
Abstract
Background
Macrophages seem to play an important role in the development of glomerulosclerosis. In both human and experimental animal models of focal glomerulosclerosis (FSGS), infiltration of macrophages in the mesangium has been considered key in the development of FSGS.
Methods
In the present study, we evaluated the effect of vasoactive agents on the migration of monocytes across a filter in a modified Boyden chamber as well as across a cultured glomerular endothelial cell layer (in vitro model of glomerular mesangium). Both light as well as scanning electron microscopic studies were performed. We evaluated the effect of vasoactive agents including histamine, prostaglandin (PG) E2, angiotensin II, endothelin-1, platelet-activating factor, and interleukin-1 (IL) on the migration of monocytes/macrophages across an endothelial cell layer as well as a gelatin-coated filter. In addition, we evaluated the effect of cyclic adenosine 3',5' cyclic monophosphate (cAMP) and PGE2 on vasoactive-induced migration of monocytes.
Results
Histamine increased (P<0.003) the migration of monocytes across the filter. This effect of histamine was dose- dependent. Histamine at concentrations of 10−8 to 10−5 mol/L induced optimal migration across the filter (control, 16.6±1.1 vs histamine, 10−8 mol/L, 40.9±0.9 monocytes/high power field). Cimetidine, an H2 receptor blocker, attenuated (P<0.001) the effect of histamine on the migration of monocytes. PGE2 inhibited the migration of monocytes in a dose-dependent manner. Histamine increased (P<0.001) the passage of monocytes across the glomerular endothelial cell layer (control, 1012±37 vs 1711±163 cpm/well). Histamine also increased the migration of murine macrophages across the glomerular endothelial cell layer. PGE2 inhibited the migration of monocytes across the endothelial cell layer under basal as well as histamine-stimulated states. Dibutyryl cyclic (DBc) AMP also attenuated the migration of monocytes under basal as well as histamine-stimulated states. Both PGE2 and DBcAMP also attenuated the IL-1β-stimulated migration of monocytes. Angiotensin II, endothelin-1, and platelet-activating factor did not modulate the migration of monocytes.
Conclusions
Vasoactive agents directly modulate the transmigration of monocytes. The present in vitro study provides a basis for a hypothesis that vasoactive agents may also be modulating the migration of monocytes across the glomerular endothelial cell layer (into the mesangium).
Introduction
Focal glomerulosclerosis has been demonstrated to be a common pathway for the progression of renal injury. 1 We studied the role of histamine on the migration of monocytes across glomerular endothelial cells because the accumulation of macrophages in the mesangium has been shown to be an important event in the development of glomerulosclerosis.1–4 Moreover, the glomerular mesangium normally contains variable numbers of resident macrophages (resident mesangial cells, 85 to 95% smooth muscle-like cells, and 5 to 15% macrophages). 5 These mesangial macrophages have been shown to be marrow-derived and presumably migrate across the glomerular endothelial cell layer. 6
The glomerular capillaries contain arterioles (afferent on the arterial side and efferent on the venous side) at both ends. This characteristic allows glomerular capillaries to have a higher pressure than other visceral capillaries. Because the glomerular filtration rate is dependent on glomerular pressure, glomerular filtration can be maintained even under low perfusion states. In addition, the glomerular capillary lumen is separated from the mesangium by a fenestrated endothelial monolayer.7–9 Because the size of these endothelial pores is 365 to 500 A, plasma and its contents freely perfuse the mesangium. 9 These characteristics distinguish glomerular capillaries from other capillaries.
In the present study, we examined the migration of monocytes loaded with IgG complexes across the glomerular endothelial cell layer. IgG complexes have been demonstrated to express properties similar to immune complexes. 10 Localized inflammation such as glomerulonephritis is a common site for the aggregation of mononuclear cells. Because histamine is one of the key mediators of inflammation, we studied the effect of histamine on the migration of monocytes. And because endothelial cells as well as mesangial cells synthesize prostaglandins, we also examined the effect of PGE2 on the passage of monocytes across endothelial cells under basal as well as histamine-treated states.
Methods
Monocyte Culture
We used an established human monocyte cell line U 937 and murine macrophage cell line J774.16 (American Type Culture Collection, Bethesda, Md) in the migration studies. Monocytes were grown in RPMI containing 10% fetal calf serum and 1% penicillin/streptomycin.
