
Abstract
Background:
The increasing prevalence of multidrug-resistant Pseudomonas aeruginosa limits treatment options and highlights the need for new antimicrobials. Although agents such as ceftazidime–avibactam (CZA), ceftolozane–tazobactam, imipenem–relebactam, and cefiderocol have expanded therapeutic choices, resistance to these antibiotics is also emerging. Combination therapies therefore remain an important strategy. This study evaluated the in vitro synergistic activity of the CZA–colistin (COL) combination in carbapenem-resistant P. aeruginosa (CRPA) isolates.
Methods:
Twelve clinical CRPA isolates obtained from Kocaeli University Hospital (2021–2022) were included. Minimum inhibitory concentration values were determined by broth microdilution, and synergy was assessed using the checkerboard method. Synergy categories were defined as follows fractional inhibitory concentrations index (FICI) ≤ 0.5 synergism, 0.5 < FICI ≤ 1.0 partial synergism, 1.0 < FICI ≤ 4.0 indifference, and >4.0 antagonism.
Results:
All isolates were susceptible to colistin, whereas four were resistant to CZA. Checkerboard analysis showed partial synergy in nine of 12 isolates (75.0%), with no antagonism detected. Partial synergy was more frequent in CZA-resistant isolates (100%) than in CZA-susceptible ones (62.5%) and was notably associated with isolates carrying blaNDM and blaOXA-48.
Conclusions:
CZA–COL combination may offer partial synergy, especially in CZA-resistant CRPA strains; however, broader in vivo and prospective studies are needed to support clinical use.
Introduction
Pseudomonas aeruginosa is an aerobic, non-spore-forming, Gram-negative opportunistic bacterium that is widely distributed in the environment, including water (particularly in the freshwater sources), soil, and animals. In urban settings, it can persist in reservoirs such as hot tubs, whirlpools, and swimming pools. P. aeruginosa can cause a wide variety of community-acquired and nosocomial infections. These infections include folliculitis, osteomyelitis, otitis externa, urinary tract infections, ventilator-associated pneumonia, bacteremia, central catheter-associated bloodstream infections, septicemia, and catheter-associated urinary tract infections. Hospital reservoirs include drinking water systems, faucets, sinks, toothbrushes, ice machines, disinfectant solutions, soap, respiratory therapy equipment, endoscopes, and associated reprocessing devices. P. aeruginosa infections carry a higher risk of mortality and morbidity compared to other pathogens.1–3 In its 2024 report, the World Health Organization included P. aeruginosa in the list of “critical priority pathogens,” emphasizing the need to develop new therapeutic strategies. 4
β-Lactam antibiotics, such as penicillins, cephalosporins, monobactams, and carbapenems, remain the first-line agents for treating P. aeruginosa infections. However, widespread use of these drugs has led to the emergence of β-lactam-resistant strains, significantly increasing both the cost of treatment and the risk of mortality. 5 P. aeruginosa has a remarkable ability to develop antimicrobial resistance through chromosomal mutations, particularly the acquisition of β-lactamase genes, including carbapenemases. This, combined with other resistance mechanisms, has led to an increased incidence of multi-drug resistant (MDR) strains. 6
Multiple mechanisms contribute to carbapenem resistance in P. aeruginosa, including the production of metallo-β-lactamases (e.g., blaVIM, blaNDM), class D β-lactamases (e.g., blaOXA-48), porin loss (particularly OprD), efflux pump overexpression, and AmpC β-lactamase induction. 7 Moreover, resistance to newly introduced β-lactam/β-lactamase inhibitor combinations has also been reported in recent years.8–11 Resistance to ceftazidime–avibactam (CZA) is primarily associated with the production of metallo-β-lactamases (notably blaVIM and blaIMP), mutations/derepression of the chromosomal AmpC (PDC) β-lactamase that reduce susceptibility to avibactam, and, in some cases, alterations in penicillin-binding proteins (e.g., PBP3) or porin/efflux changes.12–15 Resistance to ceftolozane-tazobactam (C/T) is largely explained by AmpC overexpression or structural mutations, with increased efflux pump activity also contributing.16,17 Imipenem-relebactam (I/R) resistance is often linked to carbapenemase production (especially MBLs), OprD porin loss, and overexpression of efflux pumps such as MexAB-OprM.18,19 Reported resistance mechanisms for cefiderocol (FDC) include, in some isolates, mutations in iron transport genes, carbapenemase production, porin alterations, and increased efflux. 20
These findings indicate that the use of new antibacterial agents as monotherapy may carry a risk for resistance development. Therefore, combination therapies have emerged for the treatment of MDR and extensively drug-resistant (XDR) P. aeruginosa isolates. Combinations of colistin (COL) with newer β-lactam/β-lactamase inhibitors have gained importance. However, data on the combination of CZA with COL remain limited, highlighting the need for further experimental studies. 21
Investigating the synergistic efficacy of the CZA-COL combination against carbapenem-resistant P. aeruginosa (CRPA) isolates may contribute to treatment options for resistant infections.
