
Abstract
We study curious dynamical patterns appearing in networks of one ring of cells coupled to a ‘buffer’ cell. Depending on how the cells in the ring are coupled to the ‘buffer’ cell, the full network may have a nontrivial group of symmetries or a nontrivial group of ‘interior’ symmetries. This group is
Keywords
1. Introduction
Golubitsky, Stewart, and co-authors have developed, in recent years, a theory for networks of coupled cells (Golubitsky et al., 2004a, 2005; Golubitsky and Stewart, 2006). These networks are schematically identified with directed graphs, where the nodes (cells) represent dynamical systems, and the arrows indicate the couplings between them.
Networks have applications in many areas of science, from biology, economy, ecology, neuroscience and computation, to physics (Boccaletti et al., 2006; Pinto and Golubitsky, 2006; Antoneli et al., 2010; Michiels, 2010; Mahmoud and Ahmed, 2011; Pinto, 2012; Mahmoud et al., 2013; Zhu et al., 2013; Gong et al., 2014). Particular attention has been given to patterns of synchrony (Pikovsky et al., 2001; Aguiar et al., 2011), phase-locking modes, resonance, and quasiperiodicity (Antoneli et al., 2008, 2010; Pinto, 2012).
In this paper, we study strange patterns produced by the networks in Figure 1. We were motivated by the work of Antoneli et al. (2007). There, the authors give the full analogue of the equivariant Hopf theorem for networks with ‘interior’ symmetries, extending previous results by Golubitsky et al. (2004b). They obtain states whose linearizations, on certain subsets of cells, near bifurcation, are superpositions of synchronous states with states having spatiotemporal symmetries. The equivariant Hopf theorem by Golubitsky et al. (1988) proves the existence of certain branches of spatiotemporal symmetry-breaking time-periodic states in systems with exact symmetry groups. In addition, Antoneli et al. (2007) simulate the dynamical patterns produced by the coupled cell network in Figure 1(b), using two-dimensional internal cell dynamics for the cells in the ring. Here, we use more complex internal dynamics for each cell, namely, the Chen oscillator. We simulate the networks in Figure 1 and obtain the solutions with spatiotemporal symmetries, as in the case of Antoneli et al. (2007), and more complex features, such as quasiperiodicity and chaos. The role of the ‘buffer’ cell can be viewed in the sense of dynamical systems where a master–slave relation is observed and where phenomena like synchronization and chaotic features are studied (Lin et al., 2005; Matias et al., 2011; Boulkroune and Msaad, 2012, and references therein).
Networks of one ring of cells coupled to a ‘buffer’ cell. Each node represents a cell or a dynamical system. The arrows indicate the couplings between them.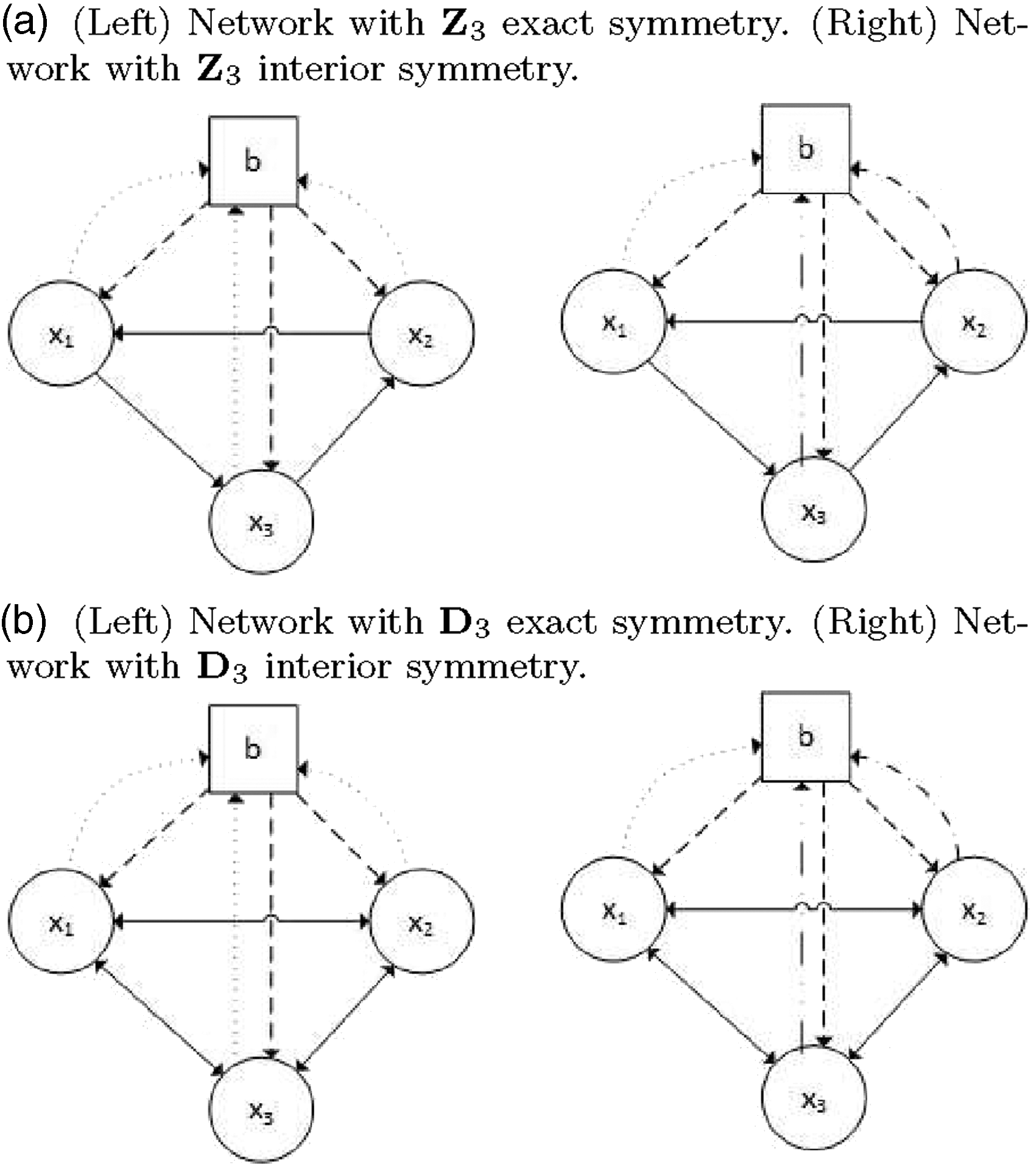
In Section 2, we briefly review the theory of coupled cell networks and bifurcation theory for symmetric dynamical systems. In Section 3, we simulate the coupled cell systems associated with the networks in Figure 1. In Section 4, we conclude this work and unveil future research directions.
2. Coupled cells and symmetry
A network of cells may be drawn as a directed graph, whose nodes represent the cells and whose edges distinguish couplings between them. Cells and arrows may be classified according to certain types (Stewart et al., 2003; Golubitsky et al., 2005). Cells of the same type have the same internal dynamics, and arrows with the same label identify equal couplings. Each cell is a dynamical system. The input set of a cell is the set of edges directed to that cell. Input isomorphisms are label-preserving bijections between input sets of cells. These isomorphisms obey the local symmetries of the network. These local symmetries have the structure of a groupoid. The latter means that they have the properties of a group, even though some products of elements may not always be defined. Figure 1 depicts four examples of coupled cell networks, where the nodes are drawn as circles (cells in the rings) and squares (‘buffer’ cell). In Figure 1 (left) there are three different types of couplings in the two networks, whereas in Figure 1 (right) there are five different types of couplings in the two networks.
Coupled cell systems are dynamical systems consistent with the architecture or topology of the graph representing the network. Each cell cj of the network has an internal phase space Pj: here we always assume that Pj is a Euclidean space
A vector field f on P is called admissible, for a given network, if it satisfies the two following conditions (Golubitsky and Stewart, 2006):
Domain condition: each component fj corresponding to a cell cj is a function only of the variables associated with the cells ck that have edges directed to cj; Pull-back condition: two components fj and fk corresponding to cells cj and ck with isomorphic input sets are identical up to a suitable permutation of the relevant variables.
In this work, we consider an important class of networks, namely, networks that possess a group of symmetries, either exact or ‘interior’ symmetry. A symmetry of a coupled cell system is the group of permutations of the cells (and arrows) that preserves the network structure (including cell labels and arrow labels) and its action on P is by permutation of cell coordinates. It is thus a transformation of the phase space that sends solutions to solutions. The networks in Figure 1(a) are examples of networks with
The notion of ‘interior’ symmetry was introduced by Golubitsky et al. (2004b), and it generalizes the usual definition of symmetry. The network in Figure 1(a), right, is a network with interior
2.1. Bifurcations in coupled cell systems
A bifurcation of a coupled cell system is associated with a change in the qualitative behavior of that system, when a parameter is varied. Generically, there are two local bifurcations from a synchronous equilibrium: the synchrony-breaking bifurcation and the synchrony-preserving bifurcations. The first bifurcation is characterized by a loss of symmetry of the solution when the bifurcation occurs. These bifurcations may be viewed as generalizations of symmetry-breaking bifurcations in symmetric coupled cell systems. The equivariant branching lemma proves the existence of certain branches of symmetry-breaking steady states (Golubitsky et al., 1988). The equivariant Hopf theorem guarantees the existence of families of small-amplitude periodic solutions bifurcating from the origin (Golubitsky et al., 1988). Generalizations of the equivariant branching lemma and the equivariant Hopf theorem for coupled cell systems with interior symmetries are given by Golubitsky et al. (2004b) and Antoneli et al. (2007).
We can produce a list of all spatio-temporal patterns satisfying the conditions of the above results and that are expected to occur when one of the coupled cell systems associated with the networks of Figure 1 undergoes an (interior) symmetry-breaking Hopf bifurcation.
Returning to the example with
Considering the networks in Figure 1(a), when the symmetry is exact the periodic solution of type
In the second case (Figure 1(b)), there are three solutions corresponding to symmetry types
3. Numerical simulations
The coupled cell systems associated with the networks depicted in Figure 1 are simulated. We use XPPAUT (Ermentrout, 2006) and MATLAB (http://www.mathworks.com) to numerically compute the relevant states. We consider the Chen oscillator as the internal dynamics of each cell in the 3-ring and a unidimensional phase space for the ‘buffer’ cell. The total phase space is thus ten-dimensional. The dynamics of a singular ring cell are given by (Chen and Ueta, 1999; Lu et al., 2002)
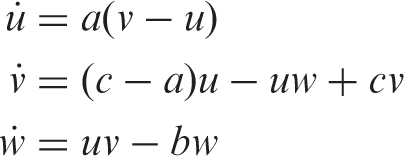
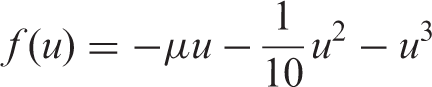
The coupled cell system of equations associated with the networks in Figure 1 is given by
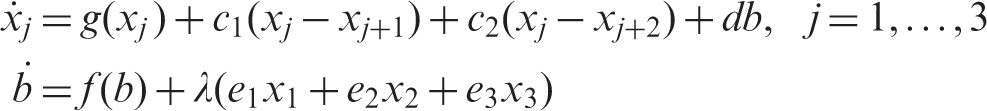
The system has exact symmetry for λ = 0 (networks in Figure 1 (right)) and ‘interior’ symmetry for λ ≠ 0 (networks in Figure 1 (left)). If c2 = 0 then the coupled cell system is admissible for the networks in Figure 1(a) with
3.1. Unidirectional networks: dynamics
We vary parameter c ∈ [15, 27], going from lower to higher values, and start from a steady state of the whole system.
We consider c2 = 0: this means that we will simulate the coupled cell systems associated with the unidirectional rings (Figure 1(a)), for λ = 0 and λ ≠ 0.
