologic system that models key structures of the BBB and enables 3D mapping of nanoparticle distributions in vascular and perivascular regions (Ahn et al 2020). The model demonstrated reduced reactive gliosis of 3D cultured astrocytes and polarized aquaporin 4 distribution, closely replicating BBB water transport and homeostatic conditions. Engineered high-density lipoprotein mimetic nanoparticles demonstrated successful BBB transport, supporting the model’s valuable application to quantify and analyze CNS drug delivery through an active 3D astrocytic network and aquaporin 4 transport mechanisms.
A recent study provided empirical evidence supporting the reliability and efficiency of BBB-on-a-chip technology for identifying functional shuttle moieties as compared with traditional transwell models that lack physiologic shear stress (Choi et al 2024). With phage display peptide biopanning across both models with mouse endothelial cells, pericytes, and astrocytes, the BBB-on-a-chip model showed greater reliability for shuttling peptides by closely emulating brain endothelial glycocalyx. Further studies may involve a more comprehensive comparison between organ-on-a-chip technology and in vivo animal models, thereby supporting the use of BBB-on-a-chip systems for vascular targeted drug delivery.
A recent model successfully applied stroke-mimicking conditions to a high-throughput tetraculture BBB model with parallel chips that utilize human microvascular endothelial cells, astrocytes, microglia, and pericytes (Wu et al 2025). This study, using microfluidic organ-on-a-chip technology, reported cell damage characteristics, BBB disruption, oxidative stress, and inflammatory responses following ischemia and demonstrated its utility as a drug screening model for ischemia-related injury. With advanced data analysis and machine learning, these high-throughput chips can be leveraged to rapidly and efficiently increase understanding of ischemic stroke pathophysiology. An integrated BBB chip was combined with transcriptomic profiling and coexpression network analysis to identify core molecular pathways and enable computational drug screening for ischemic stroke (J Liu et al 2025). This study provided a proof-of-concept framework for the integration of organ-on-chip technology and machine learning. In summary, further work is needed to validate complex models and patient-derived modeling approaches.
iPSC-Derived BBB-on-a-Chip Models for Patient-Specific Disease Modeling
The BBB-on-a-chip most commonly employs iPSC-derived brain microvascular endothelial cells to re-create a functional NVU that closely resembles the in vivo BBB. These models are built in microfluidic devices that allow 3D culture and fluid flow, which can mimic the physiologic environment of the BBB. Advanced features such as TEER measurements and assays including fluorescence and immunocytochemistry are used to test barrier tightness and drug permeability (Alves et al 2023). Some studies have also utilized patient-derived iPSC lines to model specific diseases or genetic conditions, advancing the potential for more personalized treatment approaches (Vatine et al 2019). Notably, recent research demonstrates that these in vitro BBB chips can replicate key barrier properties and have been applied to studying drug transport, stroke, and immune responses. Brain- or BBB-on-a-chip can be created using various iPSC-derived brain cells from the same subject or patient for more specificity. However, the BBB-on-a-chip models still face limitations and challenges that must be addressed before they can be used most effectively. Limitations include a lack of standardized protocols (Alves et al 2023). Future models would benefit from embedded sensors to monitor neuronal responses to drug administration and BBB penetration. Such developments would drastically increase the accuracy, efficiency, and speed of CNS drug development.
BBB-on-a-Chip Platforms for Nanoparticle Transport and CNS Drug Delivery
BBB-on-a-chip platforms provide a physiologically relevant framework for evaluating CNS drug delivery strategies by enabling direct assessment of therapeutic transport across a human-like NVU (Kantawala et al 2024). BBB-penetrating nanotechnology-based drug delivery systems, such as polymeric nanoparticles, liposomes, inorganic nanomaterials, and biomimetic carriers, may provide brain-targeted treatment (Gao et al 2025). To evaluate the therapeutic potential and transport properties of polyoxometalate-decorated gold nanoparticles across an in vitro BBB, investigators used a BBB-on-a-chip model incorporating human brain microvascular endothelial cells, astrocytes, and pericytes (Kantawala et al 2024). This system demonstrated efficient nanoparticle penetration through the barrier and confirmed the inhibition of Aβ aggregation, a key pathologic event in AD. When compared with static in vitro assays, this model allows for direct assessment of nanoparticle trafficking and interactions with BBB components under dynamic flow conditions. Although the system provides promising insights into nanoparticle-mediated neuroprotection, it does not incorporate patient-specific hiPSC-derived cells or immune elements, limiting its ability to fully capture personalized disease responses and inflammatory dynamics. Future studies could integrate additional NVU components and biosensing technologies to expand the applicability of the platform in drug testing and mechanistic studies.
