
Abstract
Purpose:
A supplement, OPTIADE® DE, containing specific lactic acid bacteria, Enterococcus faecium WB2000, has been shown to provide significant ameliorative effects in patients. However, the mechanism of the supplement has not been elucidated. In this study, we investigated the mechanism of the supplement on dry eye symptoms using an air stress-induced dry eye mice.
Methods:
Stress-induced dry eye in mice was produced by exposing the mice to an air stream. OPTIADE® DE was administered for 5 days during the daily exposure to the air stress, and then the tear volume and the expression of specific mRNA were measured. Furthermore, the effects of WB2000 and the other nutritional ingredients were also investigated, respectively. The contribution of peroxisome proliferator-activated receptor α (PPARα) to the regulation of tear secretion was investigated by an inhibitor of PPARα.
Results:
Exposure of the mice to air stress displayed a remarkable decrease in the tear volume with a concomitant reduction in the PPARα expression in the lacrimal gland. OPTIADE® DE significantly inhibited the decrease in the tear volume and the expression of the PPARα. WB2000 and the other nutritional ingredients additively inhibited the decrease in the tear volume. Inhibition of PPARα completely canceled the effects of the OPTIADE® DE and WB2000 without influencing the effect of the mixture of other nutritional ingredients on the tear volume.
Conclusion:
These results suggest that both the PPARα-dependent pathway and the independent pathway contributed to the ameliorative effect of OPTIADE® DE in the dry eye mice.
Introduction
Dry eye disease (DED) is a multifactorial disease characterized by an instability of the tear film leading to ocular surface inflammation.1,2 Recently, new therapeutic drugs have been extensively investigated for DED. Topical medications have been reported to improve dry eye-caused symptoms, such as decreased tear volume, tear film instability, and inflammation. Furthermore, there has been an increase in the development of therapies targeting meibomian gland dysfunction. 2 Despite this increase in the development of newly available treatments, novel and effective treatments are still expected. With this background, microbiota has been considered as another target of dry eye treatment, in addition to topical medications. 3 Many reports suggested the effect of probiotics on the treatment of DED. For this reason, some food and biomaterials have been investigated to improve dry eye symptoms in human and animal models.4–6 A dietary supplement, OPTIADE® DE, containing lactoferrin, omega-3 fatty acids, lutein, and Enterococcus faecium WB2000, one of the lactic acid bacteria, has been reported to improve dry eye symptoms in humans. 7 The mechanism of this supplement in stress-induced dry eye symptoms in rats was investigated. However, the mechanism and contribution of the lactic acid bacteria in the therapeutic effect of this supplement have not been elucidated yet. In contrast, peroxisome proliferator-activated receptor α (PPARα), one of the nuclear receptor family, has recently been reported to be involved in the progression of eye disease, although PPARα has been considered to be involved in modulating cellular lipid metabolism, inflammation, and differentiation so far. Especially, PPARα likely contributed to the onset and development of some types of stress-induced dry eye.8,9 Therefore, we studied the contribution of PPARα pathway to the tear secretion in our experimental mouse model, since, similar to these reports, our present study used a stress-induced dry eye model in mice.
In this study, we investigated the effects of OPTIADE® DE and each nutritional ingredient of OPTIADE® DE on stress-induced dry eye symptoms in the dry eye mouse model. Furthermore, the mechanisms and contribution of each ingredient to the therapeutic effects of OPTIADE DE were considered, focusing on the expression and function of PPARα.
Methods
Animals
Male C57/BL6 mice (six-week-old, Charles River, Japan) were used in this study (n = 4∼8 in each experiment). Animals were housed under a 12-h light/12-h dark cycle (light phase: 7 am∼7 pm; dark phase: 7 pm∼7 am) in a temperature-controlled room (20∼26°C).
Drugs
OPTIADE DE (Wakamoto Pharmaceutical Co., Ltd.) encapsulated content was removed, suspended in distilled water, and then used in this study. GW6471, a specific PPARα inhibitor, was purchased from Abcam Limited (USA).
Mouse stress-induced dry eye model and the administration of the test substances
Stress-induced dry eye was induced in mice according to the method previously reported.10,11 Briefly, mice were physically restrained in a 50 mL plastic conical tube and exposed daily to a stream of air from the front side of the mice, at a rate of 0.5∼1.0 m/s for 4 h. The dietary supplement, OPTIADE® DE (Table 1), was suspended in distilled water and orally administered to mice at 10 mg/kg for 5 days before the stress exposure. Lactic acid bacteria and the other nutrients at an equivalent amount to those contained in OPTIADE® DE were also administered to mice. GW6471 was dissolved in distilled water and administered to mice at doses of 2 or 20 mg/kg before the stress exposure. Distilled water was administered as a vehicle control.
Composition of the Dietary Supplement, OPTIADE® DE

Measurement of tear volume
Basal tear secretion volume was measured with a cotton thread test using a phenol red-impregnated thread (Zone-Quick, Showa Yakuhin Kako, Tokyo, Japan). The mice were restrained without anesthesia, and a cotton thread was placed on the temporal side of the lower eyelid margin for 30 s. The length of the moistened fragment was measured. The measurements from the right eye were used for statistical analysis.