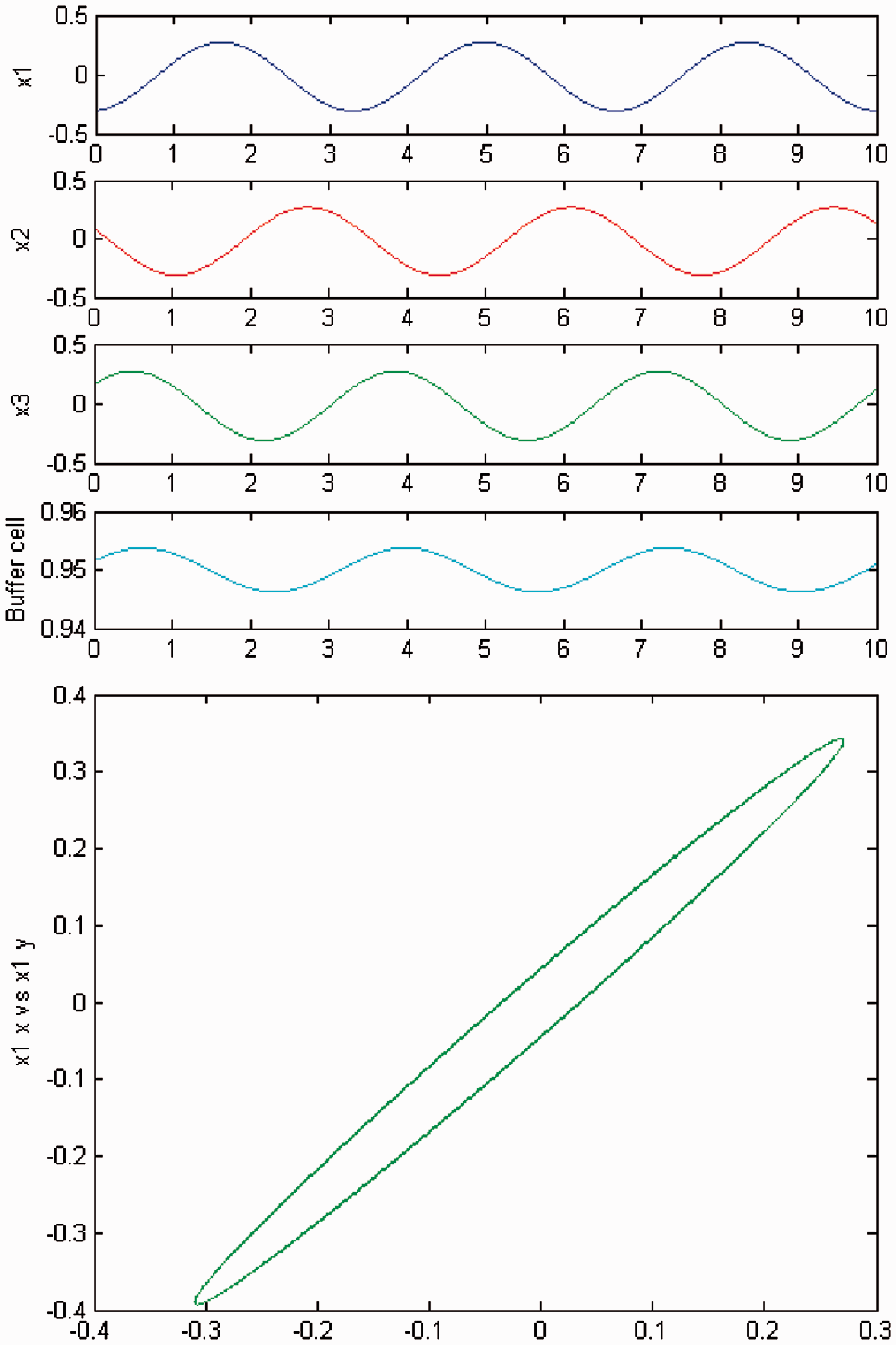
We increase c and a primary Hopf bifurcation occurs (HB1). In Figures 2 and 3, we plot (top) the time series of the solution of the coupled cell system (3), and (bottom) the phase space of the oscillator x1 of the ring. The solution is a Simulation of the coupled system (3) for λ = 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 15.59, after the first Hopf bifurcation point (HB1). The cells in the ring exhibit a Simulation of the coupled system (3), for λ ≠ 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 15.59, after the first Hopf bifurcation point (HB1). The cells in the ring depict a 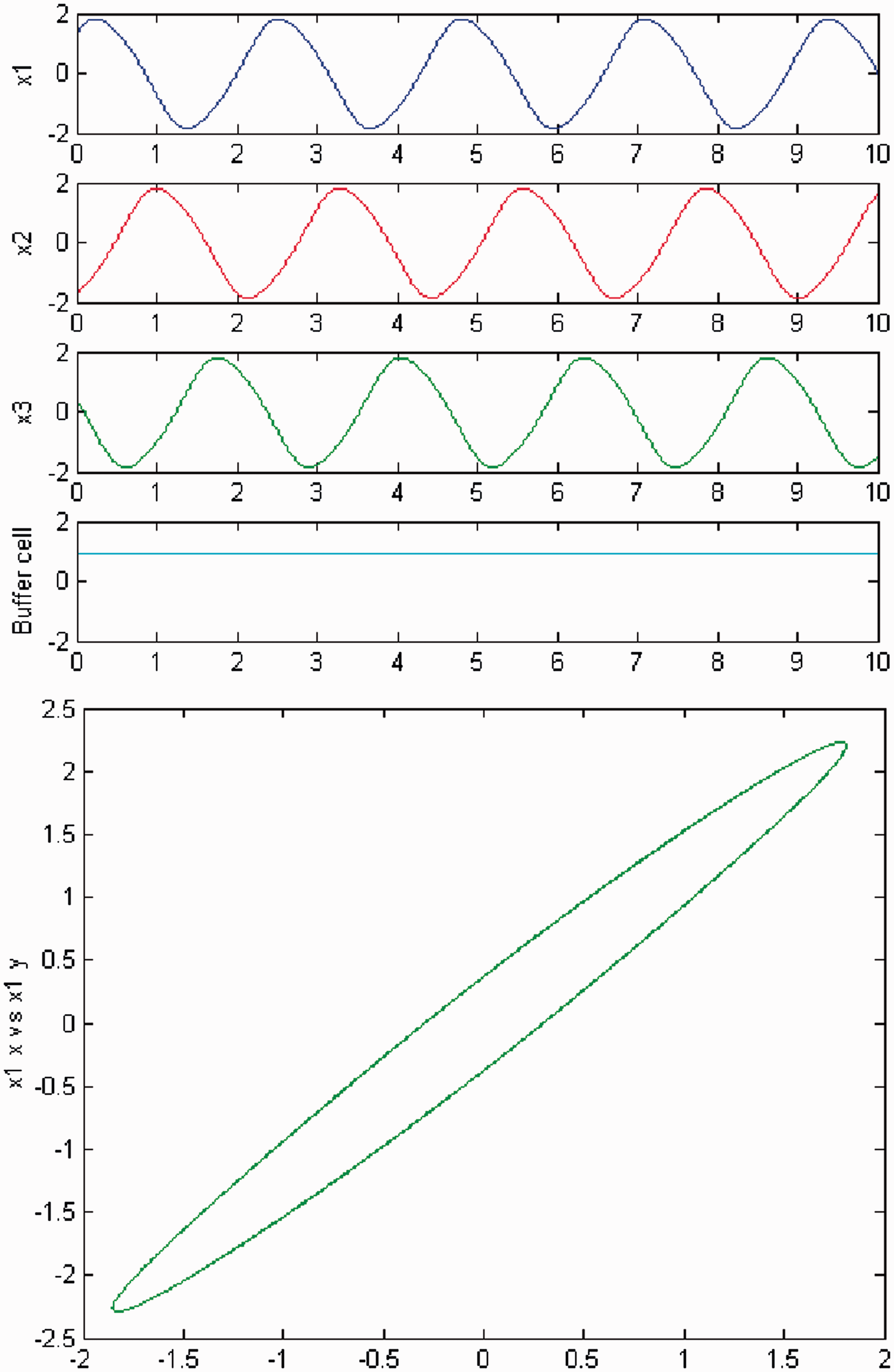
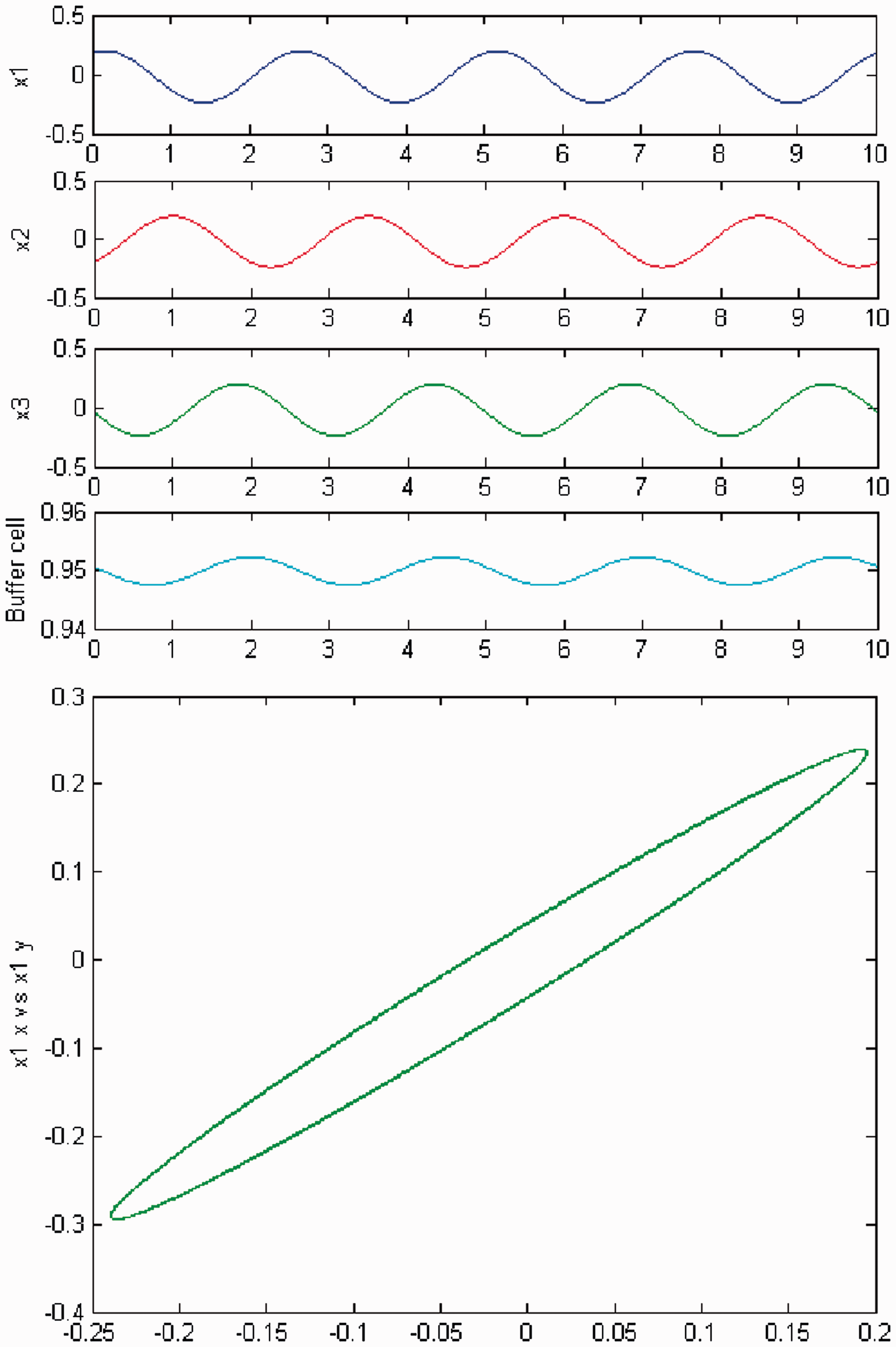
We increase c again, and a second Hopf bifurcation occurs (HB2). In Figures 4 and 5, we plot (top) the time series of the solution of the coupled cell system (3), and (bottom) the phase space of the oscillator x1 of the 3-ring. The solution is a Simulation of the coupled system (3) for λ = 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 19.54, after the second Hopf bifurcation point (HB2). The cells in the 3-ring exhibit a Simulation of the coupled system (3) for λ ≠ 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 19.54, after the second Hopf bifurcation point (HB2). The cells in the 3-ring show a 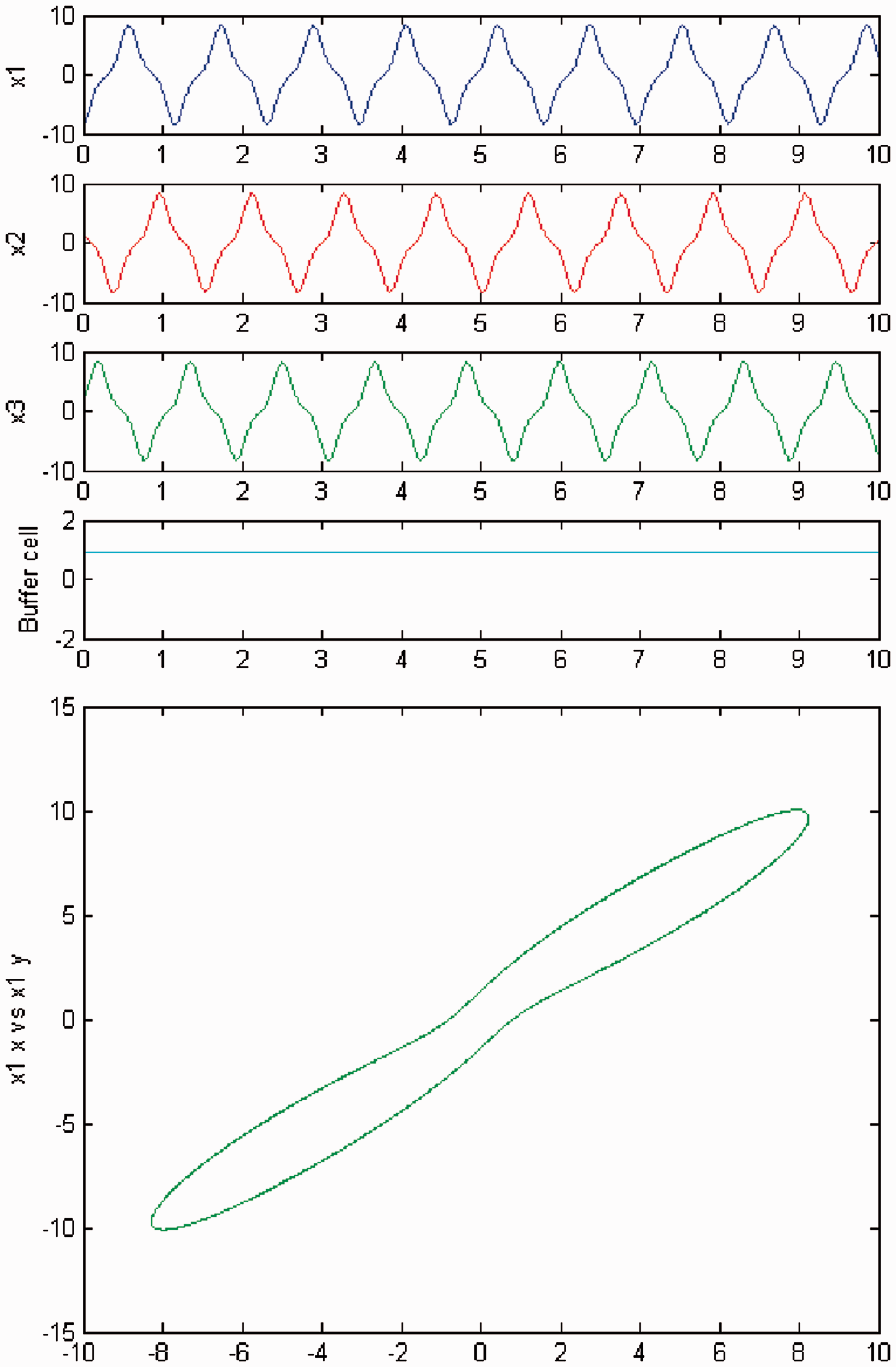
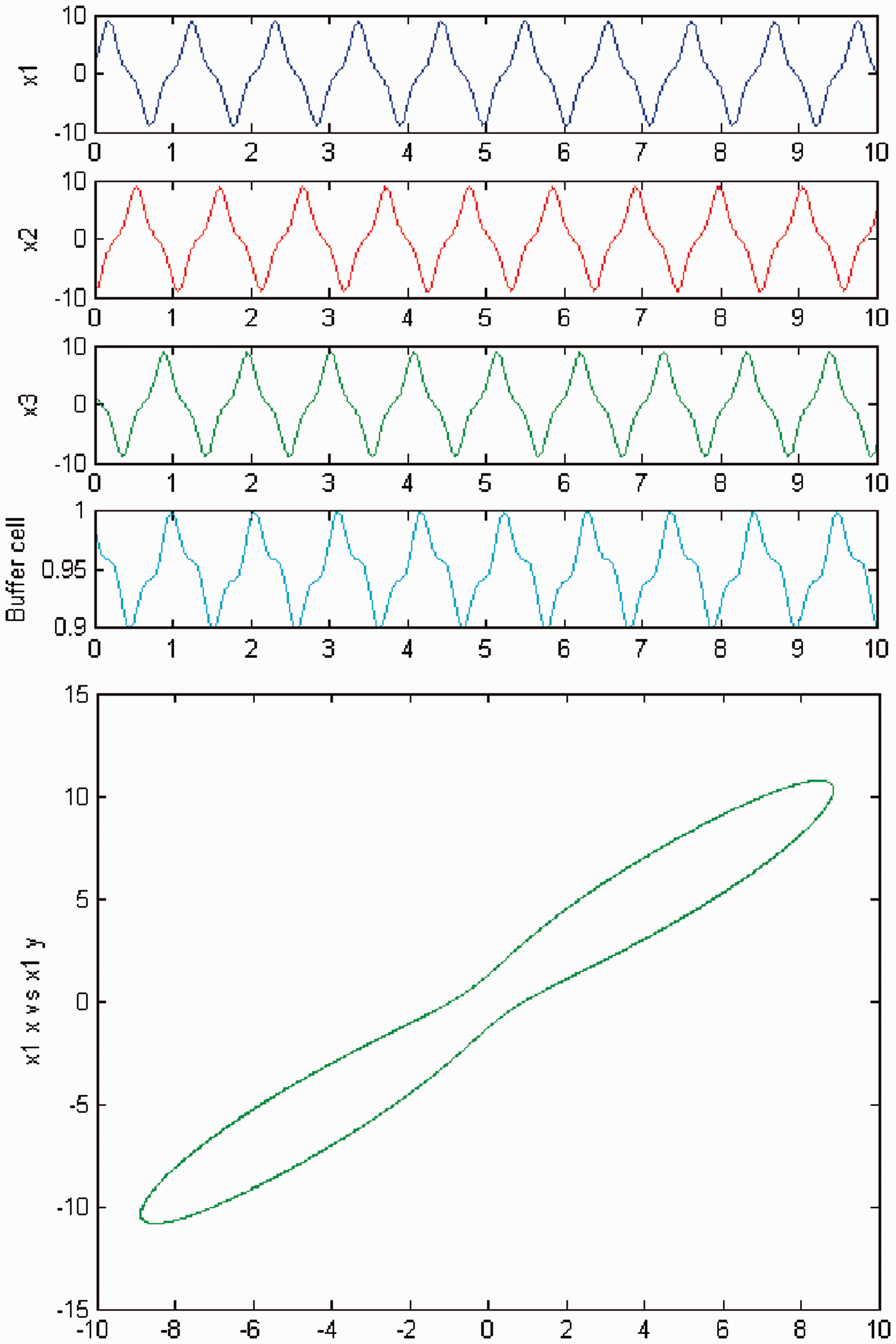
We increase parameter c again and a third Hopf bifurcation occurs (HB3). In Figures 6 and 7, we plot (top) the time series of the solution of the coupled cell system (3), and (bottom) the phase space of the oscillator x1 of the 3-ring. The solution is a Simulation of the