The aim of this study is to evaluate the in vitro synergistic activity of CZA and COL against clinical carbapenem-resistant CRPA isolates, providing experimental evidence to support combination therapy strategies.
Materials and Methods
This study was designed as a retrospective study and approved by the Kocaeli University Non-Interventional Clinical Research Ethics Committee (Approval No: 2025/21/23). All procedures were conducted in accordance with the ethical standards of the Declaration of Helsinki. As this study was based on existing microbiological and clinical records without patient identifiers, no informed consent was required.
The study included 12 CRPA isolates obtained from various clinical samples (urine, sputum, wound, tracheal aspirate, and bronchoalveolar lavage) of patients in Kocaeli University Hospital wards and intensive care units between 2021 and 2022. The isolates were selected to represent different resistance phenotypes and genotypic characteristics, including CZA-resistant and CZA-susceptible isolates, different carbapenemase gene profiles (blaVIM, blaNDM, blaOXA-48), and class-1 integron positivity. All isolates had previously been characterized for carbapenem resistance genes and class-1 integrons. 7
PCR-positive P. aeruginosa isolates for blaOXA-48, blaVIM, and blaNDM were selected for sequence-level confirmation. PCR amplicons were purified and sequenced using an Oxford Nanopore Technologies (ONT)-based amplicon sequencing workflow. Raw reads were basecalled, demultiplexed when applicable, and quality-filtered. High-quality reads corresponding to each target amplicon were used to generate consensus sequences. Consensus amplicon sequences were compared with representative carbapenemase gene reference sequences deposited in NCBI GenBank using BLASTn. The blaOXA-48, blaVIM, and blaNDM amplicons obtained in this study showed significant similarity to the corresponding reference carbapenemase gene sequences, including blaOXA-48 (AY236073.2), blaVIM-2 (AF191564.1) and blaNDM-1 (FN396876.1). Sequence confirmation was accepted when BLASTn analysis showed ≥99% nucleotide identity, ≥95% query coverage, and an E-value of 0.0 or ≤1 × 10−100 against the expected carbapenemase gene target.22,23
Broth microdilution tests were performed to determine the minimum inhibitory concentrations (MICs) of CZA and COL. CZA and COL were obtained from commercially available pharmaceutical formulations and prepared according to the manufacturer’s instructions under sterile conditions before use in the MIC and checkerboard assays. Ninety-six-well U-bottom sterile microplates and cation-adjusted Mueller Hinton Broth were used. One well was used for sterility control and one well for growth control. Escherichia coli ATCC 25922 was used as the control strain. Microplates were incubated at 37°C for 18–24 hours. The lowest concentration at which no visible growth occurred was determined as the MIC. Susceptibility interpretations (susceptible/resistant) were performed according to the EUCAST clinical breakpoints (Version 5.0, 2024). 24 The experiments were repeated three times.
The synergistic effect of CZA-COL combinations was investigated using the checkerboard method. Checkerboard assay concentrations were prepared individually for each isolate based on the MIC values previously determined by the broth microdilution method. For synergy testing, dilutions ranging from 2–3 times higher to 4–8 times lower than the MIC values were studied, corresponding to concentration ranges of 0.125–128 μg/mL for CZA and 0.125–8 μg/mL for COL. During the checkerboard assay, MIC values of each antibiotics in combination were re-evaluated within the checkerboard matrix, and FICI calculations were performed using these combination-associated MIC values.
In microplates, the first row and first column were used to evaluate CZA and COL MICs, and the other rows and columns were used to evaluate combinations. The A1 well was used as a growth control. Microplates were incubated at 37°C for 16–20 hours. For each combination, fractional inhibitory concentrations (FICs) and the FIC index (FICI) were calculated separately. FICI ≤ 0.5 was interpreted as synergism, 0.5 < FICI ≤ 1.00 as partial synergism, 1.00 < FICI ≤ 4.0 as indifference, and FICI > 4.0 as antagonism. 25 The experiments were repeated three times, and the mean FICI values obtained from the repeated experiments were used as the final reported values.
