
Abstract
A model for the mechanics of lipid membranes with non-uniform (coordinate-dependent) properties is discussed. The coordinate-dependent responses of the membranes are incorporated via the augmented non-uniform energy function and material parameters, which are dependent explicitly on the surface coordinates. We formulate the associated normal and tangential Euler equilibrium equations through which the coordinate-dependent responses of membranes are characterized. The admissible boundary conditions are taken from the existing non-linear model but reformulated and adopted to the present framework. Within the prescription of superposed incremental deformations, a compatible linear model is also formulated, from which a complete analytical solution describing the non-uniform responses of the membrane subjected to substrate–membrane interactions is obtained.
Keywords
1. Introduction
The morphological responses of lipid membranes have been the subject of intense research that has significantly enhanced our understanding of a wide range of essential cellular functions, such as fusion, budding, and vesicular transport [1–3]. Since lipid membranes are quite fragile and negligibly thin (typically 3–5 nm), analyses of their various mechanical properties are, most often, achieved via the use of an artificial “model”. This includes the development of continuum-based models describing the mechanics of the membranes, which is also in a period of intense study (see, for example, [4–7]). Lipid bilayer membranes are complex assemblies of lipid molecules (phospholipids), which are characterized by hydrophilic head groups and hydrophobic tails. These molecules arrange themselves into a two-layered sheet (a bilayer) with opposing orientations that form a critically important interface within biological cells, providing a selective permeability barrier for each cell [6–9]. This occurs, driven by the hydrophobic effect, over length scales of the order of molecular dimensions. Therefore, a lipid bilayer can be regarded as a closed membrane, much like a thin-film sandwich structure, where a fluid-like substance is present between the two films. Within this context, the development of theoretical prediction models for the mechanics of lipid membranes is facilitated by the differential geometry of a surface and the theory of an elastic surface, such that the deformation energy of a thin membrane can be expressed by the mean and Gaussian curvature of a surface Gaussian curvature of a surface [10, 11]. The energy potential that accommodates the bilayer symmetry with ensuing free energy minima was proposed by Helfrich [12], from which a well-known “membrane shape equation” is formulated. The Helfrich model has been successfully implemented in a wide range of membrane problems, including budding formations [13, 14], membrane–substrate interactions [5, 15, 16], and spontaneous curvatures [17, 18].
Recent efforts emphasize the refinement of the membrane’s energy potential and the associated mathematical framework to obtain more comprehensive and efficient prediction models for the descriptions of lipid membranes. Rangamani et al. [19] developed the general non-linear model of membranes, which incorporates the effects of intramembrane viscosity from the elastic model of surfaces [20, 21], and predicted the deformations of membranes subjected to a uniformly distributed pressure. A compatible linear model was formulated by Zeidi and Kim [22], within the prescription of superposed incremental deformations. Zeidi and Kim [22] obtained a complete analytical expression to describe the deformations of a lipid membrane, subjected to intramembrane viscous flow and substrate–membrane interactions. A constitutive framework that incorporates the tilt and distention of lipid membranes in involved responses of membranes was established by Steigmann [23]. The model is further adopted in studies of variable tilt [24] and thickness distension (without tilt) [25] of the membranes. To this end, Steigmann [6] developed a comprehensive model (including shape equations and admissible boundary conditions) for the mechanics of lipid membranes, which accommodates tilt, distension, diffusion, and viscous flow, from the theory of three-dimensional liquid crystals [26–28].
Most of the aforementioned studies presume uniform properties throughout the membrane (i.e. coordinate-independence) in an effort to obtain mathematically tractable systems and analyses. In general, though, the responses of membranes are coordinate-dependent, owing to the complex nature of such membrane systems and processes as diffusion and non-uniform protein distributions [29]. For example, drug-induced protein diffusion may be considered an energy-dissipative process [30] and therefore may further induce non-uniformity in the membrane. In addition, phase separations or local enrichments of particular lipid species arising in membrane–protein substrate interactions [31] may be considered a source of non-uniformity. Protein-induced pre-deformations may also give rise to the possible non-uniformity. For example, BIN/amphiphysin/Rvs (BAR) proteins behave as a scaffold-like structure, acting as a local tension field, which may be viewed as a non-uniformity [32]. A class of problems pertaining to the non-uniform properties of lipid membranes is discussed in [4] and [20], where the non-uniformity is accommodated via a coordinate-dependent energy function of membranes. Further, Agrawal and Steigmann [17, 29] investigated the non-uniform properties of membranes induced by surface diffusion and possible non-uniform protein distributions, and predicted the non-uniform spontaneous curvature of the membranes. However, deformation analyses of membranes accounting for explicit coordinate-dependent non-uniformities, particularly those arising in the membrane shape equation, remain lacking in the literature. In this work, we study a model that describes the non-uniform properties of lipid membranes subjected to membrane–substrate interactions and lateral pressure. The non-uniformity of the membrane is incorporated via the introduction of non-uniform energy distribution functions and material parameters, which are explicitly dependent on the surface coordinates. The corresponding normal and tangential Euler equilibrium equations are then derived, through which the coordinate-dependent responses of membranes are characterized. The admissible boundary conditions are adopted from the work of Agrawal and Steigmann [5], yet they are reformulated in the present context with the refined energy potential of the membrane. Within the prescription of superposed incremental deformations [33, 34], a compatible linear model is developed from which a complete analytical solution describing the non-uniform responses of the membrane undergoing substrate–membrane interactions is obtained.
More importantly, we found that, unlike those from the non-linear theory, the coordinate-dependent material parameters of membranes do not necessarily result in non-uniform responses of membranes. In fact, it is shown through the tangential and normal Euler equations that the terms associated with the non-uniform material parameters vanish for small deformations superposed on large deformations. This result further suggests that the descriptions of the non-uniform responses of membranes are intrinsically limited to those achieved via the augmented energy potential, which is explicitly dependent on the surface coordinates. Nevertheless, the solutions obtained from the proposed linear model demonstrate reasonable agreement with those obtained from the non-linear analysis for the small deformation regime. Both circumferential and radial non-uniform energy distribution functions are considered in order to assimilate possible non-uniformities, and the resulting deformation fields demonstrate clear signs of coordinate dependency. In particular, we have shown that the principle of superposition from linear elasticity remains valid for the present application. The result may promote the relevant morphological study of membranes, in that the solution of combined non-uniform energy distribution cases can be directly obtained via the summation of the solutions obtained, respectively, from the circumferential and radial non-uniform cases. Numerical non-linear solutions are also obtained for the purposes of comparison with those from the linear analysis and demonstration of the more general coordinate-dependent responses of membranes. The non-linear solution predicts the off-centered biconcave morphology of lipid [35, 36] and the multiple peak formations of abnormal cell membranes (burr cell), which are commonly observed in uremia and chronic kidney disease [37, 38]. Lastly, the solution presented accommodates the results in [5] and [22].
Throughout the paper, we make use of a number of well-established symbols and conventions. Thus, unless otherwise stated, Greek indices take the values in {
2. Preliminary: Surface geometry
In this section, we summarize the results from the differential geometry for use in the derivation of the non-uniform membrane formulation. Let

The local surface orientation

where the dot denotes the conventional Euclidean inner product. The matrix of the surface metric is positive-definite (i.e

where

and

Further, equations (1) to (5) furnish the useful relations

Here,

which may also be used in later sections.
Finally, the mean and Gaussian curvatures are given by

which satisfy the following equalities
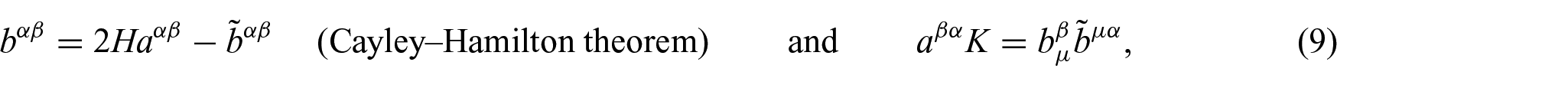
where

3. Non-uniform membrane shape equation
The membrane shape equations are given by [5]
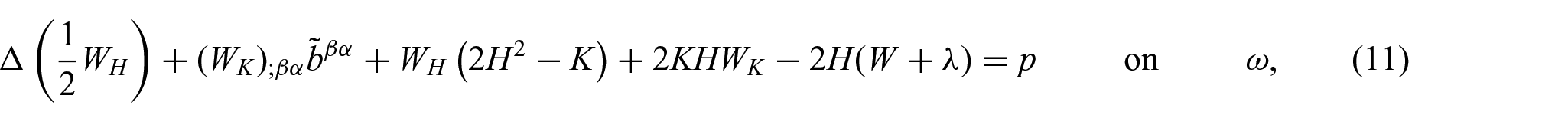
and
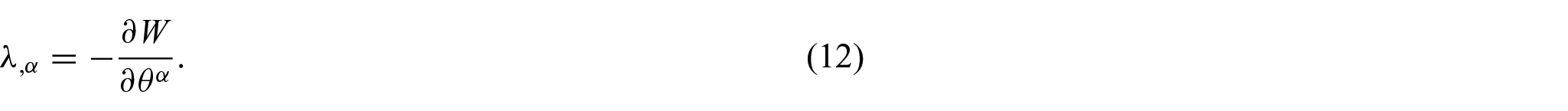
If the membrane energy potential W does not depend explicitly on the coordinates (i.e. uniform membranes), this yields [17]

and the values of

Equation (14) arises as a result of the explicit coordinate dependence in the function W, which arises from the possible non-uniformity of the membrane properties. Hence, to accommodate a particular state of non-uniformity,
In this study, we propose the following form of the modified Helfrich energy potential to assimilate non-uniform responses of membranes:

It is also noted that, the uniform membrane case (i.e.
In view of equation (15), we evaluate

and thereby obtain from equation (11)
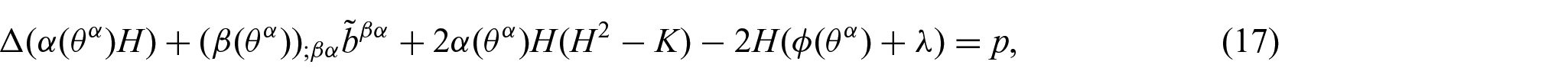
which serves as the shape equation for non-uniform membranes. Here, since

and
Further, from equation (15), we compute

where

Thus, the associated Euler equilibrium equations are found to be
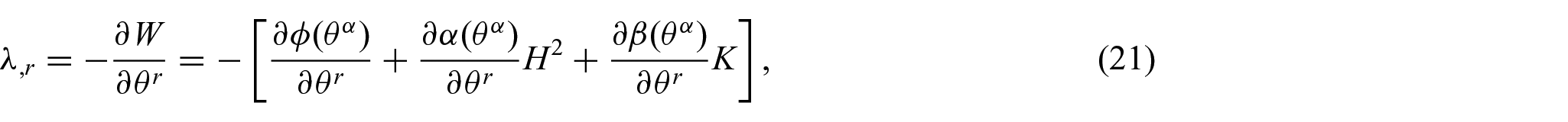
and
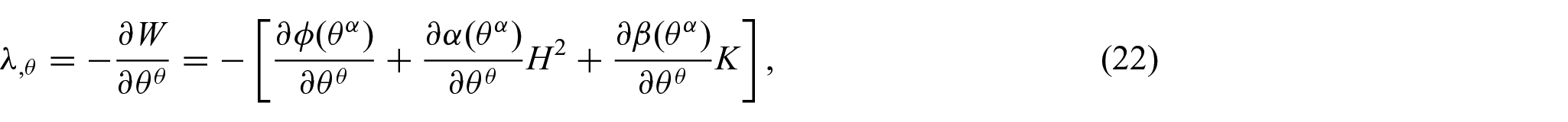
where
It is noted here that the proposed energy density function, equation (15), can be directly used in conjunction with the existing results (equations (11) and (12)) to yield the associated Euler equations (i.e. equations (17), (21), and (22)) without further modification (see Remark 1). This case, in which the refinement of the energy density function W results in non-standard forms of the shape equation, is discussed in [4, 17, 29].
To see this, we consider the virtual displacement of the equilibrium position field

The decompositions of

where
Now, the variations of the energy function W are evaluated using the chain rule:
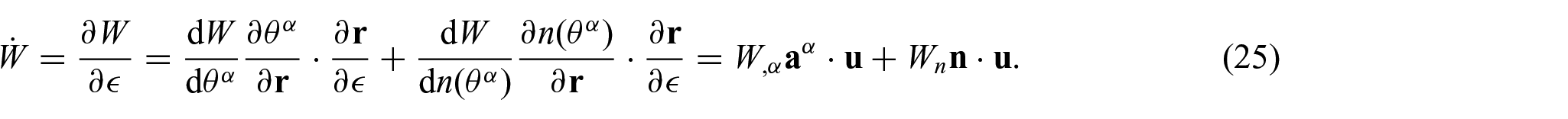
Substituting equation (24) into equation (25) and using

Thus,
Further, the associated Euler equations are given by (see [5] and [25])
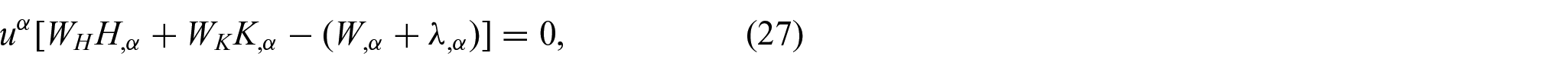
and
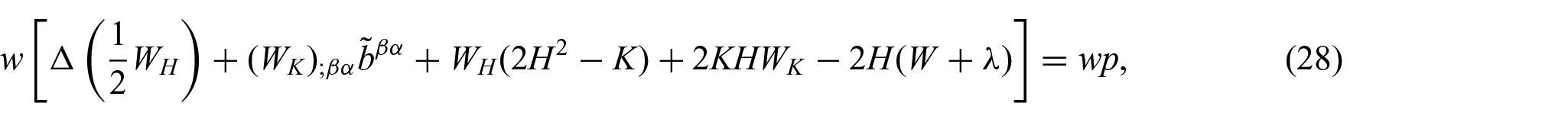
where
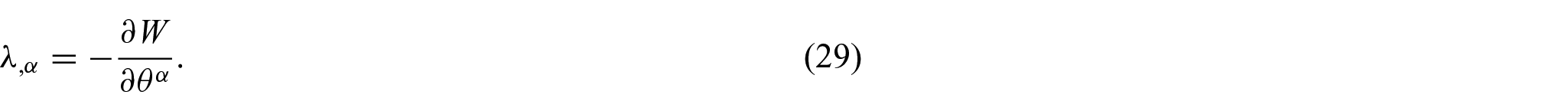
Therefore, it is evident from equations (28) and (29) that the resulting Euler equations remain intact even with the presence of the non-uniform distributions of
3.1. Formulations under Monge parametric representation
Using the Monge representation, the parametric position vector can be expressed as

where
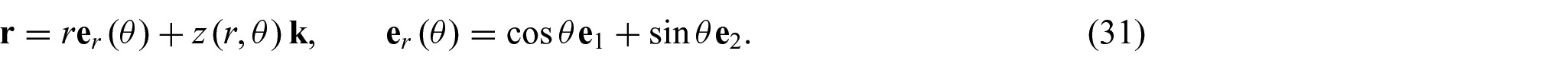
Substitution of equation (31) into equations (1) and (2) yields
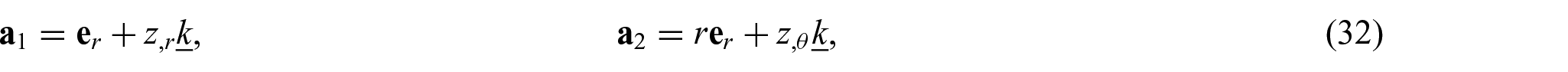
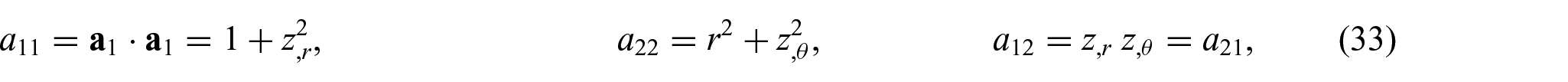
and

