
Abstract
Purpose:
Up to 46%–64% of individuals with diabetes exhibit corneal abnormalities, including epithelial dysfunction and reduced sensitivity, which may gradually impair corneal transparency. Corneal fibroblasts regulate scar formation and play a critical role in maintaining corneal transparency. This study investigated the effects of high-glucose levels on fibroblasts and elucidated the underlying mechanisms.
Methods:
Human corneal stromal fibroblasts were isolated by collagenase digestion of fresh corneal stromal lenticules, obtained from small incision corneal lens extraction surgery, and cultured with different glucose concentrations (5.5–30 mM). The phosphorylated platelet-derived growth factor receptor beta (p-PDGFRβ)/PDGFRβ) ratio, fibroblast proliferation, migration, fibrotic markers, and interleukin-6 (IL-6) expression were assessed.
Results:
Human corneal stromal fibroblast proliferation increased with increasing glucose concentrations, and a glucose concentration of 30 mM was selected for subsequent high-glucose treatment experiments. The p-PDGFRβ/PDGFRβ ratio, fibroblast proliferation, migration, fibrotic markers, and IL-6 expression were enhanced in the high-glucose group compared with those in the control group. These effects were inhibited by treatment with AG1295, a PDGFRβ tyrosine kinase inhibitor.
Conclusions:
In patients with diabetes, fibroblasts may aggregate at sites of corneal injury, potentially promoting scar hyperplasia and reducing corneal transparency. These changes may be reversed by PDGFRβ phosphorylation inhibitors, suggesting new therapeutic avenues.
Introduction
Diabetes is the most prevalent metabolic disorder, and the International Diabetes Federation projects that approximately 640 million individuals will be living with diabetes globally by 2040.1,2 This staggering figure underscores the urgent need to prioritize the prevention and treatment of diabetes as a critical public health priority.
Among the various complications associated with diabetes, diabetic keratopathy is frequently encountered, 3 affecting approximately 46%−64% of patients. 4 However, the manifestations are predominantly epithelial and neurotrophic in nature, including epithelial defects, decreased corneal sensitivity, reduced subbasal nerve density, and delayed epithelial wound healing.5,6 Persistent epithelial alterations, such as delayed wound healing, are a key feature of diabetic corneal disease. This impaired healing process increases susceptibility to vision-threatening complications, including secondary corneal injuries and potential scar formation. It also contributes substantially to the elevated risk of postoperative complications following procedures such as cataract extraction, corneal transplantation, and refractive surgeries. Patients with diabetes exhibit a notable decline in corneal transparency, 7 which correlates with the degree of glycemic control. 8 Corneal keratocytes play a pivotal role in maintaining corneal transparency.9,10 These specialized cells reside between the lamellar layers of the corneal stroma and typically remain in a quiescent state under normal conditions. However, upon injury, a cascade of cellular events is triggered. 11 Stromal cells at the site of damage undergo rapid apoptosis, inducing neighboring keratocytes to transform into fibroblasts. These activated cells migrate to the injured area, where they proliferate and initiate repair. 12 As fibroblasts accumulate at the wound edge, they further activate into myofibroblasts and secrete disorganized extracellular matrix components.13,14 This irregular matrix deposition leads to corneal scarring and ultimately results in loss of transparency. Therefore, controlling fibroblast accumulation and preventing their transformation into myofibroblasts during wound healing are crucial factors for minimizing corneal scaring and restoring visual function. 9 Consequently, this process has emerged as a key focus of corneal repair research. Nevertheless, the precise mechanisms by which hyperglycemia affects corneal stromal fibroblasts remain poorly understood. Elucidating the effects of high-glucose conditions on corneal fibroblasts may provide insight into the pathogenesis of diabetic corneal stromal lesions and support the development of novel strategies to improve corneal transparency in patients with diabetes.
Platelet-derived growth factor (PDGF), a key protein stored in platelets, acts as a crucial chemotactic agent for fibroblasts, facilitating their migration to wound sites.15,16 Its receptor, PDGFR, comprises three distinct domains: an extracellular N-terminal ligand-binding domain for PDGF recognition, a hydrophobic transmembrane domain, and an intracellular C-terminal domain with tyrosine kinase activity.17,18 PDGFR exists primarily in two isoforms, α and β, and elevated PDGFR expression in fibroblasts has been shown to be closely associated with scar formation. 19 Moreover, the activation of PDGF-PDGFR signaling is implicated in diabetes progression and its associated complications, 20 including insulin resistance, diabetic retinopathy, impaired wound healing, and renal tissue fibrosis. However, whether PDGFRβ signaling mediates high-glucose–induced fibrotic responses in human corneal stromal fibroblasts has not been fully elucidated.
Therefore, the objective of this study was to elucidate the mechanisms by which high glucose exacerbates corneal fibrosis through PDGFRβ signaling in human corneal stromal fibroblasts, and to evaluate the therapeutic potential of its inhibition.
Methods
Isolation, culture, and immunostaining of human corneal stromal fibroblasts
Corneal stromal tissue specimens were obtained from 40 eyes of 20 patients with myopia undergoing small incision lenticule extraction (SMILE) surgery at the Refractive Surgery Center of Tianjin Eye Hospital from August 2023 to June 2024. The study adhered to the Declaration of Helsinki guidelines and was approved by the Ethics Committee of Tianjin Eye Hospital (protocol number: KY-2023042; approval date: June 15, 2023). Written informed consent was obtained from all participants.
The cohort comprised 13 male and 7 female patients, aged 18–35 years, with preoperative spherical equivalent refractive errors ranging from −3.00 D to −10.00 D and astigmatism under 2.00 D. Exclusion criteria comprised keratoconus, ocular inflammation, thyroid dysfunction, prior ocular surgery, contact lens use, systemic or ocular diseases, and pregnancy or breastfeeding. After surgical aseptic extraction of corneal stromal lenticules, the samples were immediately placed in a sterile Dulbecco’s Modified Eagle’s Medium (DMEM)/F12 (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) culture medium at 4°C and transported to the laboratory for fibroblast isolation within 1 h. For stromal cell isolation, lenticular tissues were subjected to enzymatic digestion using 2 mg/mL type I collagenase at 37°C. The resultant cell suspension was centrifuged (1,500g, 5 min, 4°C), and the cell pellet was resuspended and seeded into T25 culture flasks. Cell cultures were sustained in complete DMEM/F12 medium (Gibco) enriched with 10% fetal bovine serum (FBS) and 1% antibiotics. Upon achieving a cell density of 4 × 106 cells per flask, the cultures were passaged, and cells from passages 3–5 were used for subsequent experiments. Cells from different experimental groups were derived from the same batch of primary cells.