The following experimental agents were used: histamine stocked in a concentration of 10−2 mol/L and used in concentrations of 10−4 to 10−12 mol/L (Sigma); diphenhydramine stocked in a concentration of 10−3 mol/L in normal saline and used in a concentration of 10−5 mol/L (Sigma); cimetidine stocked in a concentration of 10−2 mol/L and used in a concentration of 10−5 mol/L (Sigma); PGE2 was stocked in a concentration of 10−1 mol/L in ethanol and used in concentrations of 10−6 to 10−10 mol/L (Sigma); dibutyryl adenosine 3',5'-cyclic monophosphate (DBcAMP; Sigma), stocked at 10−2 mol/L and used at a concentration of 5×10−4 mol/L. Angiotensin II (Sigma) was stocked in 0.01% acetic acid and used in concentration of 10−6 mol/L (Sigma); endothelin-1 and platelet-activating factor (PAF; Sigma) were used in a concentration of 10−6 mol/L; interleukin-1β (IL-1-β, R & D Systems, Minneapolis, Minn) was used in a concentration of 5 µg/mL. 125I was obtained from NEN Products (Boston, Mass), and human IgG (Armour Pharmaceutical Co., Kankakee, Ill) was stocked in a concentration of 165 mg/mL.
Migration Studies across the Filter
To determine the effect of histamine on the migration of monocytes into the mesangium, we created a situation where monocytes were allowed to migrate from an adjacent compartment separated only by a porous 5-µm pore size, polycarbonate, PVP-free Nucleopore filter (Poretics, Livermore, Calif). Filters were prepared by soaking overnight in a solution of 100 µg/mL gelatin in 0.1% acetic acid and air dried. We used a modified Boyden chamber (Neuro Probe Inc., Cabin John, Md) in these studies.
Two hundred microliters of RPMI-BSA (RPMI with 0.1% BSA) containing either vehicle (control) or histamine (10−5 mol/L) was added to the lower compartment of a Boyden chamber. The upper compartment was filled with 100 µL of RPMI-BSA containing 1×10−5 monocytes/mL. At the end of the incubation period (120 minutes), the cell suspension was aspirated by pipette and the cells were detached from the upper surface of the filter with the help of a cotton-tipped swab. Subsequently, cells at the under surface of the filter were stained with Diff Quick (Baxter, Chicago, Ill) and the migrated monocytes counted under a light microscope. Seven fields on each filter were recorded by two observers who were unaware of the experimental conditions.
Preparation of in Vitro Model of Glomerular Mesangium
To simulate in vivo conditions, we performed migration studies across glomerular endothelial cell monolayers grown on collagen gels. In these experiments we used isolated blood peripheral mononuclear cells (to simulate in vivo conditions). Because uptake of IgG complexes by circulating monocytes is a common occurrence, we used radiolabeled aggregated IgG (AHIgG125I) for labeling monocytes and murine macrophages. We also performed transmission electron microscopic studies to confirm the uptake of IgG complexes by monocytes. However, we used IgG-coated gold particles in those studies. Ten milliliters of blood from healthy volunteers was collected and mononuclear cells from peripheral blood (PBMC) were isolated.
Colloidal gold particles 5-10 nm in diameter were prepared by the method of Horisberger and Vonlanthen.¹¹ Gold particles were coated with IgG as described previously. ¹β,¹³ In addition, iodinated aggregated human IgG (AHIgG¹β5I) was prepared as described previously.¹4 PBMCs (106) were labeled with AHIgG¹β5I (1 µCi/mL). To confirm the uptake of aggregated IgG complexes by monocytes in parallel studies, aliquots of PBMC were incubated with IgG-coated gold particles for 1 hour at 37°C. At the end of the incubation, cells were thoroughly washed with PBS and prepared for transmission microscopy studies as described previously. ¹β,¹³
Rat glomerular endothelial cells were kindly provided by Dr. Harry Holthofer (Department of Bacteriology and Immunology, University of Helsinki, Helsinki, Finland). These cells were characterized by staining positively with antibodies against factor VIII-related antigen, monoclonal anti-rat endothelium (OX-43), anti-gp350, and with FITC-BSI-B4 lectin.¹5 Before the use of these cells in the migration studies, we also stained these cells with antibodies against von Willebrand factor (Accurate Chemical Corp., Westbury, NY). These cells showed uniformly positive staining for von Willebrand factor.