coupled system (3) for λ = 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 20.09, after the third Hopf bifurcation point (HB3). The cells in the 3-ring exhibit a Simulation of the coupled system (3) for λ ≠ 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 20.09, after the third Hopf bifurcation point (HB3). The cells in the 3-ring exhibit a 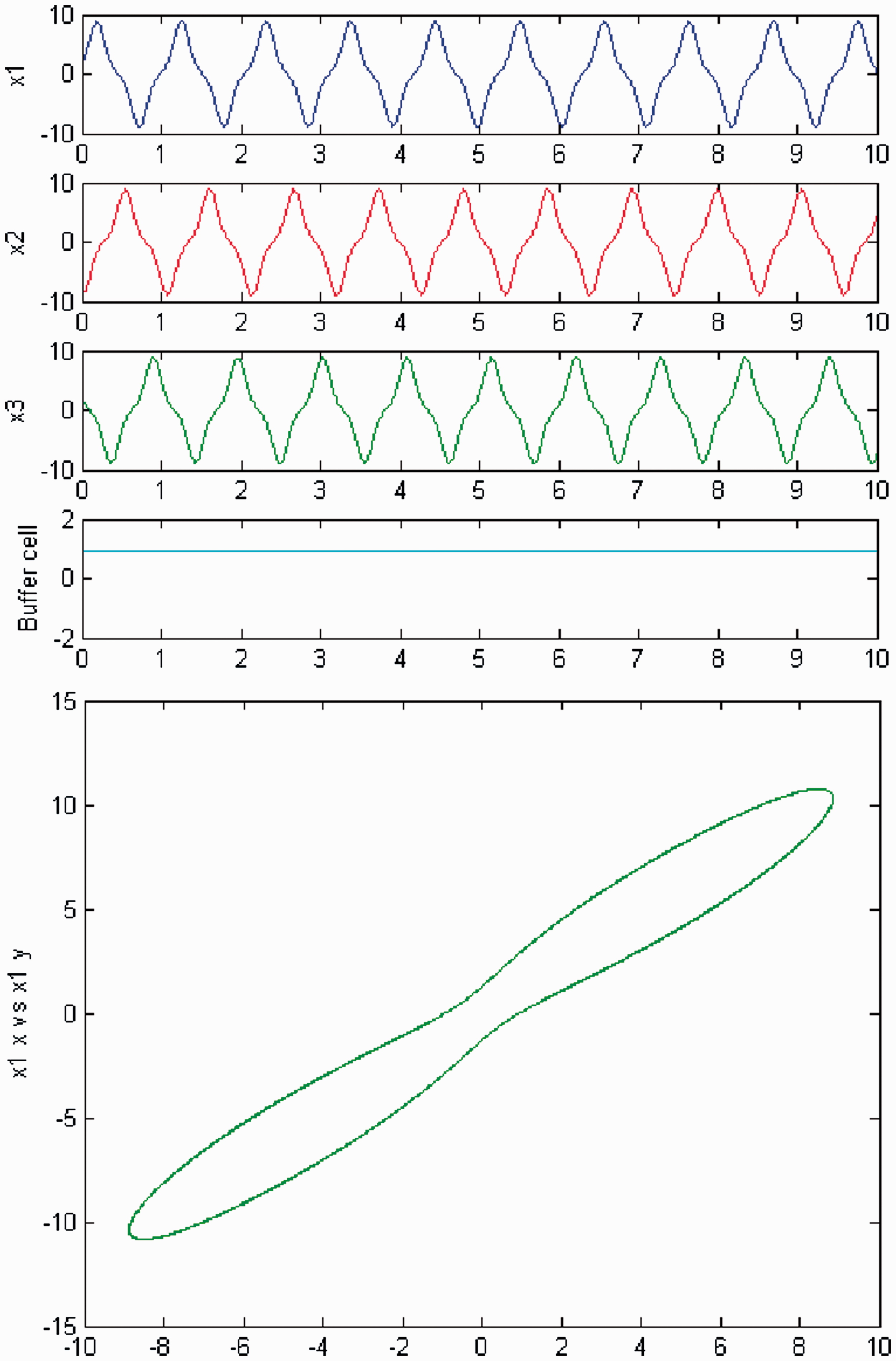
Increasing parameter c further away from the third Hopf point, a change in the qualitative behavior of the system (3) is observed, for the cases of exact (Figure 8) and ‘interior’ (Figure 9) symmetry. Namely, the cells in the 3-ring appear to show quasiperiodic motion. The ‘buffer’ cell is in equilibrium in the case of exact symmetry and appears to be quasiperiodic in the case of ‘interior’ symmetry. The full solution is quasiperiodic.
Simulation of the coupled system (3) for λ = 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 22.6, further away from the third Hopf bifurcation point (HB3). The cells in the 3-ring depict quasiperiodic motion and the ‘buffer’ cell is in a steady state. (Bottom) Phase space of the oscillator x1. Simulation of the coupled system (3) for λ ≠ 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 22.6, further away from the third Hopf bifurcation point (HB3). All cells depict quasiperiodic motion. (Bottom) Phase space of the oscillator x1.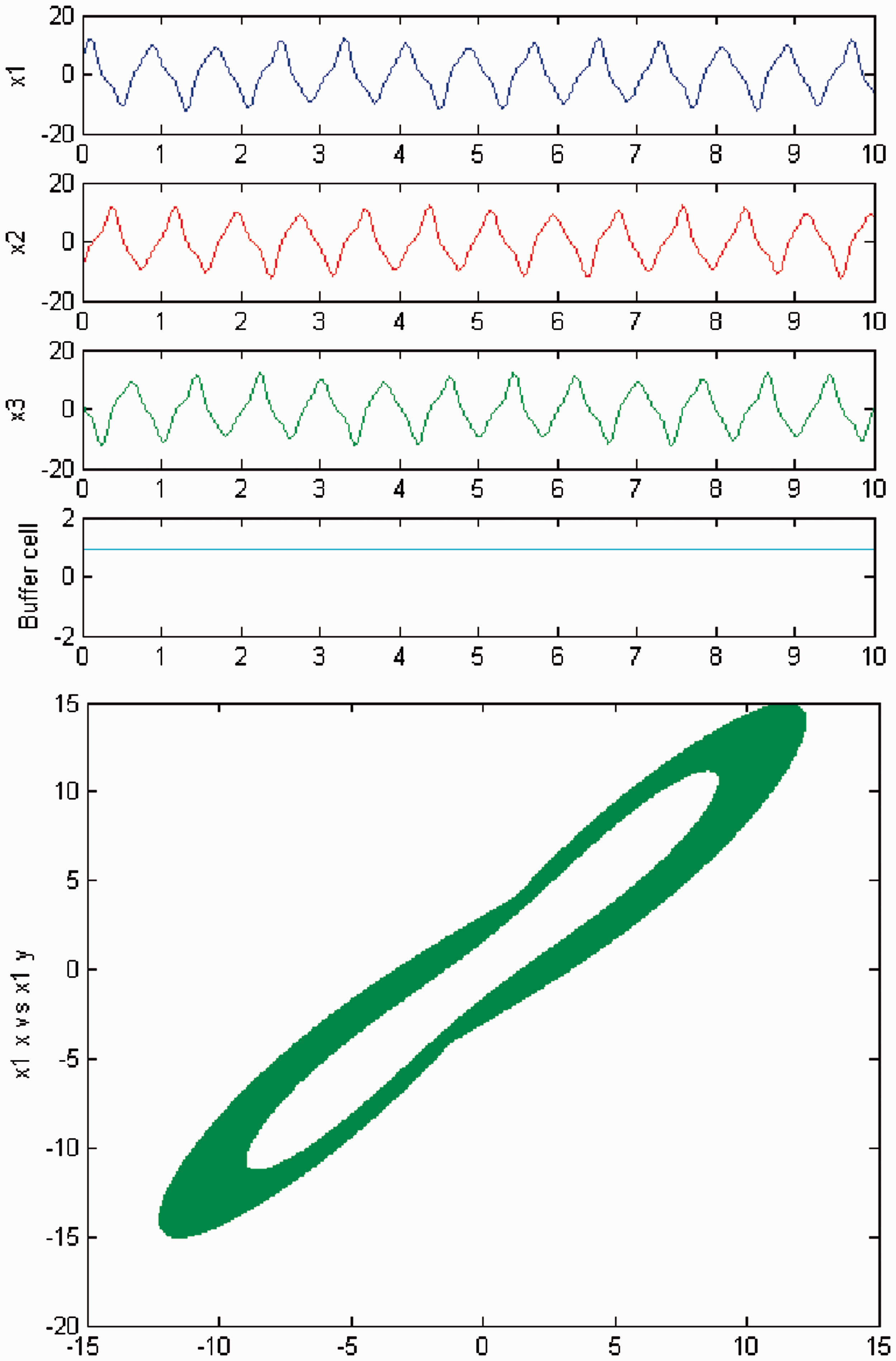

We continue to increase c. The cells in the 3-ring depict a quasiperiodic state (Figures 10 and 11), looking more ‘complex’ than the one in Figures 8 and 9. The ‘buffer’ cell is in a steady state for λ = 0 and shows quasiperiodic motion for λ ≠ 0. The full solution is quasiperiodic.