Cell morphology was assessed via microscopic imaging. Vimentin and Aldehyde dehydrogenase-1A1 (ALDH1A1), well-established markers for corneal fibroblasts, were used to confirm the identity of cultured cells. Passage-3 cells were cultured in 48-well plates until they reached 70% confluency for fibroblast verification. After three 5-min washes with phosphate-buffered saline (PBS), cells were fixed with 4% paraformaldehyde for 15 min and rinsed with PBS. Permeabilization was performed using 0.2% Triton X-100 for 20 min, followed by another round of PBS washing. Nonspecific binding sites were blocked with 5% bovine serum albumin for 30 min at 15°C, followed by an overnight incubation at 4°C with the following primary antibodies: Vimentin (1:100 dilution, WL01960, Wanleibio, China) and ALDH1A1 (1:100 dilution, 60171-Ig, Proteintech, USA). After re-equilibration to 15°C for 1 h and thorough PBS washes (5 washes, 5 min each), cells were incubated with the following secondary antibodies for 2 h at 15°C: goat anti-rabbit (1:100 dilution, SA00003-2, Proteintech, USA) and goat anti-mouse secondary antibody (1:100 dilution, SA00013-1, Proteintech, USA). Following additional PBS washes, samples were mounted in a DAPI-containing medium and imaged using a fluorescence microscope (Zeiss, Oberkochen, Germany).
Measurement of aqueous and blood glucose levels in rats
Twelve healthy male Sprague–Dawley rats (8 weeks old, specific pathogen-free, weighing 260–300 g) were purchased from Changzhou Caverns Experimental Animal Co., Ltd. (Jiangsu, China). Following a 1-week acclimatization period, diabetes was induced in the experimental group via intraperitoneal injection of streptozotocin (55 mg/kg in 10 mg/mL citrate buffer). Control rats received equivalent volumes of citrate buffer alone. Diabetes confirmation required tail vein blood glucose levels ≥16.7 mM. After the successful establishment of the model, rats were randomly allocated to three groups: healthy control (n = 4), diabetic (n = 4), and AG1295-treated diabetic groups (n = 4), which received topical ocular administration with AG1295 (a PDGFRβ tyrosine kinase inhibitor) eye drops at a concentration consistent with that used in the cellular experiments.
All experiments were performed in accordance with the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research. All animals were maintained at the Animal Center of the Second Affiliated Hospital of Anhui Medical University, with the study protocol approved by the hospital’s Institutional Animal Care and Use Committee. Fasting plasma glucose and body weight were measured once weekly. Upon administration of a lethal dose of anesthetic, rats were euthanized via an intraperitoneal injection of an overdose of pentobarbital sodium (150 mg/kg), and the relevant tissues were excised for analysis.
Determination of the effects of glucose on human corneal stromal fibroblast proliferation
The cell-counting-kit-8 (CCK8) assay was used to evaluate the effects of glucose on human corneal stromal fibroblast proliferation. Corneal fibroblasts in the logarithmic growth phase were trypsinized and resuspended in a DMEM/F12 medium supplemented with 10% FBS to obtain a density of 15 × 106 cells/L. The cell suspension was seeded into 96-well plates at 100 μL per well. After 12 h of culture, the medium was replaced with a serum-free DMEM/F12 for a 12-h starvation period, after which the medium was discarded. The control group was treated with 100 μL of DMEM/F12 medium containing a physiological glucose (D-glucose, 9637, Hushi, China) concentration of 5.5 mM. The experimental groups were treated with either 15, 30, or 50 mM glucose. Each concentration was tested in triplicate, and an additional blank control group (DMEM/F12 medium without cells) was included for background correction. Following a 24 h incubation, 10 μL of the CCK8 reagent was added to each well, and the plates were incubated for an additional 2 h. Absorbance was measured at 450 nm using a microplate reader (Thermo Fisher Scientific), and the optical density (OD) values were recorded. The proliferation rate was calculated as follows:

The glucose concentration that yielded the highest proliferation rate was used for subsequent high-glucose experiments.
Determination of the optimal concentration of AG1295
AG1295 was obtained from the MCE Company (China). To determine the optimal concentration of AG1295, human corneal stromal fibroblasts in the logarithmic growth phase were trypsinized and resuspended in DMEM/F12, supplemented with 10% FBS at a density of 15 × 106 cells/L. The cell suspension (100 μL per well) was seeded into 96-well plates. After 12 h of culture, the medium was replaced with serum-free DMEM/F12 for a 12-h starvation period, after which the medium was discarded.
Cells were divided into five experimental groups: (1) control, treated with 100 μL of complete culture medium; (2) high-glucose, treated with 100 μL of complete culture medium containing the highest glucose concentration that yielded the highest proliferation rate in the previous step; (3) high-glucose + 5 μM AG1295 group, treated with 100 μL of complete culture medium containing high glucose and 5 μM AG1295; (4) high-glucose + 10 μM AG1295, treated with 100 μL of complete culture medium containing high glucose and 10 μM AG1295; and (5) high-glucose + 20 μM AG1295, treated with 100 μL of complete culture medium containing high glucose and 20 μM AG1295.
Each concentration was tested in triplicate, and an additional blank control group (containing only complete culture medium without cells) was included for background correction. A volume of 100 μL was added to each well, and the plates were incubated at a constant temperature (37°C) for 24 h. After incubation, 10 μL of the CCK8 reagent was added to each well, and the plates were returned to the incubator for an additional 2 h. Absorbance was measured at 450 nm using a microplate reader (Thermo Fisher Scientific), and OD values were recorded. The proliferation rate was calculated using equation (1).
The lowest concentration of AG1295 that significantly affected cell proliferation was selected for subsequent drug treatment experiments.
5-Ethynyl-2′-deoxyuridine assay-based experimental detection of the percentage of proliferative cells
Human corneal stromal fibroblasts (1 × 105 cells) in the log phase were trypsinized and seeded in 96-well plates. Upon reaching 70% confluence, cells underwent serum starvation for 12 h before treatment with the following experimental media: control DMEM/F12, high-glucose (30 mM) DMEM/F12, or high-glucose (30 mM) DMEM/F12 supplemented with AG1295. After 12 h of incubation, cell proliferation was evaluated by adding a 2 × EdU working solution for 2 h. Cells were then fixed in 4% paraformaldehyde, permeabilized with 0.3% Triton X-100 for 15 min at 15°C, and incubated in the dark for 30 min to facilitate the click chemistry reaction. Each step was followed by three PBS washes. Nuclei were then counterstained with a DAPI-containing mounting medium. Finally, fluorescence microscopy was performed to determine the proliferation rates by calculating the percentage of EdU-positive cells relative to the total DAPI-stained nuclei.
Cell migration assays
Cell migration was evaluated using scratch and Transwell assays across three experimental groups: control (complete medium with 10% FBS), high-glucose (complete medium with 10% FBS plus 30 mM glucose), and high-glucose with AG1295 treatment (complete medium with 10% FBS, 30 mM glucose, and AG1295).
Fibroblasts (1 × 105 cells) in the logarithmic growth phase were trypsinized and seeded onto 12-well plates. Upon reaching confluence, a uniform scratch wound was created using a 200-μL pipette tip, followed by gentle PBS washes to remove detached cells. The medium was replaced with serum-free DMEM/F12. Cell migration was recorded at 0, 24 h, and 48 h post-scratching. The migration closure was calculated as follows:

After 24 h of serum starvation, 1 × 105 fibroblasts in the log phase were trypsinized and plated in Transwell inserts. The experimental setup consisted of 200 μL DMEM/F12 in the upper chamber and 1 mL of test medium in the lower chamber. After 24 h of culture, the medium was removed, and the cells were washed, fixed with 4% paraformaldehyde, and stained with crystal violet (500 μL) for 15 min. Non-migratory cells remaining in the upper chamber were carefully removed through cotton swab abrasion, followed by two PBS washes. Migrated cells, stained with crystal violet, were quantified using microscopic examination to analyze the migration rate.