Aliquots of the suspended cells were plated on fibronectin-coated petri dishes. Once cells were confluent, they were trypsinized (0.05% trypsin + 0.5 mmol EDTA solution) and plated on collagen gels.
Eight milliliters of purified collagen (Vitrogen, Santa Clara, Calif), 1 mL of 10 X M199, and 5 mL of 0.1 N NaOH were mixed in a 15-mL tube. A 50-µL aliquot of this mixture was instilled in each well of a 96-well culture plate. The culture dish was kept in an incubator at 37°C for 1 hour. A 100-µL aliquot of M199 was added to each well and the plate was kept in an incubator for overnight equilibration. Subsequently, wells were coated with fibronectin and plated with endothelial cells. Endothelial cells were able to cover the collagen gel during the next 5 to 7 days. The gel was checked for continuous growth (100% coverage of gel by cells) of endothelial cells before starting the migration studies.
Migration Studies across the Endothelial Cells
Muller et al in their previous elegant studies demonstrated that among PBMC, only monocytes migrate across the endothelial cells during a period of 60 minutes.¹ 6 Therefore, we used labeled PBMC in these studies and described them as labeled monocytes for migration purposes. Incubation media from cells grown on collagen were drained gently and cells were washed with HBSS buffer twice; 100-µL aliquots of freshly labeled PBMC in HBSS containing either vehicle alone (control) or histamine (10−5 mol/L) were added to each well, and cells were kept at 37°C for 60 minutes. Six sets of experiments were performed, each in triplicate. At the end of the incubation, the reaction was stopped by shaking the supernatant out over an absorbent mat. Each well was then filled with about 400 µL warm 1 mmol EGTA in HBSS. A seal plate film was then placed over the plate, the plates were inverted and centrifuged at 250g for 5 minutes. This process removed both nonadherent cells as well as cells adherent to the apical surface.¹ 6 Cells beneath the gel and cells in the process of migrating were retained. The collagen gel was collected with swab sticks and radioactivity counted for each well.
Statistical Analysis
For comparison of mean values between two groups, the unpaired t test was used. To compare values between multiple groups, ANOVA was applied and a Newman-Keuls multiple range test was used to calculate a q value. All values are mean±SE, except where otherwise indicated. Statistical significance was defined as P<0.05.
Results
Transmission Electron Microscopic Studies
Transmission electron microscopic studies confirmed intracellular localization of IgG-coated gold particles in monocytes. A representative electron micrograph showing uptake of IgG-coated gold particles is shown in Figure 1.
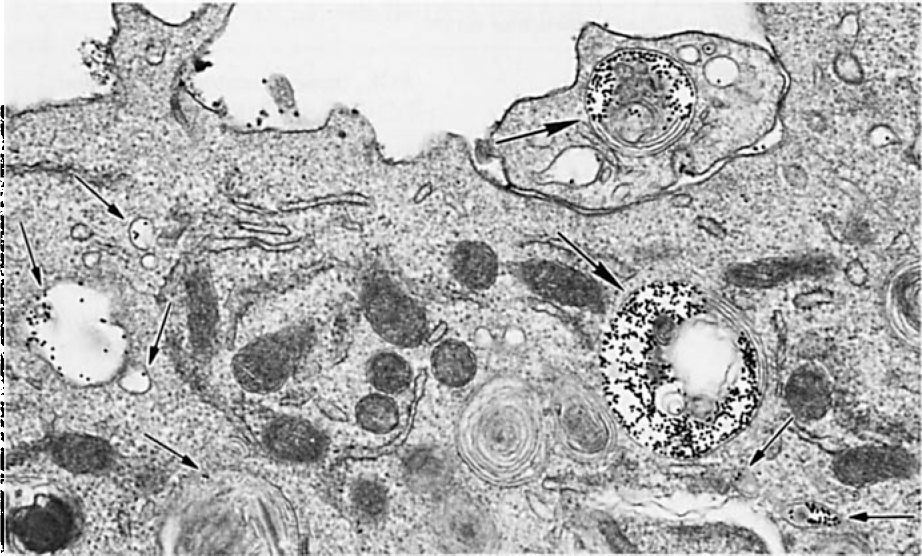
Representative electron micrograph showing uptake of IgG-coated gold particles by a monocyte. Uptake of gold particles is indicated by arrows (in vesicles and endolysosomes). Magnification×10,000.
