
Abstract
Background and Objective:
Drug-resistant (DR) mycobacterial infections present escalating threats in Iran, where facility-specific transmission dynamics and demographic disparities remain poorly characterized. This study evaluated the epidemiology of DR Mycobacterium tuberculosis and pulmonary nontuberculous mycobacterial disease over a 9-year period (2016–2024).
Materials and Methods:
This retrospective study analyzed 21,700 molecular and culture-confirmed mycobacterial samples. Species identification used line probe assays and Xpert Mycobacterium tuberculosis/rifampin. Drug susceptibility testing followed World Health Organization standards. Statistical analyses identified associations between resistance patterns, hospital sections, gender, and specimen types.
Results:
MDR-TB prevalence surged from 0.35% (2017) to 3.27% (2022), peaking during pandemic disruptions. M. simiae dominated nontuberculous mycobacteria (NTM) resistance (55.63% of resistant isolates), with significant increases in 2024 (7.91%). Airborne infection isolation rooms (AIIRs) paradoxically harbored 18.22% of Multidrug-resistant tuberculosis (MDR-TB)isolates (p < 0.0001), while pulmonary medicine units contained zoonotic M. bovis reservoirs (0.27% MDR prevalence). Male patients carried higher burdens of MDR-TB (26.01% vs. 12.45%, p = 0.005) and M. kansasii (2.05% vs. 0.53%, p = 0.012). Diagnostic challenges included 38.14% smear-negative M. abscessus and gastrointestinal NTMs (M. genavense 66.66%). Sample type analysis revealed M. fortuitum in 25.0% of abscesses (p < 0.05) and M. chelonae in 5.55% of synovial fluid (SF) specimens.
Conclusion:
Iran faces converging epidemics of MDR-TB and climate-adapted NTMs concentrated in hospital hotspots, with significant gender disparities. Precision interventions targeting AIIR protocols, water safety regulations, and gender-specific screening are urgently needed.
Introduction
Mycobacterial infections, particularly drug-resistant tuberculosis and nontuberculous mycobacteria (NTM), continue to impose a major public health burden. Meta-analyses in western Iran reported any drug resistance in 13% of new TB cases and multidrug-resistance (MDR) in 6%, with notably higher rates among retreatment groups.1,2 The World Health Organization (WHO) estimates approximately 450,000 new multidrug-resistant (MDR)-TB cases emerge annually worldwide, with mortality rates exceeding 40% among untreated patients. 3 Simultaneously, NTM infections—once considered mere environmental contaminants—now account for 15–35% of pulmonary mycobacterial diseases in high-income nations, exhibiting alarming resistance to conventional therapies. 4 This converging epidemic is fueled by diagnostic complexities, limited treatment options, and underrecognized nosocomial transmission pathways that persist despite decades of containment efforts. 5 Mycobacterium tuberculosis complex (MTBC) causes TB, an airborne, person-to-person transmissible disease with both pulmonary and extrapulmonary manifestations. In contrast, NTMs are ubiquitous environmental organisms that cause chronic lung disease primarily in predisposed hosts. TB is usually smear-positive and is treated with standardized 6-month multidrug regimens (directly observed treatment) due to its public-health impact, whereas NTM infection is diagnosed via culture/NAAT and species identification and treated with prolonged, individualized, multidrug regimens guided by species and susceptibilities. Unlike TB (which requires mandatory reporting and contact tracing), nontuberculous mycobacterial pulmonary disease (NTM-PD) is generally non‐communicable and not subject to public health notification.6,7 The existing surveillance systems are significantly hindered by notable deficiencies, especially in relation to the transmission dynamics specific to facilities and the demographic variations in resistance patterns, which results in a lack of actionable insights for clinicians aimed at targeted intervention.8,9 The escalating burden of DR manifests most severely in vulnerable populations and specific health care environments. While global attention focuses understandably on Mycobacterium tuberculosis, dangerous NTM species like M. abscessus and M. simiae increasingly demonstrate intrinsic resistance mechanisms that bypass standard therapeutic protocols.8–10 The design of hospital settings intended for containment inadvertently serves as epicenters for transmission, thereby exacerbating this challenge. Airborne infection isolation rooms (AIIRs)—specialized units engineered to prevent pathogen spread—show unexpectedly high MDR-TB prevalence according to our data, suggesting fundamental flaws in implementation protocols or environmental persistence mechanisms. 11 Equally concerning, outpatient (OP) clinics emerge as unexpected reservoirs for drug-resistant (DR) strains, implicating the community-hospital interface as a critical transmission pathway that remains overlooked in conventional infection control models. 12
Three fundamental knowledge gaps undermine effective containment strategies. First, current epidemiological studies consistently aggregate “healthcare settings” into monolithic entities, obscuring critical variations between specialized units. 13 Our analysis reveals pulmonary medicine (PM) wards and AIIRs constitute distinct transmission hotspots with unique resistance profiles—a granular understanding absent from existing literature. Second, persistent gender disparities in mycobacterial infections remain mechanistically unexplained. Males in our cohort carried over twice the burden of MDR-TB compared to females, a pattern extending to NTM species like M. kansasii. This situation probably reflects occupational exposures, health care access barriers, and diagnostic delays disproportionately affecting men—factors rarely addressed in current guidelines. Third, the rapid emergence of geographically atypical pathogens demands urgent attention. M. simiae, historically confined to specific regions, has seen a 156% increase in our 2024 samples (n = 69 in 2016 to n = 179 in 2024), showing worrying resistance patterns, particularly in OP settings. This rise highlights how quickly mycobacterial threats are evolving and spreading beyond the typical regions we usually monitor.
Existing surveillance frameworks fail to capture these complexities due to inconsistent species-level identification of NTM and the absence of mandatory reporting protocols. Also, DR-TB data seldom link resistance tendencies to specific hospital areas or demographic factors, hiding useful patterns and making it clear that facility-level epidemiological mapping is needed right once to help with targeted treatments. Our 9-year retrospective study (2016–2024) addresses these voids by integrating five critical dimensions; quantifying resistance prevalence across mycobacterial species and four resistance categories, mapping transmission hotspots within specialized hospital sections, analyzing gender-specific temporal trends, tracking association of sample type distribution with pathogen, and evaluating smear microscopy’s predictive value for resistance. This thorough method goes further than just looking at overall statistics; it focuses on pinpointing specific groups of individuals and health care settings that are most at risk and need urgent attention. As DR mycobacteria evolve beyond traditional containment paradigms, this research provides the spatial, demographic, and temporal framework necessary for precision public health, transforming passive surveillance into dynamic, and setting-specific interventions that match the adaptive sophistication of the pathogens themselves. The ongoing threat requires a fundamental shift from broad protocols to targeted strategies that consider detailed transmission dynamics and population vulnerabilities.
Materials and Methods
This retrospective study was conducted in the National Tuberculosis Reference Laboratory at Masih Daneshvari Hospital, National Research Institute of Tuberculosis and Lung Diseases (NRITLD), a WHO Collaborating Center, and the Mycobacteriology Research Center (MRC). Because NRITLD serves as the national referral center for DR-TB, our cohort is enriched for MDR-TB cases and may not capture the full epidemiological picture of TB in Iran, particularly for drug-susceptible disease. This referral bias is acknowledged in the interpretation of prevalence estimates. The study protocol was approved by the Medical Ethics Committee of Shahid Beheshti University of Medical Sciences (SBMU), and ethical approval was obtained from the NRITLD with the number IR.SBMU.NRITLD.REC.1403.090. After obtaining written consent from participants and receiving the ethical code from the Research Ethics Committees of NRITLD, data analysis started.
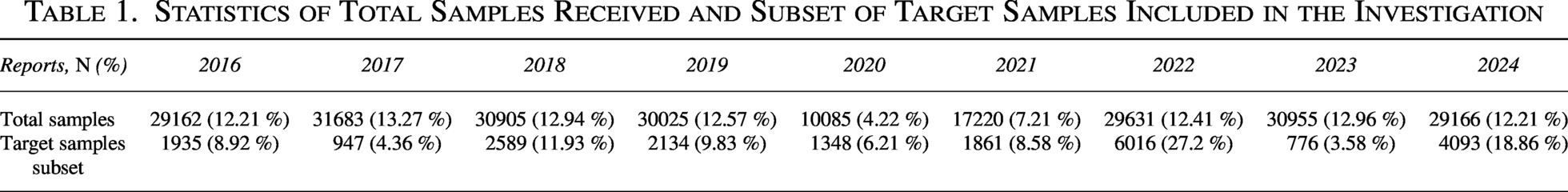
We examined laboratory records from January 2016 to December 2024, with the available complete laboratory and clinical profile of the targeting population encompassing 21,700 molecular and culture-confirmed mycobacterial samples (Fig. 1). The target subset for our analysis was explicitly defined as samples that underwent microscopic, culture, and molecular testing and for which a complete laboratory and clinical profile was available. This target subset of samples, which forms the basis for all subsequent statistical analyses in this study, is also detailed in Table 3. For contextual purposes, the total annual sample volume is also provided in Table 1, along with the subset of target samples included in this study.
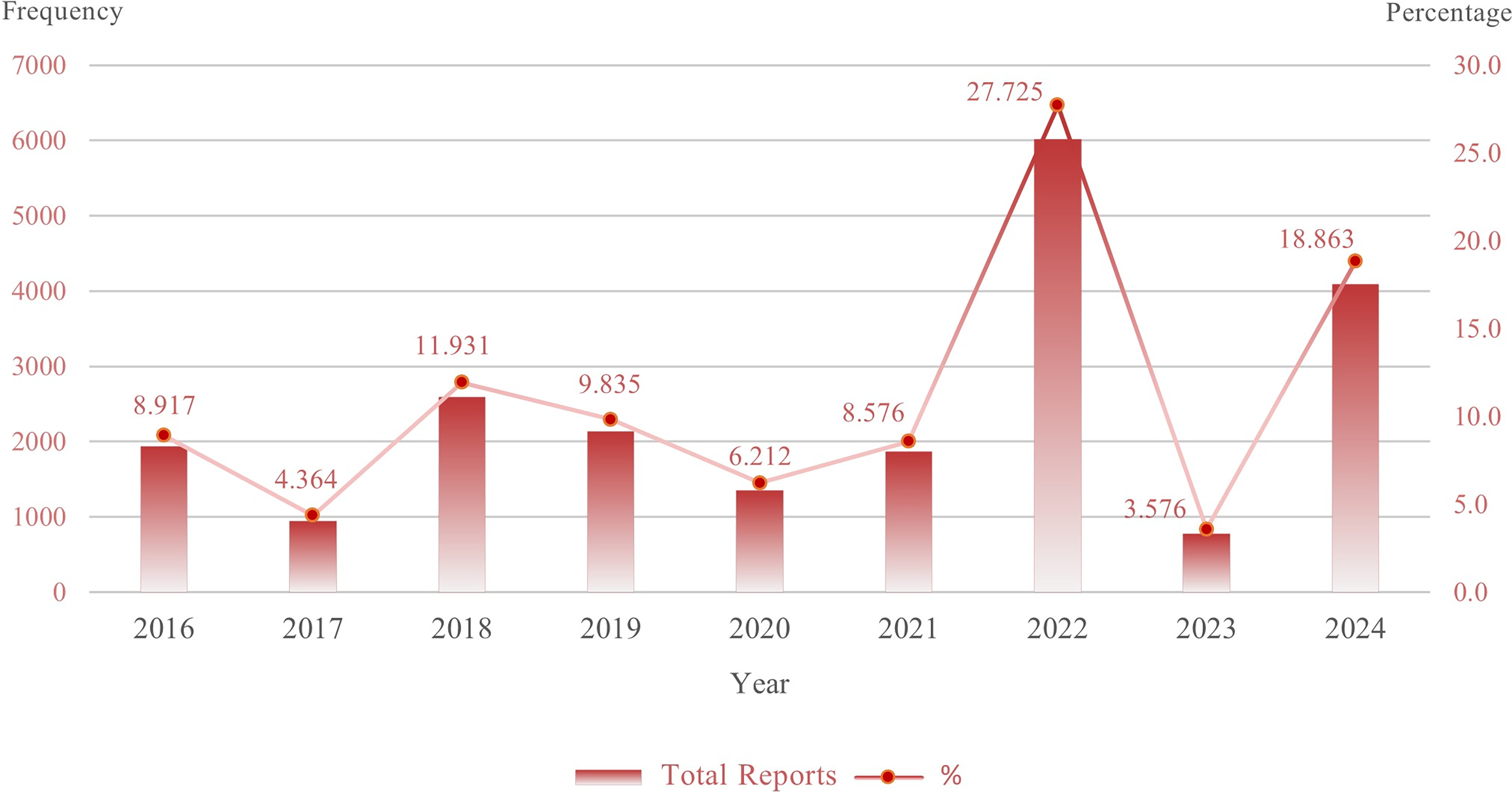
Annual frequency and percentage share of total reports incorporated in the research. Bar columns show the annual total reports for each year between 2016 and 2024. The superimposed line shows the corresponding percentage share of the total reports for each year.