which are, respectively, the surface coordinates, the induced metric, and its determinant. Similarly, the dual basis vectors and the matrix of dual metric components (
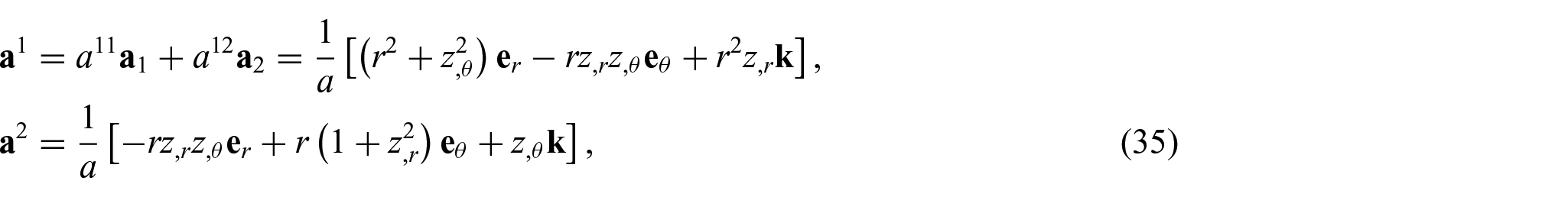
and
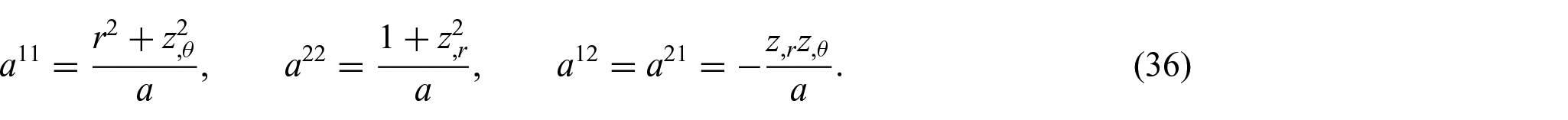
Further, from equations (32) to (36), the associated surface normal and the curvature tensor are now defined by
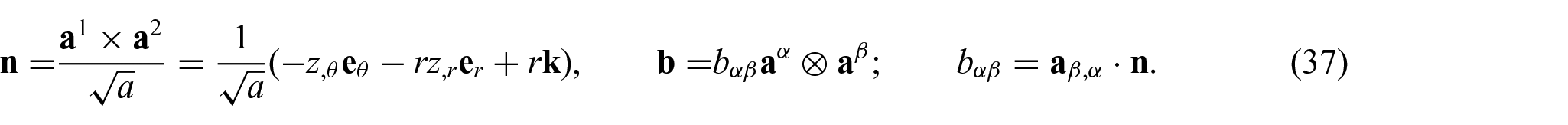
Thus, the complete expression of
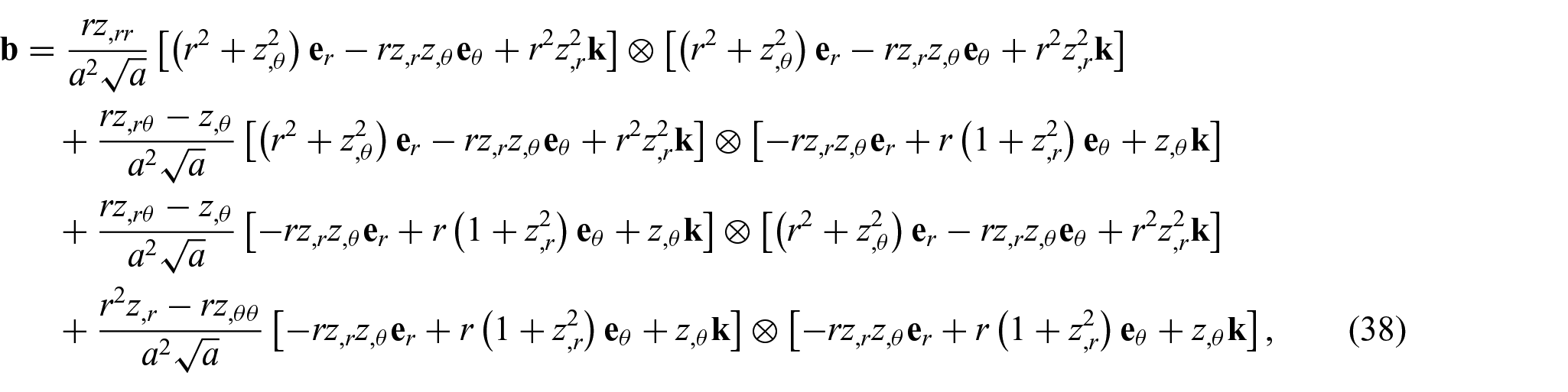
where the coefficients of the second fundamental form
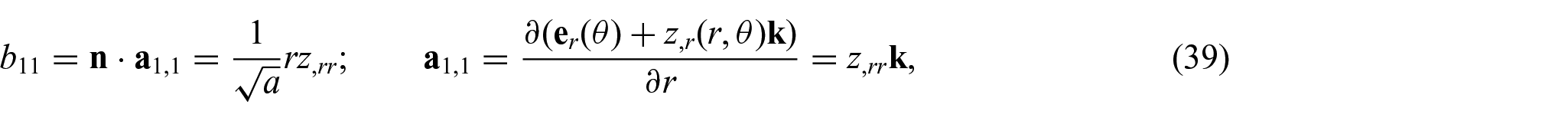
and similarly for
Lastly, from equations (8) and (36) to (38), we find the following expressions for the the mean and Gaussian curvatures:
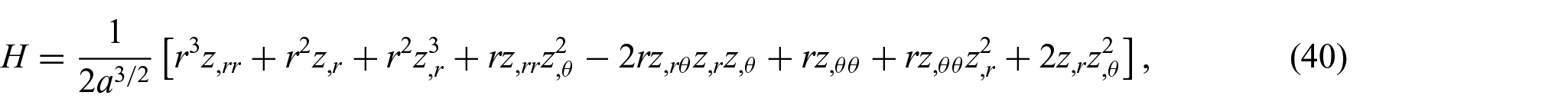

In the actual implementation of the Monge parameterization, equation (17) can be readily reformulated by using the results in equations (32) to (41). The corresponding formulations are relatively straightforward and hence omitted for the sake of simplicity.
Our intention is to assimilate the substrate–membrane interactions in the presence of non-uniform energy distributions. For this purpose, the following interaction boundary conditions are adopted from the work of Agrawal and Steigmann [5]:
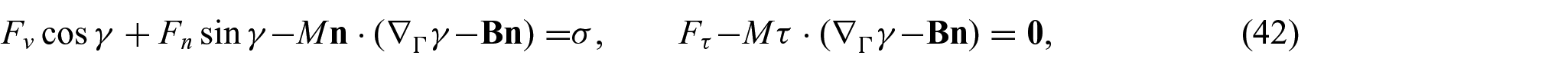
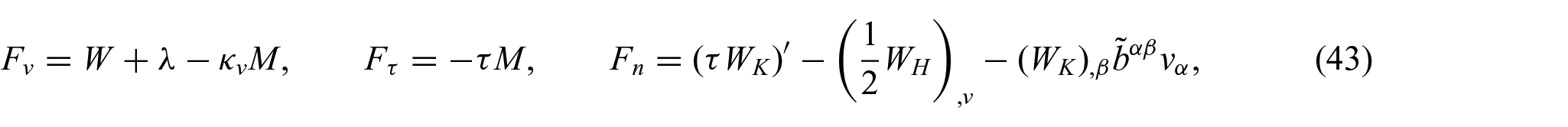
where
In the present case, in which the non-uniform membrane interacts with a circular cylindrical substrate of radius

Accordingly, equation (42) reduces to
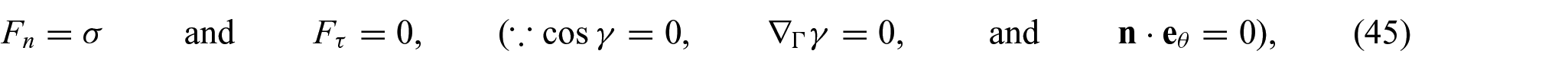
at the interacting boundary
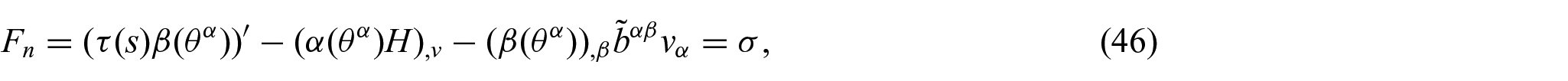
where
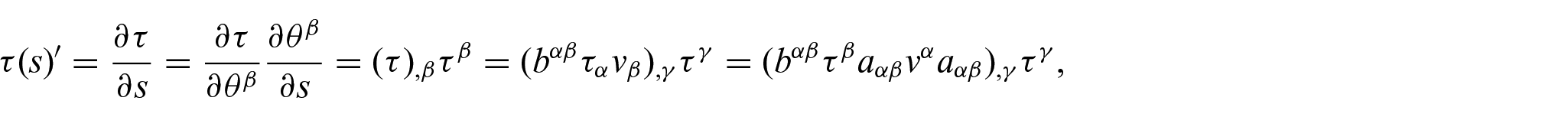
and

