
Abstract
α
Introduction
ανβ3 integrins have been implicated in regulating smooth muscle cell (SMC) growth responses following vascular injury. Expression is prominent in diseased human coronary arteries1 and following balloon injury in animal models.2–4 Inhibition of ανβ3 integrins by cyclic Arg-Gly-Asp (RGD) peptide antagonists decreased neoin-timal formation by ≈70% in a rabbit carotid artery injury model,5 by 99% in a rat carotid injury model,3 and by 43% in a porcine coronary injury model.4 Finally, ανβ3 integrins regulate proliferative and migratory3,5–9 responses of cultured SMC to a variety of agonists.
ανβ3 integrins may regulate cellular responses via formation of cytoplasmic protein complexes involving nonreceptor kinases. This hypothesis postulates that association of proteins in these complexes regulates tyrosine phosphorylation and recruitment to focal adhesions of nonreceptor kinases, leading to activation of downstream signaling pathways that stimulate proliferation or migration. The formation of protein complexes has been shown to regulate various intracellular signaling pathways, most notably those associated with tyrosine kinase10 and G protein-coupled receptors.11 There is presumptive evidence that they are important in integrin signaling processes.12
ανβ3 integrin have been shown to associate with cytoplasmic proteins under specific conditions. Vuori and Ruoslahti13 reported that tyrosine phosphorylated insulin receptor substrate-1 associated with ανβ3 integrins upon exposure of HIRCB cells to insulin. Moreover, insulin-induced DNA synthesis was 2.5-fold higher in cells expressing the ανβ3 integrin when plated on vitronectin (a ligand for ανβ3) than on other substrates. In cultured fibroblasts, Bartfeld et al14 showed that treatment with platelet-derived growth factor-BB resulted in phosphory-lation of a 190 kD protein that co-immunoprecipitated with ανβ3 integrins.
Nonmuscle myosin heavy chain-A (NMHC-A) has many characteristics suggesting it may function in ανβ3 integrin-mediated signaling. First, NMHC-A binds peptides corresponding to residues 740-762 of the cytoplasmic domain of β
On the basis of these data we hypothesized that NMHC-A plays an important role in ανβ3 integrin-mediated signaling by linking cell surface integrins with nonreceptor kinases. We tested this hypothesis by examining effects elicited by thrombospondin-1 (TSP), a glycoprotein that stimulates SMC proliferation via ανβ3 integrin-dependent mechanisms.2 Specifically, we sought to determine whether treatment with TSP caused: 1) NMHC-A to associate with ανβ3 integrins; 2) focal adhesion kinase (FAK), a nonreceptor kinase that has been implicated in integrin-mediated signaling in platelets and fibroblasts, to phosphorylate on tyrosine; and 3) NMHC-A to associate with FAK. Lastly, we also examined NMHC-A expression following vascular injury.
Methods
SMC Culture
Human aortic SMC were obtained from Clonetics Corporation (San Diego, Calif) and maintained in culture as previously described.2 Briefly, SMC between passages 4 and 15 were plated at 3.5×10
Protein Isolation and Western Blotting
Cells were lysed in a modified RIPA buffer (NaCl 150 mmol/L, Tris-HCl 50 mmol/L [pH 7.4], EGTA 1 mmol/L, Na deoxycholate 0.25%, NP-401%, Aprotinin 10 µg/mL, PMSF 1 mmol/L, Na orthovanadate 1 mmol/L, and NaFl 1 mmol/L) and the cell lysate centrifuged at 14000 rpm for 10 minutes at 4°C. Protein concentration was determined by the Bio-Rad DC protein assay kit and adjusted to 1 mg/mL. The designated antibody was added in excess and incubated for 2 hours at 4°C. Immune complexes were recovered by the addition of 50 µL of protein A-Agarose (UBI, Lake Placid, NY) and incubation for 2 hours at 4°C. The beads were recovered by a brief spin in a microcen-trifuge and washed twice with RIPA buffer without protease and phosphatase inhibitors. The immunoprecipitated proteins were eluted by boiling in sample buffer (50 mmol/L Tris-HCl [pH 6.8], 100 mmol/L DTT, 2% SDS, 0.1% bromophenol blue, and 10% glycerol), electrophoresed on a 7.5% SDS-polyacrylamide gel and transferred to nitrocellulose by electroblotting. Subsequently, the blot was blocked in 0.5% nonfat milk. After immunoblotting with the primary antibody and an HRP-conjugated sec-ondary antibody, immunodetection was performed via ECL Western blotting detection reagents (Amersham), followed by autoradiography.