Results
The resistance rates of CRPA isolates to other antibiotics were as follows: AK (n = 5, 41.7%), ATM (n = 6, 50%), FEP (n = 7, 58.3%), CAZ (n = 5, 41.7%), CIP (n = 8, 66.7%), LVX (n = 8, 66.7%), NET (n = 9, 75%), TZP (n = 7, 58.3%), PRL (n = 8, 66.7%), TOB (n = 5, 41.7%), CN (n = 6, 50%), and TE (n = 8, 66.7%). Detailed antibiotic susceptibility profiles of the isolates are presented in Table 1.
Clinical Characteristics and Antibiotic Susceptibility Profiles of CRPA Isolates Included in the Study
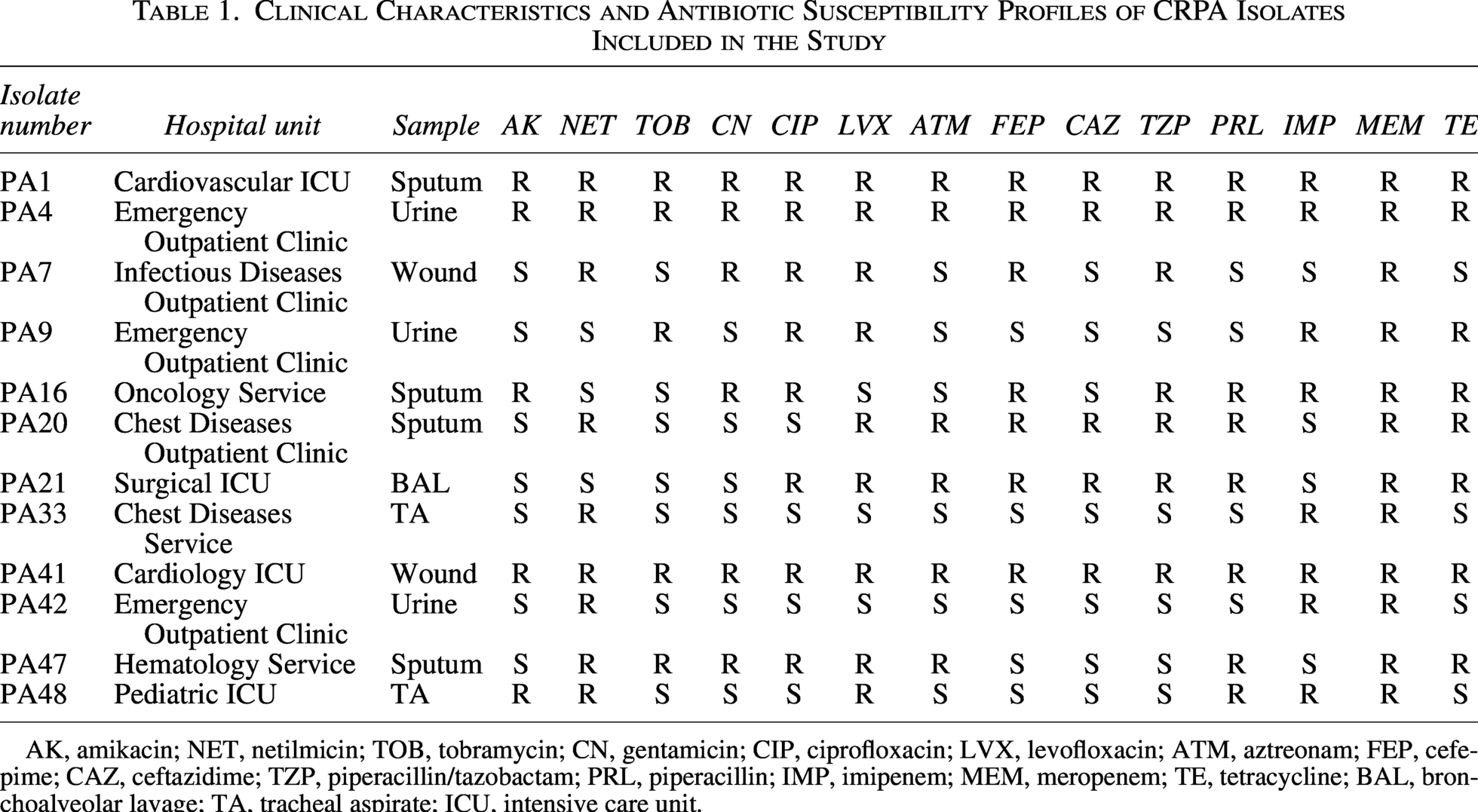
AK, amikacin; NET, netilmicin; TOB, tobramycin; CN, gentamicin; CIP, ciprofloxacin; LVX, levofloxacin; ATM, aztreonam; FEP, cefepime; CAZ, ceftazidime; TZP, piperacillin/tazobactam; PRL, piperacillin; IMP, imipenem; MEM, meropenem; TE, tetracycline; BAL, bronchoalveolar lavage; TA, tracheal aspirate; ICU, intensive care unit.