These expressions are then rewritten using the Monge representation for further analysis. For example, we have
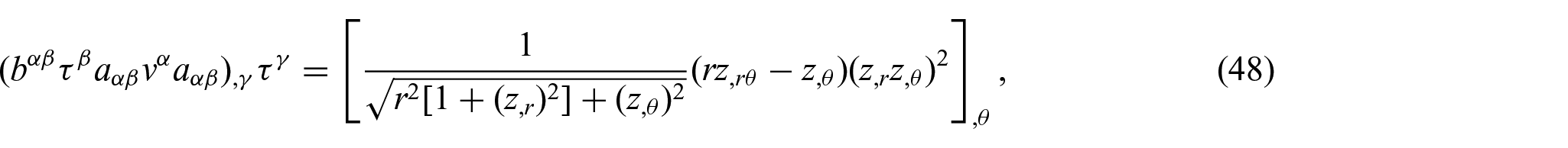
and similarly for other terms.
Detailed derivations and phenomenological implications of these boundary forces are available in [5] and [6], respectively. The interaction boundary conditions (i.e. equations (45) and (46)), together with equations (17) to (22) solve the deformed configuration of the non-uniform membrane subjected to membrane–substrate interactions. In the assimilation, we employed commercial packages (e.g. Matlab and Comsol) to solve the obtained partial differential equations (PDEs); the corresponding results are presented in later sections.
3.2. Linear model for non-uniform membranes
The formulation of the non-uniform equilibrium equations (i.e. equations (17) to (22)) in terms of equations (35) to (41) yields a highly non-linear PDE system, which often requires considerable computational resources. Alternatively, “admissible linearization” may be considered, through which one could obtain mathematically tractable systems and, more importantly, analytical expressions of solutions with minimal loss of generality. The concept has been widely and successfully implemented in studies of relevant subjects (see, for example, [5, 7, 15]). Within this setting, the derivatives of
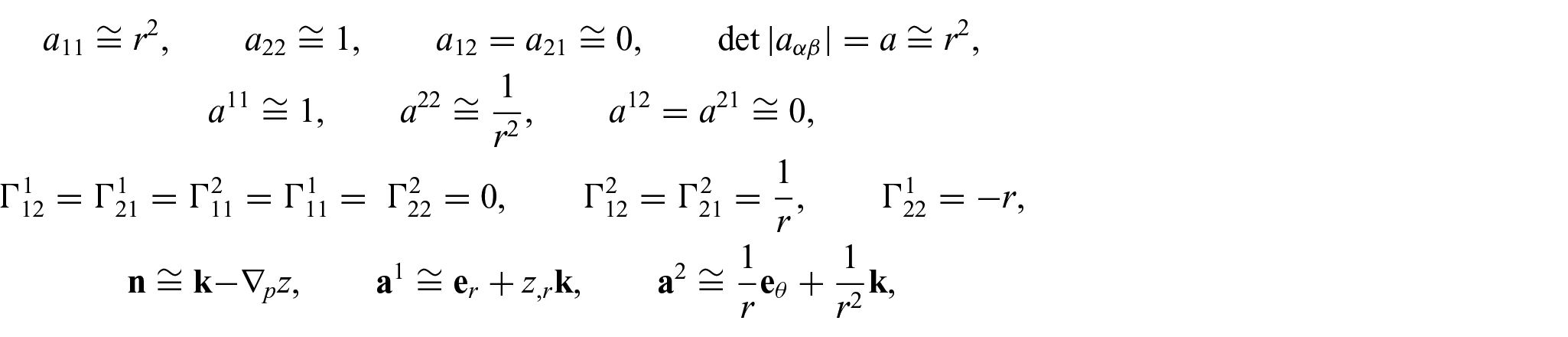
and
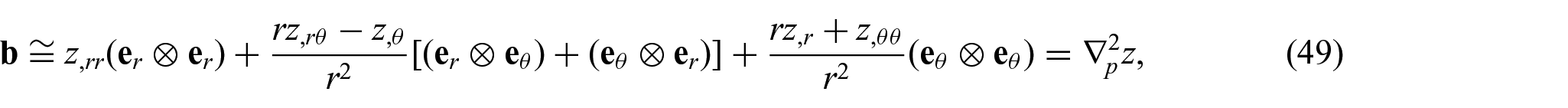
where the subscript
Thus, applying the results in equation (49), the mean and Gaussian curvatures (equations (40) and (41)) can be approximated as
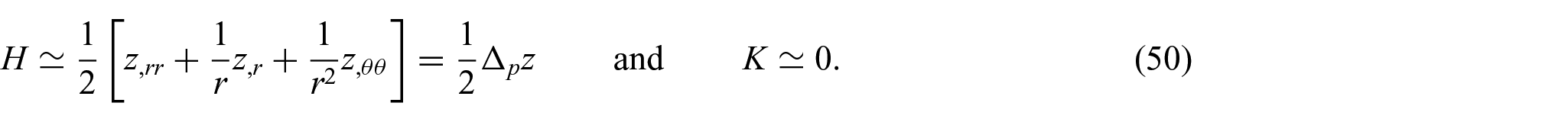
In addition, from equation (50), we reduce equations (21) and (22) to

and

which serve as the linearized Euler equilibrium equations in the coordinate directions.
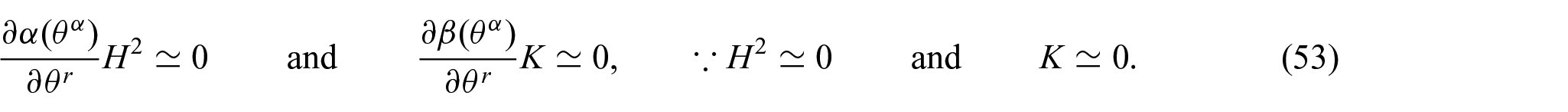
In other words, the non-uniform potentials of
Based on Remark 2, the following compact form of the membrane energy potential may be proposed to accommodate the non-uniform responses of membranes:

where
Therefore, equation (17) can be approximated in accordance with equation (54) as

which may serve as the compatible form of the shape equation for non-uniform membranes within the linear description.
3.3. Example1: Circumferentially non-uniform membranes
We consider the following form of the non-uniformity:

where
For

so that the membrane is radially uniform (

where we set

where the expression of

where
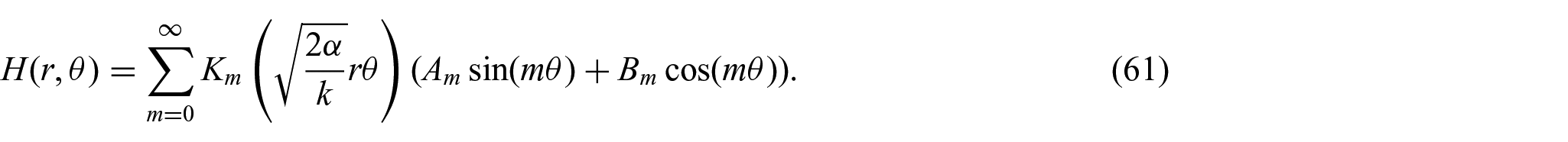
We continue by combining equations (50) and (61), and thereby obtain
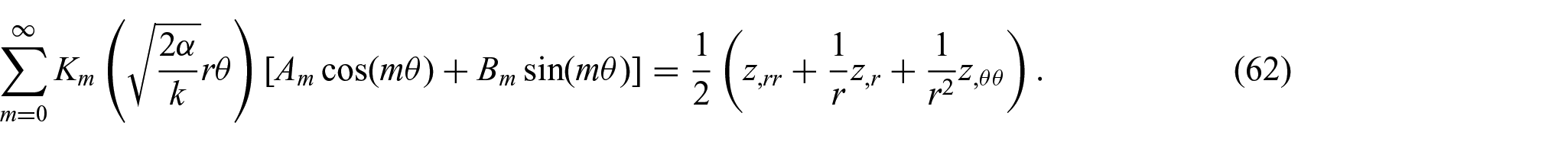
The solution of this PDE can be sought in a similar form to that in equation (59):

Further, the substitution of equation (63) into equation (62) yields
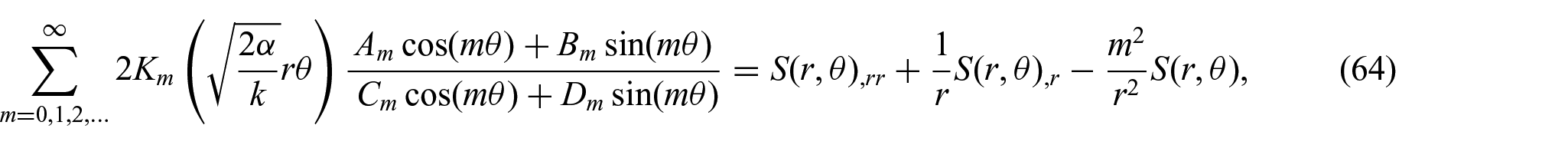
where m is the separation variable. Thus, we obtain the following expression for
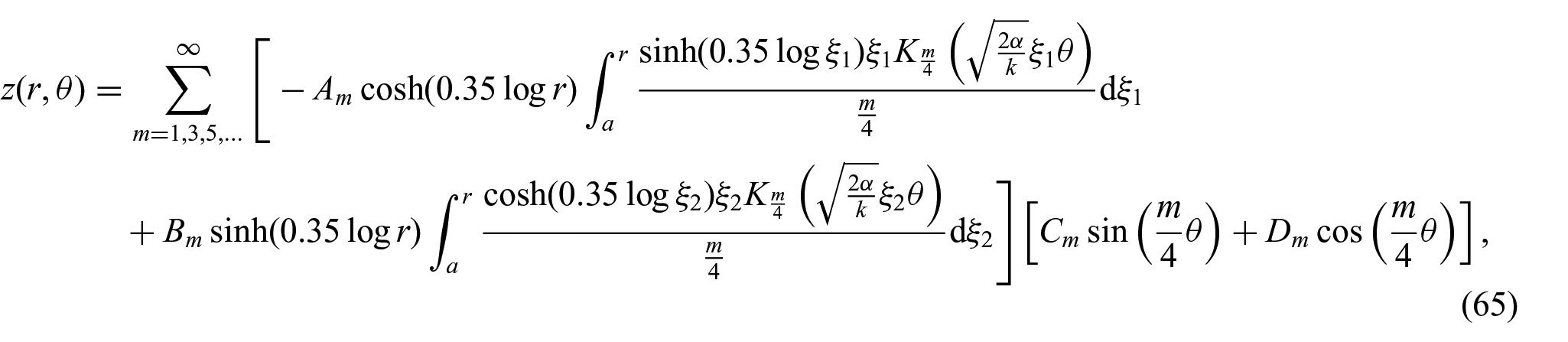
where a is the radius of the interaction boundary. The unknown constants (i.e.