Simulation of the coupled system (3) for λ = 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 23. The cells in the ring exhibit quasiperiodic motion. The ‘buffer’ cell is in equilibrium. (Bottom) Phase space of the oscillator x1. Simulation of the coupled system (3) for λ ≠ 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 23. All cells show quasiperiodic motion. (Bottom) Phase space of the oscillator x1.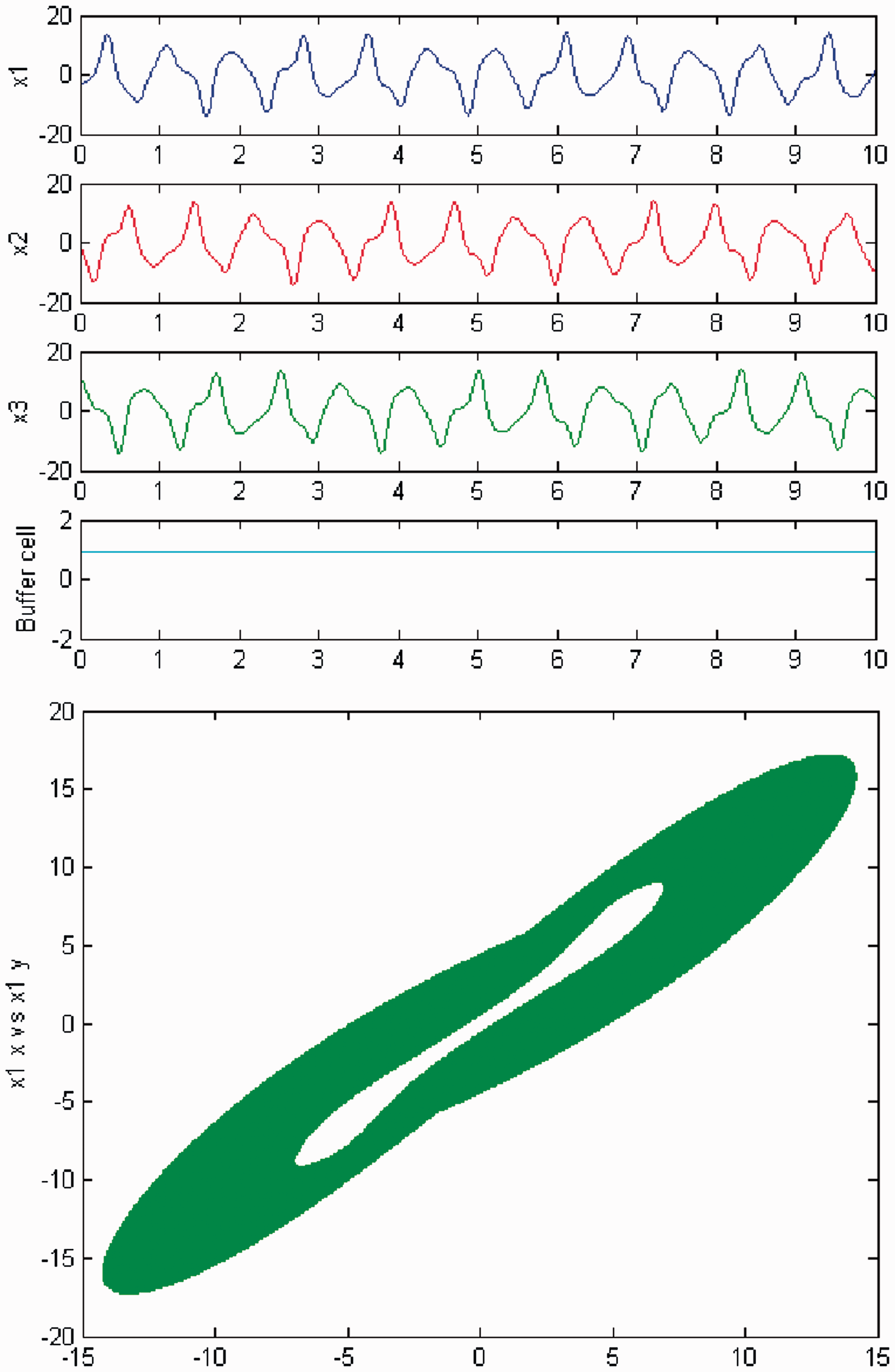
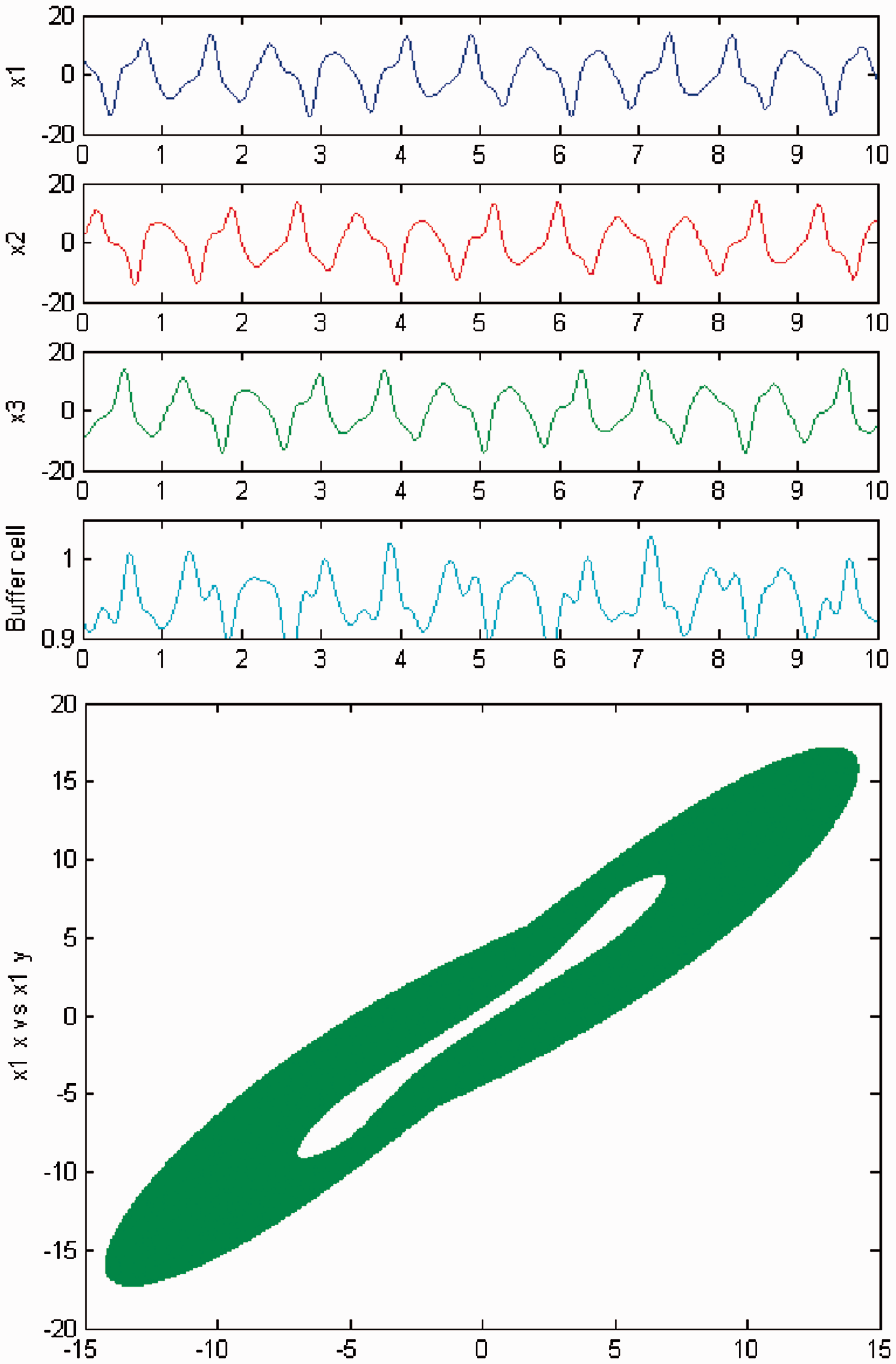
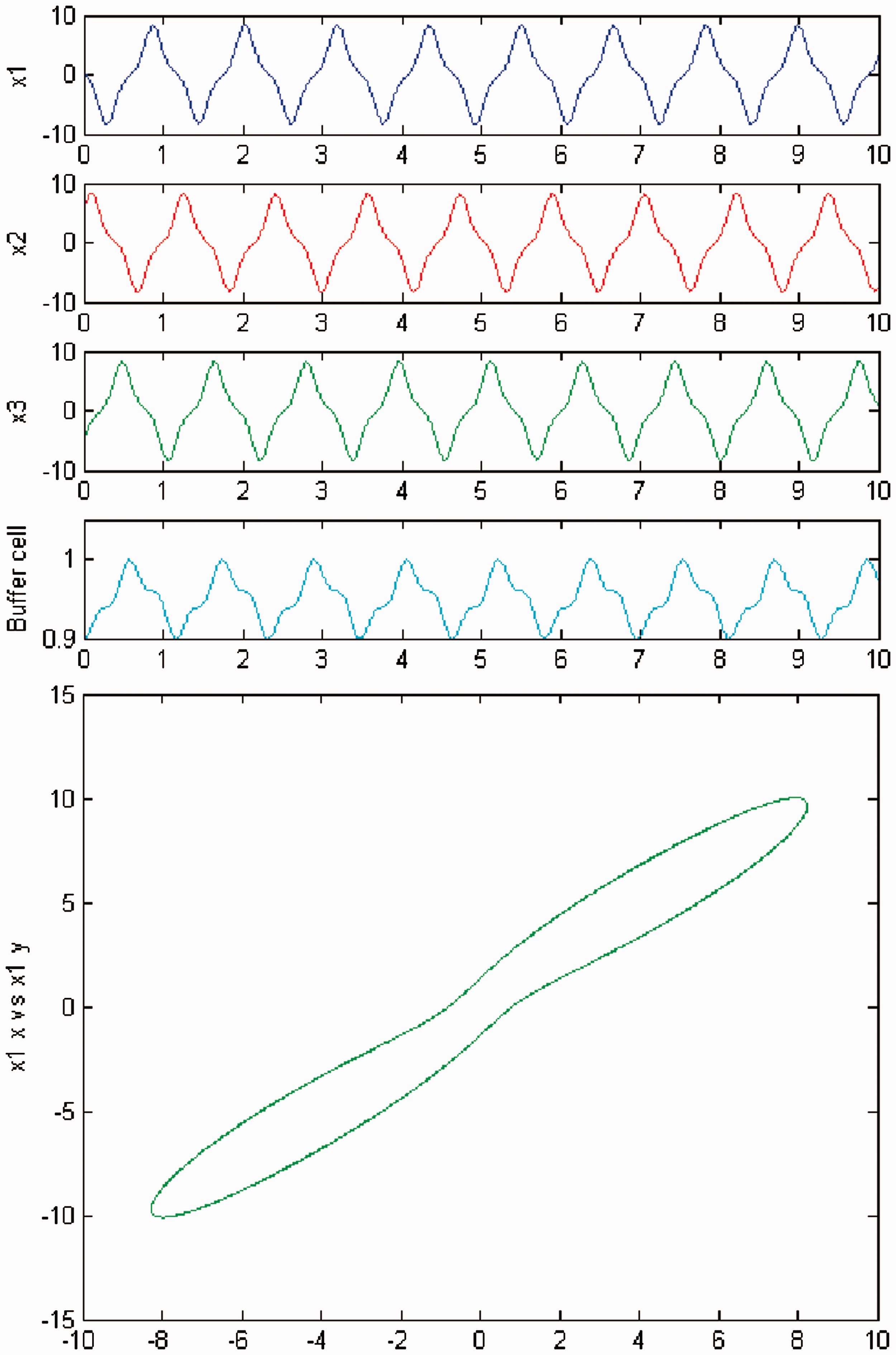
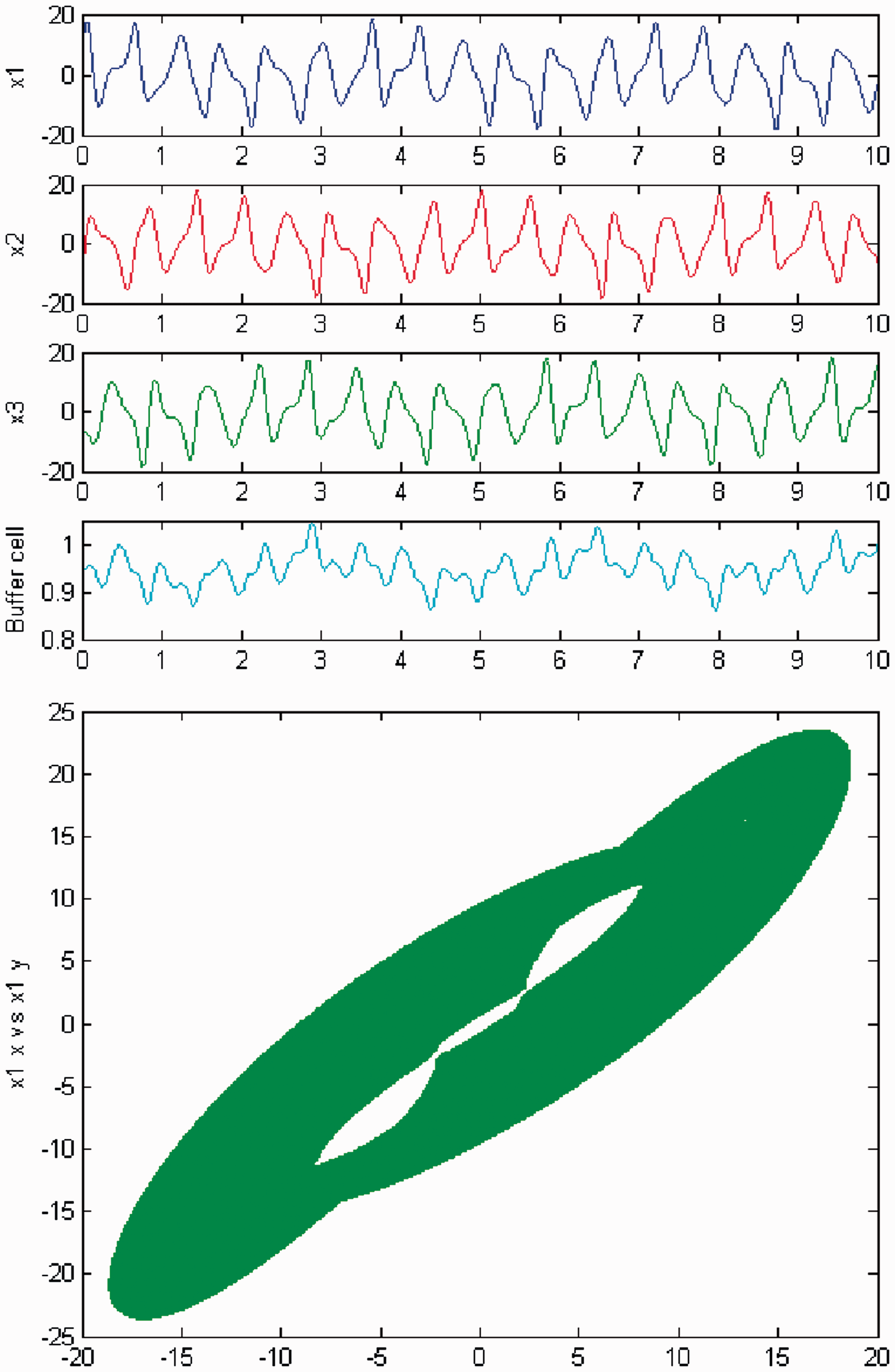
We persist in increasing the parameter c, and a new qualitative change in the behavior of the coupled cell system (3) is visible. Cells in the 3-ring show chaotic motion (see Figures 12 and 13). The ‘buffer’ cell is in a steady state in the case of exact symmetry and in a chaotic state for the case of ‘interior’ symmetry.