Reagents
Antibodies were purchased from the following sources: Anti-NMHC-A (BT561), Biomedical Technologies (Stoughton, Mass); LM609, Chemicon International (Te-mecula, Calif); anti-FAK and 4G10, Upstate Biotechnol-ogy (Lake Placid, NY); anti-β
Baboon Arterial Injury Model and Immunohistochemistry
Balloon catheter denuding injury of the left brachial artery of juvenile male baboons (Papio anubis) weighing 8-12 kg was performed at Yerkes Regional Primate Re-search Center as previously described.2 Slides of arterial sections were preincubated in 500 µg/mL trypsin for 30 minutes at 37°C, then incubated in methanol containing 1% H
Data Analysis
Numerical data are presented as mean±SE unless otherwise stated. One-way ANOVA followed by the New-man-Keuls' multiple range test was used to analyze for statistically significant differences (P≤0.05).
Results
NMHC-A Associated with ανβ3 Integrins in TSP-Treated Human Aortic SMC
Previous studies showed that peptides corresponding to the cytoplasmic domain of β
We found that NMHC-A was immunoprecipitated by LM609 in a time-dependent manner following treatment with TSP (Figure 1). Similar results were obtained when Western analysis was performed using a different anti-NMHC-A antibody20 raised by immunizing rabbits with peptides synthesized based on the deduced amino acid sequence of the carboxyl-terminal portion of human NMHC-A (GKADGAEAKPAE) conjugated to keyhole limpet hemocyanin with glutaraldehyde (data not shown).
TSP isolated from platelets is commonly contaminated by transforming growth factor-β (TGFβ), and therefore our preparations were subjected to an additional gel permeation step (in which a Bio-Gel A 0.5-m [Bio-Rad, Hercules, Calif] column is equilibrated at pH 11 to disassociate TGFβ) using the method described by Schultz-Cherry and Murphy-Ullrich.23 Levels of total and activated TGFβ were measured and were less than 0.15 ng/mL at the concentrations of TSP used in our experi-ments.
Nonspecific immunoprecipitation of myosin occurred in some (but not all) experiments (Figure 2) and persisted despite preclearing of lysates with beads alone, preclearing with IgG and beads, changing the constituents of the lysis buffer, and increasing the number of washes. This effect, however, was inconsistent and did not show a time-dependent relationship.
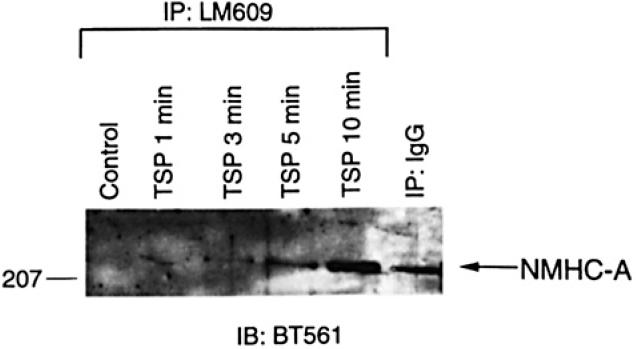
Further evidence for a specific interaction between β3 integrin subunits and NMHC-A was provided by studies in which immunoprecipitation was performed with anti-NMHC-A antibodies. β3 integrin sub-units were immunoprecipitated by BT561 and the levels were increased 5 minutes after TSP treatment (Figure 3). β3 integrin sub-units were identified using clone 1, a monoclonal antibody that recognizes an epitope between amino acids 592 and 712 of human β3 integrin. Immunoblotting resulted in a single band at the expected size for β3 integrins.