Migration of Monocytes across a Filter
Histamine increased (P<0.003) the migration of monocytes across the filter in a modified Boyden chamber (control, 11.1±0.7 vs 19.7±1.8 monocytes/HPF). To determine the dose-response effect of histamine on the migration, 200 µL of RPMI-BSA containing variable concentrations of histamine (10−4 to 10−12 mol/L) was instilled in the lower compartment; 100 µL of RPMI-BSA containing equal numbers of monocytes (2×105 cells/mL) were placed in the upper compartment. As shown in Figure 2A, histamine in concentrations of 10−12 to 10−4 mol/L enhanced (P<0.001) the migration of monocytes across the filter. This happened in a dose-dependent manner. The optimal effect of histamine on the migration of monocytes occurred at concentrations of 10−8 to 10−5 mol/L.
The effect of histamine on cells is mediated through H1 and H2 receptors.¹7 We examined whether the migration of monocytes across the filter is a receptor-mediated event. An aliquot (200 µL) of RPMI-BSA containing vehicle alone (control), histamine (10−5 mol/L), histamine (10−5 mol/L) plus diphenhydramine (H1 blocker, 10−5 mol/L), or histamine (10−5 mol/L) plus cimetidine (H2 blocker, 10−5 mol/L) was added to the lower compartment. RPMI-BSA (100 µL) containing equal numbers of monocytes (1×105/mL) was added to the upper compartment. As shown in Figure 2B, histamine increased (P<0.001) the migration of monocytes across the filter. The H2 blocker attenuated (P<0.001) the effect of histamine on migration of monocytes. The H1 blocker did not alter the effect of histamine on the migration of monocytes. These results suggest that the histamine-induced enhanced mobility of monocytes is mediated via H2 receptors.
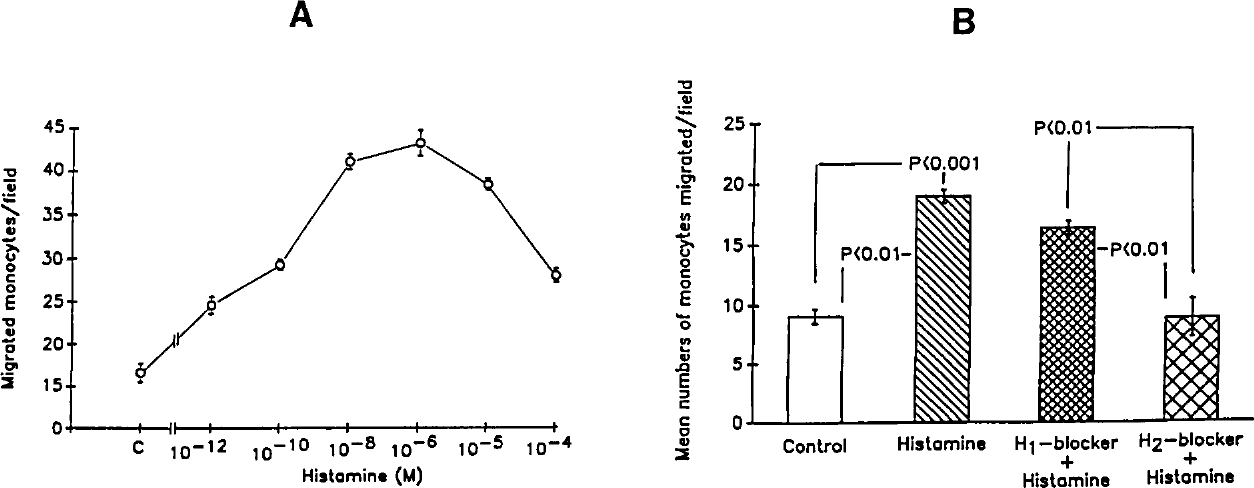
A, Dose-response effect of histamine. Results represent mean±SE from four sets of experiments. B, Effect of histamine blockers on migration of monocytes. Results represent the mean±SE from four series of experiments.