Measurement of the weight of the lacrimal gland, RNA extraction, and real-time PCR
Mice were anesthetized by inhalation of 4% isoflurane in oxygen, and then the right lacrimal gland was extracted and weighed. Mice were euthanized by cervical dislocation after the extraction of the tissues. The total RNA was extracted from the lacrimal gland and the hippocampus of the nontreated and stress-induced dry eye mice with or without administration of OPTIADE® DE using TRIzol™ Plus RNA Purification Kit (Thermo Fisher, USA), and then total RNA was inverted into cDNA using a PrimeScript™ RT Master Mix (Takara, Japan). The expression of mRNA was quantified using TB Green® Fast qPCR Mix (Takara, Japan) and a Thermal Cycler Dice® Real Time System III (Takara, Japan). The housekeeping gene β-actin was used as an internal standard to normalize the relative expression of target genes. Statistical difference was analyzed in the air stream group and the OPTIADE® DE group compared with the nontreated group as a control. The primers were designed and synthesized based on gene sequences available in the GenBank database as follows: PPARα, (F): 5′-AGAGCCCCATCTGTCCTCTC-3′, (R): 5′-ACTGGTAGTCTGCAAAACCAAA-3′: β-actin, (F): 5′-CATTGCTGACAGGATGCAGAAGG-3′, (R): 5′-TGCTGGAAGGTGGACAGTGAGG-3′, PPARγ, (F): 5′- TAGCTTGGCCATATTTATAGCTATCA-3′, (R): 5′-CGATGGGCTTCACGTTCAG-3′, brain-derived neurotrophic factor (BDNF) (total), (F): 5′-AAGGACGCGGACTTGTACAC-3′, (R): 5′-CGCTAATACTGTCACACACGC-3′, BDNF (exon1) (F):5′-CACATTACCTTCCTGCATCTGTTG −3′, (R): 5′-ACCATAGTAAGGAAAAGGATGGTCAT-3′, catalase, (F): 5′-AGTCTTCGTCCCGAGTCTCTC-3′, (R): 5′-CTGGTCGGTCTTGTAATGGAA-3′, HO-1, (F): 5′-CCTTCCCGAACATCGACAGCC-3′, (R): 5′-GCAGCTCCTCAAACAGCTCAA-3′, superoxide dismutase (SOD) 2, (F): 5′-GGCCAAGGGAGATGTTACAA-3′, (R): 5′-ACCCTTAGGGCTCAGGTTTG-3′.
Statistical analyses and justification of sample size
Statistical analysis data are presented as means and standard deviations, unless otherwise indicated. Statistical significance was calculated using the Student’s t-test, Steel–Dwass test, and one-way analysis of variance (ANOVA) or two-way analysis of variance with Dunnett’s multiple comparison test. The corrections for the multiple comparison test were made by the Bonferroni correction method.
Statement of ethics
This study was approved and monitored by the internal Animal Research Ethics Committee of Wakamoto Pharmaceutical Co., Ltd. All experiments were performed according to the ARVO Statement for the Use of Animals in Ophthalmical and Vision Research.
Results
In this study, a significant decrease in the tear volume was observed in control mice by exposing them to the stress induced by an airstream. Without stress, the tear volume level was approximately 3 mm in C57BL/6 mice. The tear volume started to decrease from the second day after the first stress was given. After the third day of stress treatment, the tear volume constantly showed 30% compared with untreated mice (P < 0.001 by Student’s t-test, n = 4), as already reported in another study10,11 (Fig. 1). When the dietary supplement OPTIADE® DE was administered, this decrease in the tear volume was significantly inhibited compared with the stress control group from the 2nd to 5th days of administration (Fig. 2). Simultaneously, stress-induced hypertrophy of the lacrimal gland was observed (P < 0.05 by one-way ANOVA with Dunnett’s multiple comparison test, n = 4, Fig. 3). An ameliorative effect of the supplement on the hypertrophy of the lacrimal gland was also observed, but it was not significant (Fig. 3). The mRNA expression of PPARα in the lacrimal gland decreased by 30% in stress-induced dry eye mice, and OPTIADE® DE counteracted the reduction of mRNA expression to the level of mice without stress, although these effects were not significant by one-way ANOVA with Dunnett’s multiple comparison test (n = 4, Fig. 4). The expression level of PPARα in the brain was constant before and after air stress (Supplementary Data S1). Notably, the inhibition of PPARα by preadministration of GW6471 dose-dependently canceled the effect of OPTIADE® DE on the stress-induced decrease in the tear volume from the 2nd to 5th days of administration in dry eye mice. The effects of GW6471 were significant at all measurement points from the 2nd day to the 5th day of the treatment (two-way ANOVA with Dunnett’s multiple comparison test, n = 4, Fig. 5B). GW6471 did not affect the tear volume in mice not exposed to the air stress (Fig. 5A). To investigate the mechanism of OPTIADE® DE in detail, the effects of one of the ingredients of OPTIADE® DE, WB2000, a specific lactic acid bacterium, and the mixture of the other nutritional ingredients, such as lactoferrin, zinc, eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), vitamin C, vitamin E, and lutein, were compared. Interestingly, WB2000 and the other nutritional ingredients additively ameliorated the decrease in the tear volume observed in the stress-induced dry eye mice. Briefly, other ingredients significantly increased the tear secretion from the 3rd to 5th days of administration compared with the stress control (P < 0.05 on the 4th day, P < 0.01 on the 3rd and 5th day by two-way ANOVA with Dunnett’s multiple comparison test, n = 4, Fig. 6). Increases in tear secretion were also observed from the 2nd to the 5th days of WB2000 administration, but the effect was significant only on the 5th day (P < 0.01, n = 4, Fig. 6). WB2000 plus other ingredients showed the most potent effects compared with the individual administration of WB2000 and other ingredients (P < 0.01 on the 4th day, P < 0.001 on the 3rd and 5th day, n = 4, Fig. 6). Furthermore, significant increases in the tear secretion observed from the 3rd day to the 5th day by the administration of WB2000 were canceled by GW6471 (Fig. 7A). In contrast, the effect of the mixture of other nutritional ingredients was not affected by the PPARα inhibition (Fig. 7B). PCR cycle conditions and the melting curves of each primer are shown in Supplementary Data S4 and S5.