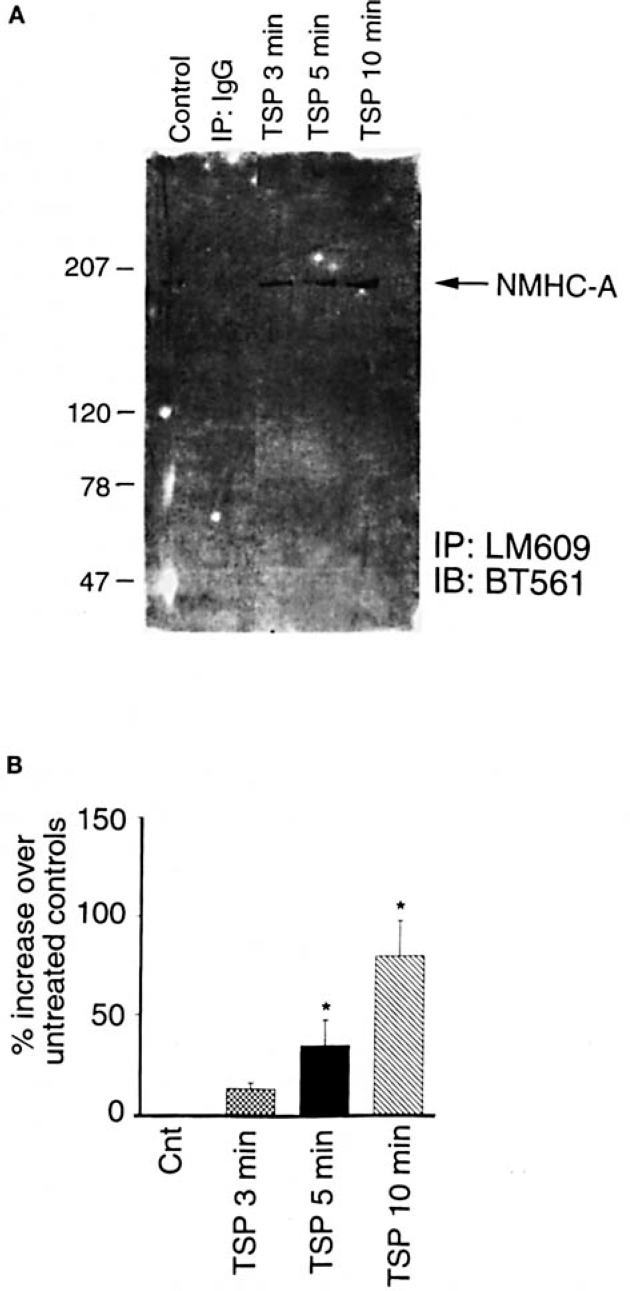
Association of NMHC-A with a •./3J integrins following treatment with TSP. Human aortic SMC were grown to confluence, growtharrested, and treated with TSP (50 p.g/mL). SMC cultured under the same conditions but not exposed to TSP served as controls. At various times, the cells were lysed and immunoprecipitation performed using LM609. The proteins isolated were subjected to Western analysis using anti-NMHC-A antibodies (BT-56}). A, Representative blot; B, result of 6 independent experiments. Cnt indicates control. *P < 0.05 compared with control group.