Revolutionizing Neurotherapeutics: BBB-on-a-Chip Technologies for Precise Drug Delivery
BBB dysfunction is associated with the progression of disorders such as AD and PD, in which barrier breakdown can worsen neurodegeneration and interfere with drug delivery. BBB-on-a-chip models closely mimic the human BBB’s structure and function by incorporating brain endothelial cells, astrocytes, and pericytes in either 2D or 3D coculture formats, often under flow conditions to cause shear stress. Importantly, some models utilize patient-derived iPSCs, allowing more personalized modeling (Vatine et al 2019). The use of TEER measurements also allows real-time assessment of barrier integrity and drug permeability. In addition, key findings from recent studies show that BBB-on-a-chip models can accurately mimic disease conditions and facilitate high-throughput screening of neurotherapeutic compounds (Kantawala et al 2024). Applications of BBB-on-a-chip models extend to the evaluation of nanoparticle transport and the investigation of neuroinflammatory processes. However, significant challenges remain, including difficulties in fully replicating the BBB’s intricate structure and physiologic complexity within microfluidic chips. Additionally, commonly used materials such as polydimethylsiloxane are incompatible with hydrophobic drugs, which can affect the accuracy of drug delivery studies.
Innovations in targeted brain delivery therapeutics have been explored through the utilization of folic acid–conjugated brush polymers that target reduced folate carriers expressed on brain endothelial cells (Mazrad et al 2024). This study utilized the in vitro BBB chip model to quantitatively test the efficacy of this approach as a potential tool in delivering cargo across the BBB.
Sensor-Integrated BBB-on-a-Chip Platforms for Real-time Barrier Assessment
This BBB-on-a-chip model incorporates human brain endothelial cells, astrocytes, and pericytes, organized in a compartmentalized 3D microfluidic device. The astrocytes and pericytes are embedded in a fibrin hydrogel, and endothelial cells were added in one of the lateral channels. This configuration enables direct cell-to-cell and cell-to-matrix interactions that are essential for physiologic BBB functions.
The primary technological advancement of this model is the integration of a micro-TEER measurement close to the barrier, allowing precise and real-time monitoring of barrier integrity without obstructing optical imaging. TEER values increased as TJs matured, paralleling in vivo conditions. The model was validated by demonstrating the permeability and functionality of the TJ. This model was successfully used to test drug permeability, specifically evaluating the ability of a multifunctionalized gold nanorod (GNR-PEG-Ang2/D1) nanotherapeutic to cross the BBB. Limitations of this system include the lack of patient-specific hiPSCs, which may limit personalized disease modeling, as well as the absence of blood flow simulation (Palma-Florez et al 2023). Thus, future studies should incorporate additional brain cell types and model immune cell migration or inflammatory responses to investigate broader disease mechanisms (Wevers et al 2021).
Isogenic BBB Model
Another key advantage of using hiPSCs lies in the ability to create isogenic systems, where the various brain cells are sourced from the same subject background or donor. This allows researchers to model certain genetic conditions or diseases with high specificity. An isogenic BBB system could consist of iPSC-derived brain microvascular endothelial cells, astrocytes, and pericytes from the same donor background (healthy subjects or patients; Figure 1). Isogenic BBB kit components are available commercially to model the BBB. One recent study developed an in vitro translational NVU model that recapitulates human pathology more accurately than previous models, which could be useful for new therapies (Shen et al 2026). The authors developed the NVU model using iPSC-derived brain cells (neurons, microglia, astrocytes, pericytes, brain-like endothelial cells) from patients with AD and compared them with iPSC-derived brain cells from control subjects. They reported that AD brain chips had decreased claudin 5 and ZO-1 and increased paracellular permeability, Aβ-dependent NVU dysfunction, neuroinflammation, and tau accumulation. Another study explored an isogenic hiPSC-derived BBB-on-a-chip platform consisting of a brain endothelial cell monolayer on top of a 3D hydrogel astrocyte culture (Motallebnejad et al 2019). An isogenic NVU model can be created from the same control- or patient-derived iPSC line (Canfield et al 2017) and can include isogenic coculture of astrocytes, brain capillary endothelial-like cells, microglia-like cells, neuronal stem cells, and pericytes (Haferkamp et al 2023). Impedance spectroscopy was used to determine TEER and membrane capacitance, with results consistent with reported TEER values from traditional transwell cultures. Transforming growth factor β1 was added to the chip and shown to disrupt the BBB, as evident through stress fibers in the treated samples, decreased junctional localization and TEER, and activation of astrocytes (Motallebnejad et al 2019). These results indicate the potential use of the model for screening of specific brain cell activation, mechanistic studies, and drug screening. Future directions will leverage isogenic models for personalized medicine applications, including patient-specific BBB-on-a-chip platforms for various neurologic diseases (Motallebnejad et al 2019; Table 1).