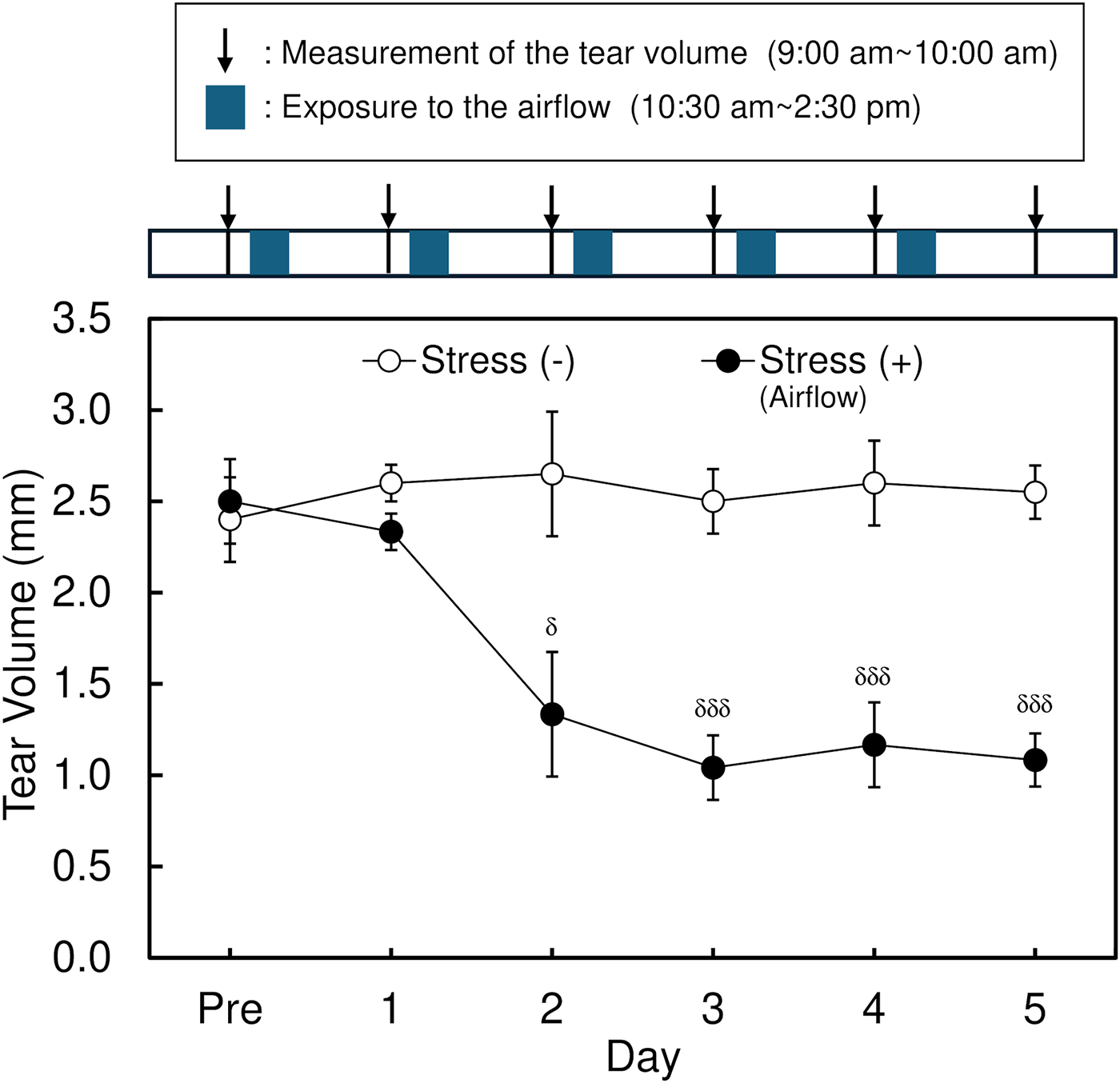
Effect of the air stress on the tear volume in mice. Each data represents the mean ± S.E. of 5∼6 mice. The mice were exposed to airflow from 10:30 am to 2:30 pm daily in the stress (+) group after measuring the tear volume. Symbols indicate the significant differences at each time point from the stress (-) at P < 0.05 (δ) and P < 0.001 (δδδ), respectively, by Student’s t-test.
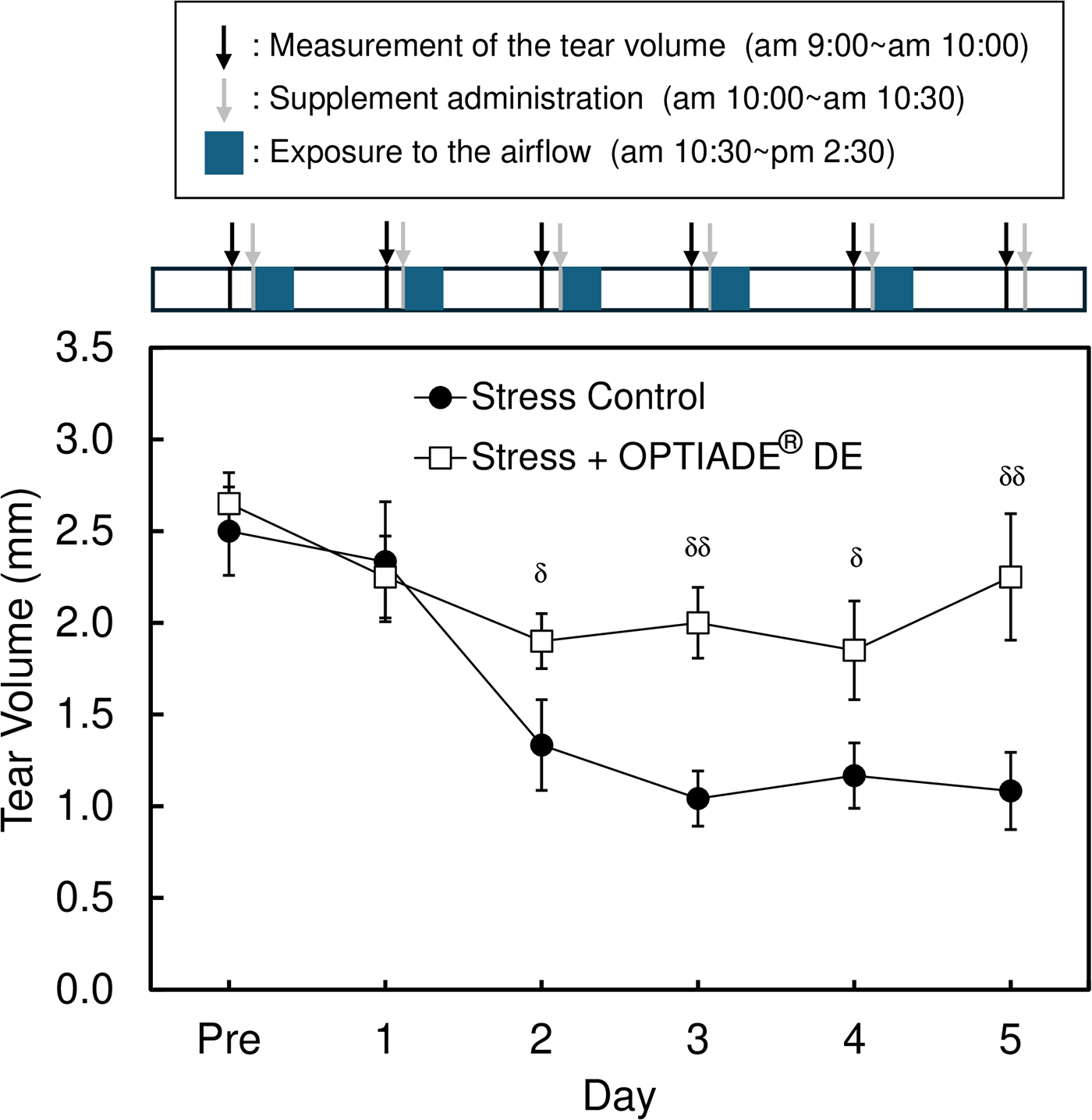
Effect of the dietary supplement (OPTIADE® DE) on the decrease in the tear volume in dry-eye model mice. Each data represents the mean ± S.E. of 5∼6 mice. The mice in the stress (+) group were exposed to airflow