Phosphorylation on Tyrosine of FAK Increased following Treatment of Human Aortic SMC with TSP
FAK is a 125 kD intracellular kinase that has been linked to integrin-mediated signaling in fibroblasts and platelets (reviewed by Hanks et al24). To determine if treatment of human SMC with TSP resulted in phosphorylation of FAK, we performed immunoprecipitation with an anti-phosphotyrosine antibody (PY20) followed by im-munoblotting with a commercially available anti-human FAK antibody raised in rabbits immunized with a pGEX-derived fusion protein containing residues 748-1053 of human FAK. We found that FAK was constitutively phos-phorylated on tyrosine in cultured human SMC (Figure 4), consistent with previous studies in quiescent SMC from rats,25,26 rabbits,27 and humans.27 Levels of FAK phosphorylated on tyrosine increased following TSP treatment in a time-dependent manner. The specificity of the inter-action between PY20 and tyrosine phosphorylated FAK was confirmed by further studies including: 1) immuno-precipitation of FAK was blocked by pretreatment of PY20 with excess phosphotyrosine peptides (100 µmol/L O-phospho-L-tyrosine; Figure 4A); 2) similar results were obtained using a different anti-phosphotyrosine antibody (4G10; Figure 4B); and 3) similar results were obtained when immunoprecipitation was performed with anti-FAK followed by Western analysis with PY20 (Figure 5).
Comparison of specific and nonspecific immunoprecipitation of NMHC-A. Human aortic SMC were grown to confluence. growtharrested, and treated with TSP (50 μg/mL). SMC cultured under the same conditions but not exposed to TSP served as controls. At various times. the cells were lysed and immunoprecipitation performed using LM609 or nonspecific IgG. The proteins isolated were subjected to Western analysis using anti-NMHC-A antibodies (BT-56/).
β3 integrins were immunoprecipitated by anti-NMHC-A antibodies. Human aortic SMC were grown to confluence. growth-arrested. and treated with TSP (50 p.g/mL).SMC cultured under the same conditions but not exposed to TSP served as controls. At various times. the cells were lysed and immunoprecipitation performed using BT56/. The proteins isolated were subjected to Western analysis using anti-B, integrin antibody (clone I).
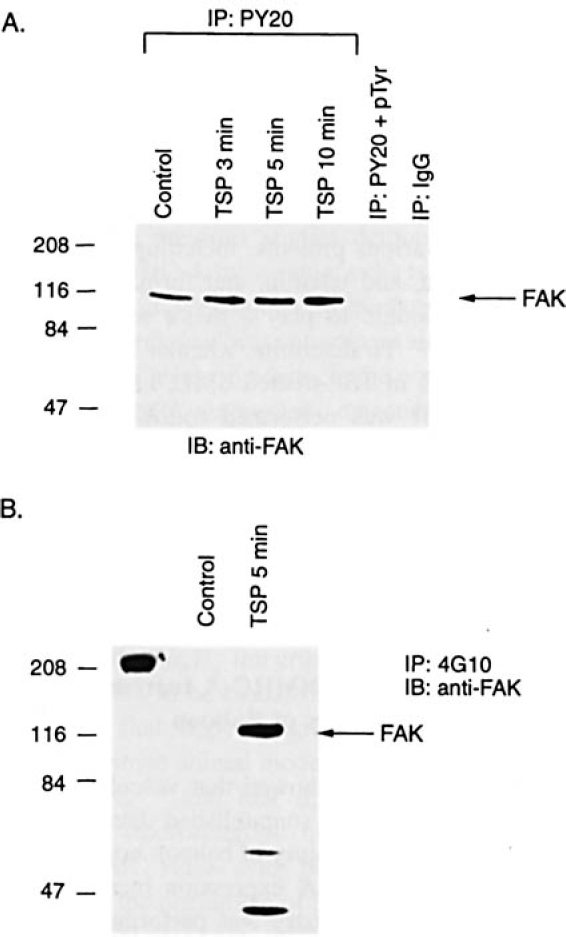
FAK phosphorylates on tyrosine following treatment of SMC with TSP. Human aortic SMC were grown to confluence. growth-arrested. and treated with TSP (50 p.g/mL).SMC cultured under the same conditions but not exposed to TSP served as controls. At various times. the cells were lysed and immunoprecipitation performed using antiphosphotyrosine antibodies (PY20 [A] or 4GlO [B]), PY20 pretreated with excess phosphotyrosine peptides (100 μmol/L: O-phopho-L-tyrosine), or nonspecific IgG. The proteins isolated were subjected to Western analysis using anti-FAK.