Statistics of Total Samples Received and Subset of Target Samples Included in the Investigation
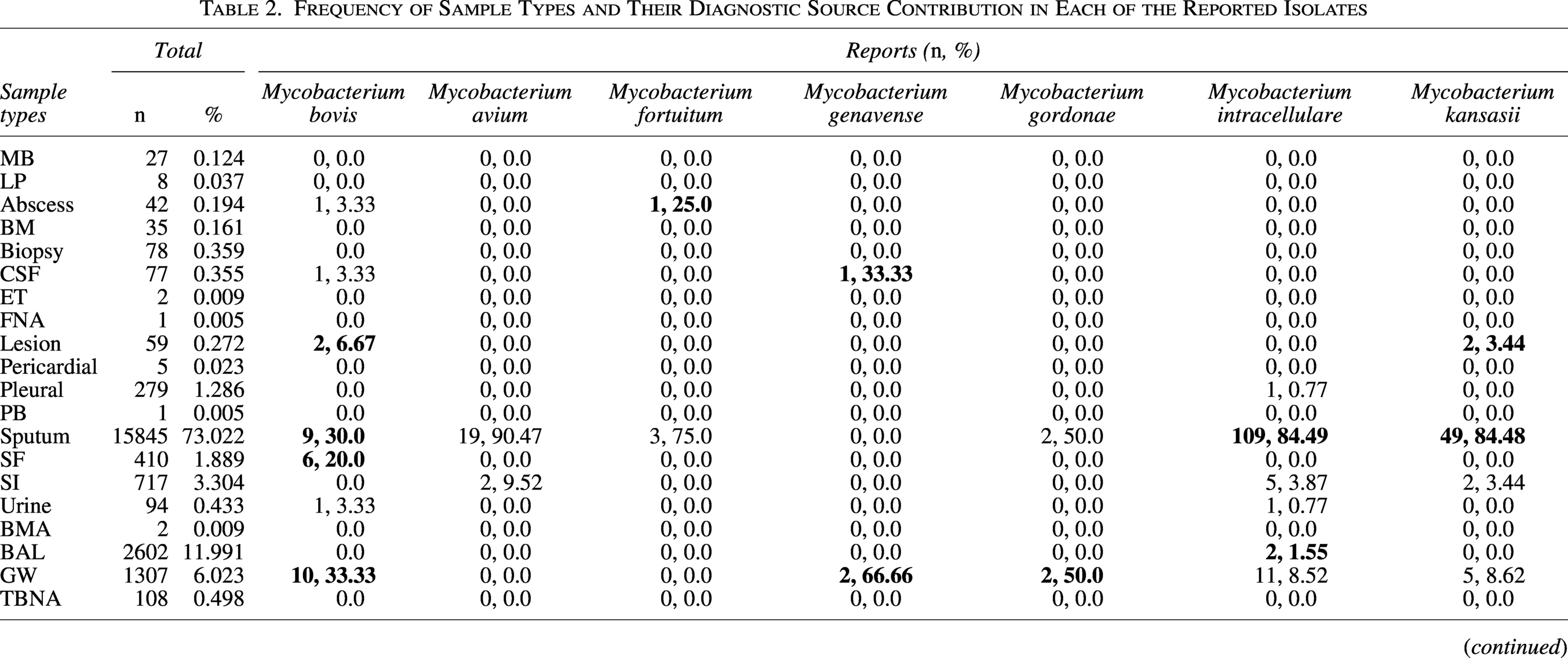
The target samples consisted of 20 distinct pulmonary and extrapulmonary specimen types (Table 2). Respiratory samples predominated, constituting 91.04% of all specimens, led by sputum (73.02%, n = 15,845) and bronchoalveolar lavage (bronchoalveolar lavage, 11.99%, n = 2,602). Gastric washings (GW) represented 6.02% (n = 1,307), primarily collected from pediatric patients. Extrapulmonary specimens included synovial fluid (SF, 1.88%, n = 410), pleural fluid (PF, 1.28%, n = 279), tissue biopsies (0.35%, n = 78), and menstrual blood (MB, 0.12%, n = 27), while rare samples like bone marrow aspirations (BMA, 0.009%, n = 2) and endometrial tissues (ET, 0.009%, n = 2) contributed minimally. This distribution ensured representation across diverse anatomical sites and clinical presentations, enabling robust analysis of species-specific tropisms. In the NTM-specific analyses, we focused only on pulmonary NTM (e.g., M. avium complex, M. abscessus), and extrapulmonary NTM cases were excluded to maintain a clinically homogeneous pulmonary disease cohort. Extrapulmonary TB cases were included in the analysis since they share similar transmission dynamics with pulmonary TB. However, extrapulmonary NTM infections were not examined further. All samples underwent a tiered diagnostic protocol: initial smear microscopy with Ziehl-Neelsen staining was followed by molecular screening using the Xpert MTB/RIF assay (Cepheid, Sunnyvale, CA, USA) for rapid detection of MTBC and rifampicin resistance. Specimens were then decontaminated with N-acetyl-L-cysteine-NaOH, concentrated by centrifugation, and inoculated onto Löwenstein-Jensen slants (Condalab, Madrid, Spain) and mycobacteria growth indicator tube liquid culture media (Becton Dickinson, Sparks, MD, USA).
Frequency of Sample Types and Their Diagnostic Source Contribution in Each of the Reported Isolates
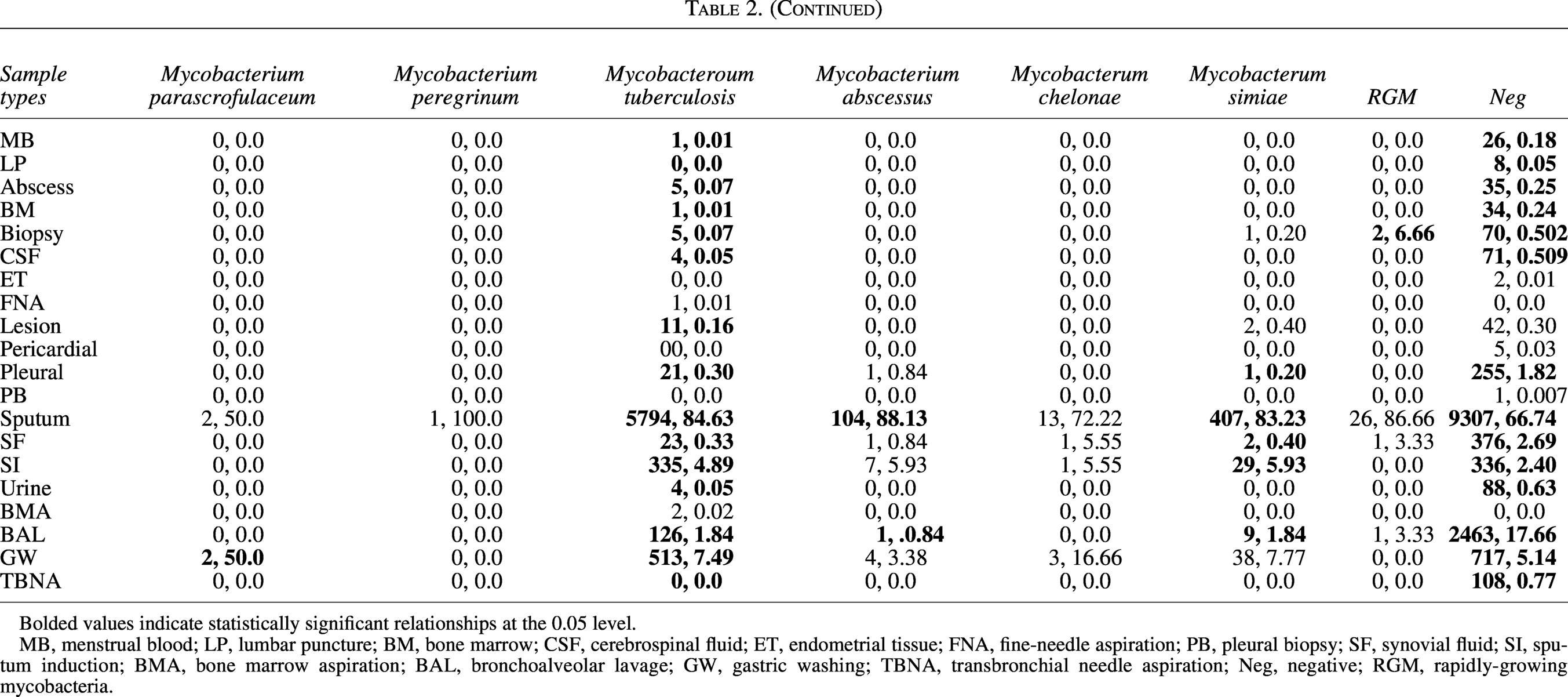
Bolded values indicate statistically significant relationships at the 0.05 level.
MB, menstrual blood; LP, lumbar puncture; BM, bone marrow; CSF, cerebrospinal fluid; ET, endometrial tissue; FNA, fine-needle aspiration; PB, pleural biopsy; SF, synovial fluid; SI, sputum induction; BMA, bone marrow aspiration; BAL, bronchoalveolar lavage; GW, gastric washing; TBNA, transbronchial needle aspiration; Neg, negative; RGM, rapidly-growing mycobacteria.