from 10:30 am to 2:30 pm daily, as shown in Figure 1. OPTIADE® DE was per-orally administered to mice before the exposure to the airflow. Symbols indicate the significant differences from the stress control at each time point at P < 0.05 (δ) and P < 0.01 (δδ), respectively, by Student’s t-test.
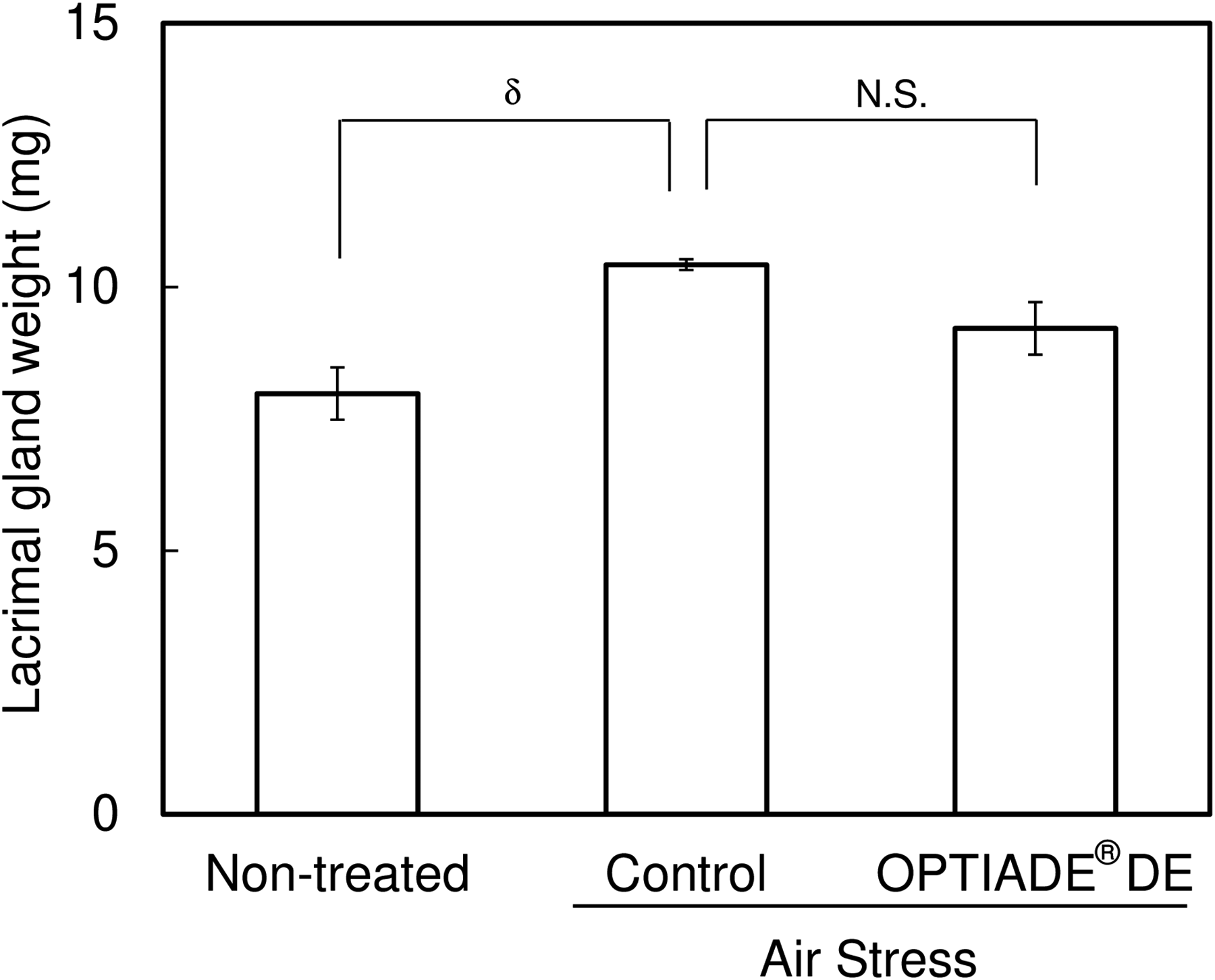
Effect of air stress and OPTIADE® DE administration on the weight of the lacrimal gland. Each column represents the mean ± S.E. of 4 mice. Symbol indicates the significant differences from the control at P < 0.05 (δ) by one-way analysis of variance with Dunnett’s multiple comparison test corrected by the Bonferroni correction method.