Western blot analysis
Western blot analysis was performed to determine the expression of p-PDGFRβ (C63G6), PDGFRβ (28E1), p-AKT(66444-1-Ig), AKT(10176-2-AP), alpha smooth muscle actin (α-SMA; 14395-1-AP), fibronectin (FN; 15613-1-AP), and interleukin-6 (IL-6; D5W4V) in the control, high glucose, and high glucose + AG1295 groups. Antibodies against these proteins were all purchased from Proteintech (Rosemount, Chicago, IL, USA). After extracting cellular proteins and quantifying their concentrations using the bicinchoninic acid assay kit (Beyotime, Nantong, China), samples were denatured and either processed immediately or stored at −20°C.
Proteins were separated using 10% sodium dodecyl sulfate — polyacrylamide gel electrophoresis gels. Membranes were blocked with 5% nonfat milk for 30 min at 15°C, followed by an overnight incubation at 4°C with the following primary antibodies: PDGFRβ, α-SMA, FN, p-AKT, AKT and IL-6 at 1:1000 dilution, and p-PDGFRβ at 1:500. After thorough washing, the membranes were incubated with HRP-conjugated secondary antibodies for 30 min at 15°C (1:5000 in TBS). Protein bands were detected using chemiluminescence and quantified using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
Each experimental group included three replicate wells, and the results are expressed as mean ± standard deviation (x ± s). Data were analyzed using SPSS 26.0 software (SPSS Inc., Chicago, IL, USA). A repeated-measures analysis of variance (ANOVA) was performed using the F-test, and pairwise comparisons among the groups were conducted using t-tests. In addition, one-way ANOVA followed by Tukey’s post-hoc test was used to evaluate differences between groups. Statistical significance was set at P < 0.05.
RESULTS
Morphology and vimentin staining of human corneal stromal fibroblasts
Human-derived corneal stromal tissue samples, obtained using SMILE surgery, were digested with collagenase, and the subsequently isolated cells were cultured. After 3–5 passages, human corneal stromal cells exhibited a spindle-shaped or hexagonal morphology (Fig. 1A). Vimentin (Fig. 1B) and ALDH1A1 (Fig. 1C) staining of the passage-3 cells confirmed that the extracted cells were human corneal stromal fibroblasts.
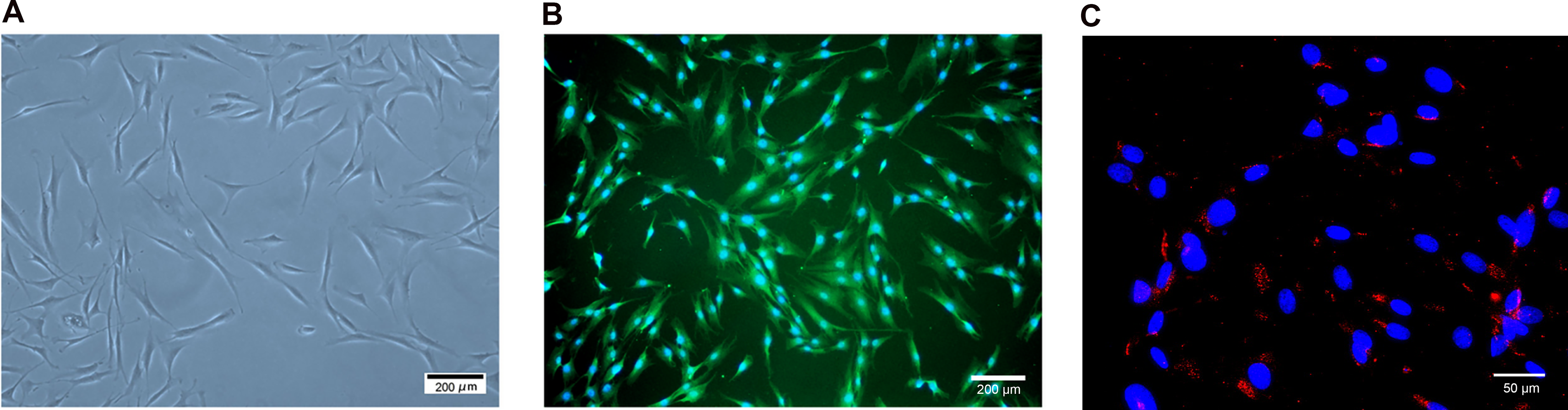
Morphological and immunophenotypical features of human corneal stromal fibroblasts (Passage-3). Corneal stromal fibroblasts exhibit a spindle-shaped or hexagonal morphology. Scale Bar: 200 µm
Effects of glucose concentration on corneal fibroblast proliferation determined using the CCK8 assay
We assessed the effect of different glucose concentrations on corneal fibroblast proliferation using the CCK8. Following treatment with glucose concentrations ranging from 5.5 mM to 30 mM for 24 h, the proliferation rate increased in a dose-dependent manner. However, when the glucose concentration was increased to 50 mM, the proliferation rate began to decline. The highest absorbance, indicating peak proliferation, was observed at a glucose concentration of 30 mM, with significant differences between the groups (Fig. 2). Although no studies have directly quantified glucose levels in the diabetic corneal stroma, stromal glucose concentrations can be reasonably inferred from measurements in tear fluid and peripheral blood, as corneal stromal cells primarily obtain glucose from these sources. In the control group, glucose concentrations in the aqueous humor and caudal vein were 4.65 ± 0.72 and 4.7 ± 0.66 mM, respectively. In the diabetes group, the corresponding values were 23.65 ± 1.5 and 24.33 ± 2.8 mM. Both aqueous humor and caudal vein glucose levels differed significantly between the control and diabetes groups (P < 0.05 for both).