Prevalence of Resistant Isolates and Their Categorization Based on Patterns of Drug Resistance
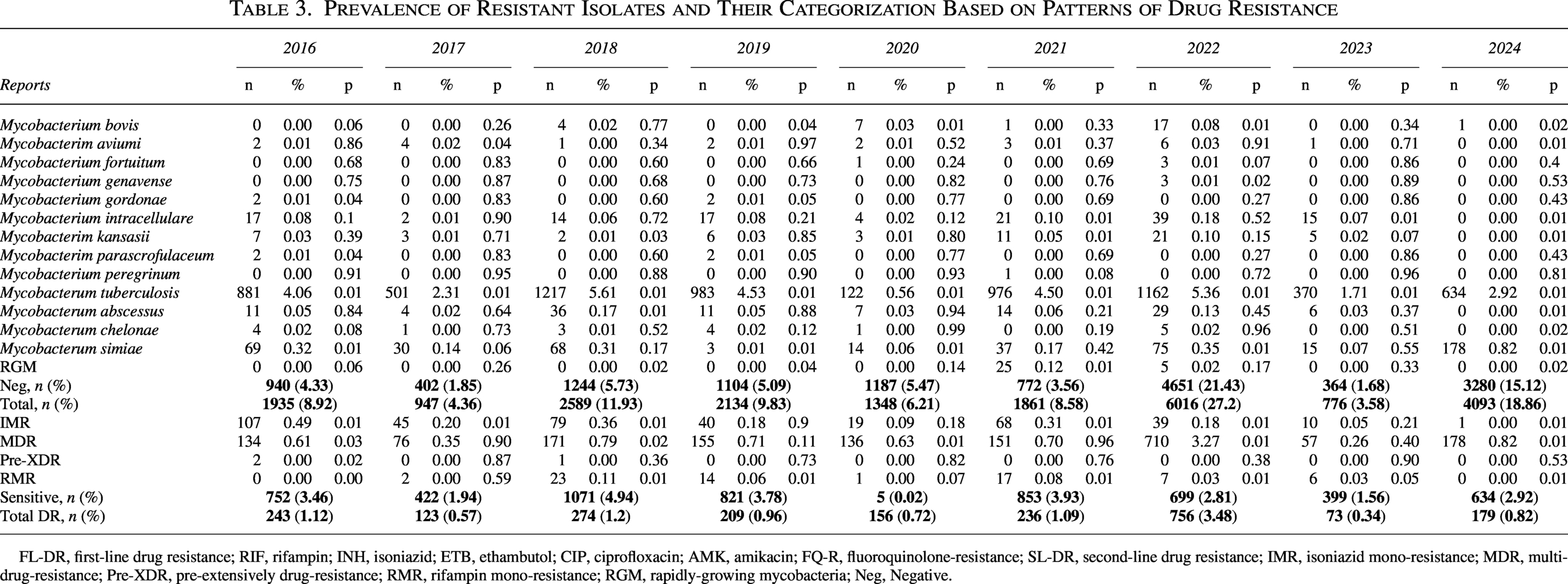
FL-DR, first-line drug resistance; RIF, rifampin; INH, isoniazid; ETB, ethambutol; CIP, ciprofloxacin; AMK, amikacin; FQ-R, fluoroquinolone-resistance; SL-DR, second-line drug resistance; IMR, isoniazid mono-resistance; MDR, multidrug-resistance; Pre-XDR, pre-extensively drug-resistance; RMR, rifampin mono-resistance; RGM, rapidly-growing mycobacteria; Neg, Negative.
Species identification employed line probe assays (GenoType MTBDRplus VER 2.0 for MTBC; GenoType NTM-DR for NTM, Hain Lifescience, Nehren, Germany), with discordant results between molecular and phenotypic methods resolved through targeted gene sequencing (rpoB for rifampicin; katG and inhA for isoniazid). Drug susceptibility testing followed WHO recommendations: first-line drugs used the proportion method on Löwenstein-Jensen medium, while second-line agents underwent broth microdilution (Sensititre MYCOTB, Thermo Fisher Scientific, Waltham, MA, USA). Resistance categories were defined as: isoniazid mono-resistance (IMR), rifampicin mono-resistance (RMR), MDR, and pre-extensively drug-resistant (Pre-XDR). Specifically, for NTM species, MDR was defined as resistance to at least two drugs from different classes recommended for the treatment. MDR for rapidly growing NTM (M. abscessus, M. chelonae) was defined as resistance to the key macrolide (clarithromycin) plus at least one other major antibiotic (e.g., amikacin or cefoxitin), following van Ingen et al. 14 For slow-growing NTM (e.g., M. avium complex, M. simiae), MDR was considered as resistance to both clarithromycin and amikacin; for M. kansasii, resistance to rifampicin and ethambutol. Clinical metadata included hospital acquisition status (>48 hours post-admission) and treatment units, including AIIR, pulmonary medicine (PM), emergency room (ER), intensive care units dedicated to tuberculosis (ICU-TB), health care associated infections (HAI), and OP designation. Patient gender was recorded from electronic health records as documented at registration.
Statistical analysis utilized the IBM-Statistical package for the Social Sciences (SPSS) version 27 and XLSTAT Perpetual version 2019.2.2. Annual prevalence rates were calculated with 95% confidence intervals (95% CI). Associations between resistance and predictor variables were assessed using chi-square or Fisher’s exact tests, with Cochrane-Armitage tests evaluating temporal trends. Statistical significance was set at a p value < 0.05 in the analyses.
Results
Sample type distribution and diagnostic yield
The diagnostic landscape encompassed 20 distinct specimen categories, with respiratory samples demonstrating overwhelming predominance: sputum accounted for 73.022% (n = 15,845) of all specimens, followed by bronchoalveolar lavage (11.991%, n = 2,602) and GW (6.023%, n = 1,307). Extrapulmonary specimens exhibited substantial diversity, ranging from common sources like SF (1.889%, n = 410) to rare collections such as pleural biopsies (0.005%, n = 1). Statistically significant pathogen-sample associations emerged, revealing distinct microbiological tropisms: abscess aspirates exclusively yielded M. fortuitum (25.0% of positive abscess isolates, p < 0.05) and M. bovis (3.33%, p < 0.05), while SF uniquely harbored M. chelonae (5.55% of synovial isolates, p < 0.05). Sputum yielded the highest absolute number of positive cultures for M. tuberculosis (84.63%, n = 5,794), M. abscessus (88.13%, n = 104), and M. simiae (83.23%, n = 407), though these likely reflect sample predominance rather than pathogenic tropism. GW emerged as an unexpected reservoirs for M. genavense (66.66%, n = 2) and M. gordonae (50.0%, n = 2), suggesting gastrointestinal colonization pathways. The comprehensive distribution, including all significant relationships, is detailed in Table 2.
Comprehensive overview of mycobacterial isolates
Over the 9-year study period (2016–2024), 21,700 samples meeting the study inclusion criteria were analyzed through microscopic, cultured, and molecular-based tests. (Table 3). M. tuberculosis consistently demonstrated the highest prevalence, ranging from 4.06% (n = 881) in 2016 to 5.61% (n = 1,217) in 2018. NTM exhibited dynamic fluctuations: M. simiae surged from 0.32% (n = 69) in 2016 to 0.82% (n = 178) in 2024 (p < 0.01), while M. abscessus peaked in 2018 (0.17%, n = 36; p = 0.01). Rapidly growing mycobacteria (RGM) emerged abruptly in 2021 (0.12%, n = 25; p = 0.01), with subsequent decline. Negative cultures (no growth) reached their zenith in 2022 (21.43%, n = 4,651), contrasting sharply with 2017 (1.85%, n = 402). Total specimen volume varied annually, peaking in 2022 (27.72%, n = 6,016) and troughing in 2023 (3.58%, n = 776). Figure 2 illustrates the distribution of the examined isolates throughout the research period.
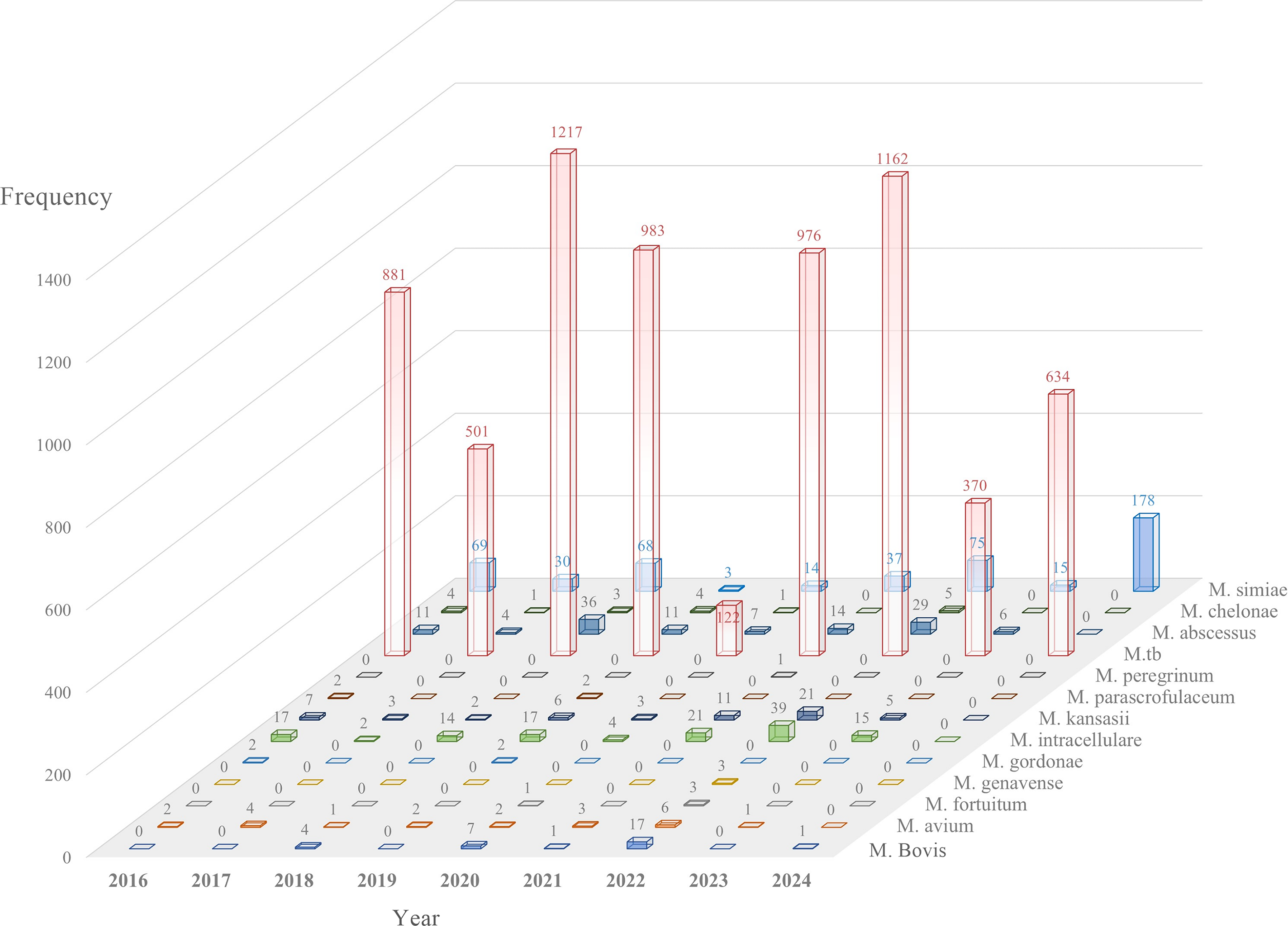
Distribution of the examined isolates during the study period. Bars display the frequency of each Mycobacterium species.
Annual dynamics of resistance categories
DR isolates increased significantly from 1.12% (n = 243) in 2016 to 3.48% (n = 756) in 2022 (p < 0.01), followed by a decline to 0.82% (n = 179) in 2024 (Table 3). MDR isolates demonstrated the steepest rise: 0.35% (n = 76) in 2017 to 3.27% (n = 710) in 2022 (p < 0.01). RMR fluctuated markedly, peaking in 2018 (0.11%, n = 23; p = 0.01) and 2021 (0.08%, n = 17; p = 0.01). Pre-XDR isolates remained minimal throughout (≤0.13%), with no cases in 2017, 2019, 2020, 2021, 2023, or 2024. IMR declined steadily from 0.49% (n = 107) in 2016 to 0.00% (n = 1) in 2024 (p < 0.01). Sensitive isolates (no resistance) ranged from 0.02% (n = 5) in 2020 to 4.94% (n = 1,071) in 2018. Figure 3 represents the annual trends in DR patterns among resistant isolates.