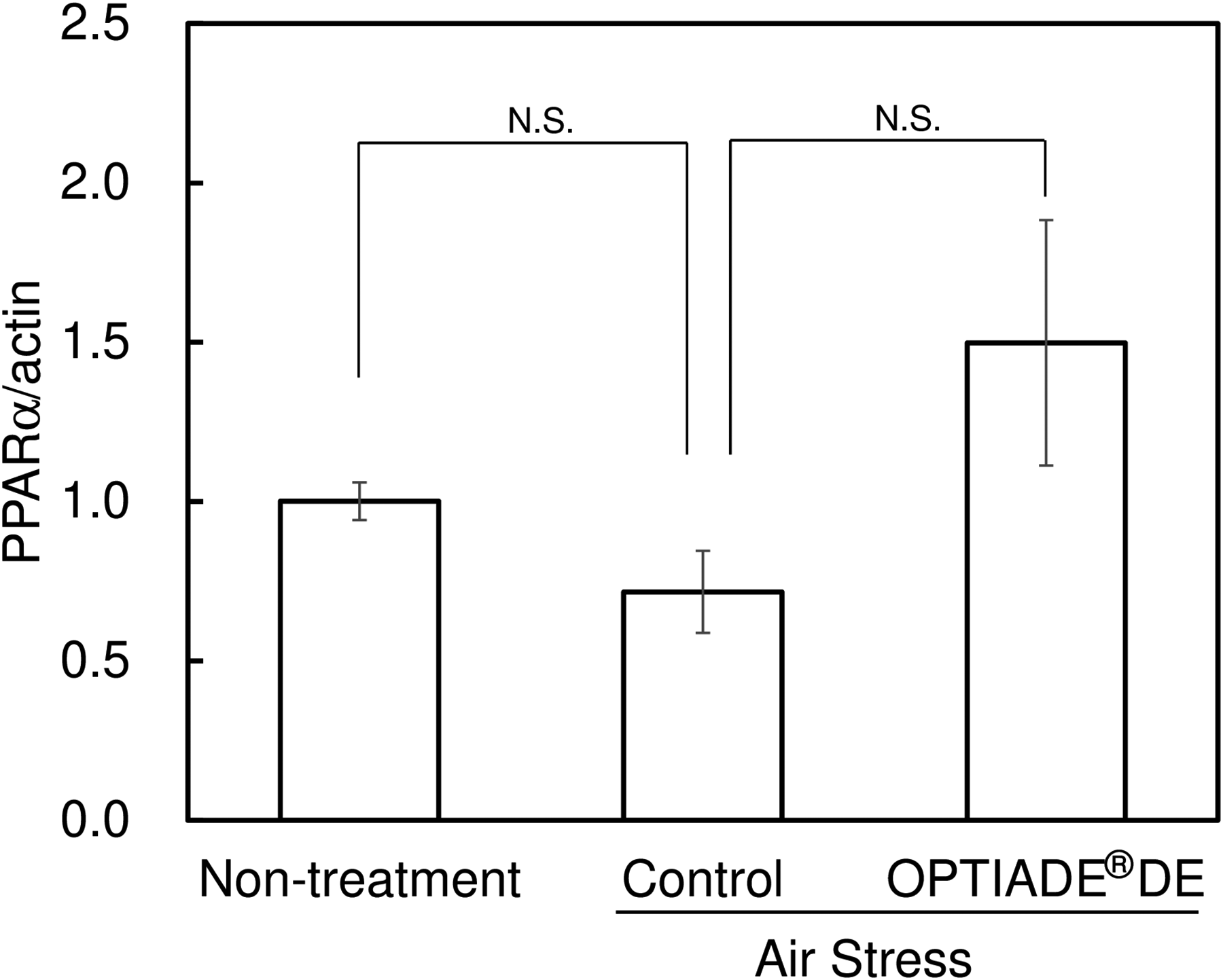
Effects of the air stress and the OPTIADE® DE on the expression of PPARα in the lacrimal gland in mice. Each column represents the mean ± S.E. of 4 mice. N.S. means nonsignificant difference from control by the Steel test corrected by the Bonferroni correction method.
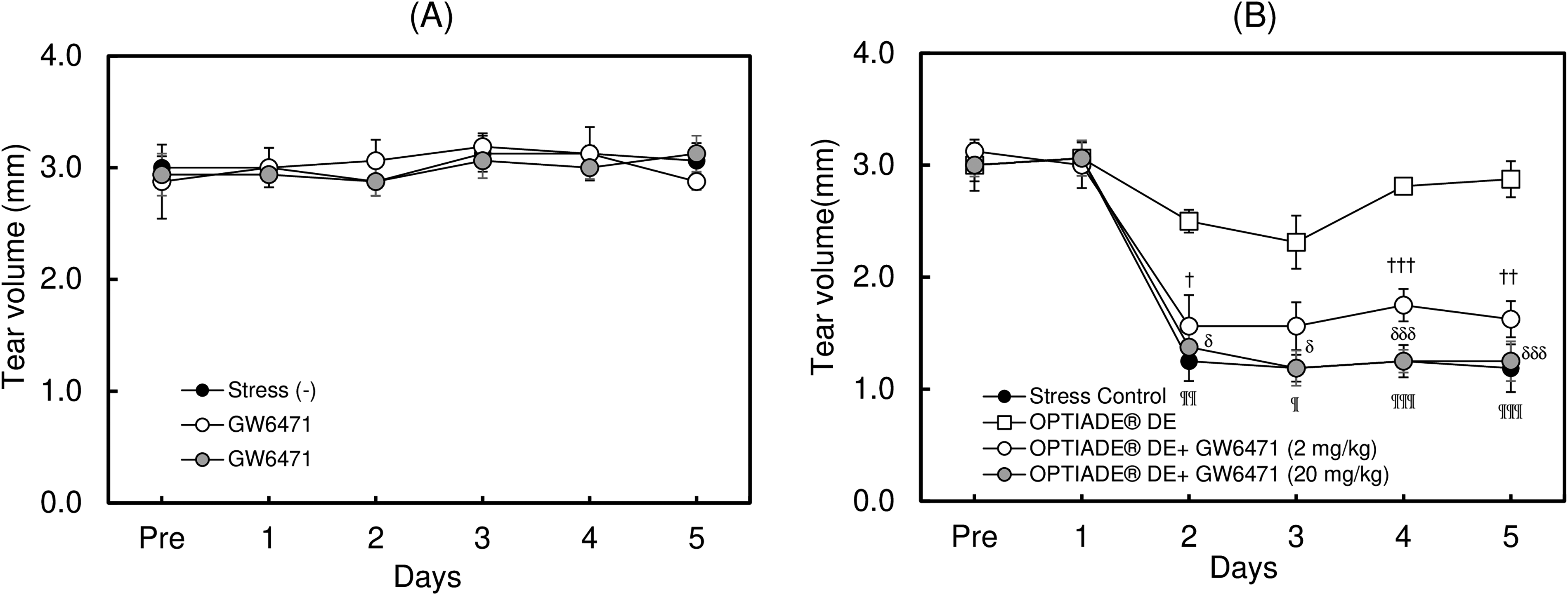
Effects of the inhibition of PPARα on the OPTIADE® DE-mediated improvement of the tear volume in mouse dry eye model. Effect of PPARα inhibitor alone
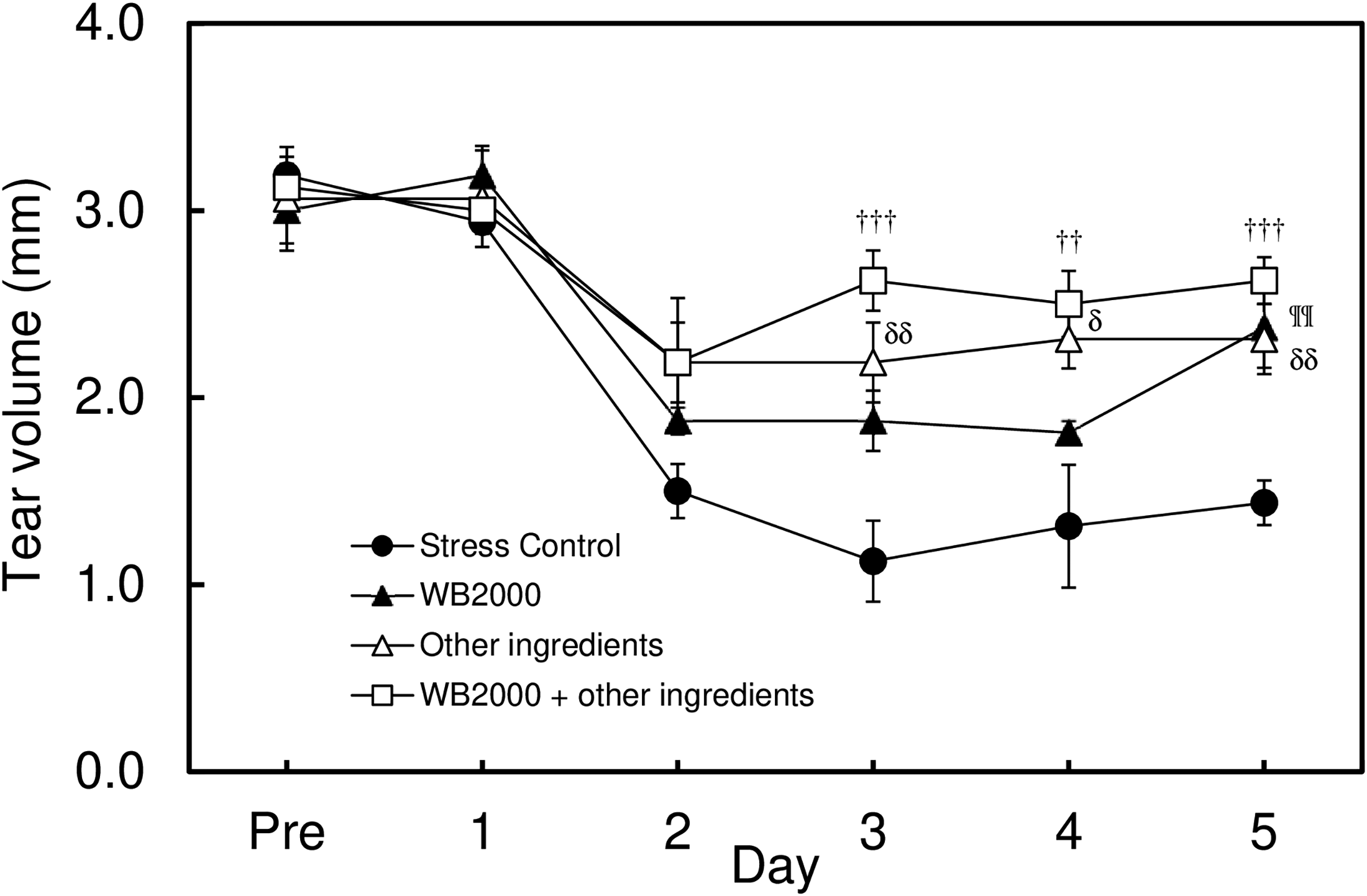
Effects of the ingredients of OPTIADE® DE, WB2000, and the other ingredients on the tear volume in the mouse dry-eye model. Each data represents the mean ± S.E. of 6 mice. The mice in the stress (+) group were exposed to airflow from 10:30 am to 2:30 pm daily, as shown in Figure 1. WB2000 and other ingredients were per-orally administered to mice before the exposure to the airflow. Symbols indicate significant differences at each time point from the stress control at P < 0.05 (δ), P < 0.01(††, ¶¶, δδ), and P < 0.001(†††), respectively, by two-way analysis of variance with Dunnett’s multiple comparison test corrected by the Bonferroni correction method.