All isolates were susceptible to COL. Four isolates were resistant to CZA. Three isolates that were resistant to CZA and also carbapenemase gene (blaVIM, blaNDM, and blaOXA-48) positive contained class-I integrons. These integrons contained gene cassettes carrying resistance genes (Table 2). All PCR-positive blaOXA-48, blaVIM, and blaNDM amplicons yielded analyzable consensus sequences. BLASTn comparison demonstrated ≥99% nucleotide identity and ≥95% query coverage with the corresponding reference carbapenemase gene sequences in NCBI GenBank. Accordingly, all PCR-positive carbapenemase gene targets were confirmed by amplicon sequencing.
FICI Values of CZA-COL Combination and Phenotypic and Genotypic Characteristics of Isolates
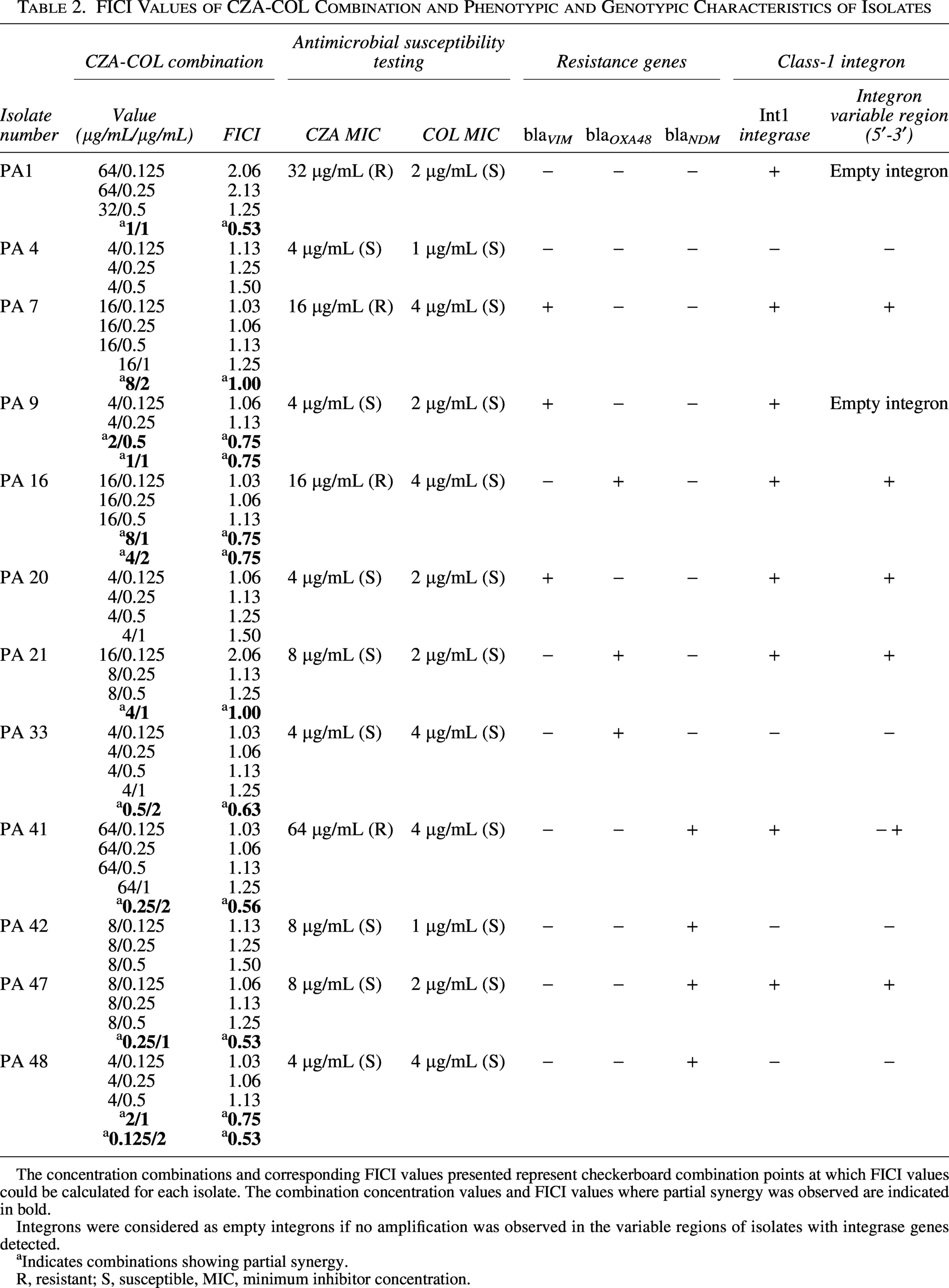
The concentration combinations and corresponding FICI values presented represent checkerboard combination points at which FICI values could be calculated for each isolate. The combination concentration values and FICI values where partial synergy was observed are indicated in bold.