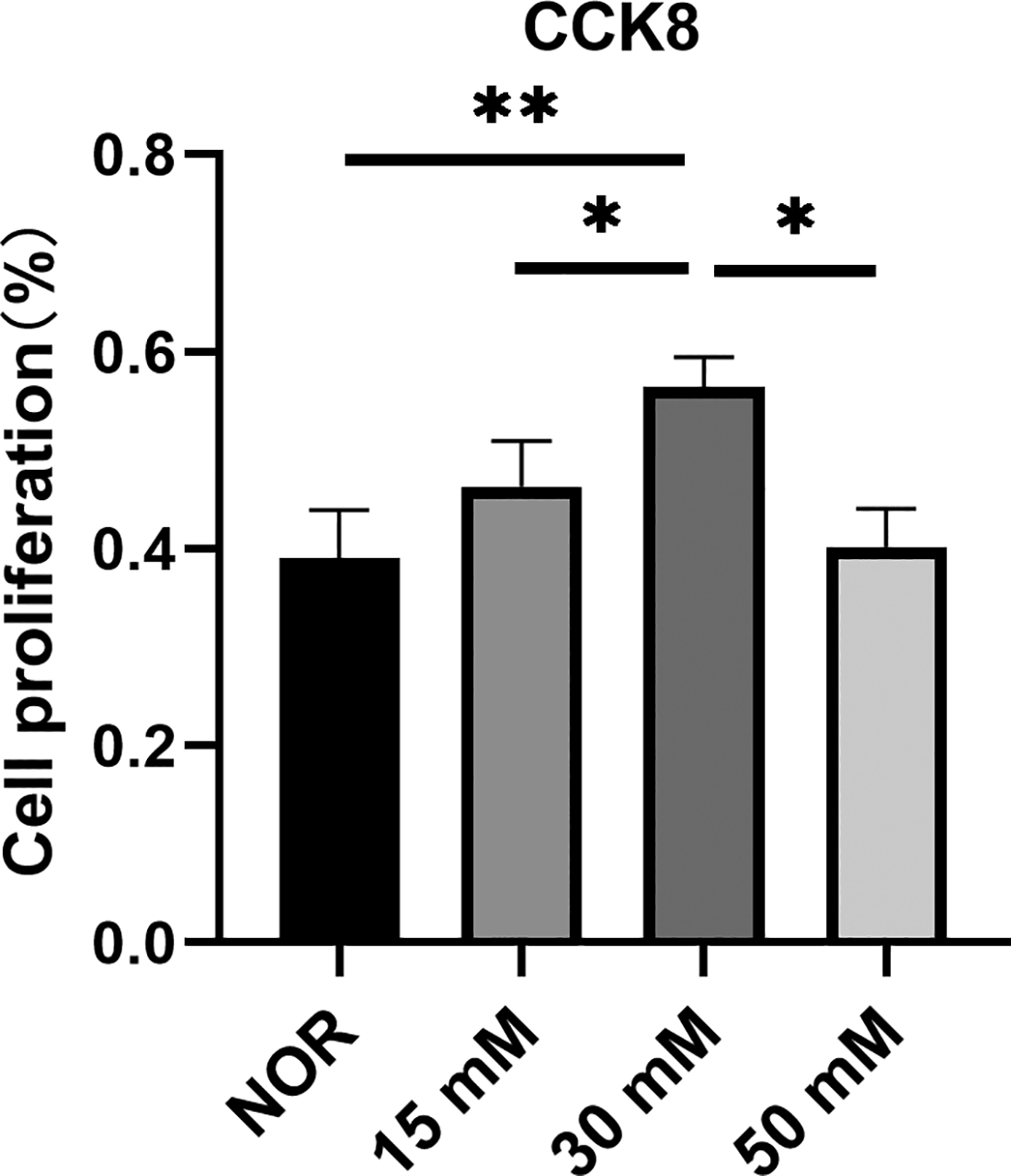
Changes in the proliferation rate of corneal fibroblasts after treatment with different glucose concentrations. Bars represent the mean ± SEM of triplicate determinations. *P < 0.05, **P < 0.01. CCK8, cell-counting kit-8; NOR, normal control group; SEM, standard error of the mean.
Based on the combined findings from the CCK-8 assay and animal experiments, a glucose concentration of 30 mM was selected as the optimal condition for establishing a high-glucose treatment model for human corneal stromal fibroblasts.
AG1295 does not affect the levels of glucose or body weight of diabetic rats
Data for body weight and glucose levels (both in plasma and aqueous humor) are presented in Figure 3. Fasting blood glucose levels were significantly higher in all diabetic groups compared with the normal control group. Furthermore, no significant difference in fasting glucose levels was observed between AG1295-treated and untreated diabetic rats. Normal rats demonstrated a steady gain in body weight throughout the experimental period, whereas diabetic rats showed a persistent decline, and this pattern was also not altered by treatment with AG1295.
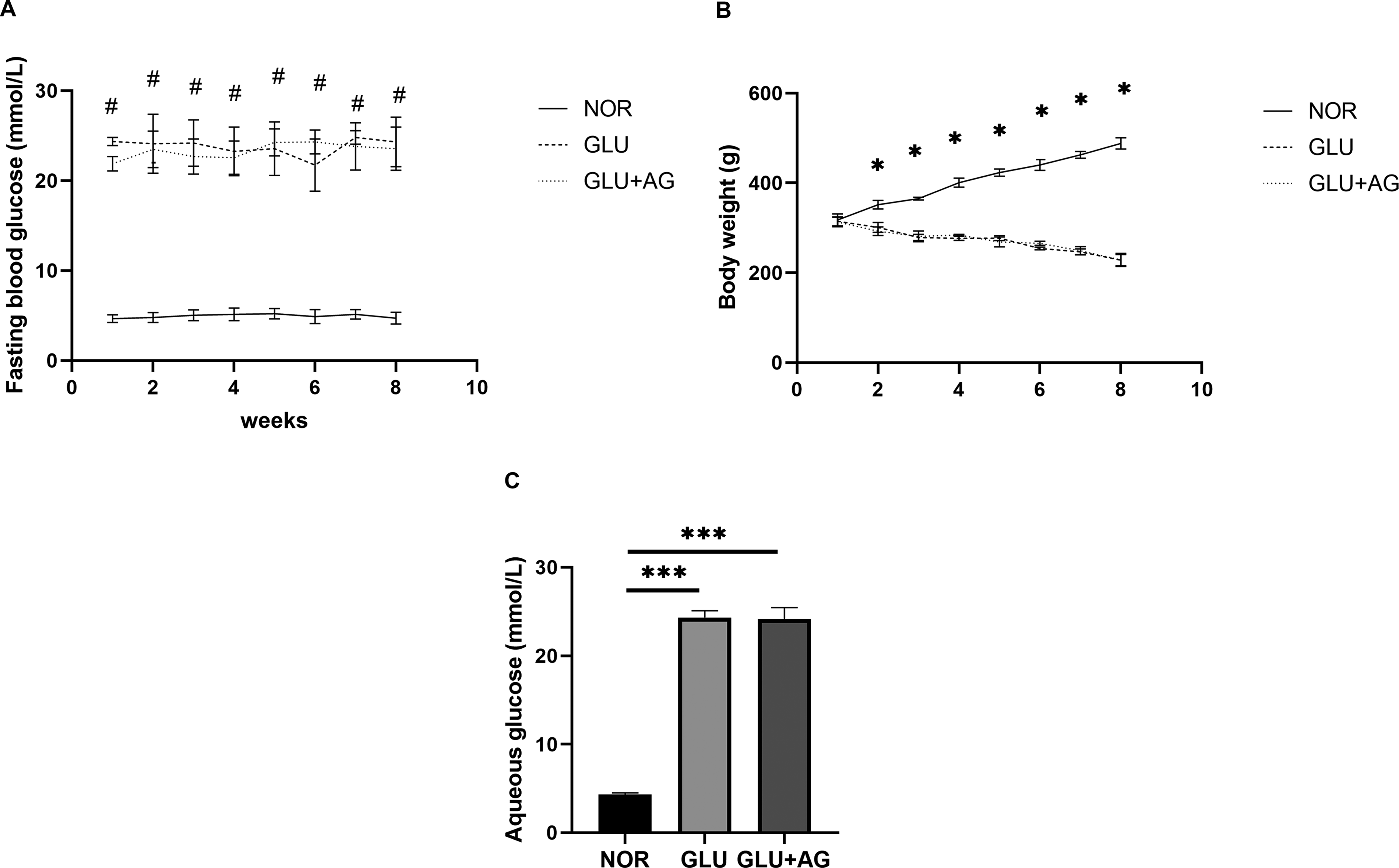
Changes in rat body weight, fasting plasma glucose levels (measured from the tail vein), and aqueous humor glucose concentrations.