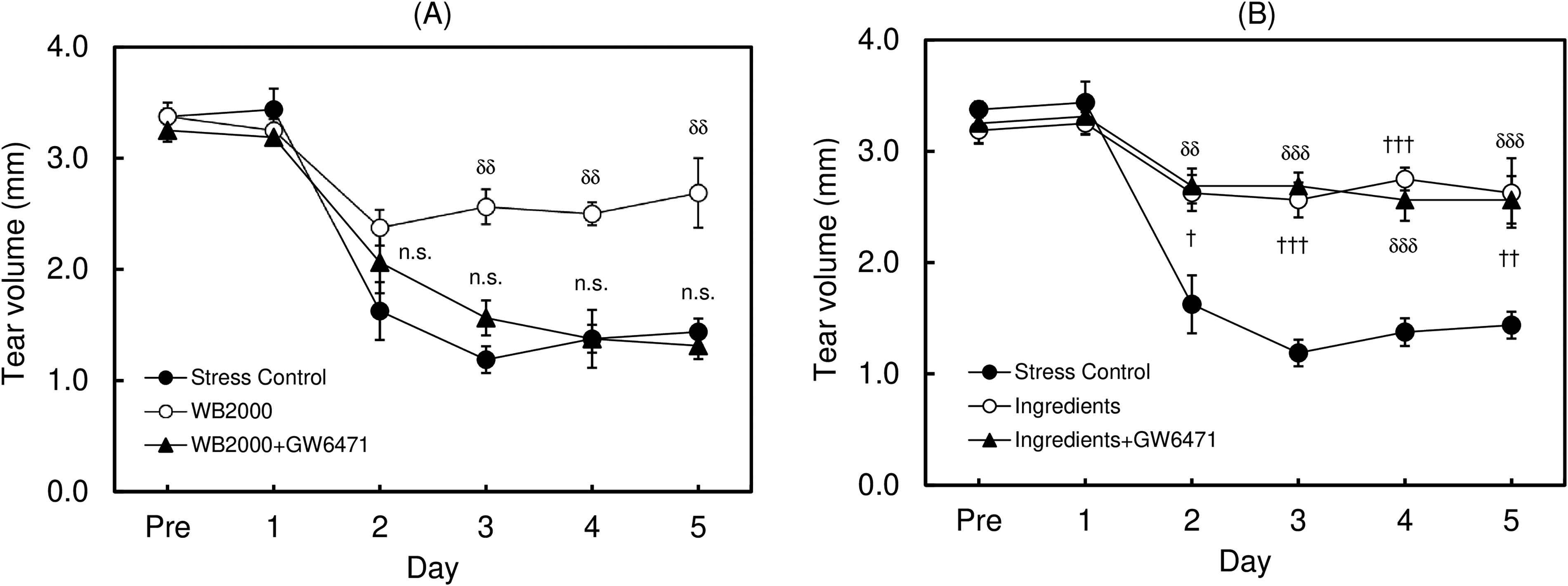
Contribution of PPARα to the ameliorative effect of WB2000 and the other ingredients on the decrease in the tear volume in the mouse dry eye model. Effect of inhibition of PPARα on the ameliorative effects of WB2000
Discussion
In this study, OPTIADE® DE significantly suppressed air stress-induced decrease in the tear volume in mice, as reported in a previous study. 7 The effect of the dietary supplement was diminished when administration was discontinued and then immediately returned by readministration in long-term experiments.12,13 As indicated in the results, the improvement of hypertrophy of the lacrimal gland was supposed to be involved in the effects of OPTIADE® DE in this dry eye model to some extent. 8 With these results, we focused on the mechanism of the ameliorative effects of OPTIADE® DE.
Notably, a reduction in the expression of PPARα in the lacrimal gland was observed in the air stress-induced dry eye mouse model. OPTIADE® DE inhibited the reduction of PPARα expression in the lacrimal gland observed in these mice. The results from the experiments that used GW6471, a specific PPARα inhibitor, showed that PPARα activation likely contributes to the effect of OPTIADE® DE on the decrease in the tear volume in these dry eye mice. These results indicate that the specific expression of PPARα in the lacrimal gland is related to the onset of the air stress-induced dry eye in mice to some extent. PPARs are ligand-activated transcription factors of the nuclear receptor superfamily that comprises three isotypes: PPARα, γ, and δ. Each PPAR subtype exhibits a distinct distribution pattern and controls the expression of various genes involved in lipid metabolism. 14 In the eyes, PPARα and PPARγ are expressed in the cornea, conjunctiva, retina, meibomian, and lacrimal glands, whereas PPARδ is expressed in the cornea, retina, and lacrimal glands. 15 Among these PPAR subtypes, the involvement of PPARα and PPARγ in the dry eye has already been investigated. Namely, fenofibrate, a PPARα agonist, has been reported to suppress the tear film instability and inflammation in the benzalkonium chloride-induced dry eye model rat. 16 PPARα also contributes to sleep deprivation-induced dry eye and Sjögren syndrome-associated dry eye in mice.9,17 Among the mechanisms of the ameliorative effects of PPARα activation on dry eye symptoms, the enhancement of tear production mediated by PPARα has been investigated.8,17 Since PPARα has been known for involvement in lipid metabolism, and the activation of PPARα results in transcriptional activation of the factors that regulate lipid catabolism and oxidation, we believed that the PPARα-mediated pathway improved the function of the lacrimal gland by regulating intracellular lipid homeostasis. In that study, the improvement of sleep-induced hypertrophy of the lacrimal gland was observed by PPARα activation, as observed in our study. Furthermore, fenofibrate increased the tear secretion by restoring the lacrimal gland function by suppressing inflammation through modulating Th1/Th17/Treg cell responses. 17
As described above, OPTIADE® DE demonstrated ameliorative effects in dry eye patients 7 at a dose equivalent to that used in mice, indicating that the PPARα-dependent pathway was activated equally in both species under both experimental conditions. However, there was a possibility that the difference in the expression level of PPARα in both species affected the intensity of OPTIADE® DE, although no report shows the difference between the species.