Integrons were considered as empty integrons if no amplification was observed in the variable regions of isolates with integrase genes detected.
Indicates combinations showing partial synergy.
R, resistant; S, susceptible, MIC, minimum inhibitor concentration.
MIC values determined by the broth microdilution method ranged from 0.25 to 256 μg/mL for CZA and from 0.25 to 16 μg/mL for COL.
Among the 12 CRPA isolates included in the study, partial synergy was observed in nine (75.0%) isolates, while no difference was observed in three (25.0%) isolates. No antagonistic effect was detected. A partial synergistic effect was detected in all four CZA-resistant isolates (100%), whereas this rate was 62.5% among CZA-susceptible isolates.
Specifically, partial synergy was detected in one of three isolates carrying the blaVIM gene, two of three isolates carrying the blaOXA-48 gene, and three of four isolates carrying the blaNDM gene. Partial synergy was also observed in two isolates, both resistant to CZA and carrying either the blaOXA-48 or blaNDM genes (Table 2). Partial synergy was observed in the 0.25/2 combination in the isolate (PA41) carrying the blaNDM gene and the class-1 integron gene cassette (Fig. 1). The FICI values of CZA-COL combination are shown in Table 2.
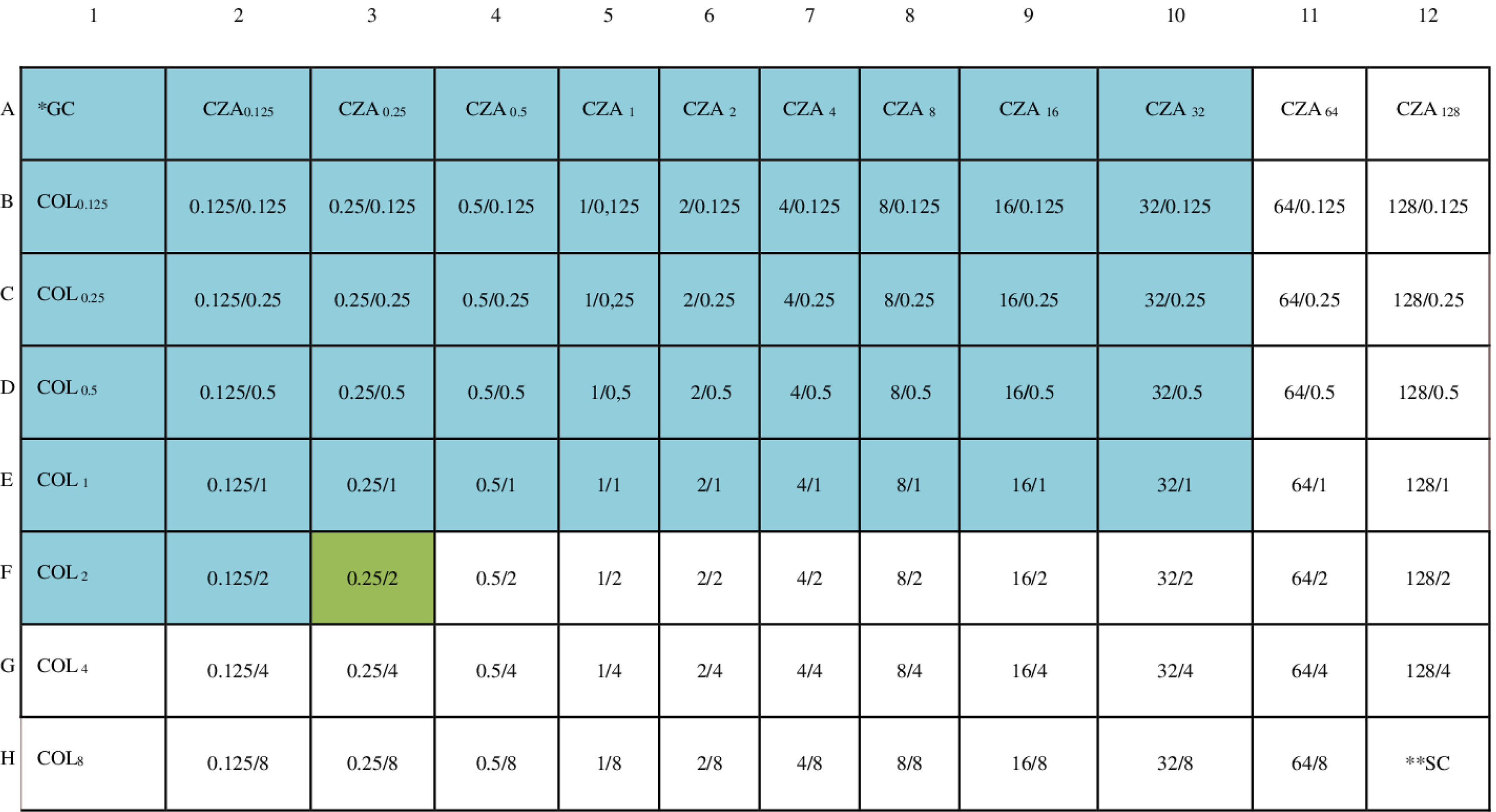
Schematic representation of the in vitro synergism of the CZA-COL combination against the CRPA isolate (PA41) in the checkerboard panel. *GC, Growth Control. **SC, Sterility Control. The numbers in the first row indicate the concentration of CZA in that column in μg/mL, the numbers in the first column indicate the concentration of COL in that row in μg/mL, and the blue areas indicate the wells where growth was observed. Green areas indicate partial synergy.
Discussion
The increasing prevalence of antibiotic resistance, particularly in infections caused by multidrug-resistant Gram-negative bacteria, has become a serious global health problem.26,27 P. aeruginosa is one of the most difficult pathogens to treat due to its inherent resistance mechanisms and easily acquired resistance genes. 