Simulation of the coupled system (3) for λ = 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 23.4. The cells in the 3-ring exhibit chaotic motion. The ‘buffer’ cell is in a steady state. (Bottom) Phase space of the oscillator x1. Simulation of the coupled system (3) for λ ≠ 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 23.4. All cells in the network depict chaotic motion. (Bottom) Phase space of the oscillator x1.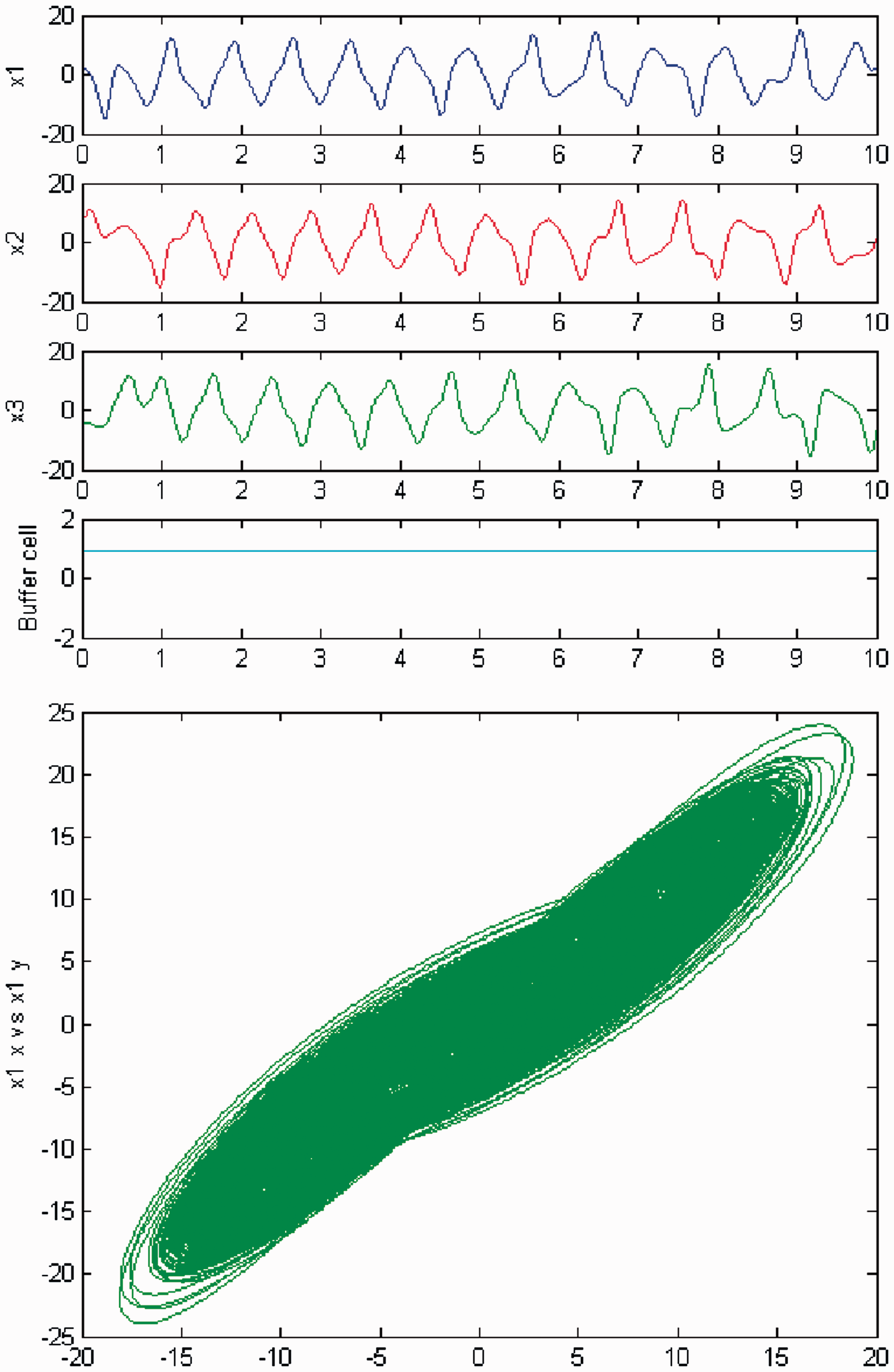
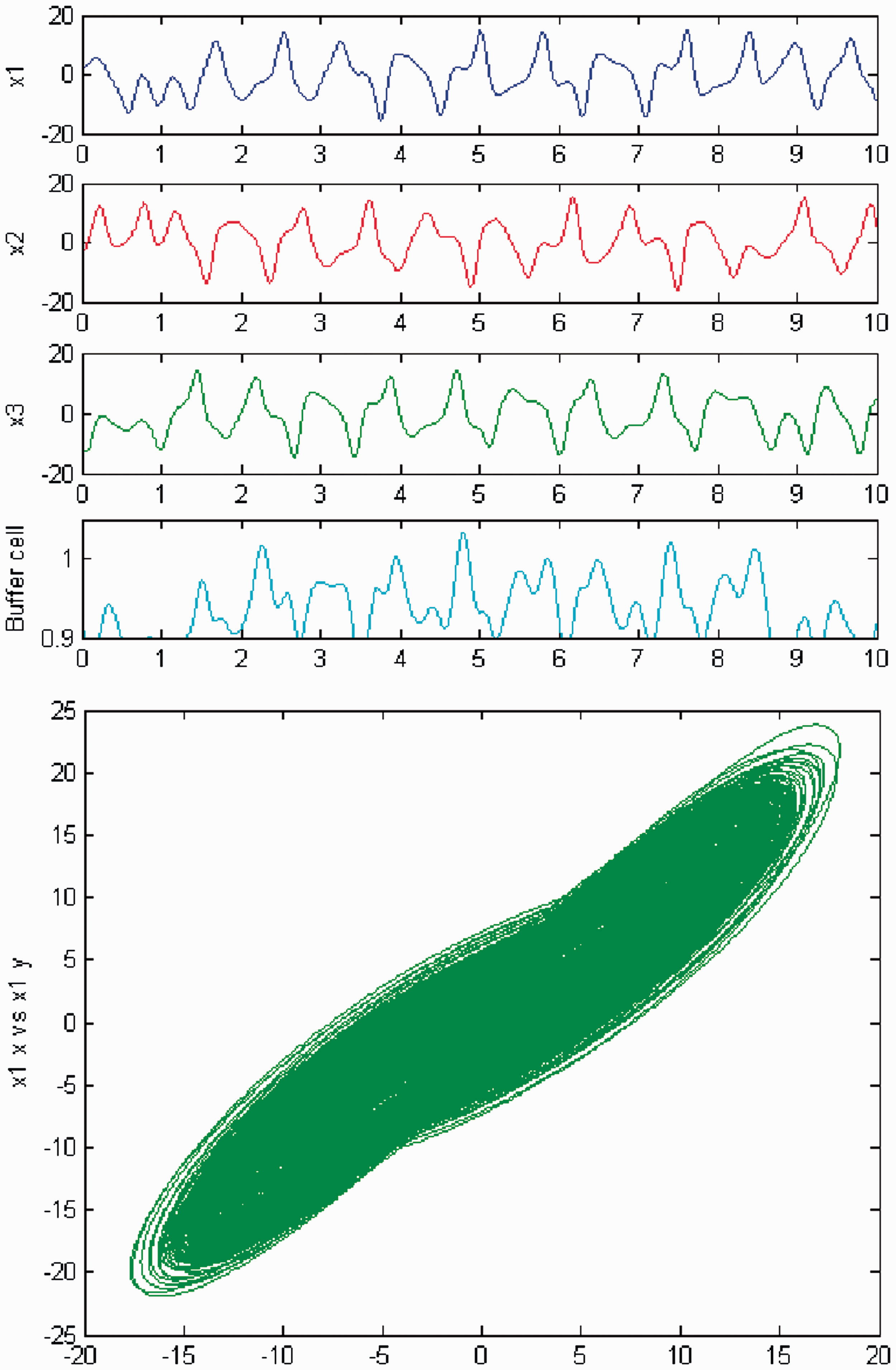
Incrementing c again, another change occurs in the dynamical behavior of the cells of the networks in Figure 1(a). The cells in the 3-ring now depict quasiperiodic motion again (Figures 14 and 15). The ‘buffer’ cell is in equilibrium for λ = 0 and in a quasiperiodic state for λ ≠ 0. The full solution is quasiperiodic.
Simulation of the coupled system (3) for λ = 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 25.6. The cells in the 3-ring show quasiperiodic motion. The ‘buffer’ cell is in equilibrium. (Bottom) Phase space of the oscillator x1. Simulation of the coupled system (3) for λ ≠ 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 25.6. All cells in the network show quasiperiodic motion. (Bottom) Phase space of the oscillator x1.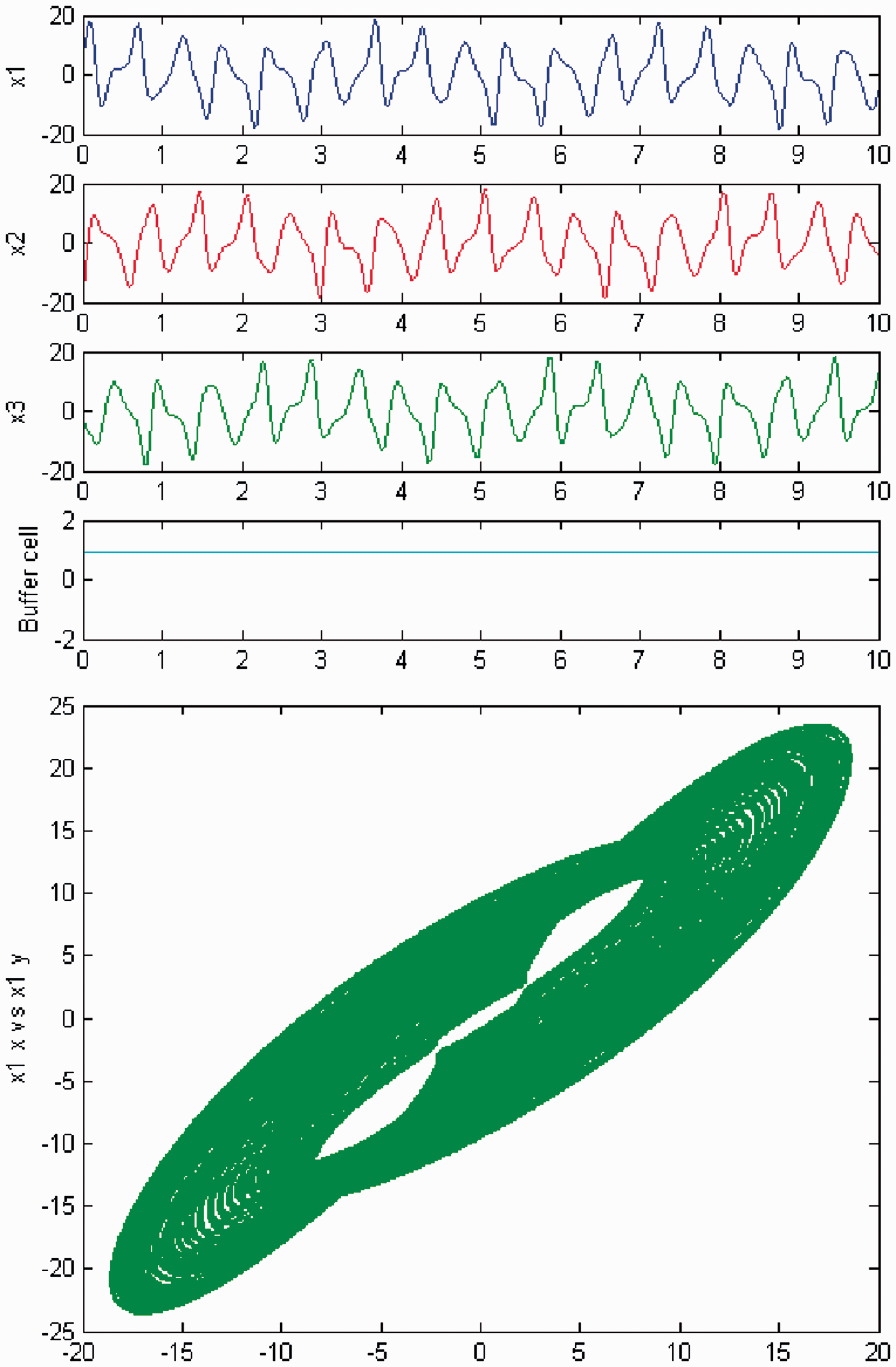
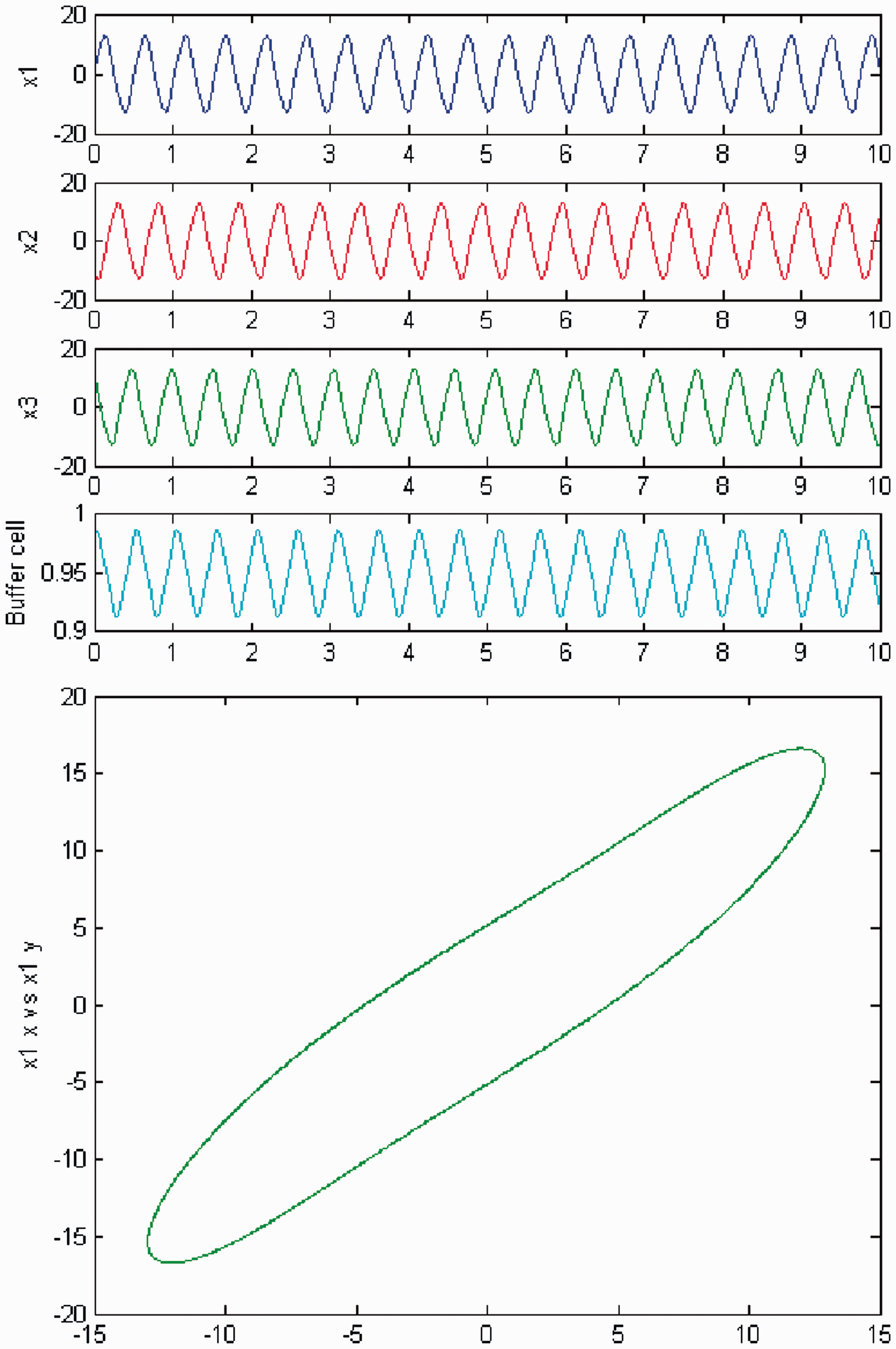
The quasiperiodic behavior disappears as c is increased once again. The coupled cell system returns to a periodic state, where the cells in the 3-ring depict a Simulation of the coupled system (3) for λ = 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 27. The cells in the ring depict a Simulation of the coupled system (3) for λ ≠ 0 and c2 = 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 27. The cells in the ring exhibit a 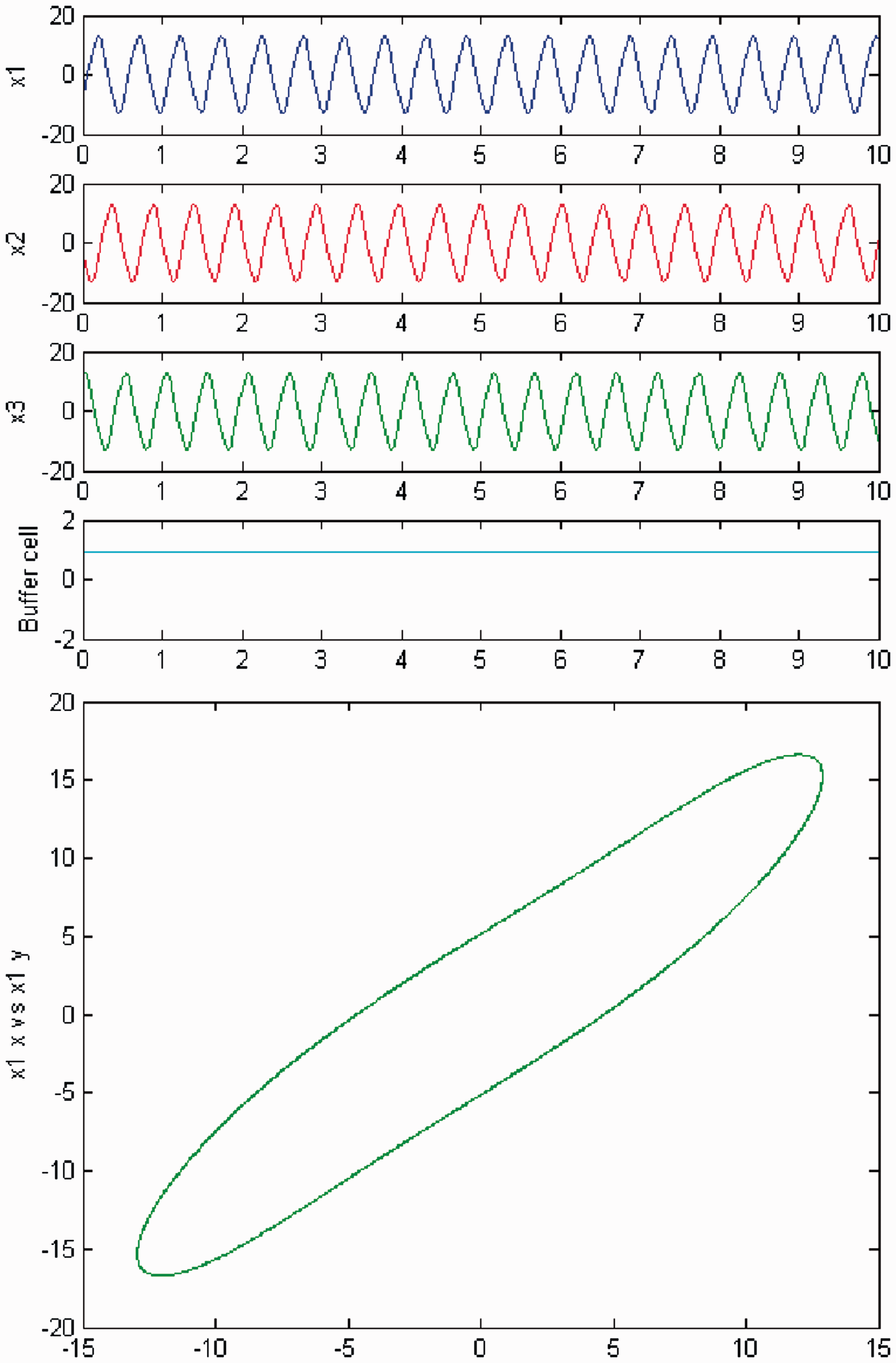
3.2. Bidirectional networks: dynamics
We now simulate the networks in Figure 1(b). We vary parameter c ∈ [15, 29], going from lower to higher values, and start from a steady state of the whole system. We now set c2 = −1, and consider the cases λ = 0 and λ ≠ 0.