Because PGE2 is the predominant prostaglandin synthesized by endothelial as well as mesangial cells under a basal state, we studied whether PGE2 may be modulating the migration of monocytes under basal or stimulated states. To determine the toxicity of PGE2 on PBMCs and U937 cells, equal numbers of cells were treated with either buffer or PGE2 (10−6 mol/L) for 120 minutes. At the end of the incubation period, cells were stained with Trypan Blue. There was no difference in uptake of Trypan Blue by control and PGE2-treated PBMCs/U937 cells (PBMCs, control, 2.8±0.2%; PGE2, 3.8±0.5%; U937 cells, control, 0.70±0.2%; PGE2, 0.97±0.3% cells). To determine the dose-response effect of PGE2 on the migration of monocytes across a filter, aliquots (200 µL) of RPMI-BSA were instilled in the lower compartment. Aliquots (100 µL) of RPMI-BSA containing variable concentrations of PGE2 (0 to 10−10 mol/L) plus an equal number of monocytes (2×105 cells/mL) were added in the upper compartment. As shown in Table 1, PGE2 in concentrations of 10−8 to 10−6 mol/L inhibited (P<0.001) the migration of monocytes across the filter.
To determine the effect of PGE2 on the histamine-induced migration of monocytes, aliquots (200 µL) of RPMI-BSA containing histamine (10−8 mol/L) were added in the lower compartment. Aliquots (100 µL) of RPMI-BSA containing variable concentrations of PGE2 (0 to 10−10 mol/L) plus equal numbers of monocytes (2×105 cells/mL) were added in the upper compartment. As shown in Table 1, histamine did not enhance the migration of monocytes in the presence of PGE2 (10−8 to 10−6 mol/L). These results indicate that PGE2 attenuates the migration of monocytes, under both basal as well as stimulated states.
Effect of PGE2 on monocyte migration under basal as well as histamine-stimulated states.
Results represent the mean±SE from 4 sets of experiments.
P<0.001 compared with control and PGE2, 10−10 mol/L.
P<0.05 compared with histamine + PGE2 (10−10 mol/L).
Because cAMP acts as a mediator of many of the effects of PGE2 in monocytes,¹8 we studied whether PGE2-modulated monocyte migration is also mediated through the generation of cAMP. To determine the effects of DBcAMP on the migration of monocytes under basal as well as histamine-induced states, aliquots (200 µL) of RPMI-BSA containing either vehicle (control) or histamine (10−8 mol/L) were instilled in the lower compartment. Aliquots (100 µL) of RPMI-BSA containing either vehicle (control) or DBcAMP (5×10−4 mol/L) plus an equal number of monocytes (2×105 cells/mL) were added in the upper compartment. As shown in Table 2, DBcAMP inhibited (P<0.05) the migration of monocytes in the basal state (control, 18.3±1.7 vs DBcAMP, 13.6±0.5 monocytes/HPF). DBcAMP also inhibited (P<0.001) the migration of monocytes in the histamine-treated state (Histamine, 28.1±2.7 vs histamine + DBcAMP, 13.0±0.3 monocytes/HPF).
Effect of DBcAMP on monocyte migration.
Results represent the mean±SE from 4 sets of experiments.
P<0.001 compared with histamine.
P<0.05 compared with control.
P<0.01 compared with histamine.
To determine whether the effect of cAMP and PGE2 on the migration of monocytes is a result of the attenuation of histamine chemoactivity or because of the modulation of the mobility of monocytes as a consequence of cytoskeletal alteration, we studied the effect of PGE2 and cAMP on basal as well as interleukin-1β-induced migration of monocytes. Aliquots (200 µL) of RPMI-BSA containing either buffer (control) or IL-1β (5 µg/mL) were instilled in the lower compartment. Aliquots (100 µL) of RPMI-BSA containing buffer (control), PGE2 (10−6 mol/L), or DB-cAMP (5×10−4 mol/L) plus equal numbers of monocytes (2×105 cells/mL) were added in the upper compartment. Four sets of experiments were performed, each in triplicate. Interleukin-1β enhanced (P<0.001) the migration of monocytes across the filter (Table 3). Both PGE2 and DBcAMP attenuated the migration of monocytes across the filter in the basal as well as the stimulated state (Table 3).
Effect of PGE2 and DBcAMP on interleukin-1β-induced monocyte migration.
Results represent the mean±SE from 3 sets of experiments.
*P<0.003 compared with control.
P<0.001 compared with control.
P<0.01 compared with IL-1.
P<0.05 compared with DBcAMP.
P<0.001 compared with IL-1.