26 The widespread prevalence of carbapenem resistance limits treatment options, increases mortality rates, and necessitates the development of new treatment strategies. Current guidelines recommend monotherapy and combination therapy for CRPA infection. However, these guidelines lack a specific method and sufficient evidence to support treatment recommendations. The increase in CRPA isolates necessitates the investigation of alternative treatment options. 3
Combination therapies are gaining importance to increase the effectiveness of existing antibiotics and delay the development of resistance; in particular, the potential synergistic benefits of “last-resort” agents such as CZA-COL are being investigated.28,29 Combination approaches should be evaluated clinically and in vitro to limit the use of single antibiotics with high toxicity risk, such as colistin.3,28
The synergistic effect of various antibiotic combinations against P. aeruginosa isolates was investigated and varying degrees of synergy were reported. In the study by Dundar and Otkun, β-lactam–aminoglycoside combinations demonstrated the highest synergistic activity (ceftazidime–tobramycin: 67%, piperacillin/tazobactam–tobramycin: 50%), whereas carbapenem–aminoglycoside (imipenem–tobramycin) and carbapenem–fluoroquinolone (imipenem–ciprofloxacin) combinations showed no synergy. Ghorbani et al. reported moderate to high synergy rates for aminoglycoside–cephalosporin (amikacin–ceftazidime: 55.6%), aminoglycoside–polymyxin (tobramycin–colistin: 58.3%), and cephalosporin–polymyxin (ceftazidime–colistin: 52.8%) combinations. In the study by Ma et al. (2019), cephalosporin–β-lactamase inhibitor combinations (ceftazidime–cefoperazone/sulbactam: 100%) produced the strongest synergistic effect against carbapenem-resistant isolates, while other β-lactam- or polymyxin-based combinations exhibited lower levels of synergy or additive effects.30–32
The combination of CZA and COL has been evaluated as a potential treatment option, particularly for MDR or CRPA infections. It has been reported in the literature that this combination may exhibit synergistic effects in vitro against both CZA-sensitive and CZA-resistant strains.