We increase the parameter c, starting at c = 15, and a first Hopf bifurcation (HB1) occurs at c = 15.39, from the full equilibrium of the system. The cells in the 3-ring depict a Simulation of the coupled system (3) for λ = 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 15.39, after the first Hopf bifurcation point (HB1). The cells in the ring exhibit a Simulation of the coupled system (3), for λ ≠ 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 15.39, after the first Hopf bifurcation point (HB1). The cells in the ring exhibit a 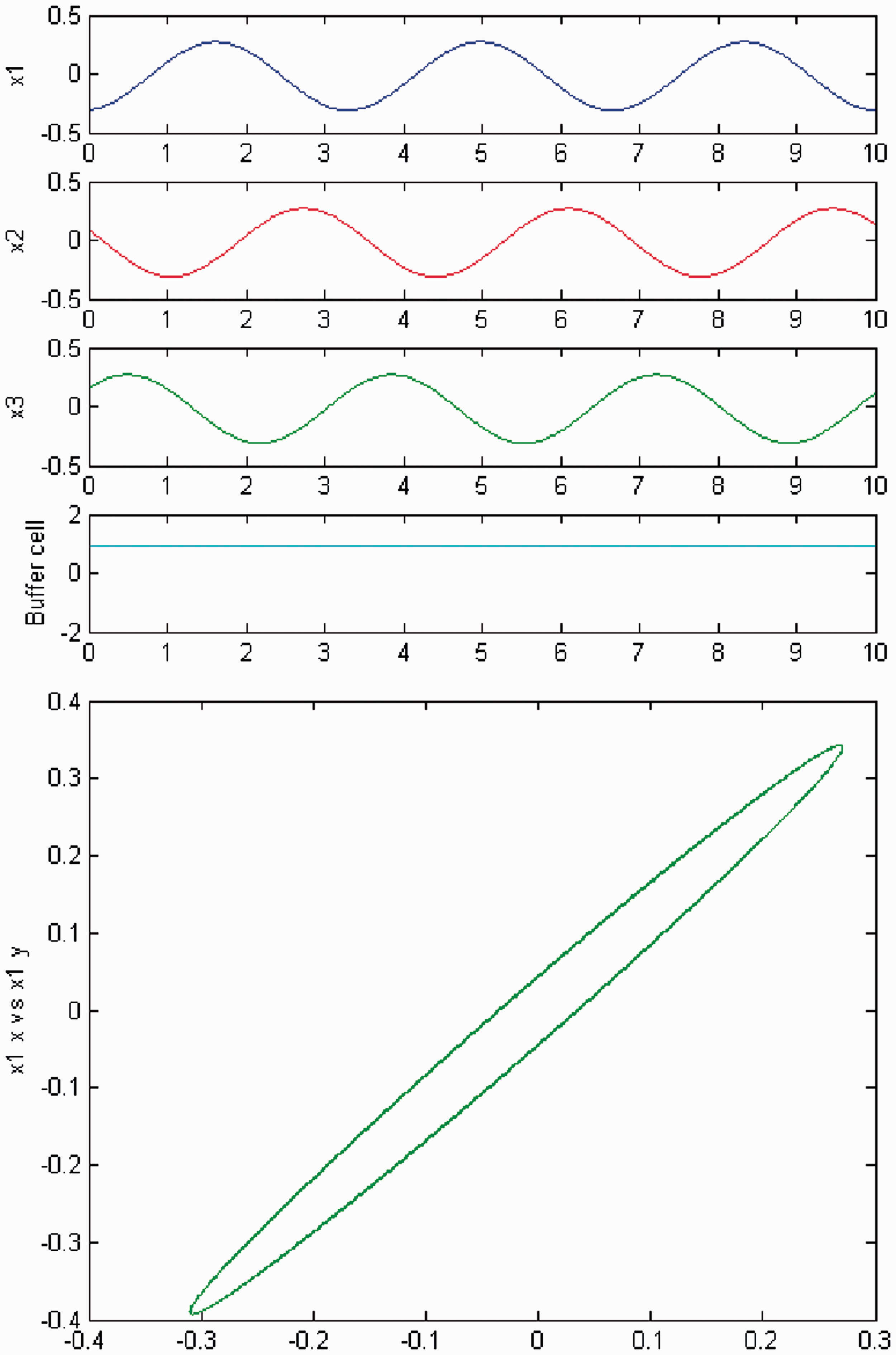
We increase c again, and a second Hopf bifurcation occurs (HB2). In Figures 20 and 21, we plot (top) the time series of the solution of the coupled cell system (3), and (bottom) the phase space of the oscillator x1 of the 3-ring. The solution is a Simulation of the coupled system (3) for λ = 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 19.03, after the second Hopf bifurcation point (HB2). The cells in the 3-ring exhibit a Simulation of the coupled system (3) for λ ≠ 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 19.03, after the second Hopf bifurcation point (HB2). The cells in the 3-ring exhibit a 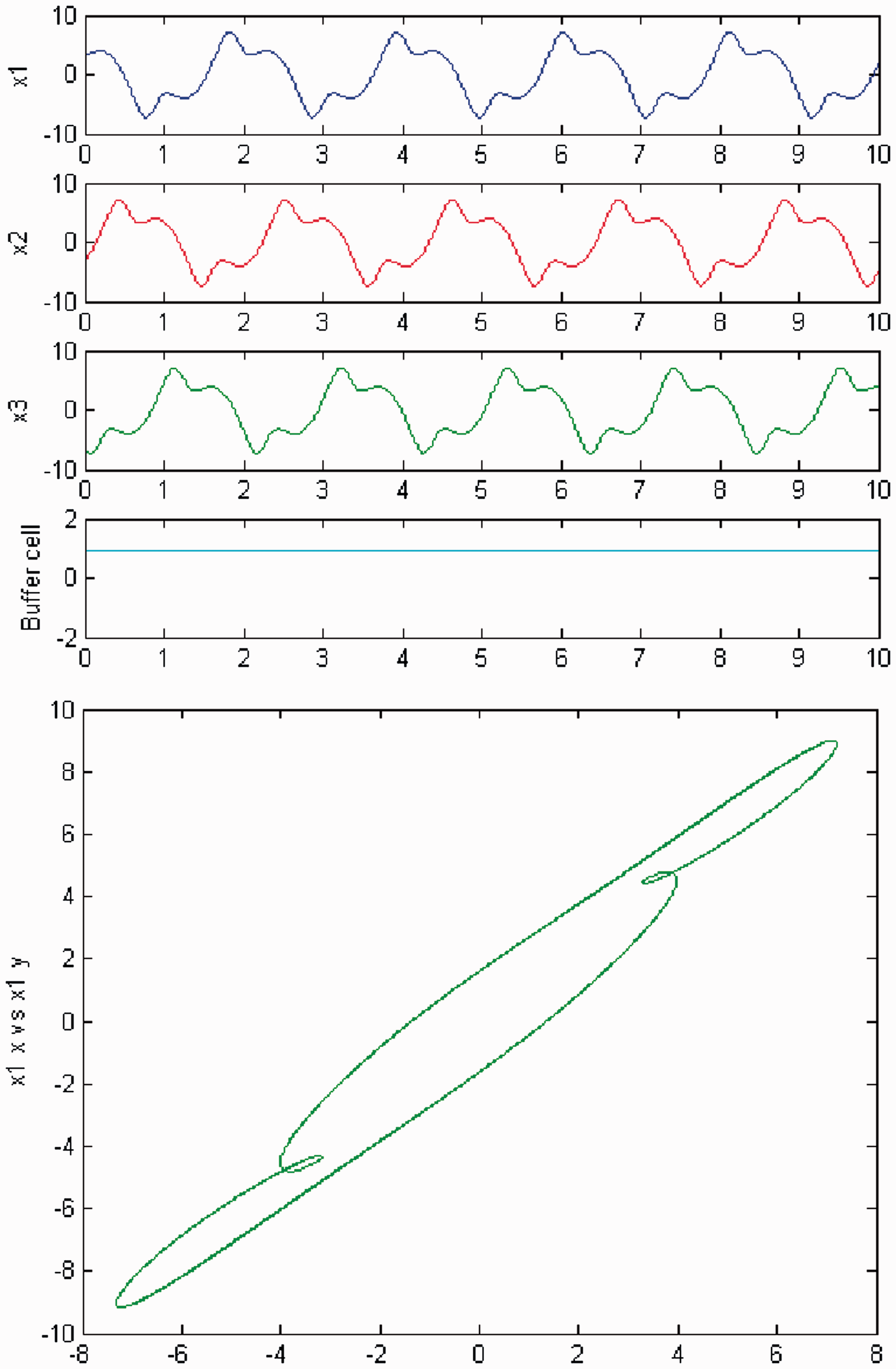
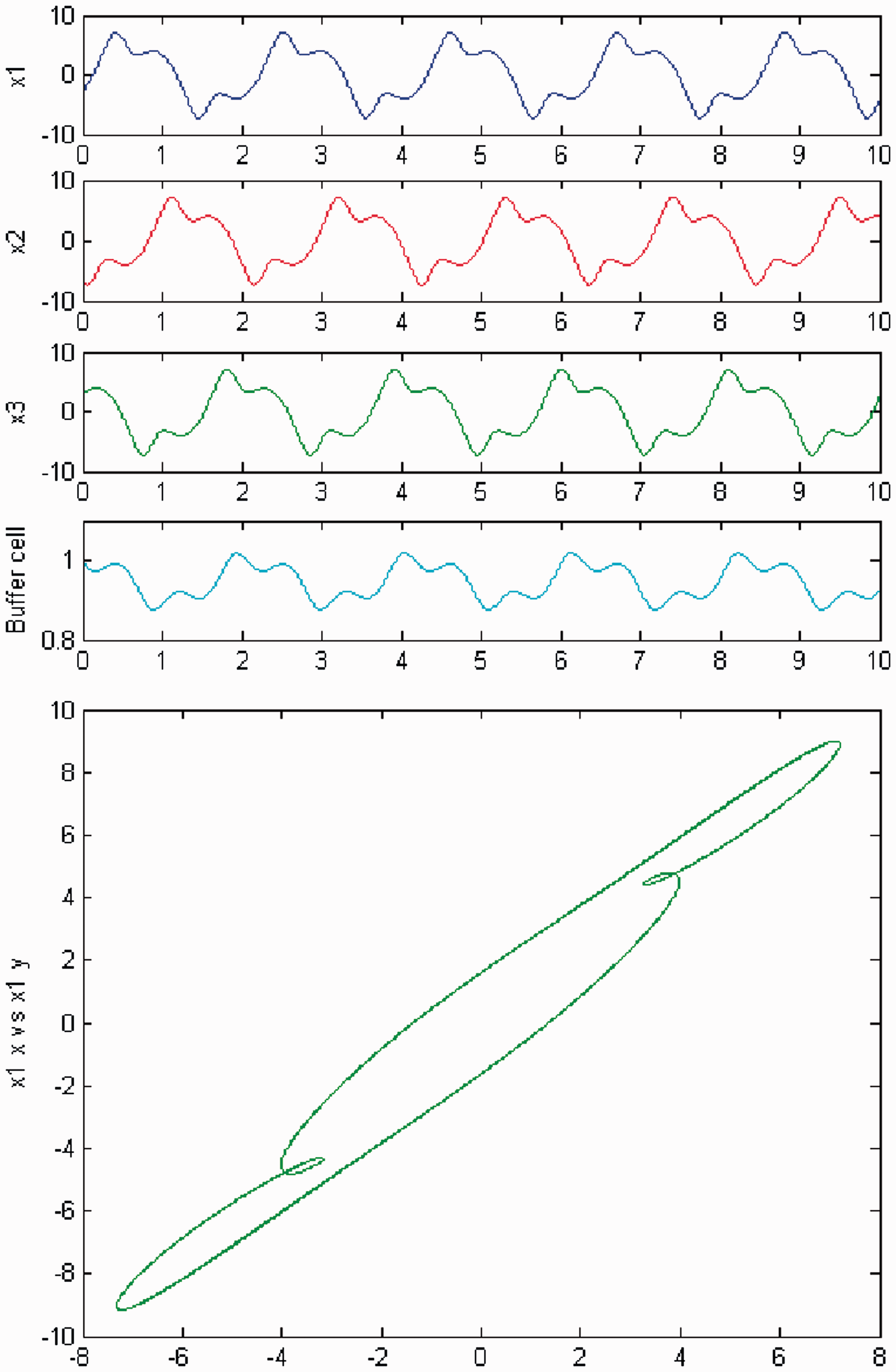
We increase parameter c again and a third Hopf bifurcation occurs (HB3). In Figures 22 and 23, we plot (top) the time series of the solution of the coupled cell system (3), and (bottom) the phase space of the oscillator x1 of the 3-ring. The solution is a Simulation of the coupled system (3) for λ = 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 20.09, after the third Hopf bifurcation point (HB3). The cells in the 3-ring exhibit a Simulation of the coupled system (3) for λ ≠ 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 20.09, after the third Hopf bifurcation point (HB3). The cells in the 3-ring exhibit a 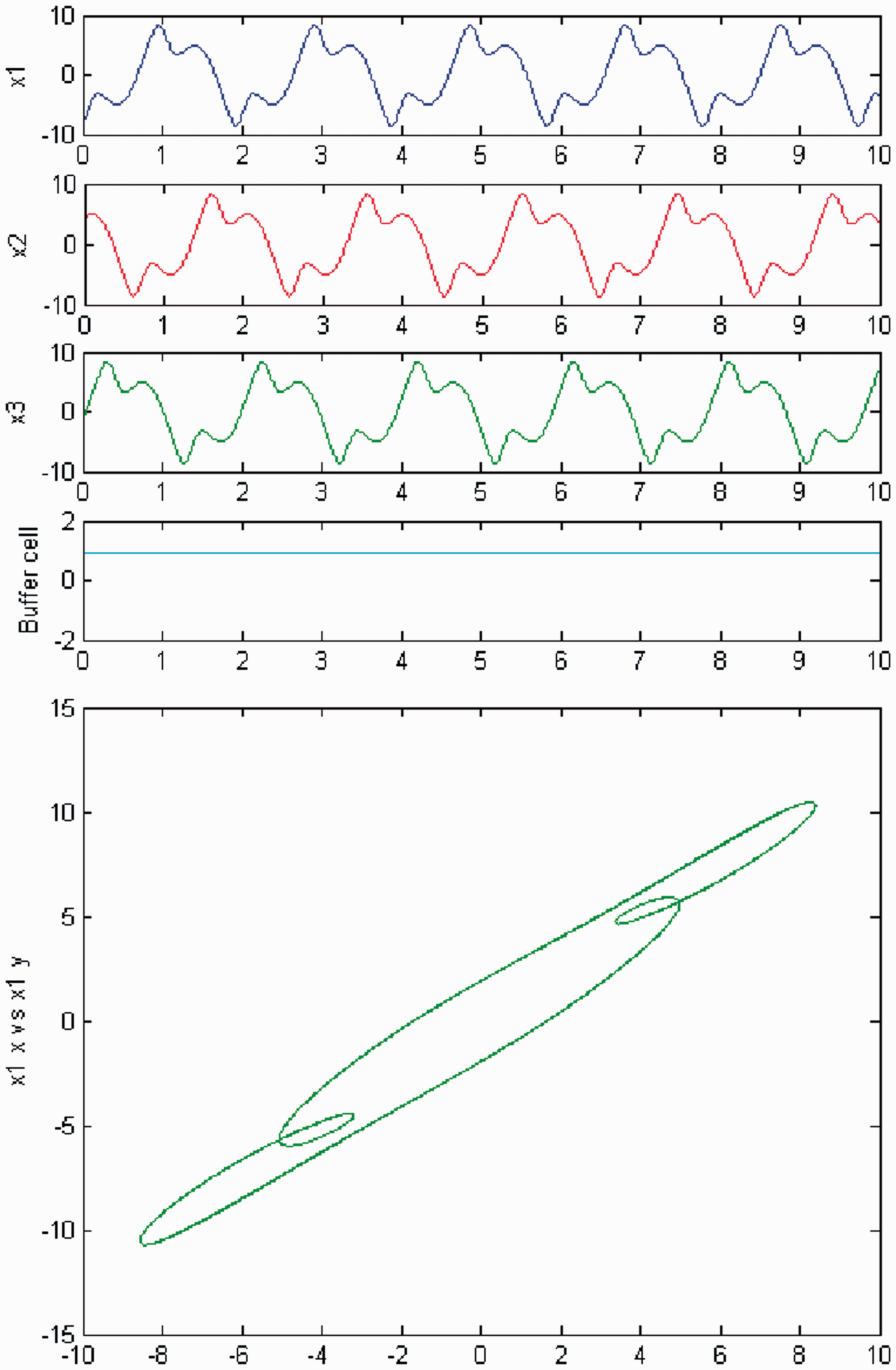
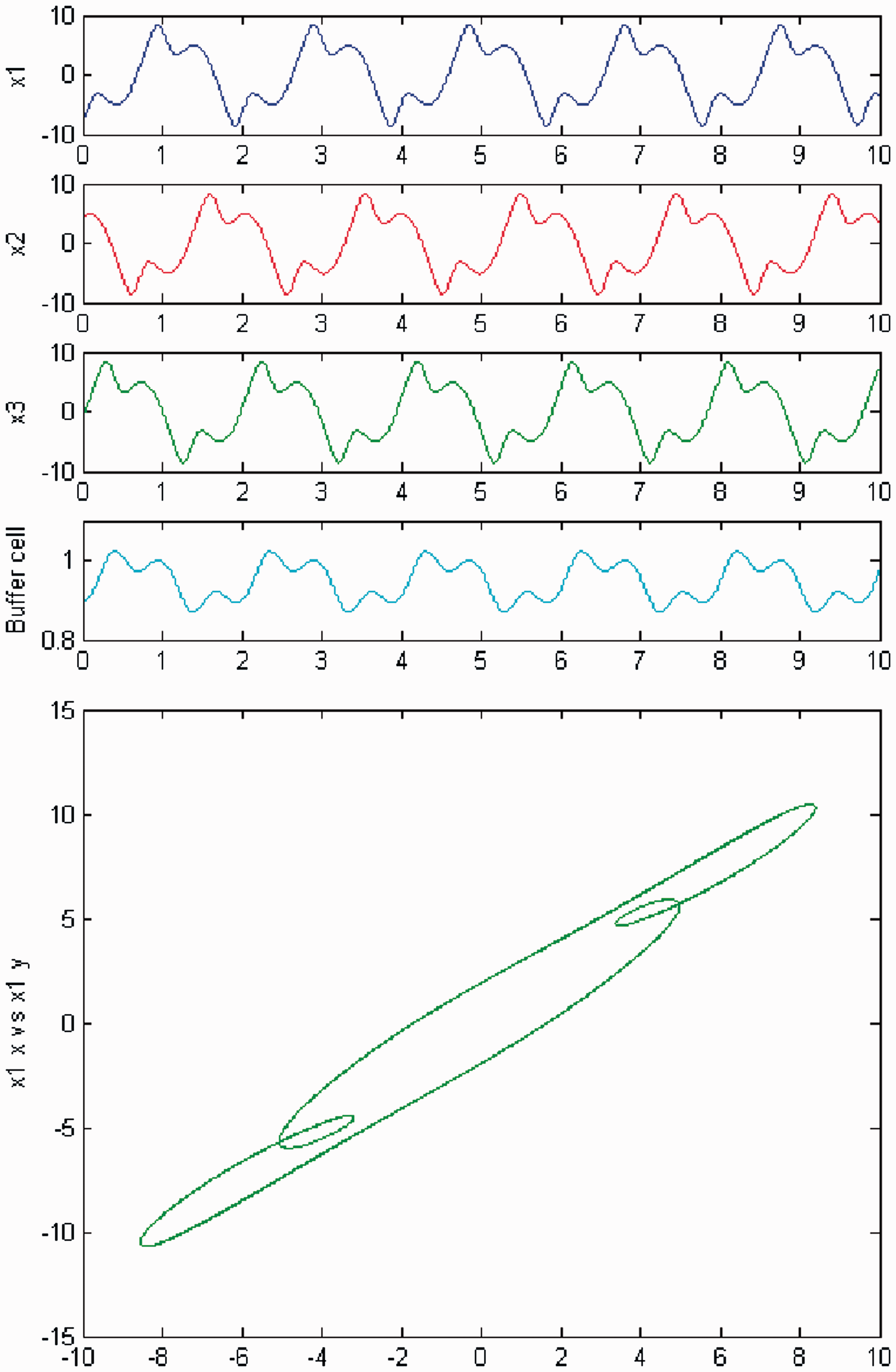
Increasing parameter c further away from the third Hopf point, a change in the qualitative behavior of the system (3) is observed, for the cases of exact and ‘interior’ symmetry. Namely, the cells in the 3-ring appear to show quasiperiodic motion. The ‘buffer’ cell is in equilibrium in the case of exact symmetry (Figure 24) and appears to be quasiperiodic in the case of ‘interior’ symmetry (Figure 25). The full solution is quasiperiodic.
Simulation of the coupled system (3) for λ = 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 23.6, further away from the third Hopf bifurcation point (HB3). The cells in the 3-ring depict quasiperiodic motion and the ‘buffer’ cell is in a steady state. (Bottom) Phase space of the oscillator x1. Simulation of the coupled system (3) for λ ≠ 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 23.6, further away from the third Hopf bifurcation point (HB3). All cells depict quasiperiodic motion. (Bottom) Phase space of the oscillator x1.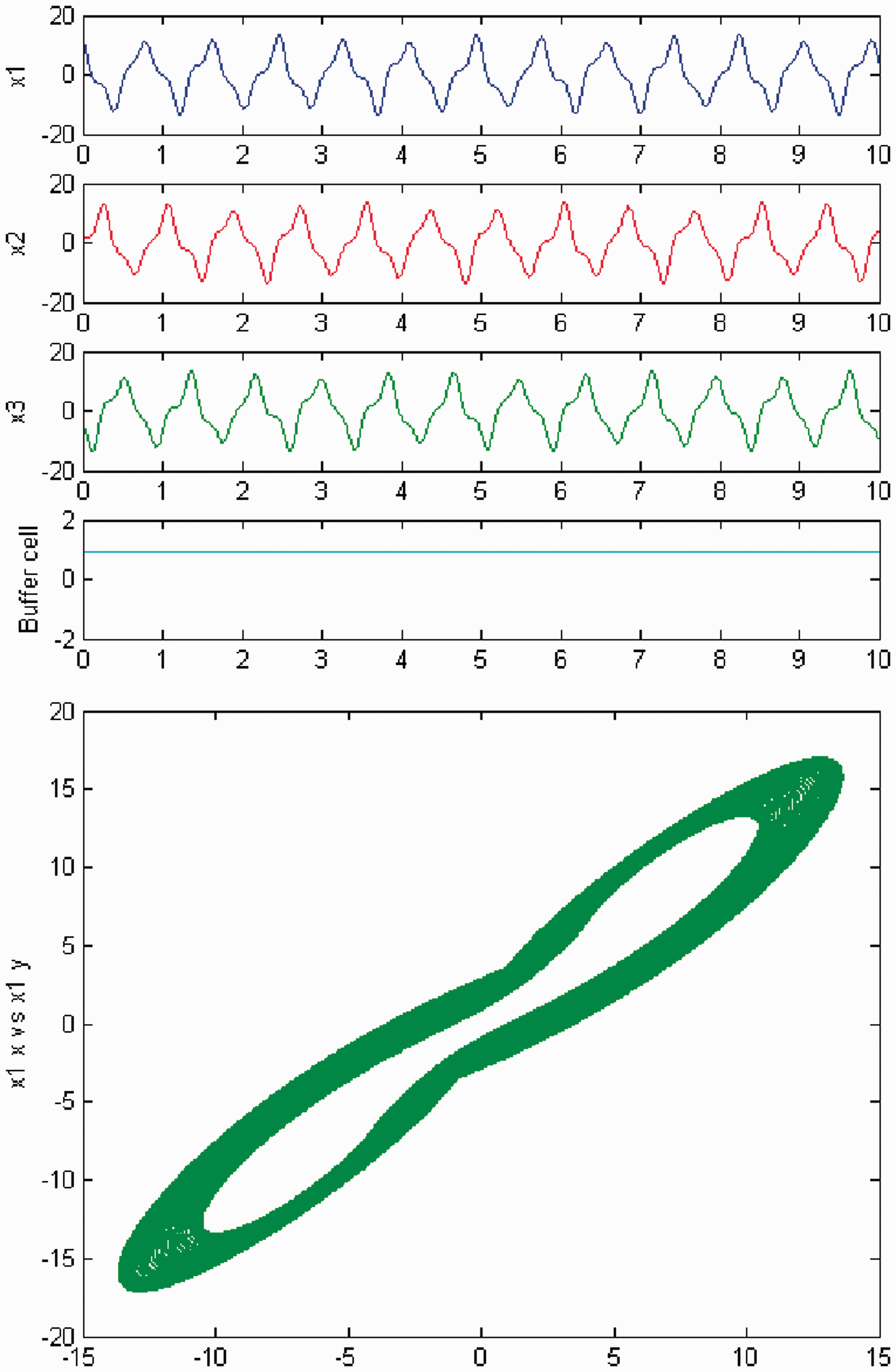