AG1295 inhibits high-glucose-induced PDGFRβ and AKT phosphorylation in human stromal fibroblasts
Western blot analysis showed that high glucose significantly increased PDGFRβ and AKT phosphorylation in human corneal stromal fibroblasts (Fig. 4A,B). However, treatment with AG1295 significantly inhibited PDGFRβ and AKT phosphorylation induced by high-glucose treatment, indicating that glucose modulates this pathway.
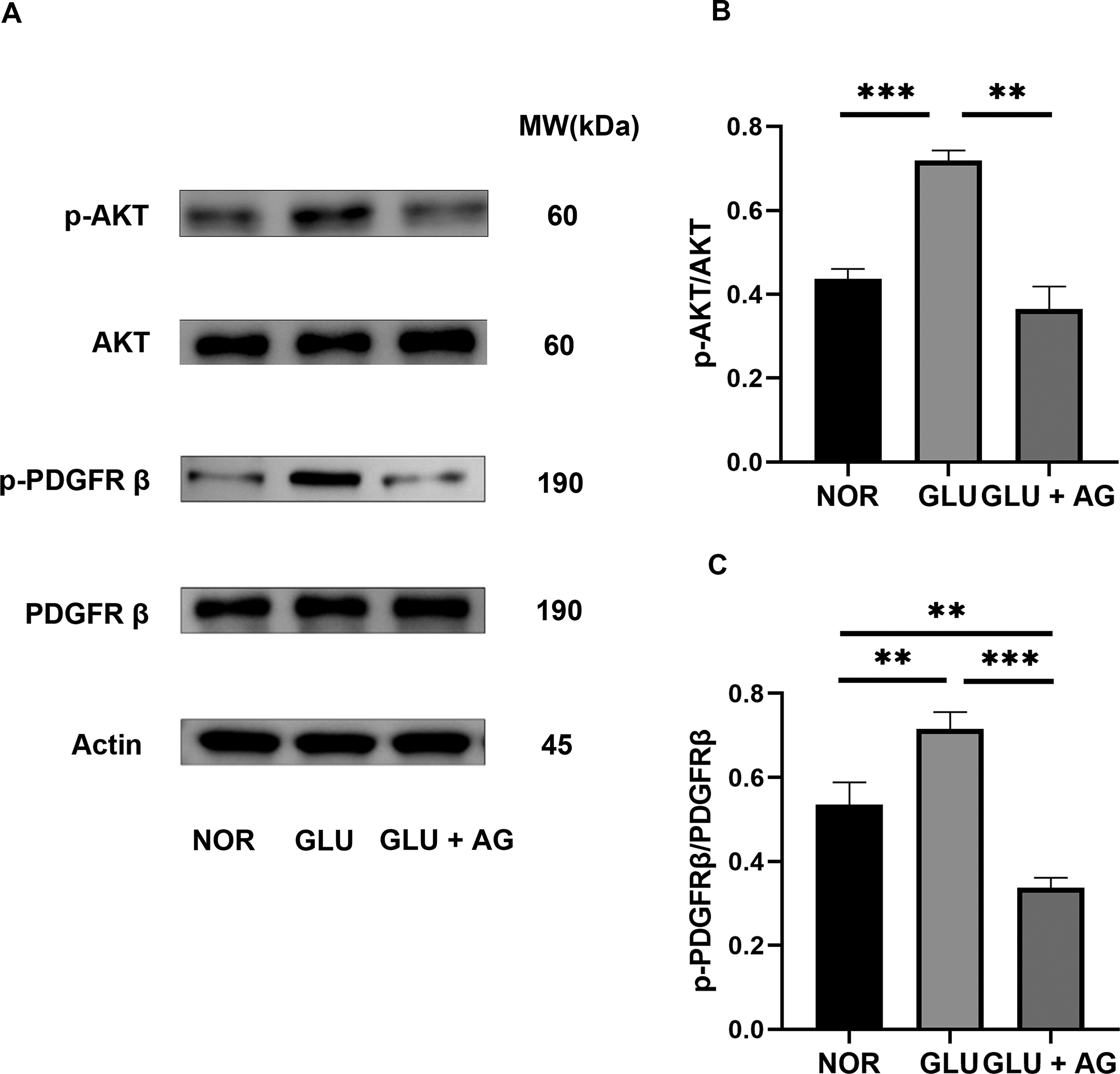
AG1295 inhibits high-glucose-induced AKT
AG1295 effectively suppresses high-glucose-induced proliferation in human corneal stromal fibroblasts
Treatment with AG1295 significantly suppressed the high-glucose-induced proliferation of human corneal stromal fibroblasts, with high AG1295 concentrations exhibiting a more pronounced inhibitory effect on cell proliferation (Fig. 5A). Therefore, we selected the minimum effective concentration of AG1295 (10 µM) for subsequent drug treatment experiments.
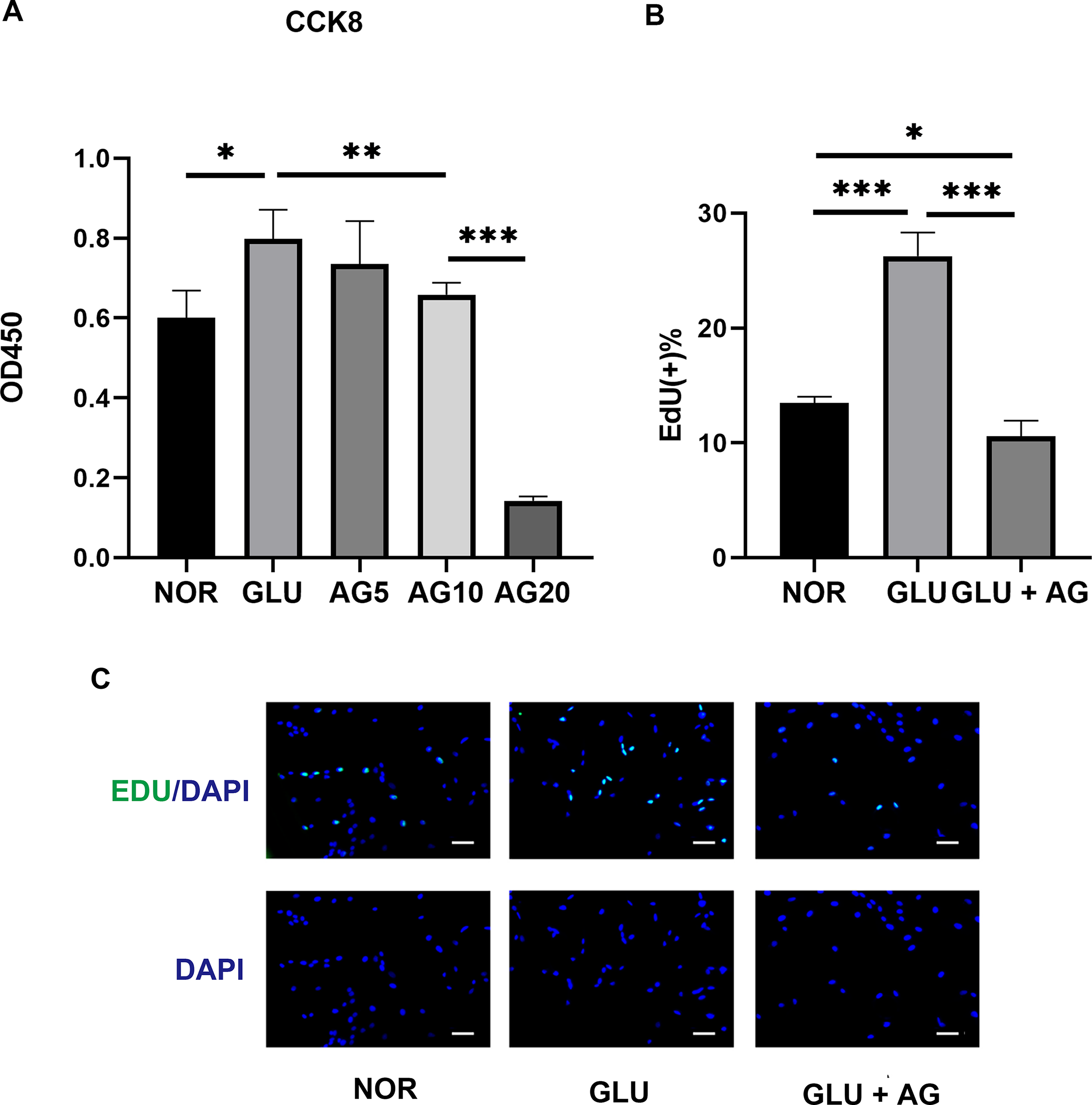
AG1295 Effectively Suppresses High-Glucose-Induced Proliferation in Human Corneal Stroma Fibroblasts, as assessed using the CCK8 assay, following treatment with varying concentrations of AG1295
The EdU assay results revealed that high glucose significantly increased the percentage of proliferating cells within the total cell population, as evidenced by an increase in the number of green fluorescent EdU-positive cells. However, AG1295 significantly reduced this proliferative response by decreasing the ratio of proliferating cells to the total cell count (Fig. 5B,C).