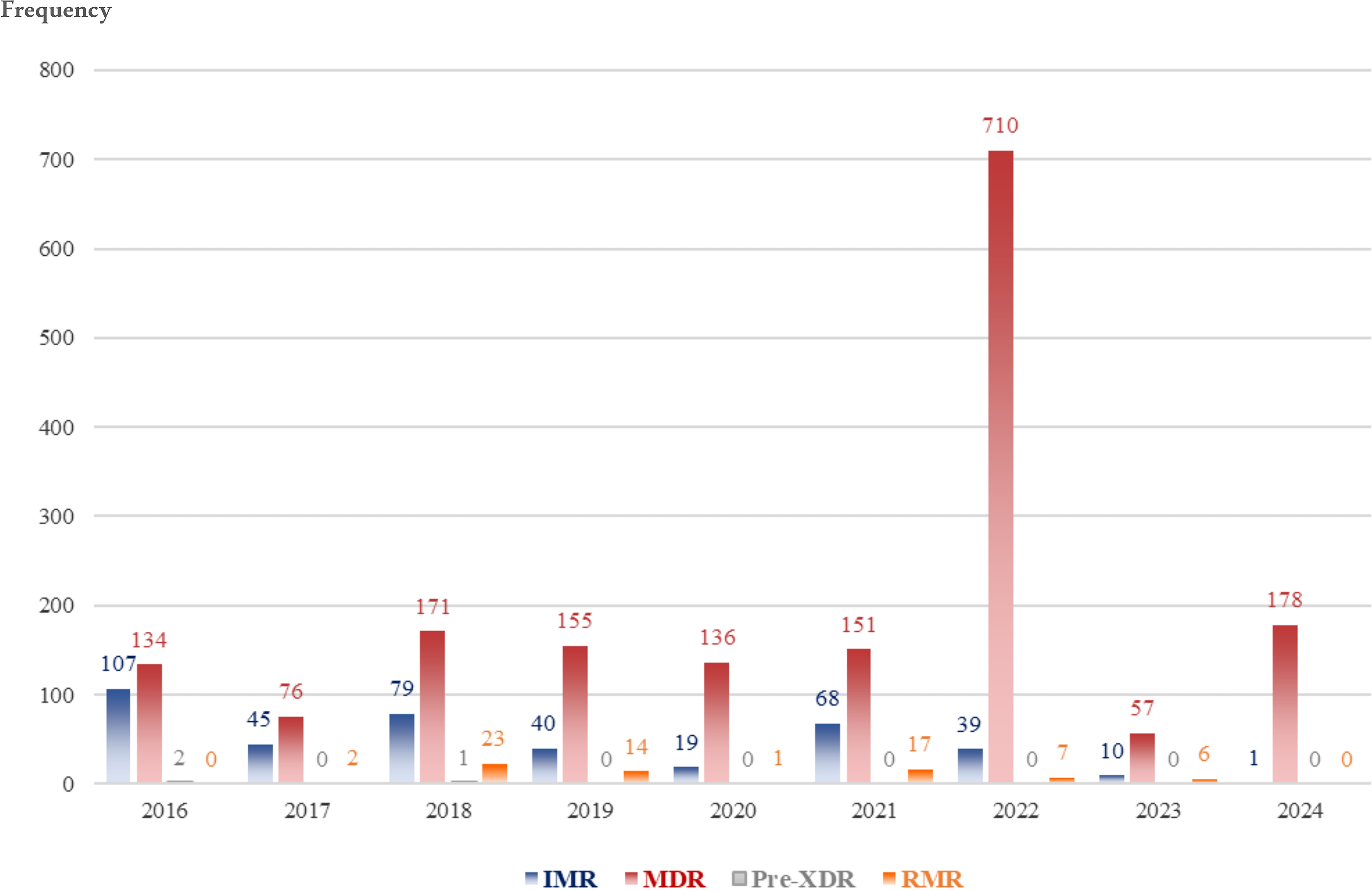
Yearly trends in drug-resistance patterns among diagnosed resistant isolates. Bars demonstrate annual frequency of each drug-resistance patterns across the years. Each bar corresponds to one resistance category. IMR, isoniazid mono-resistant; RMR, rifampicin mono-resistant; MDR, multidrug-resistant; Pre-XDR, pre–extensively drug-resistant.
Species-specific resistance profiles
Among 2,250 DR isolates categorized (Table 4), M. tb dominated resistance patterns, accounting for 59.76% (n = 1,344) of all DR cases. The prevalence of first-line drug resistance in MDR isolates was 67.5% (n = 1,519) for rifampin, 78.5% (n = 1,766) for isoniazid, and 69.7% (n = 1,568) for ethambutol. Conversely, second-line resistance was rare: ciprofloxacin and amikacin each affected 0.13% (n = 3) of isolates. For nontuberculous species, M. simiae accounted for 21.74% (n = 489) of all DR isolates, followed by M. intracellulare and M. abscessus, which constituted 5.73% (n = 129) and 5.24% (n = 118) of DR isolates, respectively (Table 4).
Prevalence of Resistant Isolates and Their Categorization Based on Patterns of Drug Resistance
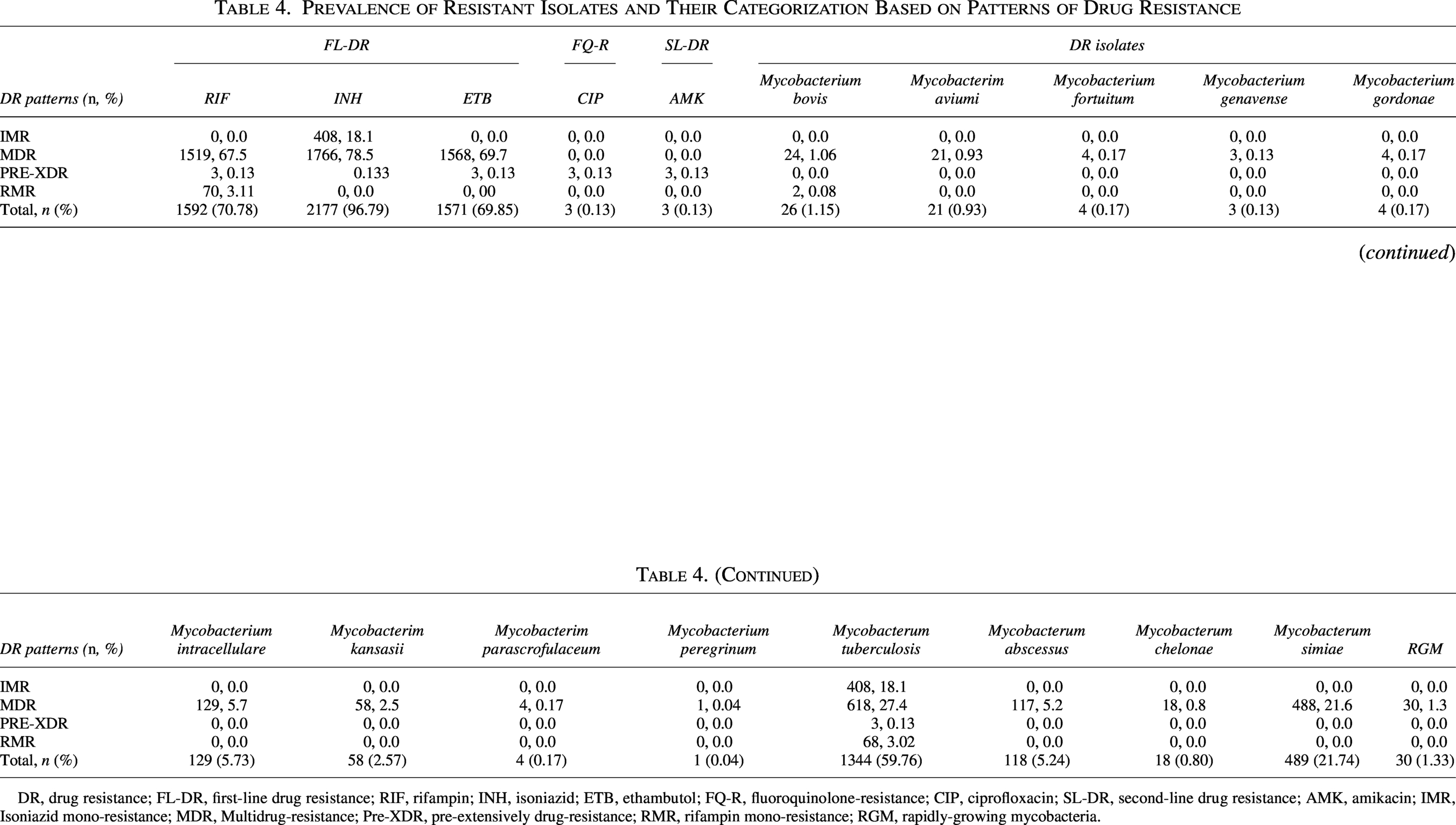
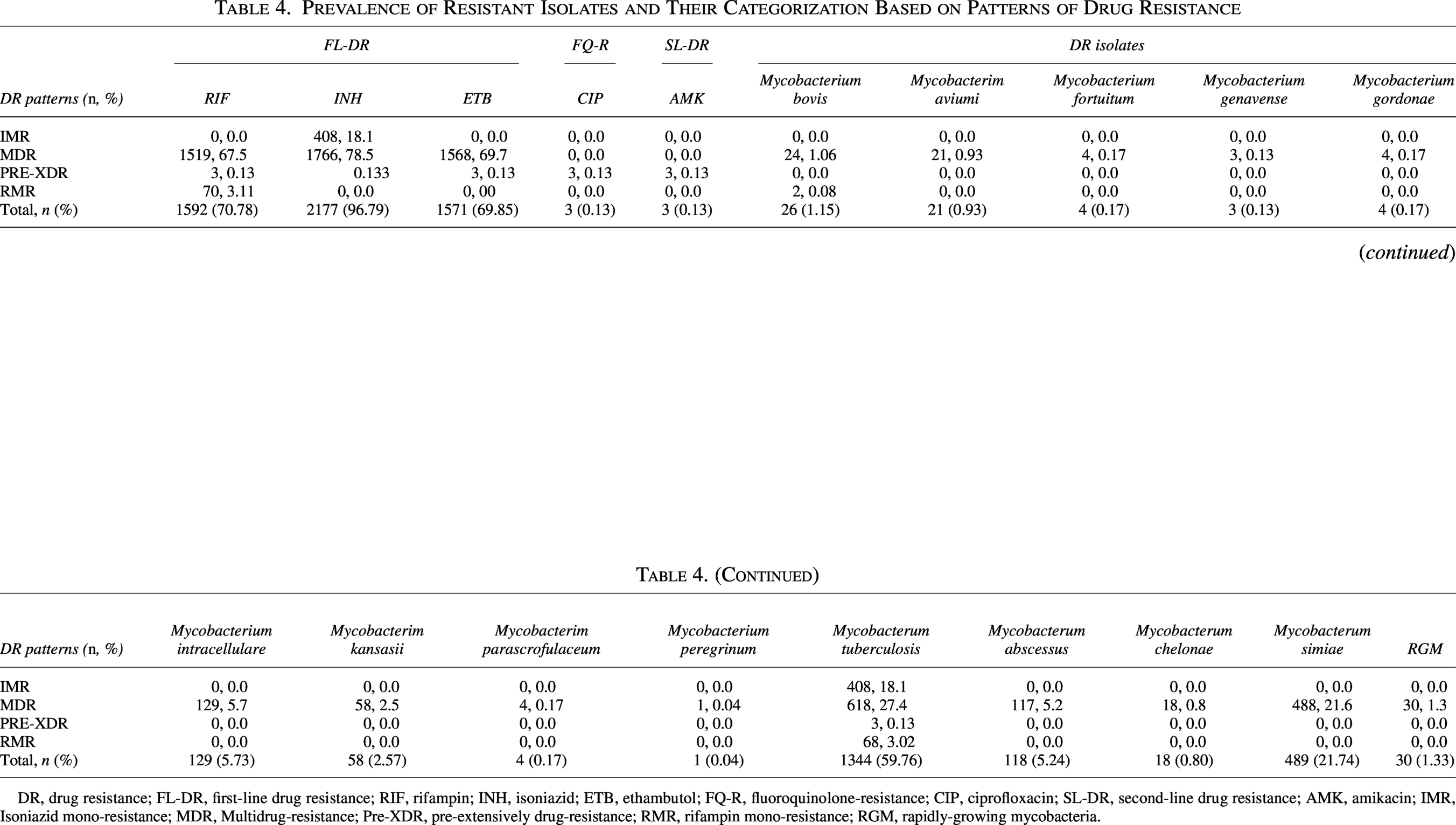
DR, drug resistance; FL-DR, first-line drug resistance; RIF, rifampin; INH, isoniazid; ETB, ethambutol; FQ-R, fluoroquinolone-resistance; CIP, ciprofloxacin; SL-DR, second-line drug resistance; AMK, amikacin; IMR, Isoniazid mono-resistance; MDR, Multidrug-resistance; Pre-XDR, pre-extensively drug-resistance; RMR, rifampin mono-resistance; RGM, rapidly-growing mycobacteria.