For example, we expand the applied interaction force in terms of Fourier series as
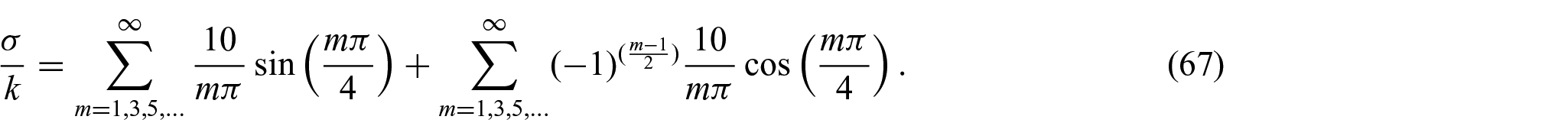
Therefore, the unknowns
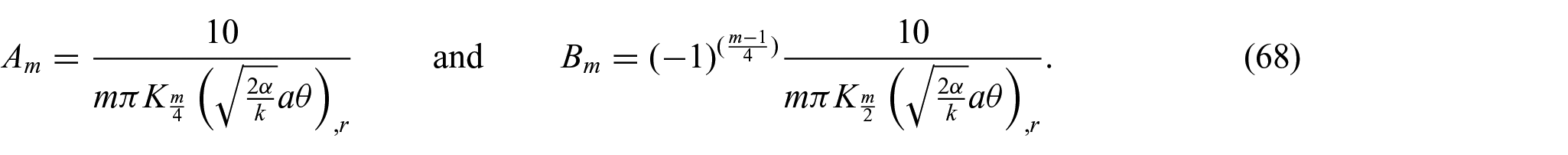
In the assimilation, we adopt the flexural modulus of the membrane as

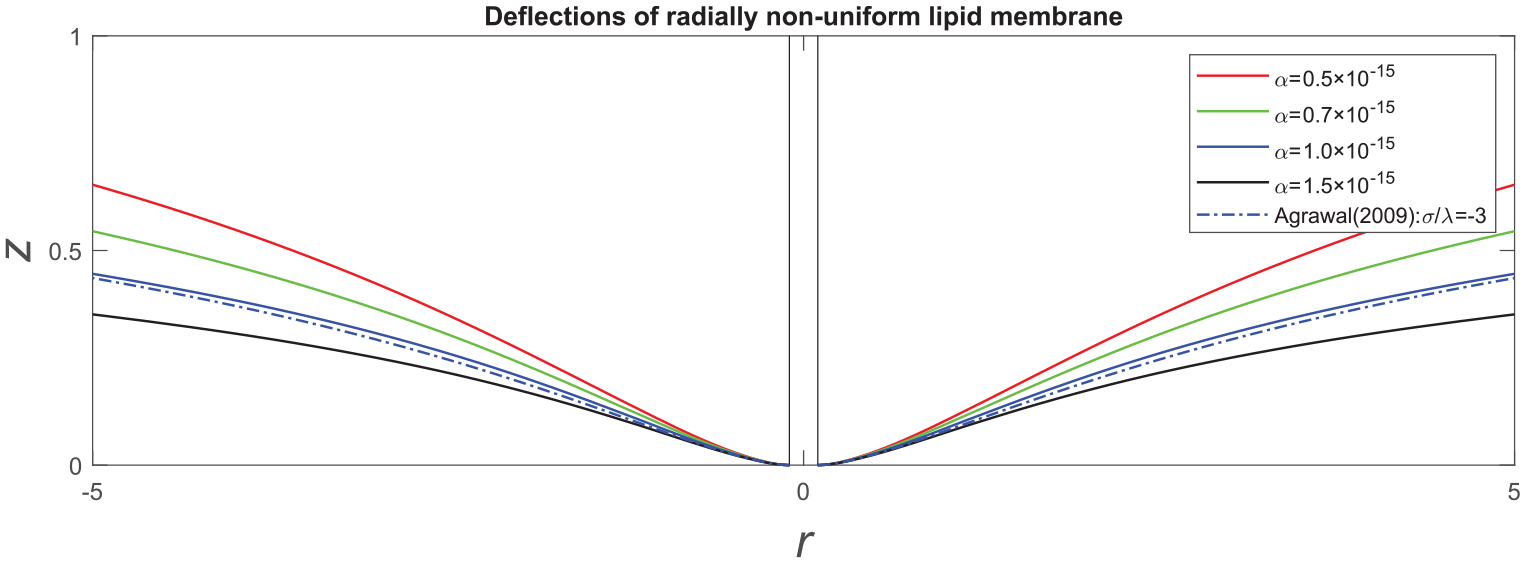
Figure 1 illustrates the deflections of a non-uniform membrane predicted by the proposed model at a particular configuration of
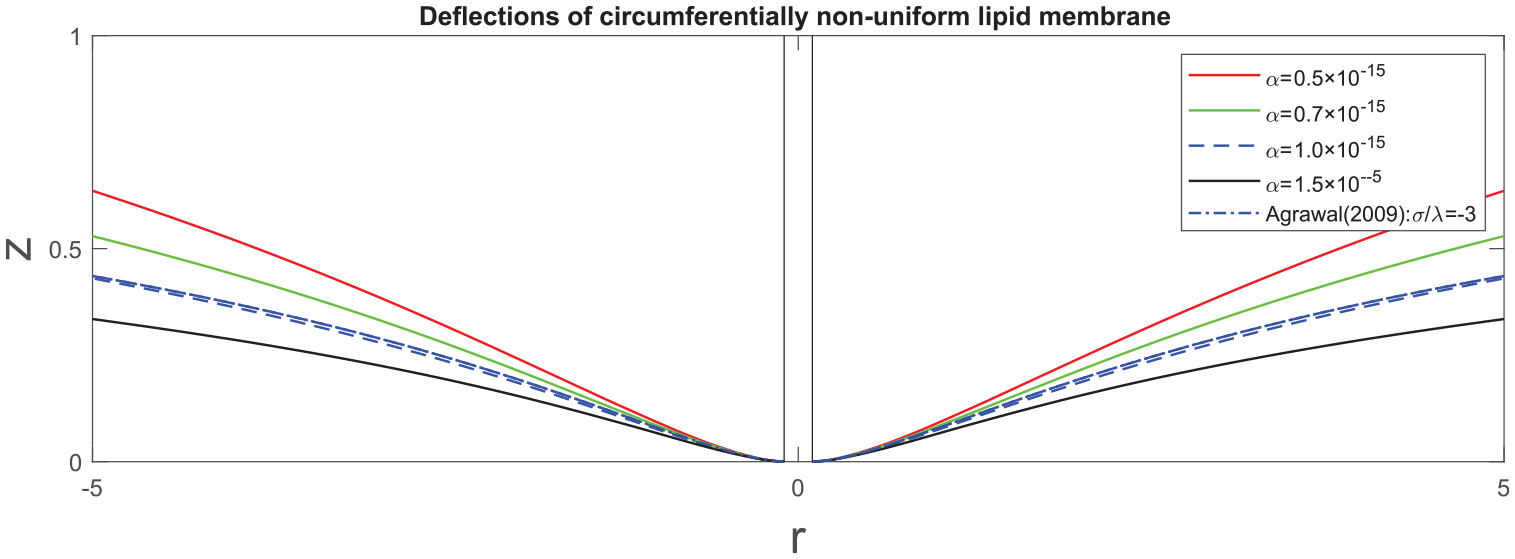
Transverse deflections of lipid membrane with respect to
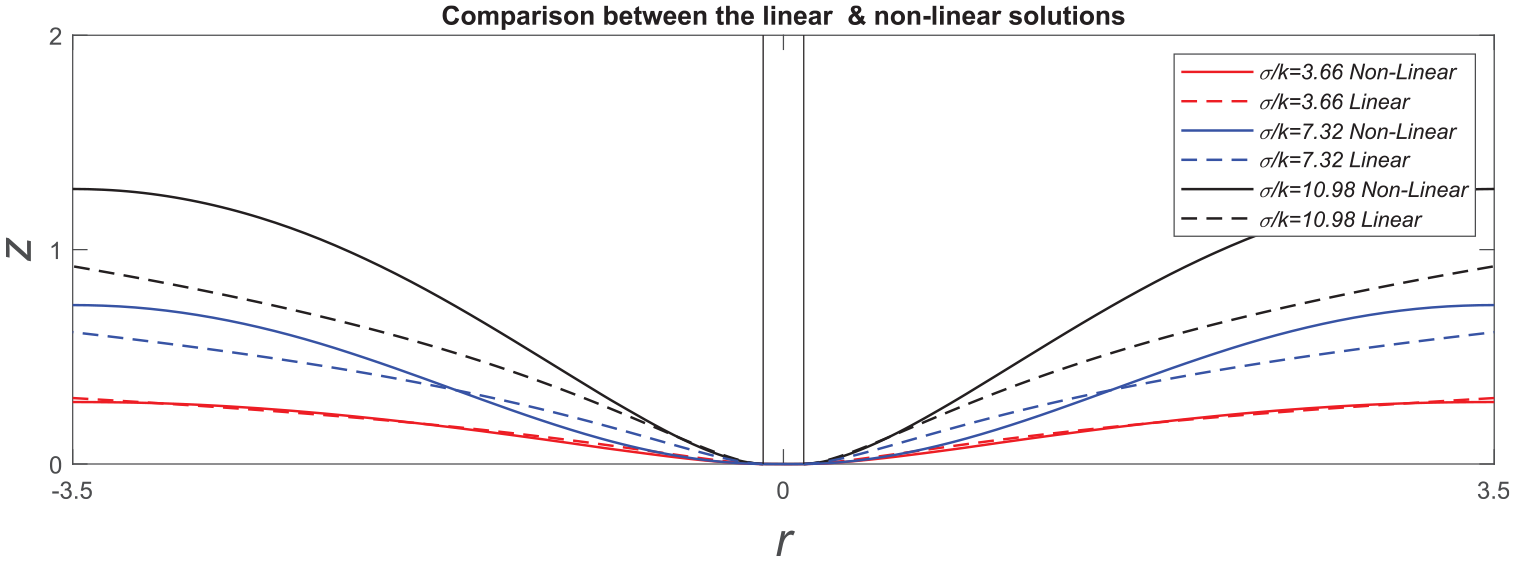
Transverse deflections of circumferentially non-uniform membranes: comparison of linear and non-linear solutions.
Lastly, the non-uniform responses of drug-treated diseased cell membranes [42] may be assimilated by using the proposed model. For this purpose, we adopt the following form of the periodic energy density distribution:

where n characterizes circumferentially non-uniform energy distributions,
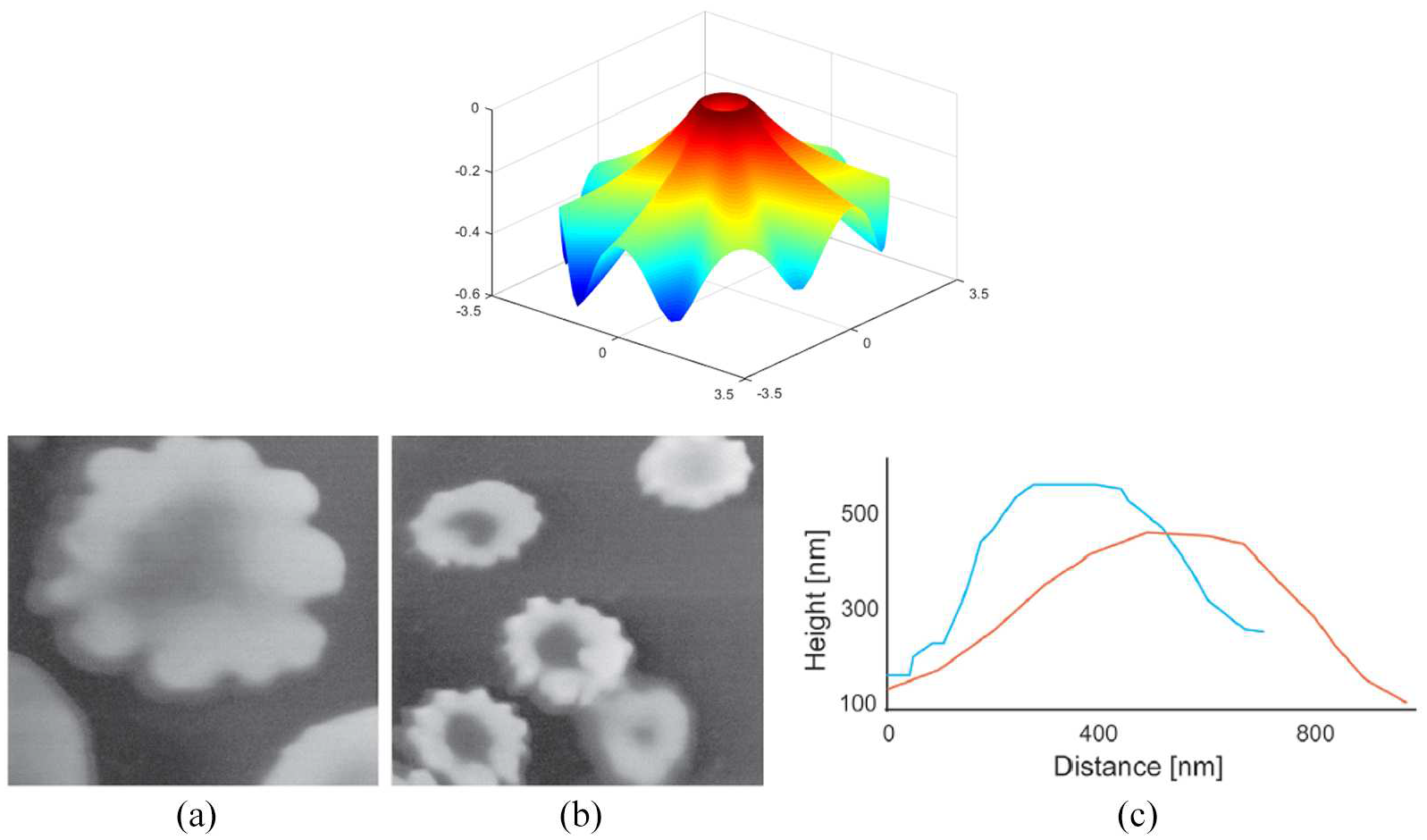
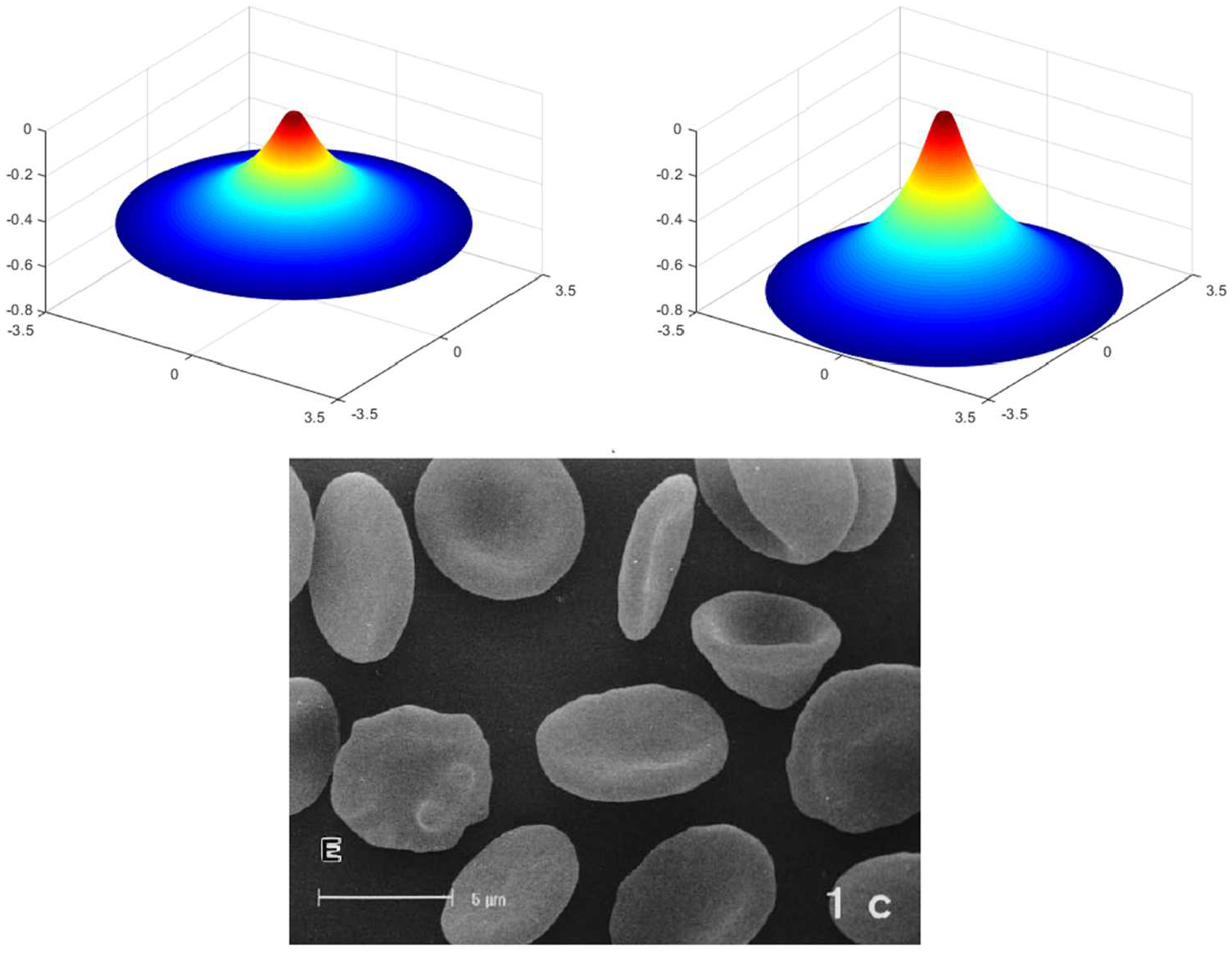
(top) Assimilation of echinocyte formation when
(left) Assimilation of the off-centered non-uniform morphology when
3.4. Example 2: Radially non-uniform membranes
The cases of radially non-uniform energy distributions are also examined, where the radial distribution function is characterized by