Recent Brain- and BBB-on-a-Chip Models for Brain Diseases.
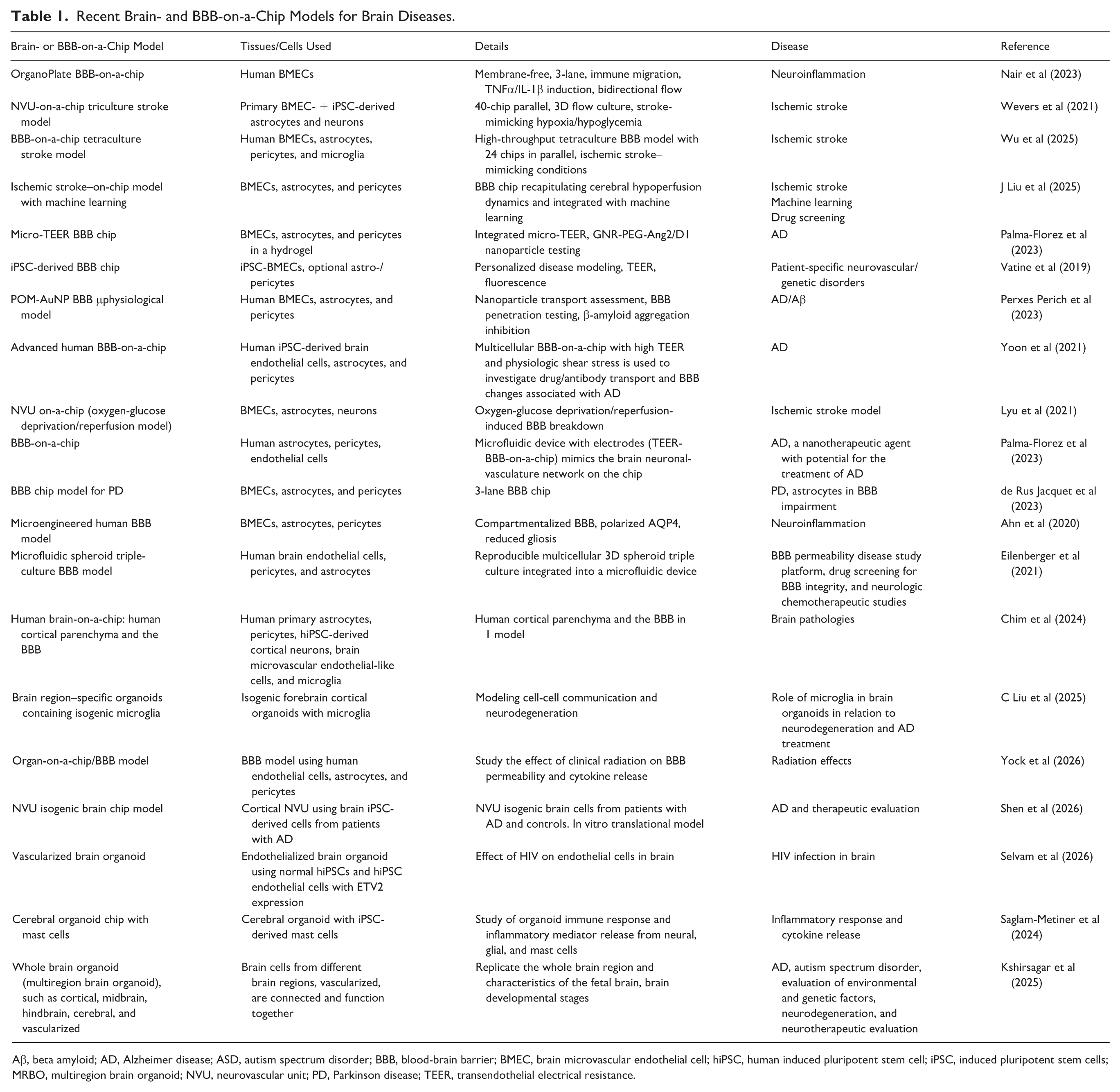
Aβ, beta amyloid; AD, Alzheimer disease; ASD, autism spectrum disorder; BBB, blood-brain barrier; BMEC, brain microvascular endothelial cell; hiPSC, human induced pluripotent stem cell; iPSC, induced pluripotent stem cells; MRBO, multiregion brain organoid; NVU, neurovascular unit; PD, Parkinson disease; TEER, transendothelial electrical resistance.