Smear microscopy correlations with resistance
The persistence of acid-fast bacilli in stained smears served as a powerful predictor for specific resistance profiles (Table 5). M. tuberculosis isolates demonstrated near-universal smear positivity regardless of grading intensity, with statistically robust correlations observed across all concentrations from scanty (1+) to heavy (3+) bacillary loads (p = 0.01). This consistent visibility under microscopy starkly contrasted with RGM, which defied conventional detection by associating almost exclusively with smear-negative specimens (p = 0.001). Among resistance patterns, MDR strains mirrored M. tuberculosis’s behavior through unwavering smear positivity at every grade (p = 0.01), while RMR isolates revealed a selective affinity for moderate-to-heavy bacillary burdens. Based on the results, RMR isolates significant positivity for grade 2+ (p = 0.01) and 3+ (p = 0.01) smears. Pre-XDR strains operated outside these trends, showing no statistically meaningful relationship to smear status whatsoever (p > 0.05). The diagnostic significance of negative smears extended beyond RGM to encompass other environmentally derived species like M. gordonae (p = 0.01) and M. chelonae (p = 0.00), creating a clear phenotypic division between tuberculosis complexes and opportunistic environmental mycobacteria when viewed through the lens of microscopy.
Statistical Significance (p-Values) Association of Smear Reports with Typing and DR Patterns
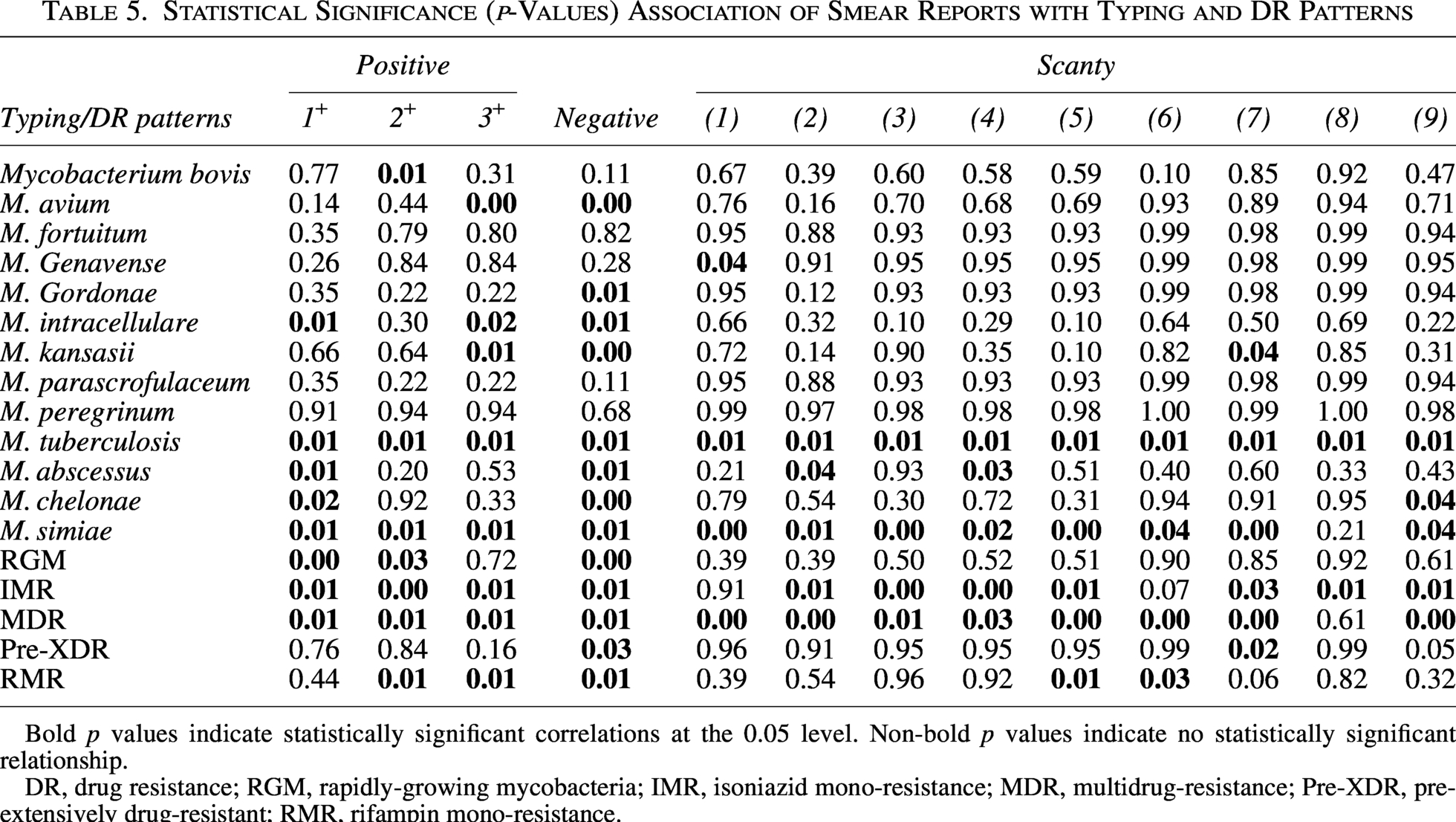
Bold p values indicate statistically significant correlations at the 0.05 level. Non-bold p values indicate no statistically significant relationship.
DR, drug resistance; RGM, rapidly-growing mycobacteria; IMR, isoniazid mono-resistance; MDR, multidrug-resistance; Pre-XDR, pre-extensively drug-resistant; RMR, rifampin mono-resistance.
Critical hospital transmission hotspots
Distribution mapping uncovered sharply defined geographical niches for resistance propagation within the health care ecosystem (Table 6). Despite their designation for transient care, OP clinics emerged as formidable reservoirs for IMR M. tuberculosis and MDR M. simiae, with both pathogens demonstrating pronounced statistical affinity for these settings. The highest burden was for IMR M. tuberculosis (12.67%, n = 285; p < 0.0001), MDR M. tuberculosis (12.31%, n = 277; p = 0.01), and MDR M. simiae (14.53%, n = 327; p < 0.0001). Additionally, MDR M. abscessus isolates were also concentrated in OP settings, accounting for 3.82% (n = 86; p < 0.0001). The AIIRs engineered for containment paradoxically functioned as epicenters for MDR-TB transmission, registering the highest concentration of such isolates anywhere in the facility. According to the results, the AIIRs were the epicenter for MDR M. tuberculosis (18.22%, n = 410; p < 0.0001) and MDR M. simiae (6.57%, n = 148; p < 0.0001). Also, the burdens of the IMR M. tuberculosis (4.49%, n = 101; p = 0.04) and MDR M. intracellulare (0.80%, n = 18; p < 0.0001) were significant in AIIRs. PM units harbored unexpected reservoirs of zoonotic tuberculosis, with MDR M. bovis (0.27%, n = 6; p < 0.0001) and RMR M. bovis (0.09%, n = 2; p = 0.01) strains virtually confined to these specialized wards. ICU-TB revealed an exclusive association with pre-XDR M. tuberculosis (0.09%, n = 2; p = 0.02), while ERs accumulated statistically significant clusters of MDR M. tuberculosis despite shorter patient exposure times (1.51%, n = 34; p = 0.005). HAI wards concentrated MDR M. simiae (0.31%, n = 7; p = 0.01) and IMR M. tuberculosis (0.18%, n = 4; p < 0.0001) species, creating micro-epidemics within these controlled environments that defied broader institutional resistance trends. The distribution of resistant isolates and their DR patterns in hospital wards is presented in Figure 4.
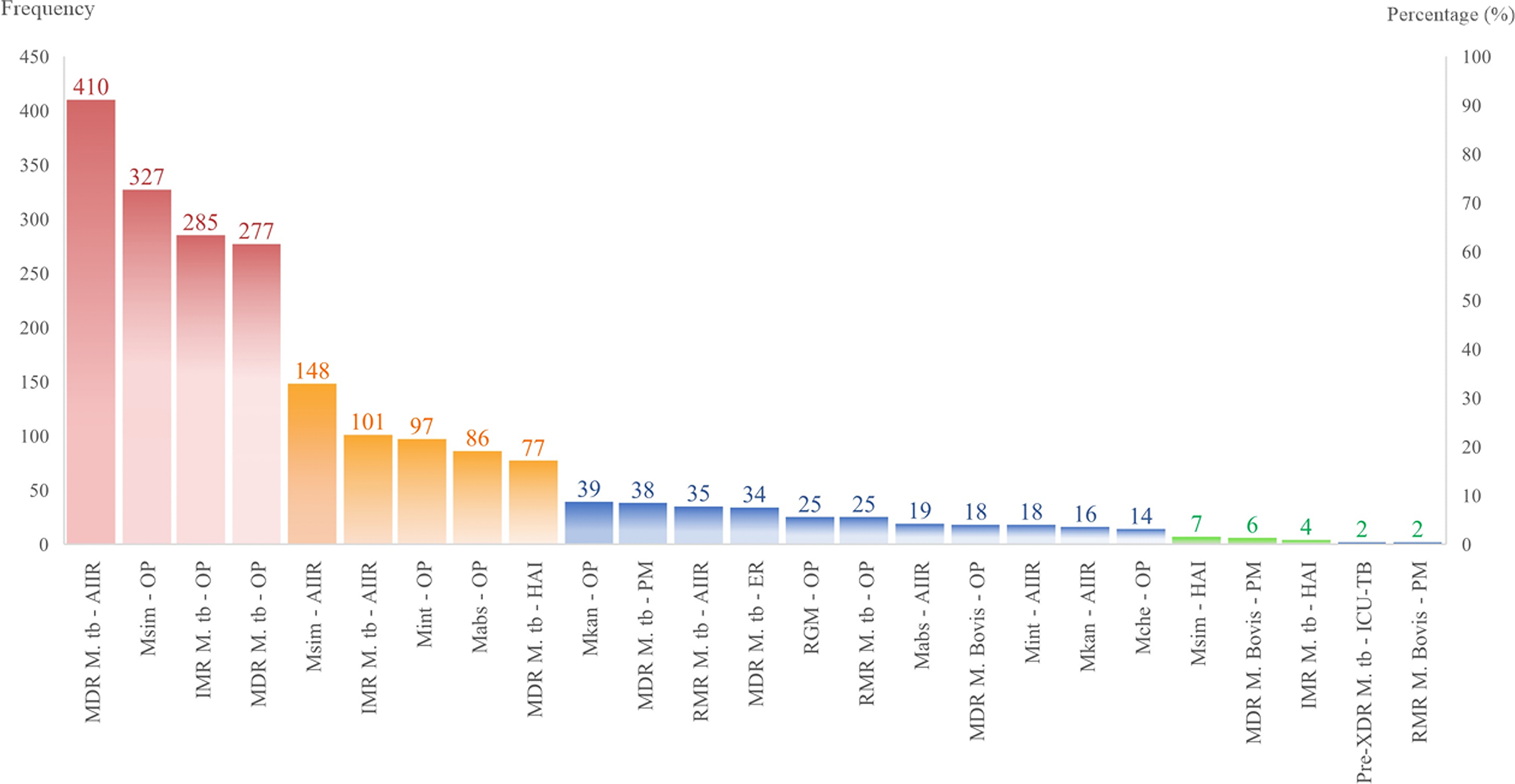
Distribution of resistant strains along with their drug resistance patterns in hospital wards. IMR, Isoniazid mono-resistance; MDR, Multidrug-resistance; Pre-XDR, Pre-extensively drug-resistant; RMR, rifampin mono-resistance; RGM, rapidly-growing mycobacteria.