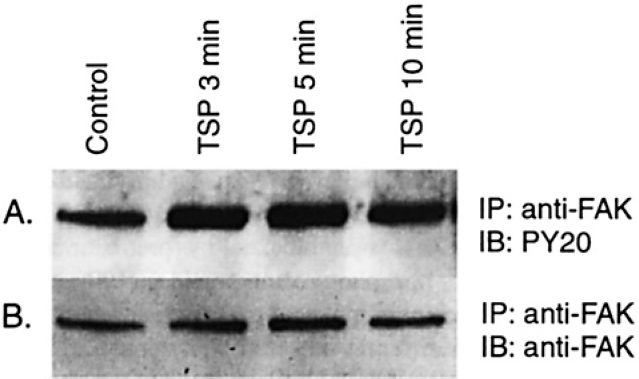
Tyrosine phosphorylation of FAK following treatment of SMC with TSP. Human aortic SMC were grown to confluence. growth-arrested, and treated with TSP (50 μg/mL). SMC cultured under the same conditions but not exposed to TSP served as controls. At various times, the cells were lysed and immunoprecipitation performed using anti-FAK antibodies. The proteins isolated were subjected to Western analysis using anti-phosphotyrosine (PY20 [A]). The membrane was stripped and reprobed with anti-FAK (B).
NMHC-A Associates with FAK in TSP-Treated Human Aortic SMC
FAK binds various proteins, including Src family ki-nases, Cas, talin, and paxillin, and formation of protein complexes is thought to play a major role in signaling through FAK.24,28 To determine whether NMHC-A associ-ated with FAK in TSP-treated SMC, immunoprecipitation with BT561 was performed followed by Western analysis with an anti-human FAK antibody. Results dem-onstrated that NMHC-A associated with FAK in a time-dependent manner following treatment with TSP (Figure 6). This association was also apparent in studies where immunoprecipitation was performed with anti-FAK fol-lowed by Western blotting with BT561 (Figure 7).
Vascular Expression of NMHC-A Increased following Balloon Injury of Baboon Brachial Arteries
Our previous studies showed that vascular expression of β3 integrins2 and TSP (unpublished data) increased 1 week following balloon injury of baboon brachial arteries. To determine if NMHC-A expression increased in this model, immunocytochemistry was performed using anti-NMHC-A antibodies (BT-561) on multiple sections of injured and uninjured brachial arteries from three animals. BT-561 has been shown to bind to NMHC-A with high specificity17 and binds to a single band at ≈200 kD on Western blotting of SMC lysates (data not shown). Our studies demonstrated that NMHC-A was expressed at low levels in the media and endothelium in uninjured vessels (Figure 8), similar to previous studies using human29,30 or rabbit31 arteries. Seven days after injury, NMHC-A ex-pression was increased in the media of injured artery and present in the neointima as well. Immunocytochemistry utilizing IgG from nonimmunized rabbits did not show any staining.