In a study of 21 XDR P. aeruginosa isolates, seven were resistant to CZA, and six of these carried carbapenemase genes (e.g., blaVIM, blaGES, blaOXA-2). Using the time-kill method, the CZA–COL combination showed synergy in six CZA-resistant and six CZA-susceptible isolates. The authors noted that the combination can be effective in both resistant and susceptible isolates, although its efficacy may vary depending on resistance mechanisms. Overall, combination therapy was considered more beneficial than monotherapy against XDR P. aeruginosa. 21 Mikhail et al. evaluated the CZA–COL combination in 21 P. aeruginosa isolates. Among five CZA-resistant isolates, three carried acquired β-lactamase genes (blaIMP-48, blaOXA-10, blaCARB-2). The combination showed limited activity in four resistant strains and strong activity in one. Similar MIC reductions were observed in CZA-susceptible isolates. Time-kill assays performed on two CZA-susceptible isolates showed synergy in one (with AmpC overexpression and OprD loss), but not in the other (AmpC overexpression alone). The authors highlighted that although the CZA–COL combination can produce bactericidal synergy, its effectiveness depends on underlying resistance mechanisms. They also noted that avibactam is ineffective against MBL-producing strains (e.g., blaIMP), limiting the combination’s usefulness. Overall, they concluded that such combinations may serve as potential therapeutic options but require further investigation. 33 These findings, compared with our study, suggest that stronger MIC reductions were observed in resistant isolates, but methodological differences (checkerboard and time-killing) played an important role in the results.
In another study involving 16 XDR P. aeruginosa isolates, the synergistic effect of the CZA–COL combination was assessed using the checkerboard method. Among the CZA-resistant isolates, blaIMP and blaVIM genes were detected in three (18.75%). Synergy was observed in four isolates (25%). An in vivo biofilm model was also established, and the effect of the CZA–COL combination was evaluated using time-kill assays. In these assays, the combination reduced the planktonic bacterial load of three XDR isolates and showed greater decreases compared to monotherapy. The researchers concluded that combining CZA and COL can enhance the bactericidal activity of each drug and simultaneously inhibit biofilm formation. Therefore, they emphasized that the CZA–COL combination represents a potential treatment option for XDR P. aeruginosa infections. 34 Similarly, our findings demonstrated notable in vitro partial synergistic activity of the CZA–COL combination, particularly among CZA-resistant isolates, while no antagonistic interaction was detected. At the genetic level, partial synergy was observed in one-third of blaVIM-carrying isolates, two-thirds of blaOXA-48-carrying isolates, and three-fourths of blaNDM-carrying isolates. Although blaOXA-48 is less commonly reported in P. aeruginosa compared with metallo-β-lactamases such as blaVIM and blaNDM, blaOXA-48-producing P. aeruginosa isolates and co-existence of blaOXA-48 with other carbapenemase genes have previously been reported in Türkiye, particularly among high-risk ST235 clones. 35 In our study, all blaOXA-48-positive isolates were additionally confirmed by ONT-based amplicon sequencing and BLASTn analysis, supporting the accuracy of molecular identification. These findings may reflect local epidemiological diversity in carbapenem resistance mechanisms among CRPA isolates.
Class-1 integrons were included in the study because of their well-recognized clinical and epidemiological importance in the acquisition and dissemination of antimicrobial resistance genes among P. aeruginosa isolates, particularly in MDR and CRPA strains. Awareness of integron-associated resistance determinants may contribute to better understanding of resistance dissemination patterns and may also be clinically relevant for optimizing antimicrobial treatment strategies. 36 In the present study, partial synergy findings appeared to be more frequent among integron-positive isolates; however, because of the limited number of isolates, a definitive correlation between integron carriage and synergy outcomes could not be established. Nevertheless, evaluation of class-1 integrons provided additional epidemiological and clinically relevant information regarding the resistance background, resistance gene dissemination potential, and genetic adaptability of the isolates included in this study.
Partial synergy was detected in the blaNDM and class-1 integron-carrying PA41 isolate at a 0.25/2 μg/mL combination. These observations demonstrate the potential efficacy of CZA combinations, particularly against CRPA isolates resistant to CZA and carrying certain carbapenemase genes. Moreover, the partial synergy rates in the current study were higher than those reported by Mei et al., 34 suggesting that different genetic determinants and antibiotic combinations may influence synergy profiles. Overall, the data indicate that CZA combinations could represent a promising option for the treatment of XDR/CRPA isolates.