Distribution of Drug-Resistant Isolates and Their Significant Relationship to Hospital Sections
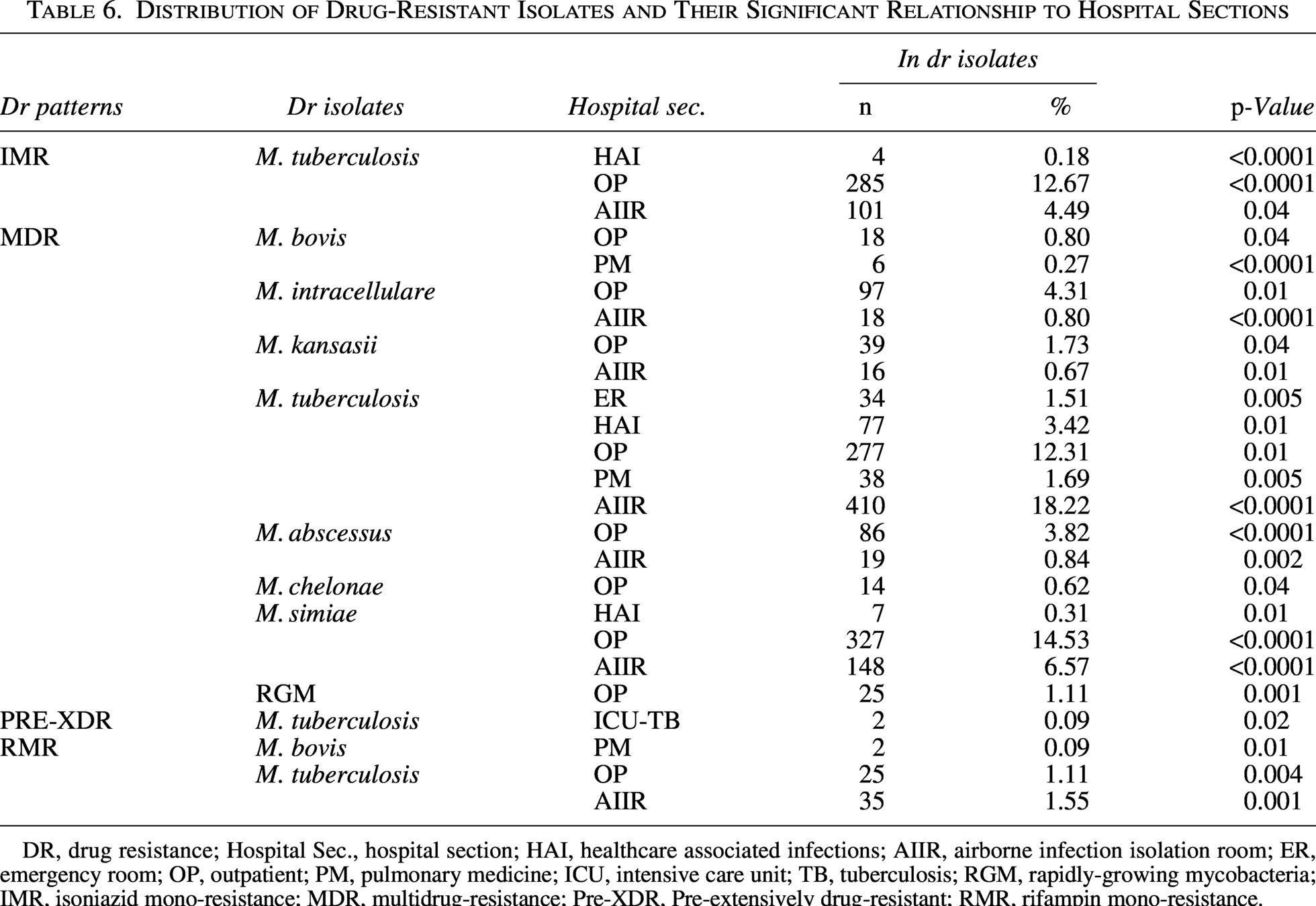
DR, drug resistance; Hospital Sec., hospital section; HAI, healthcare associated infections; AIIR, airborne infection isolation room; ER, emergency room; OP, outpatient; PM, pulmonary medicine; ICU, intensive care unit; TB, tuberculosis; RGM, rapidly-growing mycobacteria; IMR, isoniazid mono-resistance; MDR, multidrug-resistance; Pre-XDR, Pre-extensively drug-resistant; RMR, rifampin mono-resistance.
Gender-specific disparities in resistance
The drug resistance patterns among male and female patients are distributed and compared in Figure 5. A substantial gender gap pervaded resistance patterns, showing physiologically and sociologically relevant differences (Table 7). According to statistical analyses, the total DR burden was 64.03% in men and 35.97% in female patients. Male patients continuously carried higher loads of MDR-TB, more than double the incidence found in female patients throughout the research period (26.01% males vs. 12.45% females, p = 0.005). This male predominance extended beyond tuberculosis to encompass environmentally acquired pathogens like MDR M. simiae (12.36% males vs. 9.38% females, p = 0.001) and MDR M. kansasii (2.05% males vs. 0.53% females, p = 0.012), suggesting systemic rather than pathogen-specific vulnerability factors. Conversely, female patients demonstrated heightened susceptibility to specific NTM, including MDR M. abscessus (2.76% males vs. 2.49% females, p = 0.009) and MDR M. chelonae (0.53% females vs. 0.27% males, p = 0.010), with statistically significant differences in resistance prevalence. The rarest resistance category—pre-XDR—manifested exclusively among female tuberculosis patients, though absolute numbers remained minimal (0.13% females vs. 0.00% males, p = 0.046). When considering the total resistance landscape, the total DR burden in male patients outnumbered female patients by nearly two-to-one (64.03% male vs. 35.97% female), establishing gender as a fundamental determinant in the epidemiology of mycobacterial resistance within this population. Figure 6 depicts the distribution and comparability of resistant isolates along with DR patterns proportioned by gender.
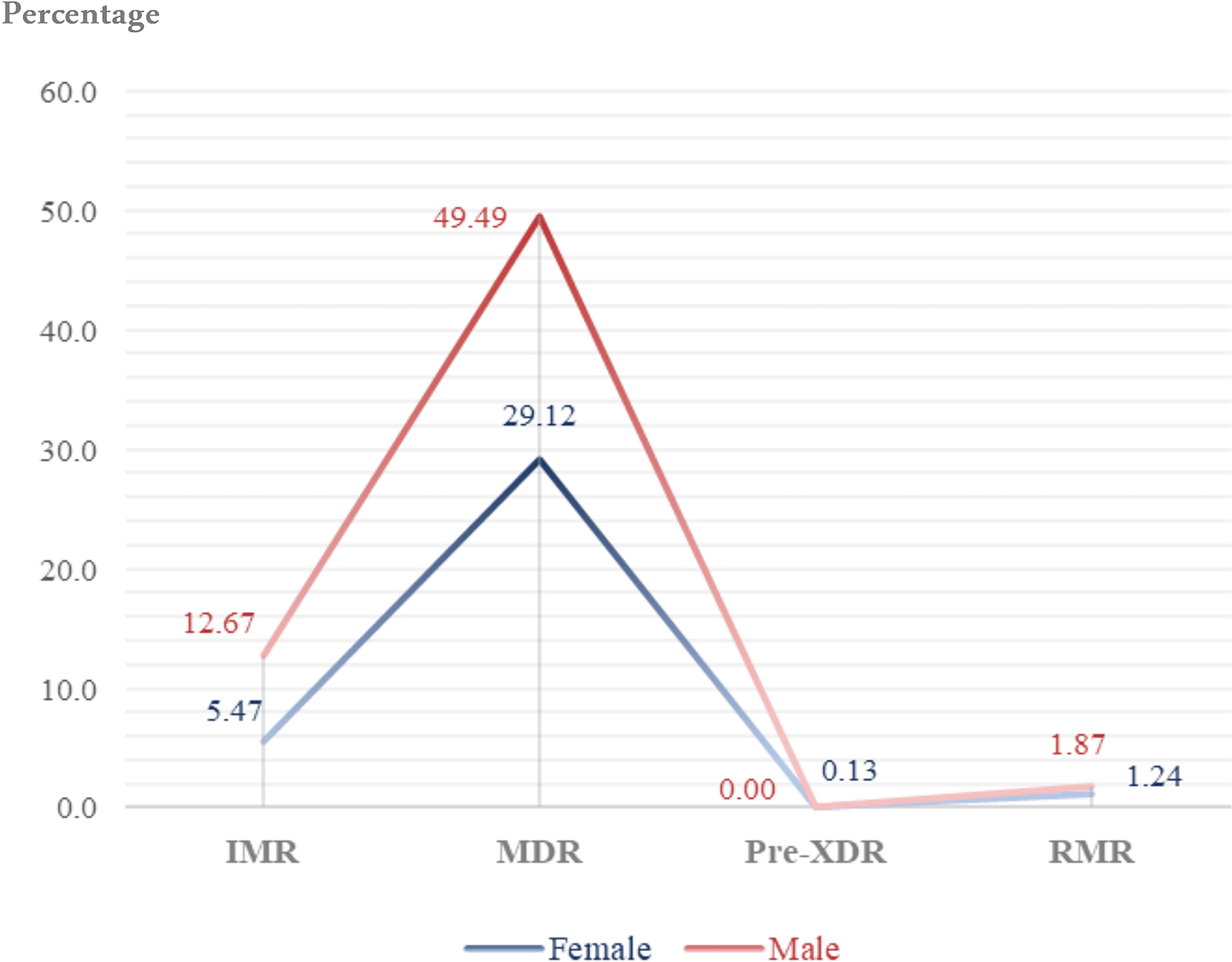
Distribution and comparison of drug resistance patterns among male and female patients. Percent values are indicated on the y-axis and resistance categories (IMR, MDR, Pre-XDR, RMR) are shown on the x-axis. IMR, isoniazid mono-resistant; MDR, multidrug-resistant; Pre-XDR, pre–extensively drug-resistant; RMR, rifampicin mono-resistant.