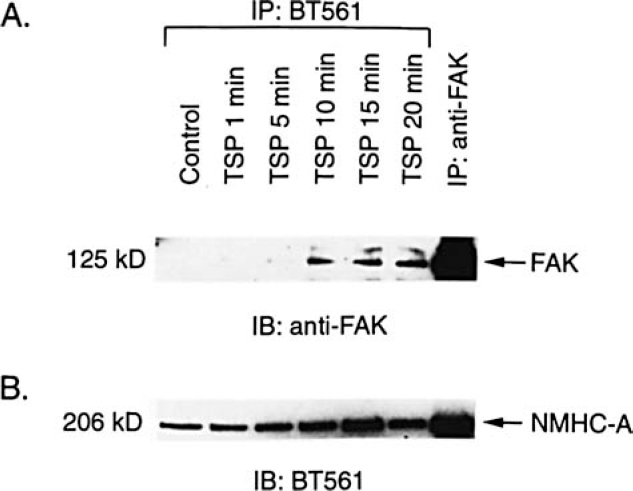
FAK was immunoprecipitated by anti-NMHC-A following treatment of SMC with TSP. Human aortic SMC were grown to confluence, growth-arrested. and treated with TSP (50 ILg/mL). SMC cultured under the same conditions but not exposed to TSP served as controls. At various times, the cells were lysed and immunoprecipitation performed using anti-NMHC-A (BT56l) or anti-FAK. The proteins isolated were subjected to Western analysis using anti-FAK (A). The membrane was stripped and reprobed with BT561 (B).
Discussion
The present studies demonstrate that NMHC-A expres-sion increased following vascular injury in the baboon and that NMHC-A associated with ανβ3 integrins and FAK in a time-dependent fashion following TSP-treatment of cul-tured SMC. These results support the hypothesis that ανβ3 integrins regulate growth responses to TSP by controlling the formation of cytoplasmic protein complexes involving nonreceptor kinases and that NMHC-A links integrin ac-tivation and intracellular signaling pathways.
Our data strongly suggest that there is a specific, phys-iological interaction between NMHC-A and β3 integrins. Support for this conclusion includes: 1) the amount of NMHC-A immunoprecipitated by LM609 consistently showed a time-dependent increase following treatment with TSP (Figure 1); 2) in the experiments in which there was nonspecific immunoprecipitation of NMHC-A, the amount of NMHC-A immunoprecipitated by LM609 was consistently greater than that immunoprecipitated by IgG (Figure 2); 3) NMHC-A was immunoprecipitated by LM609 in experiments in which NMHC-A was not im-munoprecipitated by IgG (Figure 1); 4) β3 integrins were immunoprecipitated by anti-NMHC-A antibodies follow-ing treatment with TSP (Figure 3); 5) NMHC-A binds ex vivo to peptides corresponding to the cytoplasmic domain of β3 integrins;15 and 6) NMHC-A associates with β3 integrins in thrombin-treated rat aortic SMC, human em-bryonic kidney cells transfected with pcDNA-1neo con-structs encoding full-length β3 integrin subunits, and hu-man aortic SMC transduced with pLXIN, a bicistronic retroviral vector encoding human β3 integrins with a car-boxyl-terminal FLAG recognition sequence (data not shown). NMHC-A is the second protein identified that associates with ανβ3 integrins in intact cells and the first protein identified that associates with ανβ3 integrins in SMC.
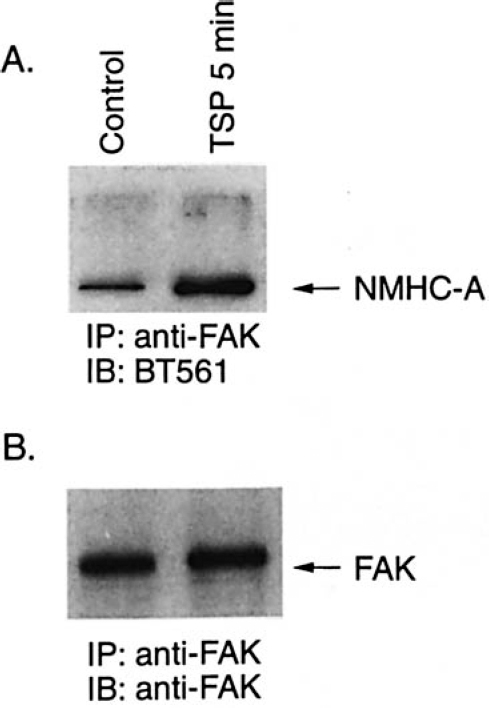
NMHC-A was immunoprecipitated by anti-FAK following treatment ofSMC with TSP. Human aortic SMC were grown to confluence, growth-arrested, and treated with TSP (50 μg/mL). SMC cultured under the same conditions but not exposed to TSP served as controls. At various times, the cells were lysed and immunoprecipitation performed using anti-FAK. The proteins isolated were subjected to Western analysis using anti-NMHC-A (BT561 [A]). The membrane was stripped and reprobed with anti-FAK (B).