To determine the dose-response effect of IL-1β on monocyte (U937) migration, aliquots containing variable concentrations of IL-1β (0.01, 0.1, and 1 µg/mL) were instilled in the lower compartment (n=7). IL-1β promoted the migration of U937 in a dose-dependent manner (control, 13.5±1.6; IL-1β, 0.01 µg/mL, 19.5±2.5; IL-1β 0.1 µg/mL, 26.1±1.5; IL-1β, 1.0 µg/mL, 33.0±2.8 cells/field). To further confirm the effect of IL-1β, aliquots of buffer, IL-1β (0.1 µg/mL), or IL-1β plus anti-IL-1β antibody (1 µg/mL) were instilled in the lower compartments. Anti-IL-1β antibody completely inhibited IL-1-β-induced monocyte migration across the filter (control, 17.7±2.2; IL-1β, 29.6±3.3; anti-IL-1β + IL-1β, 8.9±1.4 cells/field).
Migration across Endothelial Cells
Monocytes treated with histamine (10−5 mol/L) showed increased (P<0.001) migration through the endothelial cell layer (control, 1012±37 vs 1711±163 cpm/well). These results suggest that histamine also enhances the migration of monocytes across the glomerular endothelial cell layer.
To determine the effect of PGE2 in the passage of monocytes across endothelial cells, cells were incubated with freshly labeled monocytes in HBSS (100 µL) containing either vehicle alone or PGE2 (10−8 mol/L) for 60 minutes at 37°C. Three to five sets of experiments were performed, each in triplicate. PGE2 inhibited (P<0.001) the migration of monocytes across the endothelial cell layer (control, 1053±61 vs PGE2, 493±21 cpm/well).
To determine the effect of PGE2 on histamine-induced migration of monocytes, confluent endothelial cells grown on gels were incubated with freshly labeled monocytes in HBSS (100 µL) containing vehicle alone (control), histamine (10−5 mol/L) or PGE2 (10−5 mol/L) plus histamine (10−5 mol/L) for 60 minutes at 37°C. Three to five sets of experiments were performed, each in triplicate. PGE2 attenuated (P<0.001) the effect of histamine on the migration of monocytes across the endothelial cell layer (control, 1053±61 cpm/well; histamine, 10−5 mol/L, 1811±155 cpm/well; PGE2, 10−5 mol/L + histamine, 10−5 mol/L, 285±26 cpm/well).
To evaluate whether the effect of histamine on glomerular endothelial cells is specific or not, we examined the effect of other vasoconstrictors such as angiotensin II, endothelin-1 and PAF on the migration of monocytes across glomerular endothelial cells. In these studies we used labeled monocytes (U 937). One hundred-microliter aliquots of freshly labeled U937 in HBSS containing buffer (control), angiotensin II (10−6 mol/L), endothelin-1 (10−6 mol/L), or PAF (10−6 mol/L) were added to each well and cells were kept at 37°C for 60 minutes. Five sets of experiments were performed, each in triplicate. Angiotensin II (10−6 mol/L), endothelin-1 (10−6 mol/L), and PAF (10−6 mol/L) did not increase the migration of monocytes across the endothelial cell layer when compared to control (control, 962.6±80.9; angiotensin II, 1119.2±228.3; endothelin-1, 1065.8±224.4; PAF, 991.6±113.6 cpm/well). These results indicate that vasoconstrictors do not affect the migration of monocytes across the glomerular endothelial cell layer. It appears that histamine has a specific effect on the migration of monocytes across the cell layer.
To evaluate whether the effect of histamine on the migration of monocytes is species-specific, we used murine macrophages (J774.16 cells labeled with radiolabeled IgG) in additional studies. One hundred microliters of PBS containing variable concentrations of histamine (10−12 to 10−4 mol/L and zero) plus equal numbers of macrophages (¹β5IgG-labeled) were added to the wells containing endothelial cell monolayers grown on collagen gels in a 96-well plate. Incubation was terminated after 1 hour. As shown in Figure 3, histamine at concentrations of 10−8 to 10−4 mol/L enhanced (P<0.001) the migration of murine macrophages across the endothelial cell layer. These results suggest that the effect of histamine on the migration of monocytes is not species-specific.