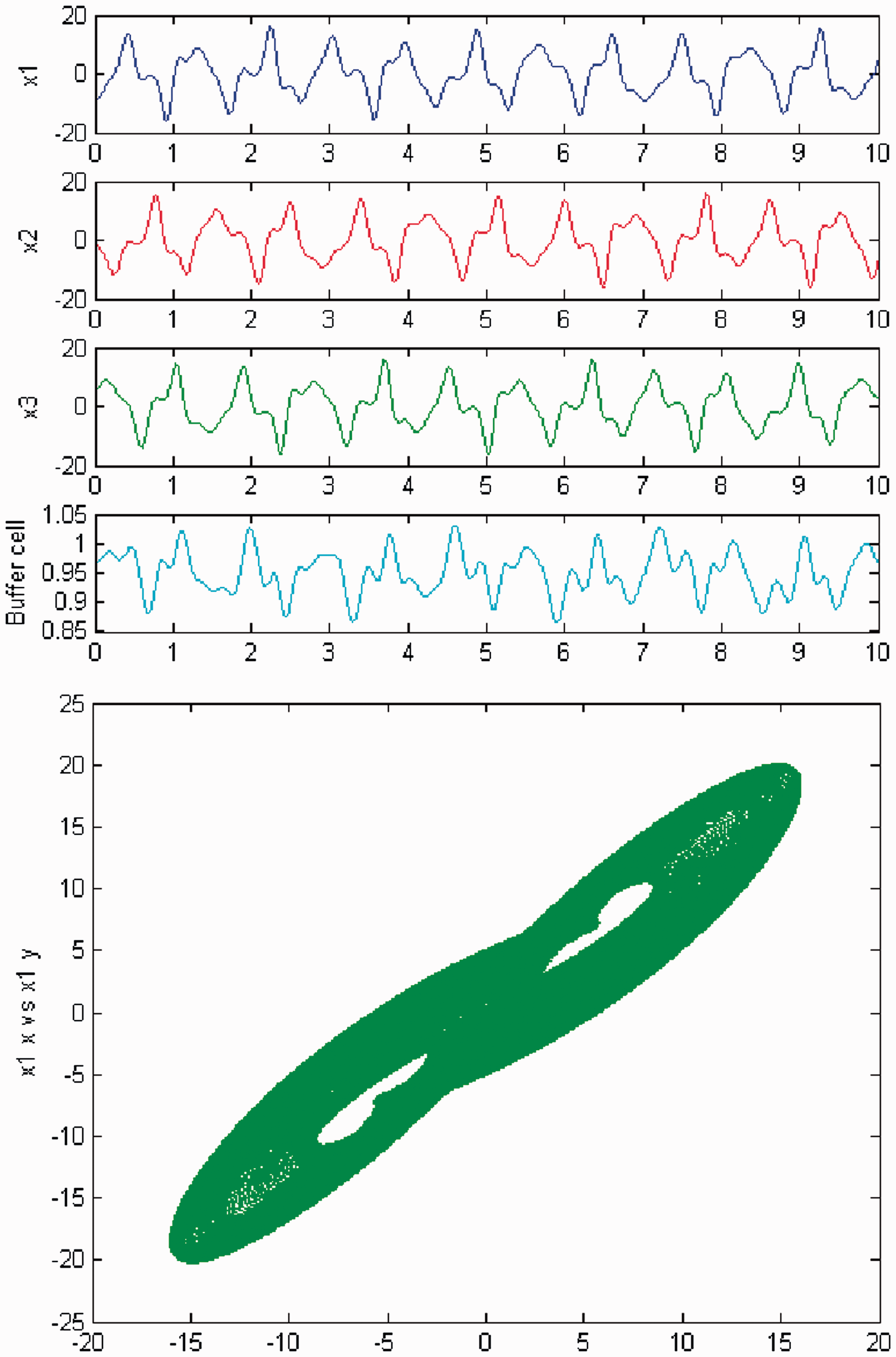
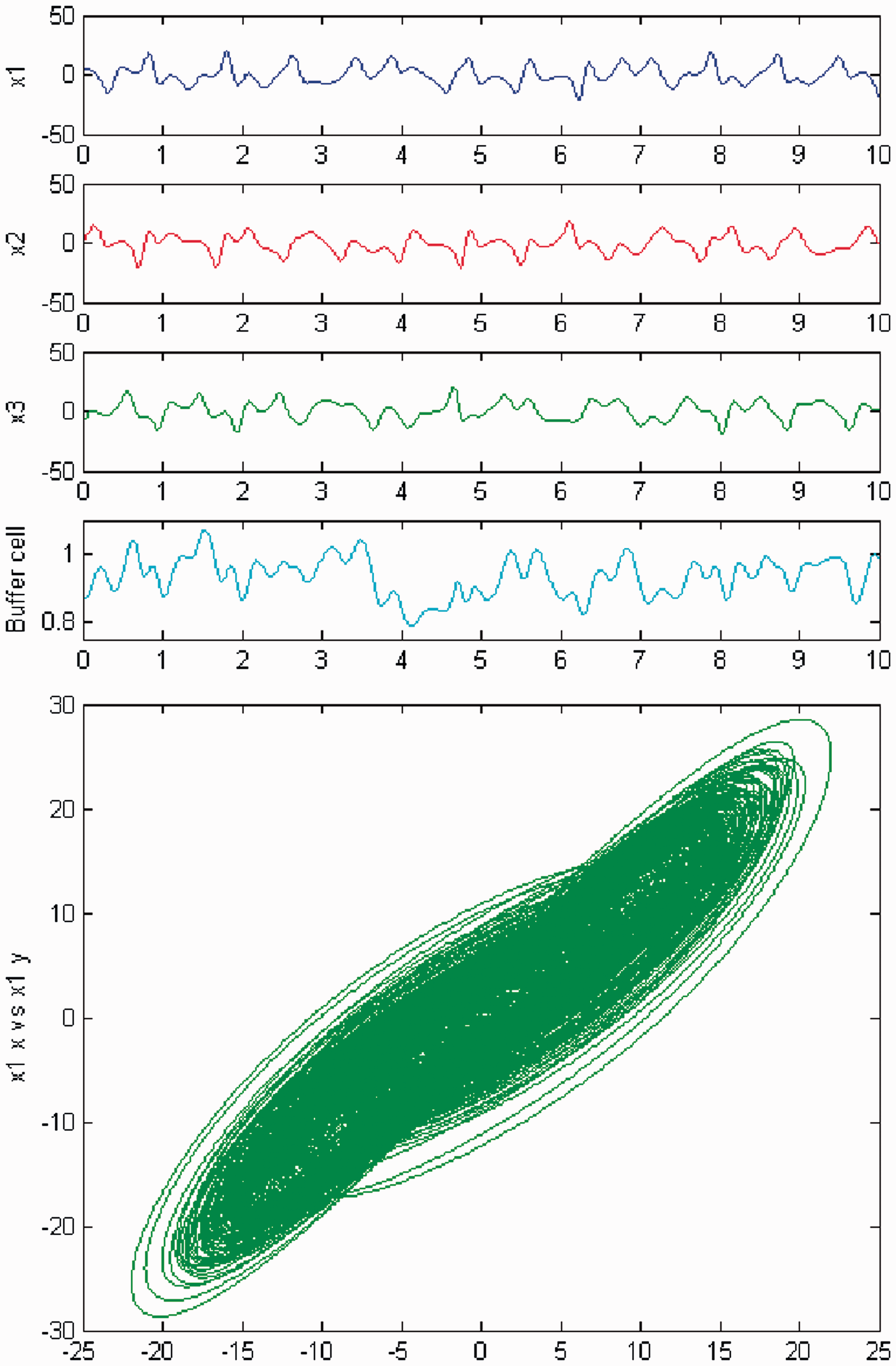
We persist in the increase of parameter c, and a new qualitative change in the behavior of the coupled cell system (3) is observed. Cells in the 3-ring show chaotic motion (see Figures 26 and 27). The ‘buffer’ cell is in a steady state for λ = 0 and in a chaotic state for λ ≠ 0.
Simulation of the coupled system (3) for λ = 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 29. The cells in the 3-ring exhibit chaotic motion. The ‘buffer’ cell is in a steady state. (Bottom) Phase space of the oscillator x1. Simulation of the coupled system (3) for λ ≠ 0 and c2 ≠ 0. (Top) Time series of the cells in the 3-ring and of the ‘buffer’ cell, for c = 29. All cells in the network depict chaotic motion. (Bottom) Phase space of the oscillator x1.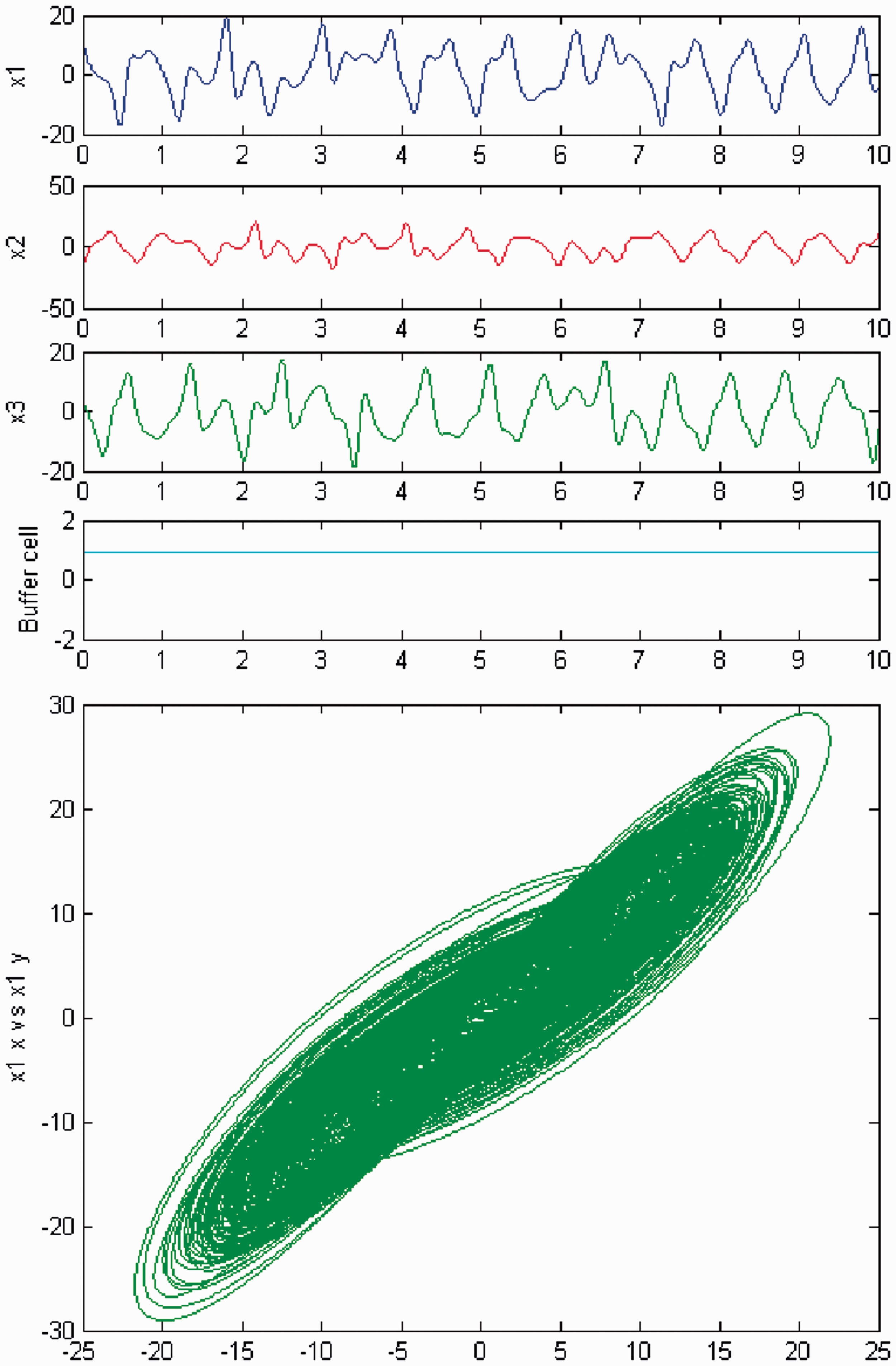
3.3. Discussion
The schematic bifurcation diagram of the coupled system (3) is reproduced in Figure 28. This is a schematic bifurcation diagram of a sequence of three Hopf bifurcations, followed by a cascade of period-doubling bifurcations, and period-halving bifurcations. The first Hopf bifurcating branch emanates from a trivial branch of steady states. Note that, for the parameter region considered here, we do not observe, in the networks in Figure 1(b), the period-halving bifurcations. We have stopped in the chaotic state, after the period-doubling bifurcations.
Schematic (partial) bifurcation diagram for the coupled cell system (3), near the equilibrium point. Solid lines represent stable solutions. HBi, i = 1,2,3, are Hopf bifurcation points, PD are period-doubling bifurcations, PH are period-halving bifurcations, PO are periodic solutions, and QP are quasiperiodic states.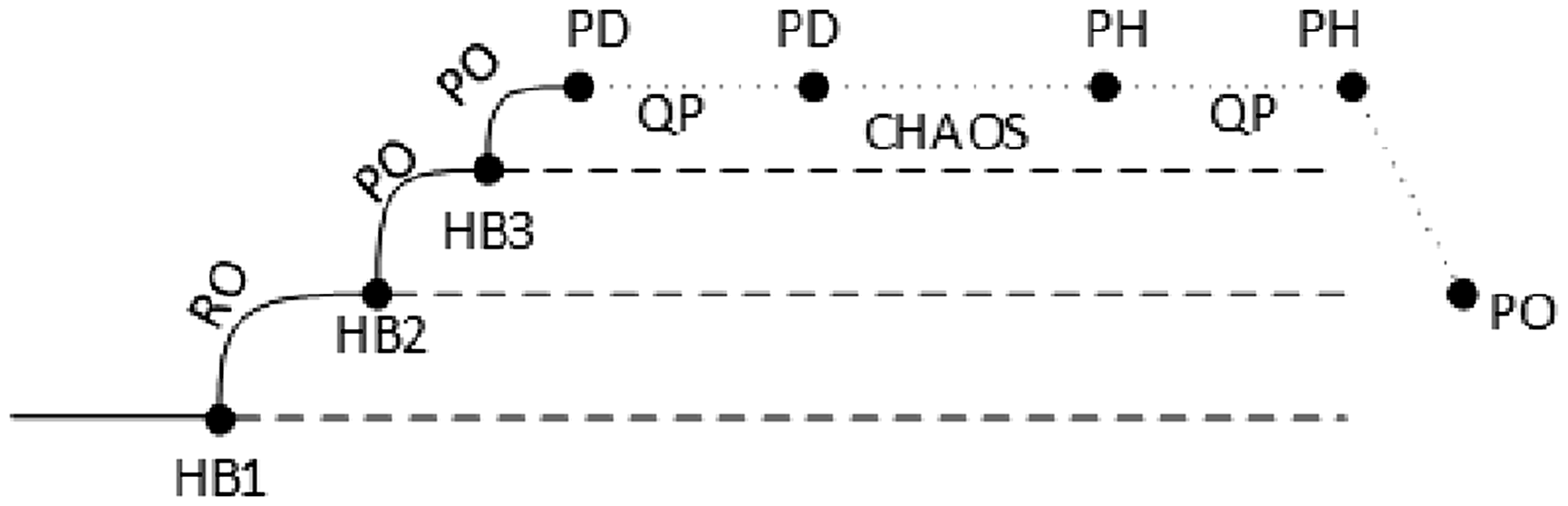
The solutions arising after the primary Hopf bifurcation point are explained by the equivariant Hopf theorem for coupled cell systems in the symmetric case, and the interior symmetry-breaking Hopf theorem for coupled cell systems with ‘interior’ symmetry.
The secondary branch represented in Figure 28 appears through a secondary Hopf bifurcation, along the primary branch at the second Hopf bifurcation point (HB2). The tertiary branch occurs along the secondary branch at the third Hopf bifurcation point (HB3). In this parameter region the solution is periodic, with the same symmetry type as in the primary and secondary branches. Further away along this branch, the coupled cell system (3) evolves to complex dynamical behavior. First, quasiperiodic solutions start to be visible; these solutions are then followed, as the parameter c is increased, by chaotic patterns. This suggests the occurrence of several period-doubling bifurcations in the system.