In the forgoing analysis, we demonstrated the case for
For

from which the radially non-uniform membrane (

Hence, combining equations (50) and (73) furnishes
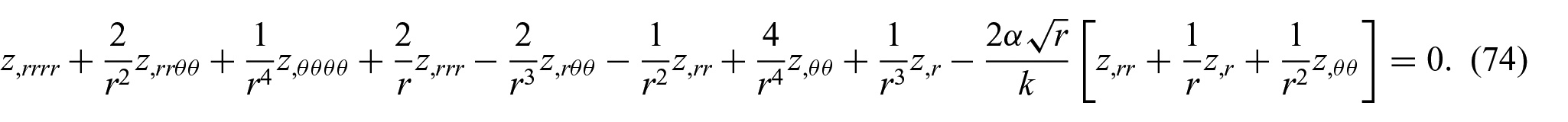
In the case of axisymmetric energy density distributions (i.e. circumferentially uniform and radially non-uniform), equation (74) may further reduce to
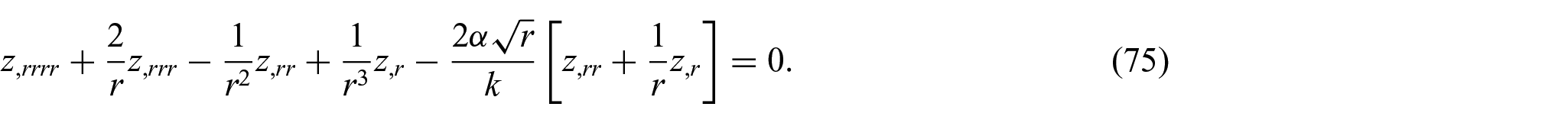
The solution of equation (75) can be found as
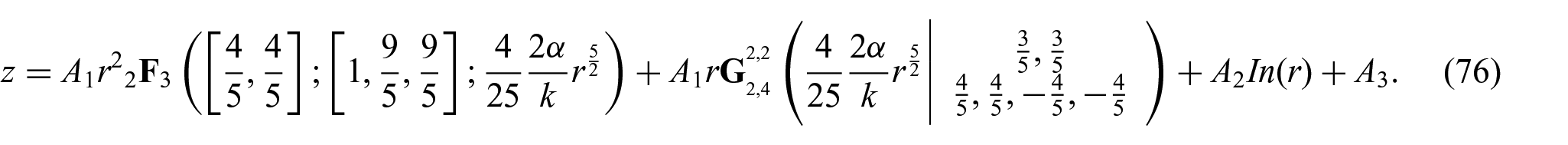
Here