AG1295 effectively suppresses high-glucose-induced migration in human corneal stromal fibroblasts
Scratch and Transwell assays indicated that while exposure to high glucose conditions significantly enhanced the migration capacity of the fibroblasts. Nevertheless, treatment with AG1295 significantly attenuated this effect (Fig. 6). These findings indicate that AG1295 effectively inhibited the high-glucose-induced migration of human corneal stromal fibroblasts.
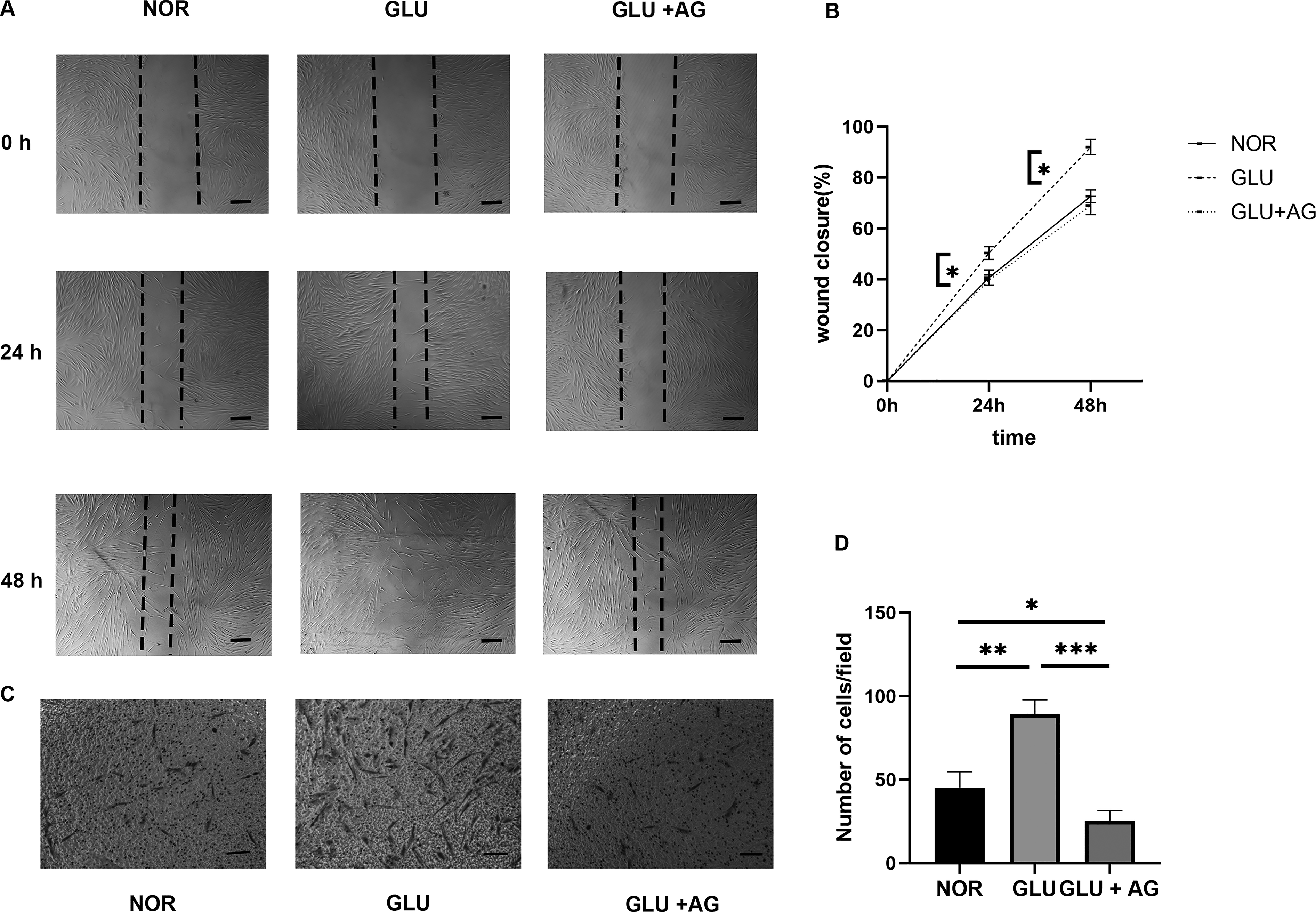
AG1295 Effectively Suppresses High-Glucose-Induced Migration in Human Corneal Stroma Fibroblasts. Scratch assays
AG1295 effectively suppresses hyperglycemia-induced phenotype transition of human corneal stromal fibroblasts
Both α-SMA and FN are well-established molecular markers indicative of myofibroblast differentiation, with their expression levels increasing as corneal stromal cells differentiate into myofibroblasts. When exposed to 30 mM glucose, α-SMA and FN expression levels increased; however, these increases were significantly mitigated by AG1295 (Fig. 7).