For the networks in Figure 1(a), proceeding with the increment of the value of c, we note that the system is pulled back from chaos, and quasiperiodic motion reappears. The bifurcations associated with the latter changes in the qualitative behavior of a system are period-halving bifurcations. Incrementing c for the last time, the quasiperiodic solutions disappear, and the coupled cell system (3) again produces a stable
Relaxation oscillations are observed for the four networks in Figure 1, after the first Hopf bifurcation point. Relaxation oscillations are solutions characterized by long periods of quasi-static behavior interspersed with short periods of rapid transition, commonly seen in fast–slow systems (Krupa and Szmolyan, 2001). In previous work (Antoneli et al., 2008, 2010; Pinto, 2012), relaxation oscillations were found after the cascade of three Hopf bifurcations points. There, the authors have considered networks of two coupled rings of cells coupled to a ‘buffer’ cell (Antoneli et al., 2008, 2010) or coupled to each other (Pinto, 2012). One of the rings possessed
Thus, the small networks of one ring of cells coupled to a ‘buffer’ cell in Figure 1 reveal extraordinary dynamical features. Some of these features are certainly explained by the network architecture; nevertheless, the others, such as the route to chaos and returning from chaos, are most certainly attributed to the internal dynamics of the cells, in this case, the Chen oscillator. The Chen oscillator possesses a rich variety of dynamical features for certain parameter regions (Ueta and Chen, 2000; Lu et al., 2002; Li et al., 2004).
4. Conclusions
Four networks of one ring of three cells coupled to a ‘buffer’ cell with exact and ‘interior’
Footnotes
Acknowledgement
We thank the anonymous reviewers for their valuable comments and helpful suggestions.
Funding
The authors wish to thank Fundação Gulbenkian, through Prémio Gulbenkian de Apoio à Investigação 2003, and the Polytechnic of Porto, through the PAPRE Programa de Apoio à Publicação em Revistas Científicas de Elevada Qualidade, for financial support. The authors were partially funded by the European Regional Development Fund through the program COMPETE and by the Portuguese Government through the FCT – Fundação para a Ciência e a Tecnologia (project PEst-C/MAT/UI0144/2013). The research of Ana Carvalho was partially supported by the FCT (grant SFRH/BD/96816/2013).