and

We seek a bounded solution within a reasonably finite domain of
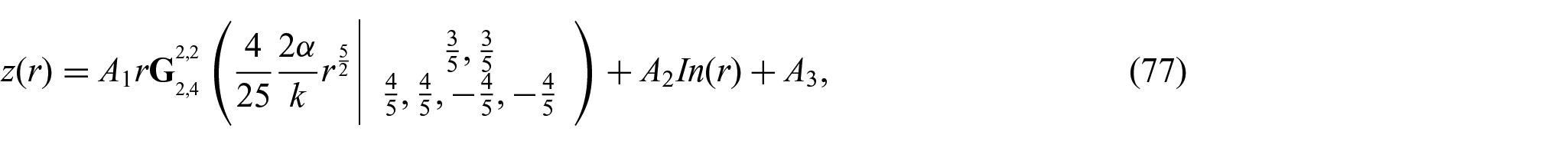
where the unknown constants
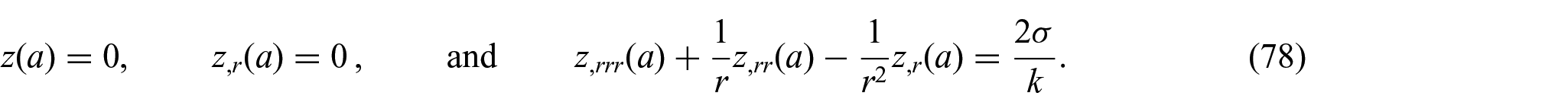
Thus, we find
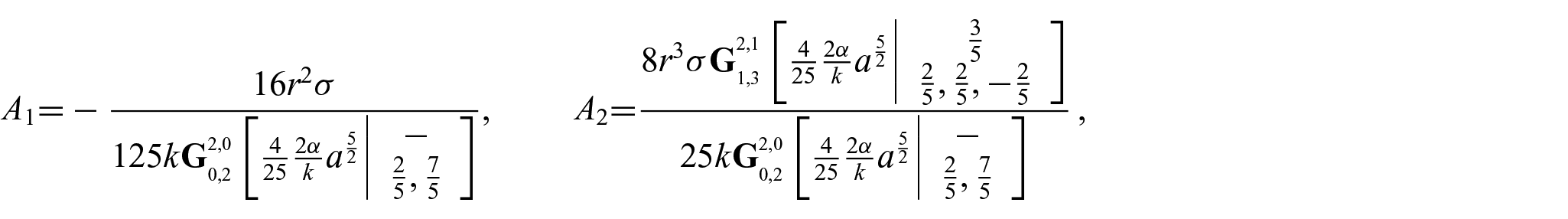
and
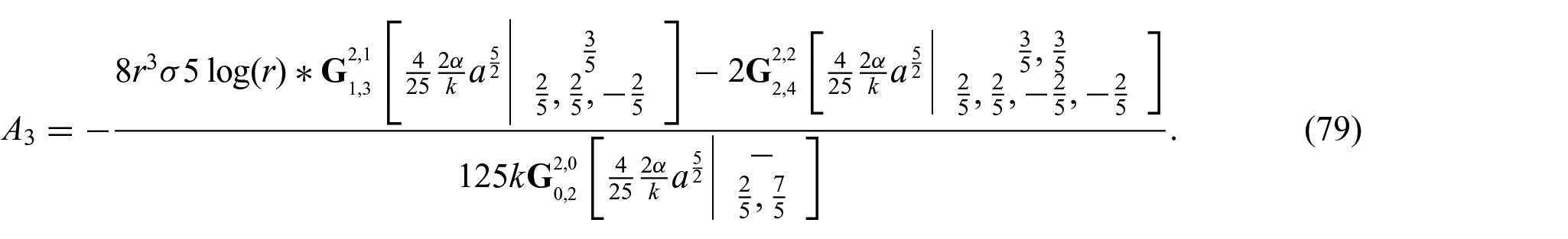
The assimilation of the obtained solution is performed within the same normalized setting, as depicted in equation (69). Like the circumferentially non-uniform cases, the transverse deflection of membranes increases as the intensity of the membrane’s energy distribution (
Transverse deflections of lipid membrane with respect to
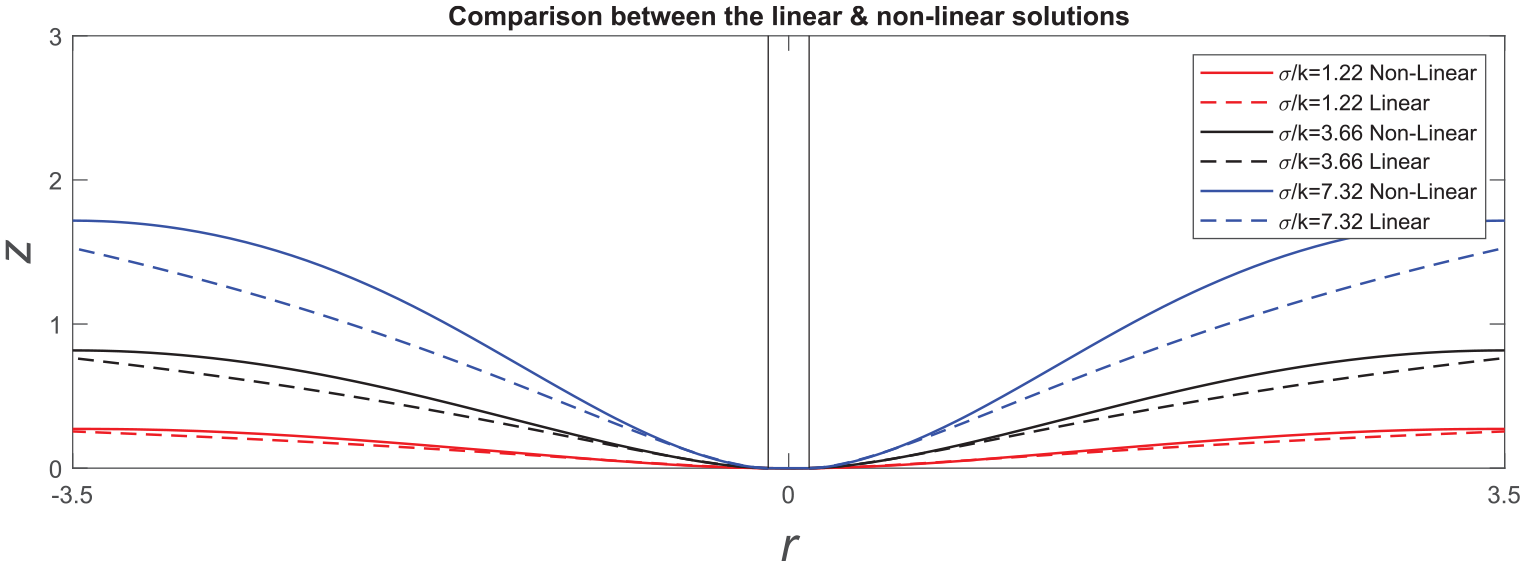
Transverse deflections of radially non-uniform membrane: comparison of linear and non-linear solutions.
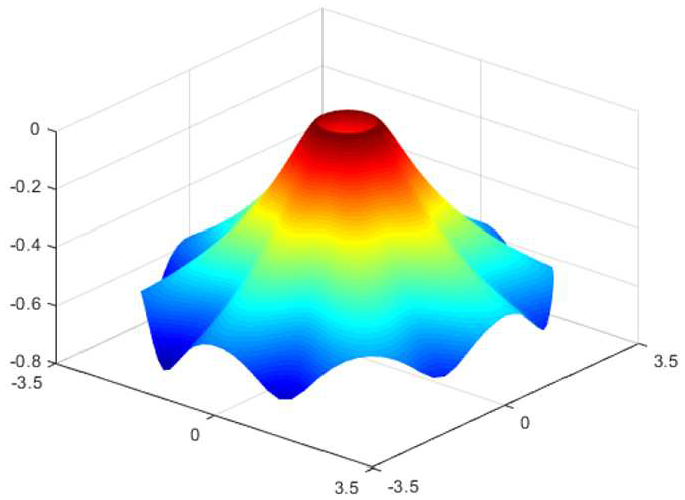
The sequences of discocyte–stomatocyte morphology in cell membranes [47] may be mapped using the proposed energy density function (equation (71)) via
(top) Sequence of deformation mapping with respect to
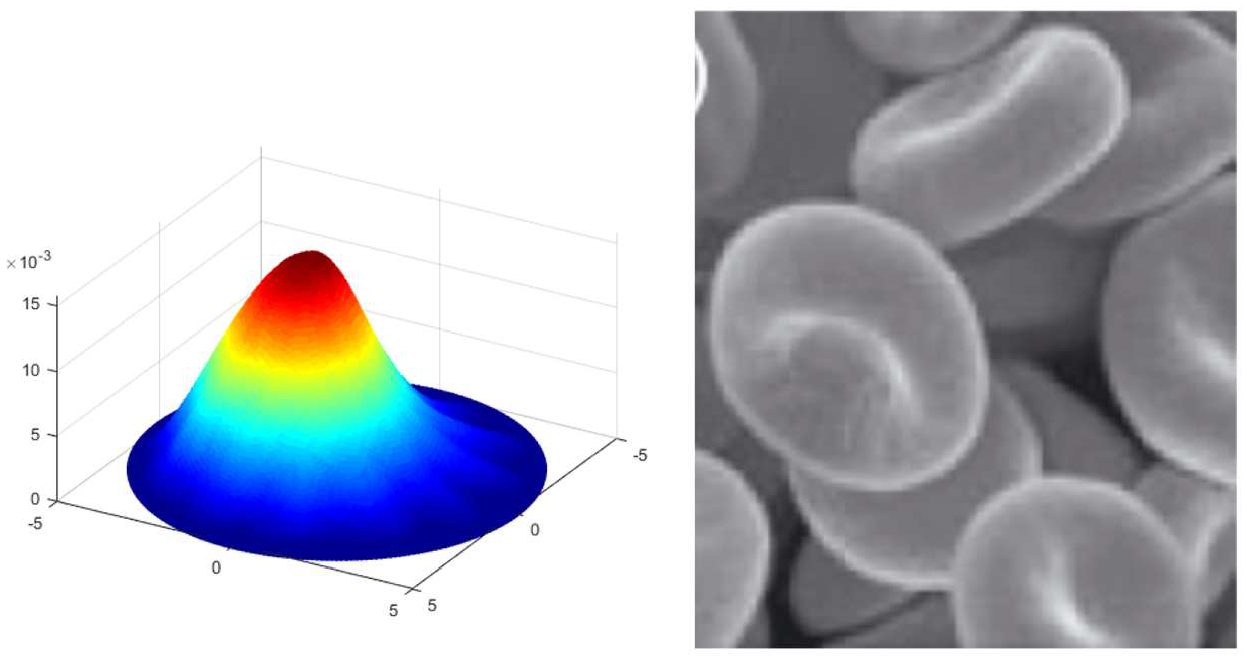
Deformation of radially and circumferentially non-uniform membrane when
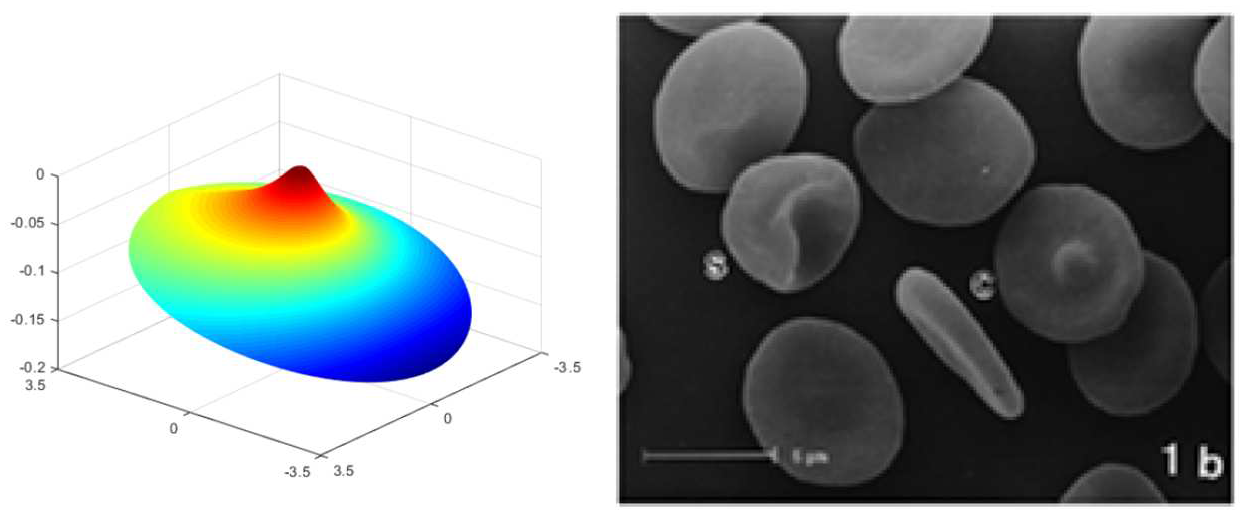
Lastly, we remark that more general membrane configurations may be characterized by combining the proposed non-uniform energy density functions. For example, the deformation contour predicted by the obtained non-linear model demonstrates close similarity to the highly off-centered protrusion of red blood cell membranes that are treated under 14 days of storage in a liquid medium [36] (see Figure 9). Also, Figure 10 illustrates that the non-linear solution predicts the arrangement of peak formations of abnormal cell membranes (burr cells) commonly observed in uremia and chronic kidney disease [37, 38]. In the assimilations, the magnitude of normal deformation is controlled by the intensity parameters (
(left) Deformation contour characterized by the potential
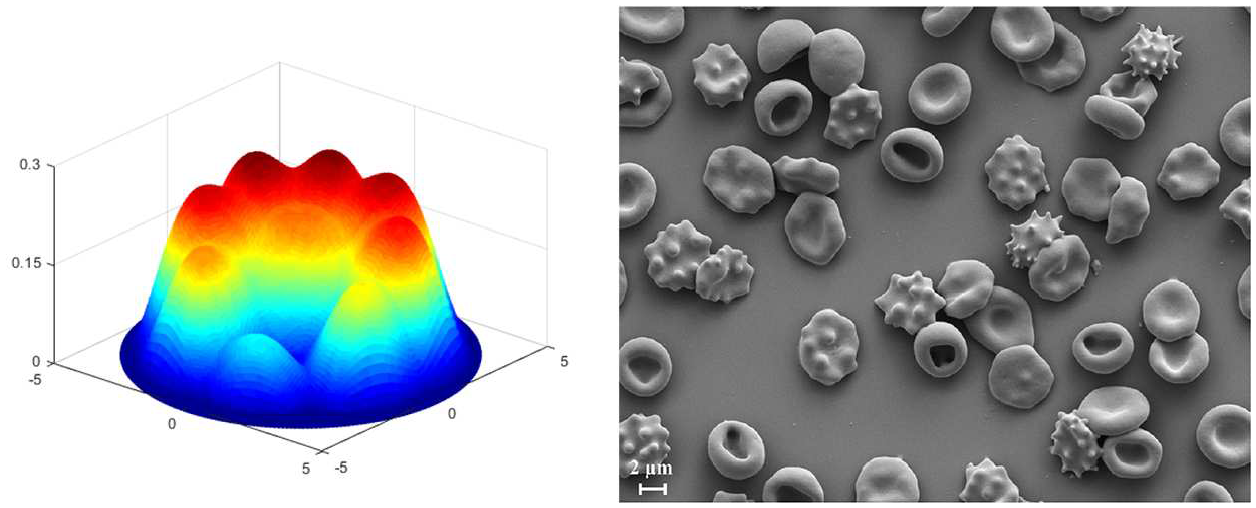
(left) Deformation contour characterized by the potential
As mentioned in earlier sections, the proposed model may not be sufficient for immediate uses in the phenomenological or clinical research of cell membrane morphology. However, it could still provide quantitative information pertaining to such membrane formation via the estimations of required elastic energy and the corresponding non-uniform energy distributions over the domain of interest.
Footnotes
Acknowledgements
Chun IL Kim thanks Professor David Steigmann for his invaluable and inspirational advice in the subject.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Natural Sciences and Engineering Research Council of Canada (Grant Number RGPIN 04742).