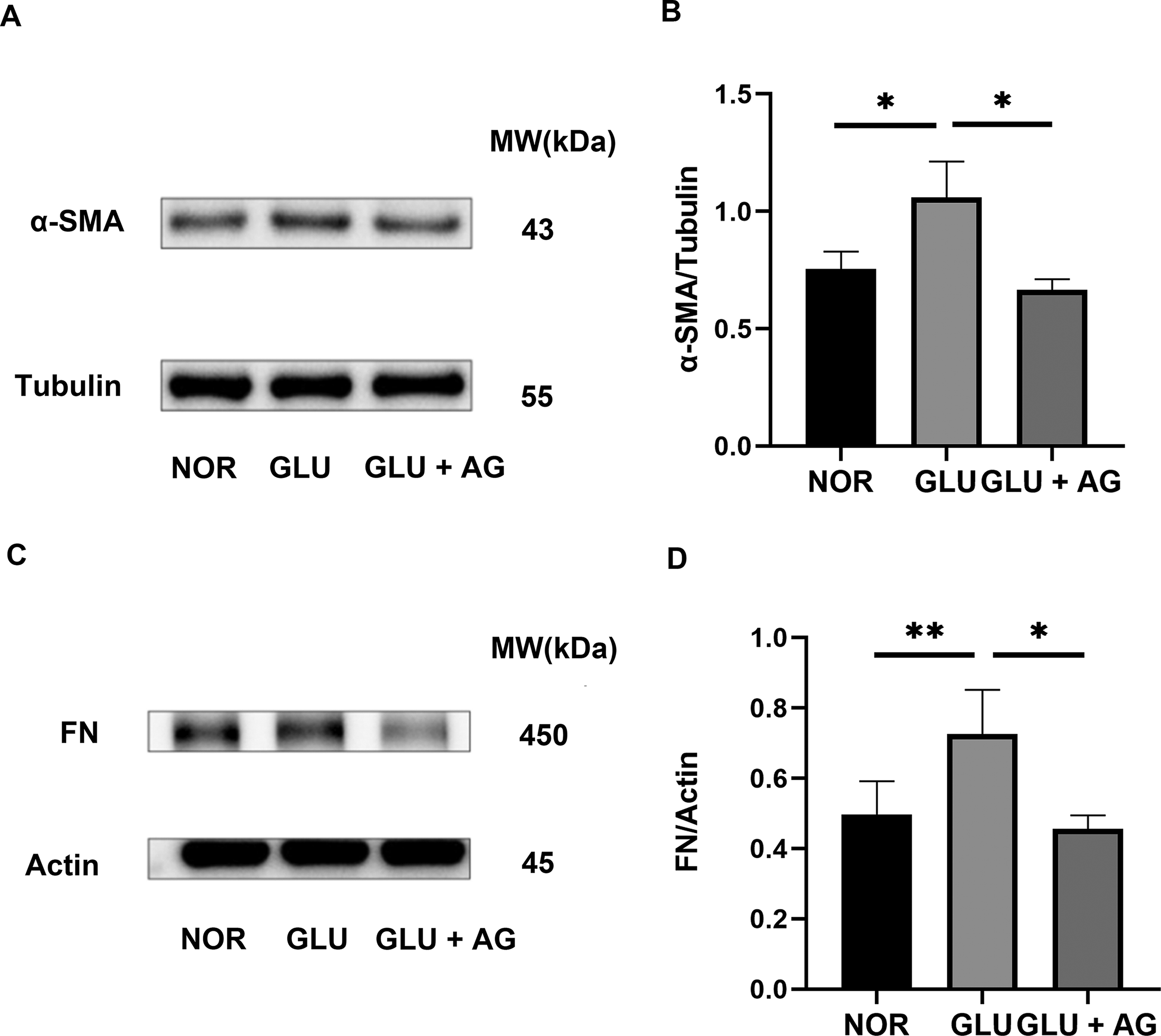
AG1295 Effectively Suppresses Hyperglycemia-Induced Phenotype Transition of Human Corneal Stroma Fibroblasts. Expression of α-SMA
AG1295 suppresses high-glucose-induced IL-6 expression in human corneal stromal fibroblasts
Exposure to a high glucose solution significantly increased the expression levels of IL-6, an important inflammatory cytokine that regulates cornea wound healing. However, treatment with AG1295 significantly reduced the high-glucose-induced IL-6 expression (Fig. 8).
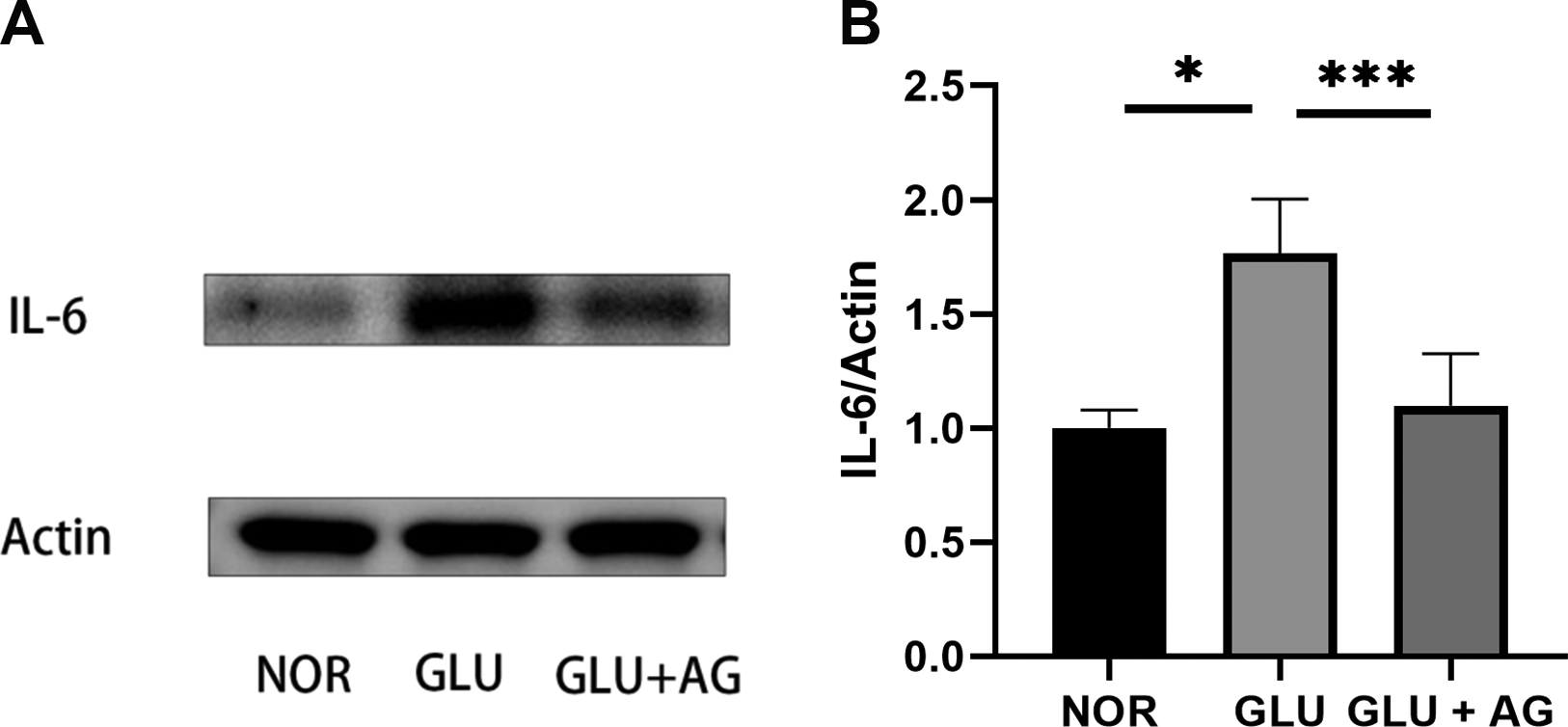
AG1295 Suppresses High-Glucose-Induced IL-6 Expression in Human Corneal Stromal Fibroblasts
DISCUSSION
The Refractive Surgery Preferred Practice Pattern, 21 issued by the Corneal Disease Group of the Ophthalmology Branch of the Chinese Medical Association, identifies diabetes as a relative contraindication for refractive surgery. However, the mechanisms by which hyperglycemia influences corneal fibroblasts remain unclear. Previous studies have shown that a serum-free, low-glucose environment helps maintain the corneal stromal cell phenotype, reduces cellular migration and contractility, and restores characteristic stromal cell markers, such as cluster of differentiation 34. 22 Despite these findings, research on the effects of hyperglycemia on corneal stromal fibroblast proliferation, migration, and inflammatory factor expression have not been fully elucidated. Therefore, our findings provide mechanistic insight into how elevated glucose directly alters corneal stromal fibroblast behavior.