Limitations of Brain-on-a-Chip and BBB Models
Even with substantial advancements, BBB-on-a-chip devices still have significant drawbacks that constrain their clinical translation potential. Many models oversimplify the cellular architecture of the BBB, frequently excluding essential contributors such as microglia, oligodendrocytes, perivascular macrophages, and circulating immune and blood-borne components, thereby restricting the ability to model neuroinflammatory signaling, immune-endothelial interactions, and disease-specific BBB dysfunction in neurodegenerative disorders (Musafargani et al 2020; Vetter et al 2025). Animal models of the BBB show poor translational success and differ from human neuropathology. Current brain- and BBB-on-a-chip platforms lack chronic simulation conditions and the incorporation of different systemic factors and pathogens implicated in brain disorders. Moreover, the widespread reliance on polydimethylsiloxane layers used to model BBB and 3D brain parenchyma to mimic natural extracellular matrix introduces nontrivial experimental artifacts, as hydrophobic small molecules and biologics can be variably absorbed into the device material, leading to unpredictable alterations in effective drug concentrations, confounding pharmacokinetic and efficacy assessments (Musafargani et al 2020; Mir et al 2022). Polydimethylsiloxane devices are used to create a vasculogenesis model (Mir et al 2022). Hydrogels used in the brain- and BBB-on-a-chip fabrication have some disadvantages, including differences in degradation, durability, difficulty in integration with electrodes, and long-term toxicity. Finally, a lack of standardization across BBB-on-a-chip platforms remains a major barrier to reproducibility and cross-study comparison. Variability in cell sourcing, differentiation protocols, extracellular matrix composition, shear stress parameters, and benchmark readouts such as TEER and permeability thresholds complicates validation and limits broader adoption (Konig et al 2025; Vetter et al 2025). Addressing these limitations will be essential for advancing brain- and BBB-on-a-chip systems from exploratory research tools toward robust, predictive platforms for translational neuroscience and drug development.
Conclusions and Future Directions
There has been recent considerable advancement in the brain organoid, brain-on-a-chip, organoid-on-a-chip, and BBB-on-a-chip platforms for the study of brain development and neurologic diseases using iPSC-derived cells. Advancements in the fabrication of brain-on-a-chip devices are making it possible to better replicate the complex 3D network of the brain in vivo. However, these technologies remain in the developmental stages and require significant improvement, especially in modeling the vascular system. Although the models contain vascular structures, they lack many essential features of the brain’s complete vascular structure and function. Current organoid, brain-on-a-chip, and BBB-on-a-chip systems lack sufficient intercellular junctions, water channel proteins, and astrocyte end feet coverage. Future priorities in brain and BBB chip modeling development include expanding patient-specific or CRISPR-engineered isogenic BBB models for high-throughput drug screening, incorporating microglia and other immune components, and integrating real-time biosensors for metabolic and transcriptomic readouts. Moreover, linking BBB chips to other organ-on-chip systems is underway, such as brain region–specific organoids/MRBO incorporating synthetic biological intelligence through the integration of NeuroAI with brain organoids. Collectively, these advancements are expected to enhance the translational relevance of the brain- and BBB-on-a-chip system technologies for our understanding of neurodegenerative diseases and accelerate the development of novel CNS therapeutics for these diseases.
Footnotes
Authors’ Note
Figures were created with BioRender Software.
Author Contributions
D.K.: conceptualized, wrote the original manuscript, created the figures, and critically edited the manuscript. H.S., T.T., Da.K., A.S., P.R.R.B., M.G.B., S.P.K., B.C., C.P., P.P., D.S.V., A.S., Di.K., D.S.V.: contributed to manuscript preparation, review, and editing. T.F.: reviewed and edited the manuscript. T.C.T.: contributed to the manuscript preparation and reviewed and edited it.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: a President’s Research Grant to D.K. from Nova Southeastern University, a Faculty Research Grant to D.K. from the Dr. Kiran C. Patel College of Osteopathic Medicine, Nova Southeastern University, as well as a grant to T.C.T. from the Gateway Institute for Brain Research.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.