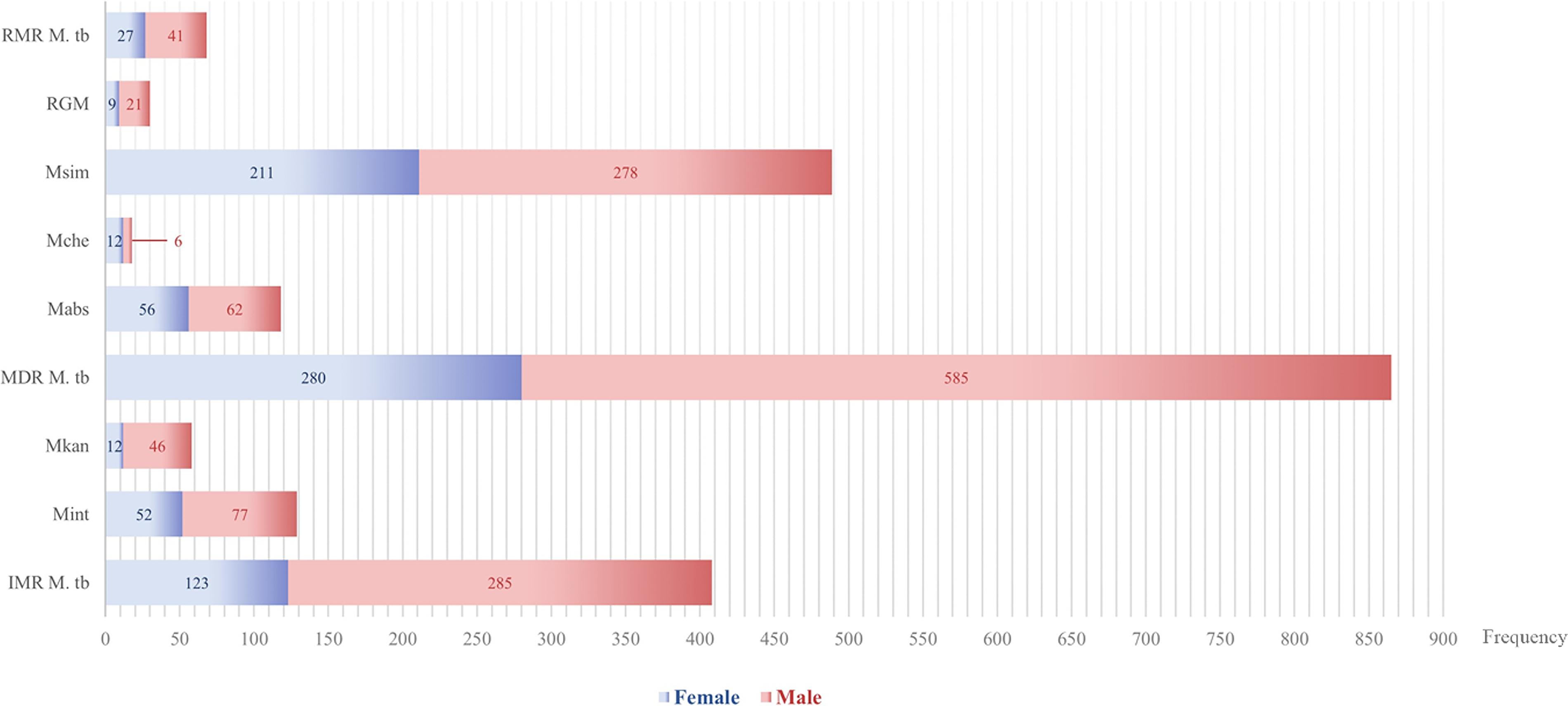
Distribution and comparison of resistant isolates along with drug resistance patterns by gender. Grouped bars indicate the distribution of resistant isolates and drug resistance patterns in male and female patients. Frequency values are shown on the x-axis, and grouped bars indicate the gender-stratified proportion for resistant isolates along with their drug resistance patterns. RMR, rifampicin mono-resistant; IMR, isoniazid mono-resistant; MDR, multidrug-resistant; RGM, rapidly-growing mycobacteria.
Yearly Trends in the Prevalence of Resistant Isolates and Their Association with Gender
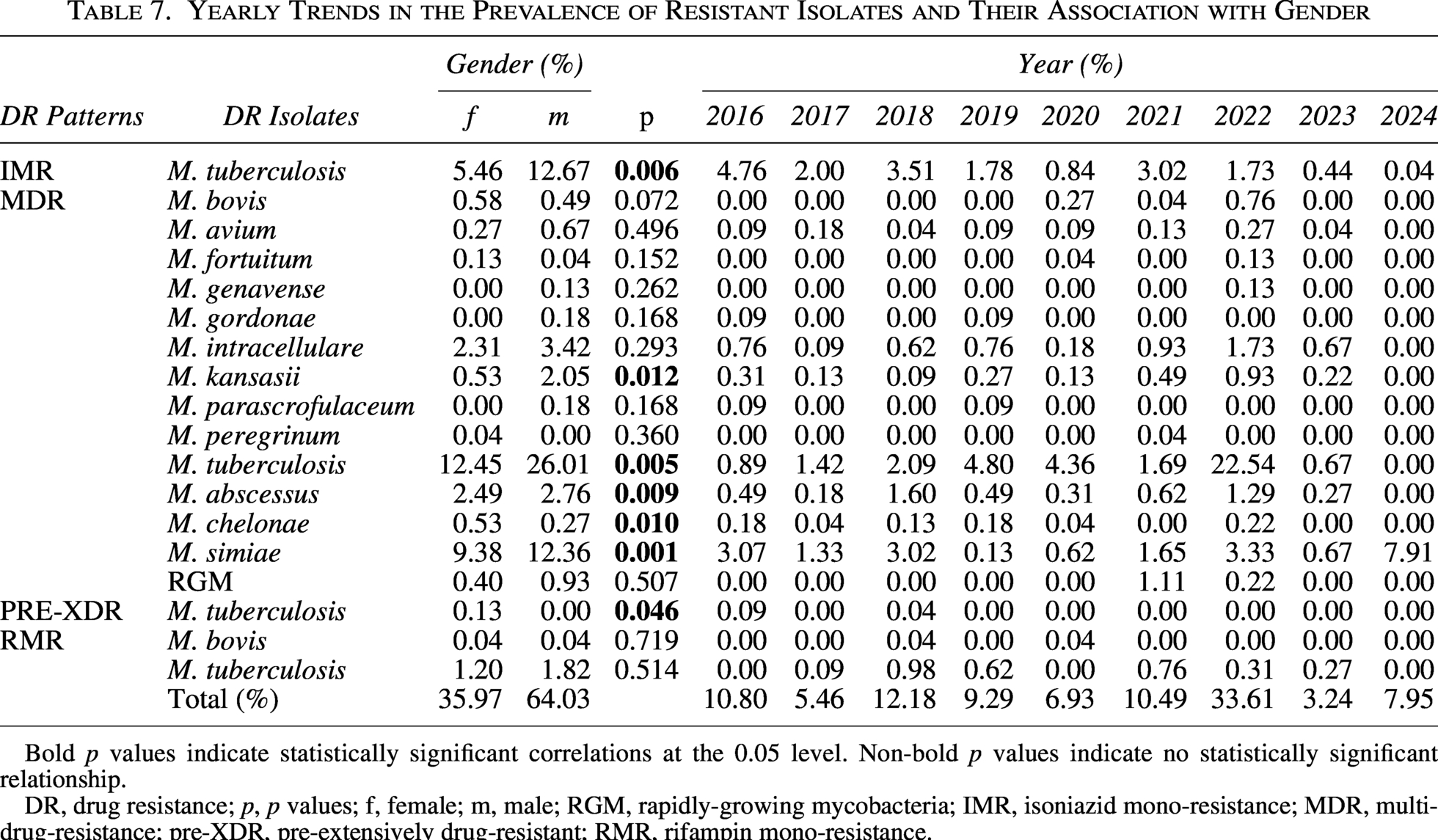
Bold p values indicate statistically significant correlations at the 0.05 level. Non-bold p values indicate no statistically significant relationship.
DR, drug resistance; p, p values; f, female; m, male; RGM, rapidly-growing mycobacteria; IMR, isoniazid mono-resistance; MDR, multidrug-resistance; pre-XDR, pre-extensively drug-resistant; RMR, rifampin mono-resistance.
Temporal evolution of species resistance
The 9-year surveillance period captured dynamic evolutionary trajectories among resistant pathogens (Table 7). The epidemiological landscape of MDR-TB experienced a significant transformation, moving from a phase of relative stability in the first 5 years of the study to a remarkable increase in the second to last year, which fundamentally altered the institutional resistance dynamics. The MDR M. tuberculosis dramatically surged in 2022 (22.54% of all isolates that year) after relatively stable levels during 2016–2019 and 2020 (0.89%–4.80% and 4.36%). Conversely, IMR M. tuberculosis displayed a consistent downward trajectory that nearly eliminated this resistance profile by the final surveillance year (4.76% in 2016 to 0.04% in 2024), while RMR M. tuberculosis fluctuated erratically without establishing a clear directional trend (fluctuating pattern with peaks of 0.98% in 2018 and 0.76% in 2021). The resistance of M. simiae exhibited a contrasting temporal pattern marked by a steady yet unyielding increase, leading to an unparalleled prevalence by the end of the study. The prevalence of M. simiae rose from 3.07% in 2016 to 7.91% in 2024, with a secondary peak in 2022 (3.33%). M. abscessus followed a bimodal trajectory with distinct peaks separated by several years (1.60% in 2018 and 1.29% in 2022), suggesting periodic selective pressures rather than continuous transmission. These temporal patterns collectively illustrated how different mycobacterial species and resistance mechanisms respond disparately to changing clinical practices, environmental pressures, and population immunity over extended observation periods.
Negative and sensitive isolate trends
An inverse relationship between negative cultures (no growth) and confirmed resistance has emerged as a defining characteristic of institutional epidemiology (Table 3). The proportion rose from 4.33% (n = 940) in 2016–21.43% (n = 4,651) in 2022, then declined to 15.12% (n = 3,280) in 2024. The expansion of negative culture rates during the study’s mid-years resulted in diagnostic gaps that temporarily hindered resistance surveillance. The rise of non-diagnostic specimens ultimately slowed down, although it stayed far higher than baseline levels. Drug-sensitive cultured isolates showed a distinctive rhythm that changed independently of resistance trends and stayed the same throughout the observation period. Sensitive isolates (no resistance) showed variability: highest in 2018 (4.94%, n = 1,071) and lowest in 2020 (0.02%, n = 5). Annual fluctuations in sensitive isolates did not correlate with DR trends (p > 0.05). The cyclical pattern of these non-resistant groups countered resistance escalation and provided context for evaluating the absolute load of resistant infections in the specimen landscape over the observation period.
Smear negativity in the NTMs group
The diagnostic challenge of smear-negative, culture-positive NTM emerged as a significant finding, with Table 8 revealing substantial proportions of DR isolates evading microscopic detection. Among 879 NTM isolates, M. simiae dominated the smear-negative landscape, accounting for 55.63% (n = 489) of total NTM isolates, yet 20.65% (101/489) of them demonstrated smear negativity, revealing that approximately one in five resistant cases escaped initial microscopic diagnosis. Extension of this pattern to other clinically significant species was also observed.; M. abscessus exhibited 38.14% smear negativity (45/118) despite representing 13.42% (n = 118) of total NTM isolates, while M. intracellulare showed 32.56% (42/129) of smear-negative resistance. RGM, though comprising only 3.41% (n = 30) of the NTM isolates, demonstrated 23.33% (7/30) smear-negative cases. Strikingly, M. fortuitum displayed the highest proportion of smear-negative resistance at 75% (3/4), suggesting particular elusiveness to conventional microscopy. Only M. gordonae defied this trend with 0% smear negativity (0/4), though its overall contribution remained minimal. These findings collectively demonstrate that smear microscopy fails to detect 11.49–75% of resistant NTM isolates across key species, creating critical diagnostic blind spots where culture-independent methods miss active resistant infections, particularly in M. simiae and M. abscessus which collectively represented 69.05% of the NTM burden yet accounted for 62.86% (101 + 45:146) of all smear-negative NTM cases.