Functional evidence for the importance of NMHC-A in regulating SMC proliferation was provided by Simons and Rosenberg.32 They showed that serum-induced prolifera-tion of rat aortic SMC could be inhibited ≈60% by treat-ment with an 18-mer antisense phosphorothiolate comple-mentary to nucleotides 232-250 of human NMHC-A and 95% complementary to human NMHC-B. The inhibitory effect on growth was observed only if the NMHC anti-sense oligonucleotides were added to growth-arrested cells before serum stimulation; proliferation of actively grow-ing cells was not inhibited by antisense to NMHC. Treat-ment with antisense to NMHC for 2 hours inhibited pro-liferation for the initial 72 hours, but proliferative responses returned to normal by 5 days, demonstrating that the inhibitory effect was reversible.
Levels of tyrosine phosphorylation of FAK increased following treatment with TSP, extending the results of previous studies that FAK phosphorylation increased fol-lowing treatment with PDGF27 or angiotensin II.25 FAK has been implicated in controlling cellular responses (in-cluding migration, proliferation, and survival) to integrin engagement in fibroblasts and other cell lines.24 Tyrosine phosphorylation of FAK is a crucial regulatory step for activation of downstream signals and, at least in fibro-blasts, leads to activation of the ras/MAP-kinase path-way.33 The functional importance of FAK in SMC has yet to be established.
Our studies demonstrating that NMHC-A expression by baboon brachial arteries increased following injury are consistent with previous studies demonstrating that ex-pression of NMHC-A by vascular cells is regulated during development and after injury. In humans29 and rabbits,31 NMHC-A expression was prominent in the fetus and present at lower levels in adults. In the normal adult rabbit, 4-10% of SMC expressed detectable amounts of NMHC-A; however, expression increased following high cholesterol feedings34 or balloon injury.31 Studies using atherectomy specimens and nonatherosclerotic tissue dem-onstrated that NMHC-A mRNA and protein are expressed in normal tissue and in primary atherosclerotic and rest-enotic lesions.30
The role that ανβ3 integrins play in human vascular disease remains to be clarified. Although there are com-pelling data that ανβ3 integrins regulate SMC growth responses in some animal models of vascular injury and in cultured SMC,2–9 and that ανβ3 integrin expression is increased in atherosclerotic human coronaries,1 the re-sponse of SMC varies with phenotype. Also, as is well documented, the phenotype of SMC within the atheroscle-rotic human plaque may differ significantly from those studied in animal models or cell culture.35 Thus, the ques-tion as to what role, if any, that ανβ3 integrins play in human vascular disease must await direct study.
In summary, treatment of human SMC with platelet-derived TSP resulted in: 1) association of NMHC-A with ανβ3 integrins; 2) association of NMHC-A with FAK; and 3) increased tyrosine phosphorylation of FAK. NMHC-A was expressed within the media of uninjured baboon bra-chial arteries and expression markedly increased following balloon withdrawal injury. Taken together, these results demonstrate that ανβ3 integrin-dependent protein com-plexes form following treatment of SMC with TSP and suggest that NMHC-A may play a pivotal role in regulat-ing SMC growth responses following vascular injury through linking integrin activation with intracellular sig-naling pathways.