In the study by Mataracı Kara et al., 40 colistin-nonsusceptible MDR P. aeruginosa isolates were examined. CZA resistance was detected in 47.5% (19/40). Among all isolates, blaOXA-48 was found in 20% (8/40); various other carbapenemase genes (blaNDM, blaOXA-23, blaOXA-51, blaKPC, blaIMP) in 12.5% (5/40); blaVIM in 5% (2/40); and blaIMP alone in one isolate. Synergy testing using time-kill assays showed that the CZA–COL combination was synergistic in 4 of 6 isolates (66.6%) at standard concentrations and in 5 isolates (83.3%) at 4× MIC. No antagonism was detected. Synergy was especially notable in blaOXA-48 - positive and carbapenemase-negative isolates, but it was not sustained in blaIMP-positive strains. The authors concluded that the CZA–COL combination may serve as an alternative treatment option for colistin-nonsusceptible and CZA-resistant P. aeruginosa. They also highlighted that lowering the colistin dose could help reduce the risk of nephrotoxicity. 37 In our study, partial synergy, rather than synergy, was observed. Efficacy may vary depending on the testing method used and the resistance mechanisms of the isolates.
In studies evaluating the clinical efficacy of CZA-COL combination, the rates of P. aeruginosa isolates were examined in different patient groups. In a series of 31 patients reported by Zheng et al., the CRPA rate was found to be 5 patients (16.1%); it was reported that CZA-COL treatment in this group provided promising results in terms of clinical improvement and microbiological eradication, and no antagonistic effect was observed. 38 In a cohort study of 95 patients conducted by Zhang et al., CRPA was isolated in 18 patients (18.9%), and the CZA-COL combination was found to be particularly advantageous in terms of clinical improvement and reduced rates of sepsis and septic shock in severe respiratory infections. 39 These data demonstrate that the CZA-COL combination provides clinically promising results also in this subgroup.
The clinical applicability of the observed in vitro synergy findings should also be evaluated in terms of pharmacokinetic/pharmacodynamic (PK/PD) compatibility. Current IDSA guidance recommends CZA at a dose of 2.5 g every 8 hours administered by prolonged infusion for severe multidrug-resistant Gram-negative infections. 40 In addition, international consensus guidelines for polymyxins suggest target steady-state plasma COL concentrations of approximately 2 mg/L, while emphasizing that higher exposures may substantially increase nephrotoxicity risk. 41 In the present study, partial synergy was observed at concentration combinations that were generally within or close to clinically achievable exposure ranges for both CZA and COL according to current dosing recommendations and PK/PD data. However, the clinical relevance of checkerboard-based synergy findings may vary depending on achievable serum and tissue concentrations, particularly in critically ill patients. Therefore, although the observed in vitro interactions are promising, their direct clinical applicability should be interpreted cautiously and confirmed by further PK/PD-supported experimental and clinical studies.
Another important point is that partial synergy findings obtained by checkerboard assays may be influenced by the inherent variability of broth microdilution methods, particularly for FICI values close to interpretive thresholds. In broth microdilution-based synergy testing, a ± 1 dilution difference is generally regarded as an acceptable methodological variation; therefore, some partial synergy findings may also fall within the indifference category. In the present study, partial synergy was observed in a substantial proportion of isolates, whereas no complete synergistic effect was detected. If complete synergy had been demonstrated, stronger conclusions regarding the therapeutic potential of the combination could have been made. Therefore, the observed partial synergy findings should be interpreted cautiously and primarily as descriptive in vitro observations rather than definitive pharmacodynamic evidence of synergy.
This study has several limitations. The number of CRPA isolates included was relatively limited, and therefore the findings should be interpreted primarily as descriptive in vitro data. Non-enzymatic resistance mechanisms, including porin loss and efflux pump overexpression, were not investigated and may also have influenced the observed synergy findings. Checkerboard assays may show methodological variability, particularly for FICI values close to interpretive thresholds. Commercially available pharmaceutical formulations rather than standard analytical-grade antibiotic powders were used in the experiments, which may have influenced the in vitro activity results to some extent.
Conclusion
In conclusion, the CZA-COL combination demonstrated partial synergistic activity against a substantial proportion of CRPA isolates, including CZA-resistant strains. The absence of antagonism suggests that this combination may represent a potentially useful therapeutic option for difficult-to-treat CRPA infections. Nevertheless, the findings should be interpreted cautiously because of the descriptive in vitro nature of the study and the inherent variability of checkerboard-based synergy testing. Further experimental, PK/PD-supported, in vivo, and prospective clinical studies are needed to better clarify the clinical applicability, safety profile, and therapeutic potential of the CZA–COL combination against CRPA infections.
Authors’ Contributions
G.A. and D.D.: Conceptualization, methodology, data analysis, data interpretation, and investigation. G.A.: Data curation, writing—original draft. D.D.: Supervision, writing—review and editing.
Footnotes
Ethical Approval
This study was designed as a retrospective study and approved by the Kocaeli University Non-Interventional Clinical Research Ethics Committee (Approval No: 2025/21/23).
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.