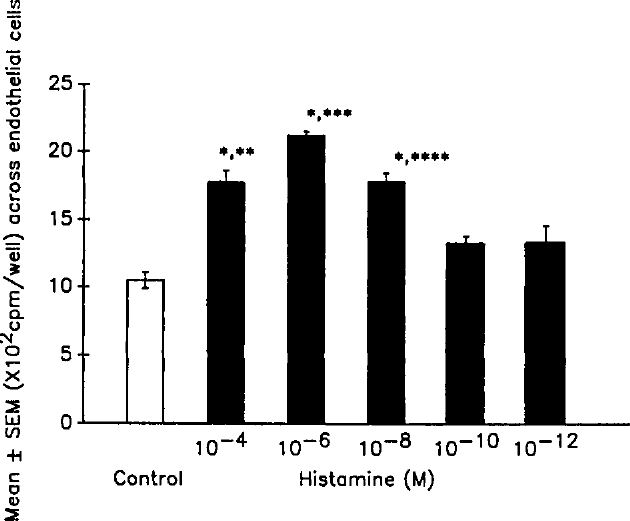
Effect of histamine on migration of murine macrophages across the endothelial cell layer. Results represent mean±SEM from three sets of experiments performed in triplicate.
Discussion
The present study demonstrates that histamine enhances the passage of monocytes across a filter. This happened in a dose-dependent manner. Because cimetidine, an H2 receptor blocker, could inhibit this effect, it appears that H2 receptors play a role in the histamine-induced mobility of monocytes. Histamine also enhanced the passage of monocytes across glomerular endothelial cells. Prostaglandin E2 inhibited the movement of monocytes across endothelial cells in both basal as well as histamine-stimulated states. Both PGE2 and DBCAMP also attenuated IL-1β-induced migration of monocytes across the filter. Because DBcAMP inhibited the migration of monocytes across the filter under basal as well as histamine-stimulated states, it appears that the effect of PGE2 may be mediated through cAMP.
Histamine has been reported to modulate the function of monocytes in a variety of ways. Histamine enhances the production of granulocyte macrophage colony-stimulating factor and IL-6 by mononuclear cells.19 It suppresses gene expression and synthesis of tumor necrosis factor by lipopolysaccharide-stimulated peripheral blood mononuclear cells and monocytes. 20 It has also been demonstrated to enhance the production of interleukin-1 by mouse peritoneal macrophages.β¹ The addition of histamine to macrophage-like P388d1 cells resulted in a dose-dependent increase in intracellular calcium.ββ Many of the effects of histamine on macrophages may be mediated through the increase of intracellular calcium.
An increased generation of histamine is a common finding in inflammatory conditions. Because there is no published data about the generation of histamine in the mesangium, the role of histamine in the migration of monocytes across the glomerular endothelium remains theoretical at the present time. The glomerular mesangium is inhabited by mesangial cells and macrophages. Uptake of immune complexes by macrophages as well as mesangial cells has been associated with the release of a variety of chemoattractants (including IL-1β and TGF-β), proinflammatory cytokines, prostaglandins, and reactive oxygen species. The present study suggests that IL-1β may induce the migration of monocytes. In addition, histamine-induced endothelial cell contractility widens the gap (fenestrae) between the cells: the widened endothelial fenestrae may provide a path for the passage of monocytes.β³ Endothelial cells have specific receptors for histamine. The binding of histamine to receptors has been demonstrated to be associated with increased cytosolic calcium.β³ Increased cytosolic calcium has been demonstrated to trigger a contractile response.β³ Because PGE2 and DBCAMP attenuated the migration of monocytes under basal as well as stimulated states, these agents may directly be affecting the mobility of monocytes rather than reducing the chemotactic action of histamine and IL-1.
In Boyden chamber experiments, histamine augmented the migration of monocytes across the filter at a concentration of 10−12 mol/L. This effect of histamine was a result of chemotaxis. However, in the experiments related to glomerular endothelial cells, the effect of histamine was significant only at concentrations of 10−6 mol/L or higher. This effect of histamine may be related to the contractile effect of histamine on endothelial cells.
In the present study, prostaglandin E2 inhibited the passage of monocytes across endothelial cells both under basal and histamine-stimulated states. Endothelial cells as well as mesangial cells secrete PGE2. It seems that release of PGE2 by these cells may be serving as a feedback mechanism to regulate the influx of monocytes into the mesangium.
We conclude that vasoactive agents may modulate the migration of monocytes across glomerular endothelial cells. The present study indicates that vasoactive agents, in addition to their hemodynamic effect, have the potential to contribute to glomerular injury through the modulation of macrophage migration into the mesangium.
Footnotes
Acknowledgment
This work was supported by grant RO1 DA 12111 from the National Institutes of Health.