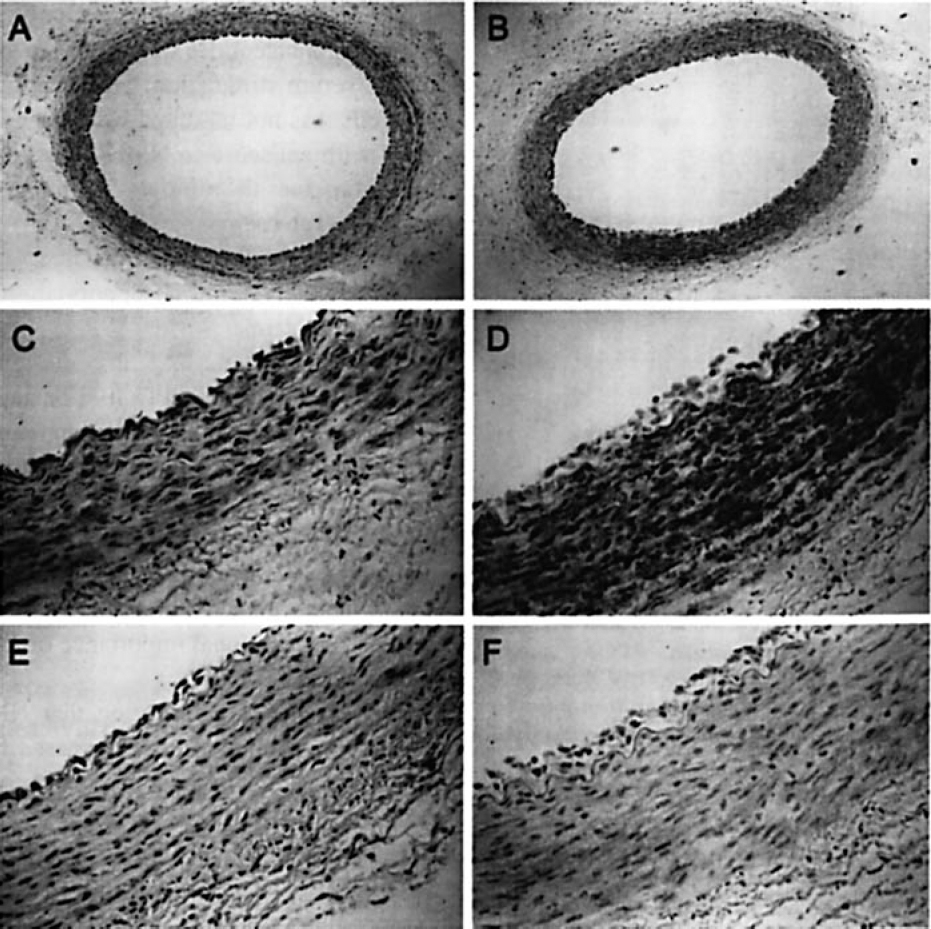
Expression of NMHC-A in injured and uninjured baboon brachial arteries. Left brachial arteries of baboons were injured by balloon withdrawal. Seven days later, left (injured) and right (uninjured) brachial arteries were harvested. NMHC-A immunoreactivity was detected using anti-NMHC-A antibodies raised in rabbits immunized with human platelet myosin (BT-561) and 3,3′-diaminobenzidine substrate. A. Uninjured artery blotted with BT-561 (magnijication × 100); B. injured artery blotted with BT-561 (magnijication × 100); C. uninjured artery blotted with BT-561 (magnijication × 400); D, injured artery blotted with BT-561 (magnijication × 400); E, uninjured artery blotted with IgG from a nonimmunized rabbit (magnijication × 400); F, injured artery blotted with IgG from a nonimmunized rabbit (magnijication × 400).
Footnotes
Acknowledgments
This study was supported in part by American Heart Association Texas Affiliate Grant-in-Aid (96G-631).