Our findings also showed that high glucose conditions (30 mM) significantly increased PDGFRβ phosphorylation, downstream AKT phosphorylation, p-PDGFRβ/total PDGFRβ ratio, and p-AKT/total AKT ratio in human corneal stromal fibroblasts, accompanied by enhanced proliferative and migratory capacities of the fibroblasts, along with increased expression of the fibrotic markers ɑ-SMA and FN. Given that fibroblast aggregation, proliferation, and phenotype transformation are closely associated with tissue scarring, 9 these hyperglycemia-induced alterations may contribute to corneal scar formation and subsequent loss of transparency.
In this study, we isolated fibroblasts from fresh corneal lens tissue obtained from SMILE surgery. Traditionally, human corneal stromal fibroblasts have been isolated from residual tissue obtained during corneal transplantation surgeries or from donor corneas deemed unsuitable for transplantation. 23 However, the use of full-thickness corneal tissue that comprises the epithelial, stromal, and endothelial layers requires meticulous manual removal of epithelial and endothelial cells before fibroblast isolation, which increases the risk of cellular contamination. Furthermore, prolonged intervals between tissue procurement and cell isolation, as well as chemical preservation, may compromise the characteristics of the fibroblasts. Notably, the advent of SMILE surgery has revolutionized the isolation of corneal fibroblasts. 24 This minimally invasive refractive procedure yields abundant stromal lenticules with robust cellular activity in predominantly young, healthy patients with myopia. Unlike traditional sources, SMILE-derived lenticules offer several advantages, including immediate laboratory processing without freezing, which preserves tissue freshness and cellular viability. Moreover, their exclusive stromal origin eliminates the risk of epithelial or endothelial cell contamination. Immunofluorescence staining for Vimentin is used to confirm the successful isolation of pure human corneal stromal fibroblasts from these lenticules. Therefore, SMILE-derived stromal lenticules represent an optimal source of human corneal stromal fibroblasts, combining superior cellular viability with unparalleled purity, thereby advancing corneal biology and tissue engineering research.
Inflammatory factors play a pivotal role in fibroblast activation and fibrotic progression. 13 IL-6, a pleiotropic cytokine, is significantly upregulated during the inflammatory response. It mediates its effects through both membrane-bound and soluble receptor signaling pathways. In the present study, high-glucose exposure significantly, whereas PDGFRβ inhibition attenuated this response. These data suggest that PDGFRβ signaling contributes, at least in part, to hyperglycemia-induced inflammatory activation.
Patients with diabetes frequently exhibit reduced corneal transparency, which correlates with the degree of glycemic control. 8 Our findings provide mechanistic support for this observation by demonstrating that hyperglycemia enhances the proliferative and migratory capacities of corneal stromal fibroblasts while upregulating the expression of fibrotic markers and inflammatory factors. Overall, these findings suggest that patients with diabetes, particularly those with poor glycemic control, are at an increased risk of excessive scar formation following corneal injury, which can compromise corneal transparency. Following refractive surgery, corneal fibroblasts in patients with diabetes tend to migrate toward the incision site, proliferate, and differentiate into myofibroblasts at an elevated rate. This can lead to increased scar formation at the incision site, reduced postoperative corneal transparency, and diminished vision. These results provide a theoretical basis for refining perioperative risk assessment and management in this population.
PDGFR activation can be effectively inhibited by receptor tyrosine kinase inhibitor AG1295. In the present study, treatment with AG1295 significantly reduced the p-PDGFRβ/total PDGFRβ ratio and suppressed fibroblast proliferation and migration under high-glucose conditions. Moreover, AG1295 blocked the phosphorylation of both PDGFRβ and its key downstream effector AKT, confirming that PDGFR is essential for PI3K/AKT pathway action. AKT activation critically promotes cell growth, and its phosphorylation is a key step in mediating downstream effects of PDGFR signaling, including fibroblast proliferation and migration. 25 Previous studies have shown that AG1295 attenuates proliferative vitreoretinopathy in diabetic rabbits and exerts therapeutic effects on liver and kidney fibrosis.25–27 Similar mechanisms have been reported in tumor-associated fibroblasts, in which PDGFRβ inhibition reduces fibroblast activation, growth, and IL-6 secretion. 28 Collectively, these findings suggest the potential therapeutic value of AG1295 in reducing diabetic corneal scar formation and improving corneal transparency.
This study has some limitations. First, because corneal stromal cells tend to differentiate into fibroblasts under in vitro conditions, our experiments primarily reflect fibroblast behavior and may not fully recapitulate stromal cell responses in vivo. Second, fibroblasts were derived exclusively from normal donors. Third, even though AG1295 demonstrated inhibitory effects in vitro, additional studies are required to evaluate its ocular safety and therapeutic feasibility. Finally, although TGF-β and PDGFR-β signaling pathways are known regulators of corneal fibrosis, we did not examine TGF-β signaling—despite observing marked upregulation of α-SMA and FN, key markers of myofibroblast differentiation and extracellular matrix deposition. Future studies should address this pathway to further clarify molecular interactions.
Conclusions
PDGFRβ phosphorylation inhibitors can effectively suppress high-glucose-induced proliferation, migration, and expression of inflammatory and fibrotic factors in corneal stromal fibroblasts. Although this approach is a promising clinical strategy for alleviating corneal scar formation in patients with diabetes, its efficacy and safety in clinical applications should be further investigated and validated.
Authors’ Contributions
Conceptualization: Y.Z. and J.W. Software: Y.Z. and H.J. Formal analysis: Y.Z. and J.W. Investigation: Y.Z. and Q.Z. Writing—original draft preparation: Y.Z. Writing—review and editing: Y.Z., J.W., Q.Z., Q.W., and Y.W. Visualization: Q.Z. and H.J. Supervision: Y.W. Funding acquisition: Y.W. and Q.W. All authors have read and agreed to the published version of the article.
Footnotes
Acknowledgment
The authors are grateful for the technical support of the Tianjin Eye Hospital and Eye Institute, Tianjin Key Lab of Ophthalmology and Visual Science.
Data Availability Statement
The data in this article are available upon request from the corresponding author.
Author Disclosure Statement
The authors report there are no competing interests to declare.
Funding Information
This study was funded by the National Program on Key Research Project of China (2022YFC2404502), National Natural Science Foundation of China (82271118), Tianjin Key Medical Discipline Construction (TJYXZDXK-3-004A-3), and Science Foundation of Anhui Medical University (2023XKJ168).