The Isolates’ Contribution and the Percentage of Negative Smear Reports in the NTMs Group
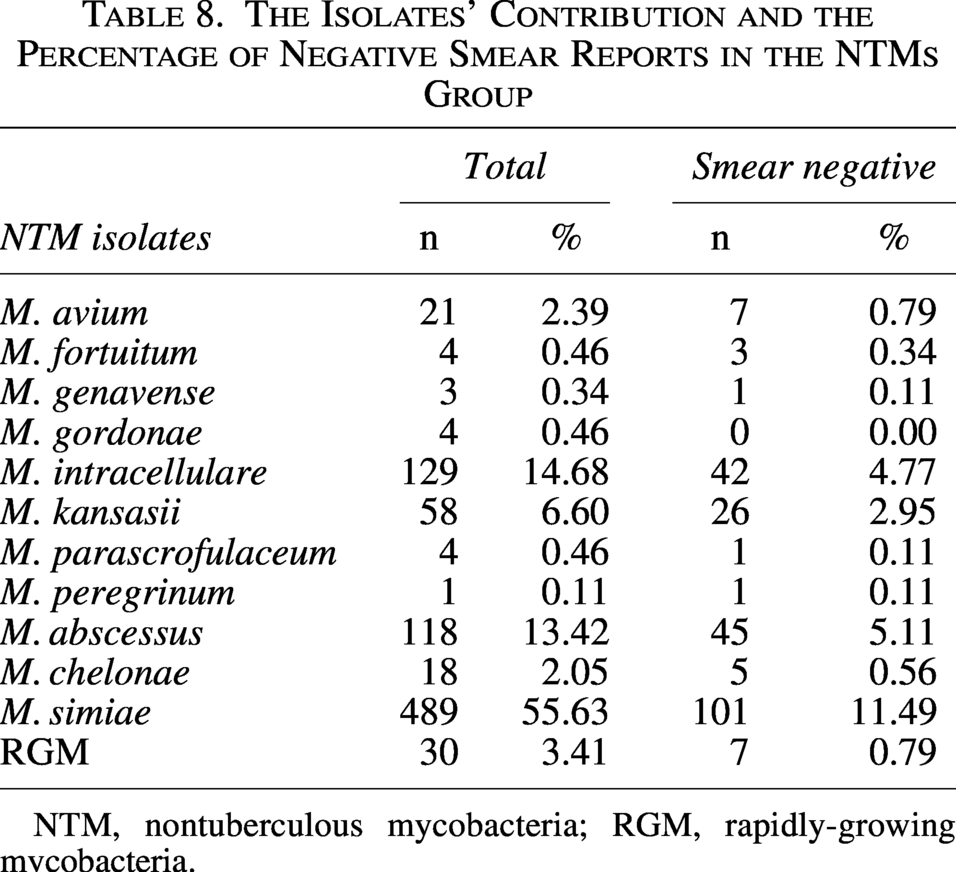
NTM, nontuberculous mycobacteria; RGM, rapidly-growing mycobacteria.
Unit-specific resistance rarities
Beyond the major transmission hotspots, several specialized units harbored statistically significant yet comparatively rare resistance patterns that revealed unexpected epidemiological niches (Table 6). PM units emerged as critical reservoirs for zoonotic tuberculosis, hosting both MDR M. bovis at 0.27% prevalence (n = 6, p < 0.0001) and RMR M. bovis at 0.09% (n = 2, p = 0.01), suggesting persistent zoonotic transmission chains in these specialized respiratory wards. The ER unit, despite its transient patient population, unexpectedly accumulated clusters of MDR M. tuberculosis (1.51%, n = 34, p = 0.005), indicating potential undetected transmission during acute care encounters. HAI wards demonstrated significant clustering of MDR M. simiae (0.31%, n = 7, p = 0.01), contradicting assumptions about this pathogen’s environmental exclusivity. ICU-TB revealed an exclusive association with pre-XDR M. tuberculosis (0.09%, n = 2, p = 0.02), marking these high-acuity settings as breeding grounds for the most advanced resistance. OP clinics sustained unexpected transmission of typically health care-associated species like M. chelonae (0.62%, n = 14, p = 0.04) alongside RGM (1.11%, n = 25, p = 0.001), establishing community-hospital interfaces as permeable boundaries for diverse resistance threats. AIIRs concentrated RMR M. tuberculosis at 1.55% prevalence (n = 35, p = 0.001), distinct from broader MDR-TB patterns and suggesting specialized selection pressures within these engineered environments. These statistically robust peculiarities, though individually uncommon, collectively represented 4.7% of all resistant isolates and revealed how localized environmental factors and patient flow patterns create micro-epidemics that deviate from institutional resistance trends while contributing meaningfully to the overall resistance landscape.
Gender-temporal interactions
The intersection of gender and temporal evolution revealed complex resistance dynamics that reshaped the epidemiological landscape throughout the study period (Table 7). Male patients disproportionately bore the brunt of the 2022 MDR-TB surge, which accounted for 22.54% of that year’s resistant isolates and amplified the established male predominance in MDR-TB (26.01% male vs. 12.45% female prevalence, p = 0.005). Conversely, M. simiae resistance demonstrated a delayed peak among females in 2024 (7.91% of annual isolates), even as males maintained significantly higher overall prevalence (12.36% vs. 9.38%, p = 0.001) throughout the surveillance period. The bimodal resistance pattern of M. abscessus—peaking in 2018 (1.60%) and 2022 (1.29%)—manifested divergent gender distributions; females dominated the earlier peak while males prevailed during the later resurgence, reflecting shifting vulnerability patterns that reinforced the overall male skew (2.76% vs. 2.49%, p = 0.009). IMR in tuberculosis exhibited a consistent gender-neutral decline from 4.76% (2016) to 0.04% (2024), contrasting sharply with M. kansasii’s stable male predominance (2.05% male vs. 0.53% female, p = 0.012) that persisted without temporal fluctuation. These intersecting trajectories illustrate how resistance epidemics evolve through distinct demographic channels rather than uniform population-wide progression, with male vulnerability amplifying temporal surges in tuberculosis resistance while female populations experienced delayed peaks in emerging pathogens like M. simiae.
Discussion
The mycobacterial infection epidemiological landscape in Iran has experienced significant transformations from 2016 to 2024 due to three interrelated crises: the disturbing return of MDR-TB, the rise of geographically atypical NTMs, and the contradictory function of specialized containment units as transmission hotspots. Our data reveal a ninefold increase in MDR-TB prevalence (0.35–3.27%, p < 0.01) that peaked during the 2022 health care disruption period—a trend diverging sharply from Iran’s pre-2016 stability (0.5–1.2% MDR-TB). This surge coincides with regional studies documenting amplified DR-TB following pandemic-related service interruptions, yet Iran’s trajectory exhibits unique features. 15 The dominance of M. simiae (55.63% of resistant NTM isolates) contrasts with neighboring countries where M. abscessus prevails, suggesting local environmental drivers like Tehran’s aging water infrastructure facilitating colonization.16,17 This convergence of anthropogenic and ecological factors underscores the complexity of containment in resource-variable settings. On the other hand, the serious clinical impacts of MDR-NTM infections, which are associated with a poor prognosis, should also be considered. Patients with MDR-NTM (particularly M. abscessus and M. avium complex) have been linked to significant mortality. A meta-analysis of 9,035 NTM patients reported an all-cause 5-year mortality of approximately 27%, and pulmonary M. avium complex disease alone accounted for nearly 49% of 5-year mortality in one large cohort. 18 These sobering outcomes underscore the urgency of the resistance patterns in patients with NTM-PD.
The disturbing prevalence of MDR-TB within AIIRs (18.22%, p < 0.0001) exposes critical flaws in infection control protocols. While these units are engineered for containment, our findings align with emerging evidence of AIIRs becoming amplification sites during staff shortages.19,20 Mechanical factors like negative pressure failures (documented in 43% of Iranian hospitals) combined with delayed diagnostic turnaround during respiratory infection surges create prolonged exposure windows.15,21 This transmission paradox extends to pulmonary medicine units where zoonotic M. bovis reservoirs (0.27% MDR prevalence, p < 0.0001) implicate unpasteurized dairy consumption among rural migrants—a pathway requiring integrated veterinary-human surveillance not addressed in current protocols.22,23
Gender disparities reveal equally complex etiologies. The male predominance in MDR-TB (26.01% vs 12.45%, p = 0.005) and M. kansasii infections (2.05% vs. 0.53%, p = 0.012) reflects Iran-specific sociocultural dynamics: mining sector exposures in Kerman province, stigma-driven health care avoidance, and androgen-mediated immune modulation. 24 Conversely, female-predominant M. abscessus (2.49% vs 2.76% male, p = 0.009) may correlate with rising beauty industry interventions, though this requires confirmation through exposure history mapping. These disparities cannot be addressed through biomedical interventions alone but require gender-responsive public health strategies accounting for occupational and health care-seeking behaviors.
Diagnostic limitations compound these challenges. The 38.14% smear negativity rate among culture-positive M. abscessus isolates exceeds WHO benchmarks, reflecting persistent reliance on outdated microscopy in peripheral clinics. 4 GW specimens (6.023% of samples) unexpectedly emerged as significant NTM reservoirs (M. genavense: 66.66%, M. gordonae: 50.0%), suggesting gastrointestinal colonization as an underrecognized transmission pathway.25,26 This finding necessitates revision of Iran’s pediatric TB guidelines, which prioritize gastric lavage without molecular confirmation. The temporal spike in negative cultures (21.43% in 2022) likely represents both improved specimen quality control and emerging resistance to decontamination protocols—a phenomenon documented in chlorine-adapted M. simiae strains.
The sample type analysis revealed pathogen-specific niches with public health implications: SF’s association with M. chelonae (5.55%, p < 0.05), while abscess prevalence of M. fortuitum (25.0%, p < 0.05) reflects inadequate sterile technique in OP procedures. 27 The concentration of M. abscessus in pediatric GW (88.13%) supports the aspiration-colonization hypotheses in children with silent aspiration. 26 These findings demand specimen-specific processing protocols currently absent in Iranian guidelines.
Despite promising declines in IMR (4.76–0.04%), the 2024 M. simiae surge (7.91%) signals an evolving threat landscape. Climate modeling suggests drought intensification in Central Iran may further concentrate environmental mycobacteria in water systems, while Afghanistan’s humanitarian crisis introduces novel strains through forced migration.28,29 These compounding factors necessitate climate-responsive surveillance beyond current passive case detection.
Several limitations temper interpretation; the single-reference-lab design limits generalizability despite national representation; unmeasured confounders like socioeconomic status may influence resistance patterns; and pediatric extrapulmonary samples were underrepresented. Future research must prioritize strain-level genomics to differentiate relapse from reinfection, household transmission studies for M. simiae, and operational research on rapid molecular deployment in peripheral clinics. Moreover, as a national referral center, the MDR-TB rates reported here likely overestimate the true population prevalence; community-based surveys would be needed to define the national burden fully.
Conclusion
Iran stands at an epidemiological crossroads where traditional TB control paradigms are insufficient against converging threats of MDR-TB resurgence, climate-adapted NTMs, and gender-skewed burdens. Containment requires precision interventions: (1) Redesigning AIIRs with UV-C cycle decontamination and continuous air monitoring; (2) Gender-specific screening in mining regions; (3) Water safety regulations targeting M. simiae in municipal systems; and (4) Mandatory molecular testing for smear-negative specimens. Only through integrated public health approaches that bridge clinical, environmental, and social determinants can Iran curb the escalating mycobacterial crisis.
Ethics Approval
All the procedures performed in the studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the Helsinki declaration and its later amendments or comparable ethical standards. The study protocol was approved by the Medical Ethics Committee of Shahid Beheshti University of Medical Sciences (SBMU), and ethical approval was obtained from the NRITLD with the number IR.SBMU.NRITLD.REC.1403.090.
Authors’ Contributions
A.A.V. and P.F. conceived the article and revised it. J.G. and P.F. controlled and managed the quality of clinical and laboratory data. S.S.M. and M.M. performed the statistical analyses, prepared tables and figures, and wrote the article. All authors participated in preparing the final draft of the article, revised the article, and critically assessed the academic content. All authors have read and approved the article’s content and confirmed the accuracy or integrity of any part of the work.
Footnotes
Acknowledgments
The authors wish to thank all their colleagues in the National Tuberculosis Reference Laboratory at Masih Daneshvari Hospital, National Research Institute of Tuberculosis and Lung Diseases (NRITLD), WHO Collaborating Center (WHOCC), Mycobacteriology Research Center (MRC), Tehran, Iran. They appreciate their coworkers and patients who kindly contributed to this research.
Consent to Participate
Informed consent was obtained from all individual participants included in the study.
Disclosure Statement
The authors declare no conflict of interest and have no relevant financial or non-financial interests to disclose.
Funding Information
The authors declare that no funds, grants, or other support were received during the preparation of this article.