Since our additional experiments indicated the additive effect of WB2000 and the other nutritional ingredients on the dry eye symptom in mice, we investigated the contribution of PPARα pathway to the effect of WB2000 and other nutritional ingredients, separately.
As a result, similar to the results of OPTIADE® DE, the effect of WB2000 on the decrease in tear volume was completely canceled by the inhibition of PPARα, indicating that the PPARα pathway was upregulated by the administration of WB2000. Although upregulation of PPARα was observed by specific lactic acid bacteria, the mechanisms have not been elucidated.18,19 We supposed that middle-chain fatty acids, the internal ligand of PPARα, produced by lactic acid bacteria contribute to the upregulation of PPARα pathway.20,21 Recent computational analysis showed the potential of probiotic-derived metabolites as agents for modulating PPAR activity. 22 In their study, the in silico analysis demonstrates that the indole metabolite produced by bacteria showed stronger interactions with PPARs compared with fatty acids, as ligands. These metabolites may be absorbed by the gastrointestinal tract and distributed throughout the body and may show anti-inflammatory effects. These reports strongly suggested the involvement of the probiotics in the activation of PPARs. In contrast, unlike WB2000, the effect of the mixture of other nutritional ingredients was not affected by the inhibition of PPARα, indicating that the effect of these nutritional ingredients is not mediated by the PPARα pathway. Many reports suggested that the effects of nutrients, such as lactoferrin, zinc, EPA, DHA, vitamin C, vitamin E, and lutein, were mediated by each specific mechanism.23,24 Lactoferrin and lutein, known to act as antioxidants, showed beneficial effects by downregulating ROS production and upregulating SOD in dry eye patients.23,24 In addition, omega-3 fatty acids showed an improvement in dry eye symptoms by inhibiting pro-inflammatory cytokine production and T-lymphocyte proliferation in patients.1,23 The effects of these nutrients have been investigated by administering them alone or in combination, including vitamins. 24 Hence, we consequently supposed that these nutritional ingredients and WB2000 additively inhibited the stress-induced decrease in the tear volume by a distinct mechanism in this model. Interestingly, although WB2000 and the other ingredients were effective on the decrease in the tear volume by distinct mechanisms, and the effects of the ingredients were shown not to be mediated by the PPARα pathway, the effect of the concomitant administration of WB2000 and the other ingredients was completely inhibited by the PPARα inhibitor. From these results, we speculated that the main pathway that regulates the tear secretion integrates into a PPARα-dependent pathway when PPARα is activated, even though both PPARα-dependent and independent pathways exist in the regulation process of the tear secretion. Indeed, a cross talk of signaling pathways mediated by PPARα, β/δ, and γ has been reported. 25 Namely, PPARα activation by GW7647, a synthetic agonist of PPARα, suppressed the effect of PPARβ/δ and γ in the rat primary astrocytes. 25 From these reports, there is a possibility that the cross talk of PPAR-mediated pathways is involved in the mechanism of the regulation of tear secretion by concomitant administration of WB2000 and other nutritional ingredients in mice.
In addition, our preliminary study showed the possible involvement of BDNF, 26 specifically in the lacrimal gland, in the ameliorative effect of OPTIADE® DE (Supplementary Data S2). An experiment using BDNF knockout mice showed that BDNF plays an important role in basal tear secretion, and an inverse relation between the expression of tissue (brain and lacrimal gland) BDNF and the tear secretion levels was reported in stress-induced dry eye mice. 26 These results support our present results, despite our results indicating that only the lacrimal gland BDNF was involved in the tear secretion. Although the direct contribution of lacrimal gland BDNF to the improvement of tear secretion has not been elucidated, increased expression of BDNF is likely mediated by PPARα activation.27,28 Since the PPARα-dependent pathway is regulated by many internal ligands and the cAMP-related transcription factors,21,29 additional studies regarding the involvement of lacrimal gland BDNF in the tear secretion and the detailed mechanism of the regulation of BDNF expression through the activation of PPARα by the supplement are needed.
As described above, OPTIADE® DE and WB2000 increased the tear secretion mediating the PPAR-dependent pathway. Another report showed that PPARα improved dry eye symptoms by enhancing radical scavenging, regulating the expression of the oxidation factors, such as SOD, catalase, and reactive oxygen species (ROS).12 Furthermore, a decrease in retinal ROS and the downregulation of the expression of antioxidant inhibitors, such as thioredoxin-interacting protein, by the administration of PPARα agonist were observed in streptozotocin-induced diabetic animal.30,31 Contrary to the report, there were no relations between some oxidation-related parameters and the effects of OPTIADE® DE (Supplementary Data S3). Differences in some experimental conditions, especially the dose of WB2000 administered to mice, are supposed to affect the discrepancy of these data since a 100 times higher dose of WB2000 was administered in the previous study compared with our study.
Broadly speaking, the research on the utility of probiotics in the treatment of ocular diseases has been in the investigative stage during the last decade; our result shows the potential of probiotics for preventing and managing DED. Especially, the decrease in the tear secretion with morphological changes in the corneal sub-basal nerve caused by air stress is representative of chronic alterations caused by environmental stress.10,11 Hence, probiotics have benefits for the treatment of lifestyle-related dry eye symptoms among the causes and risk factors of dry eye. 32
Conclusions
WB2000, a lactic acid bacterium, and other nutritional ingredients contribute to the ameliorative effect of a diet supplement, OPTIADE® DE, in distinct mechanisms in air stress-induced dry eye mice.
Furthermore, both PPARα-dependent and independent pathways are complexly involved in the effects of these ingredients of OPTIADE® DE.
Authors’ Contributions
T.M. produced all the experiments: Conducting research and investigation process, specifically performing the experiments, and data/evidence collection. M.S. assisted with the experiments and preparing the article. R.A. planned all experiments, wrote the article, and prepared the figures.
Footnotes
Acknowledgments
The authors express their sincere thanks to Dr. Kazuo Tsubota, Dr. Shigeru Nakamura, and the members of the laboratory, Department of Ophthalmology, Keio University School of Medicine, Tokyo, for assistance in establishing the experimental protocol.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This research received no external funding.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.