
Abstract
Background
Methods
Results
Conclusion
Introduction
11β-Hydroxysteroid dehydrogenase type 1 (11βHSD1) is a bidirectional nicotinamide adenine dinucleotide phosphate (NADP)/reduced NADP (NADPH)-dependent de-hydrogenase/oxoreductase that mainly converts inactive glucocorticoids (cortisone and 11-dehydrocorticosterone) to their active forms (Cortisol and corticosterone). Conversely, 11βHSD type 2 (11βHSD2) is an NAD-depen-dent unidirectional dehydrogenase that exclusively transforms active glucocorticoids to inactive ones. 1 Although the bulk of studies have been conducted in livers and kidneys, there is now evidence that the two isoenzymes are expressed in the human adrenal cortex (HAC), where they may play a role in the regulation of steroid hormone secretion.
The presence of 11βHSD1 has been demonstrated by immunocytochemistry in all three zones of HAC, with the most intense staining occurring in the zona reticularis. 2 The activity of this isoenzyme has not been assayed, but functional defects in 11βHSD1 have been reported to be associated with adrenal hyperplasia3,4, and more recently they have been reported to be associated with the absence of the classic phenotype in a patient with a pituitary-dependent Cushing's syndrome, 5 suggesting involvement in the maintenance of normal glucocorticoid secretion. The expression of 11βHSD2 gene and marked 11βHSD2 activity have been demonstrated in human adrenocortical cells,6,7 and evidence has been found that 11βHSD2 is negatively modulated by the local concentration of non-11β-hydroxylated steroid hormones either in vitro 6 or in vivo. 8
The expression and activity of both 11βHSD isoenzymes also have been studied in adrenal tumors, but far less intensively than in the normal gland. The presence of 11βHSD2 mRNA has been detected in adrenal carcinomas, aldosterone-secreting adenomas9,10 and cortisol-se-creting adenomas (CSAs). 7 In CSAs, however, 11βHSD2 activity was rather low in comparison with normal adrenal tissue, a finding in keeping with the possibility that the high tissular level of steroid hormone intermediates in adenomas may downregulate 11βHSD2. No data are available regarding 11βHSD1 expression and activity in adrenal adenomas. It therefore seemed worthwhile to study the expression and activity of both 11βHSD1 and 11βHSD2 in a series of six CSAs and to compare these parameters with those in the HAC.
Methods
Adrenal Specimens
Six adult patients with unilateral CSAs producing Cushing's syndrome were recruited. They displayed high basal levels of plasma and urinary Cortisol (20-32 μg/dL, and 325-674 μg/d, respectively) as well as low levels of plasma adrenocorticotropic hormone (ACTH; 4-6 pg/ mL). After adrenal mass identification by nuclear magnetic resonance imaging, the patients underwent surgery, and the resected tumors were histologically identified as lipid-rich-cell (presumably zona fasciculata cells), benign adenomas. Normalization of hematourinary hormonal parameters was observed within 10 days after surgery (plasma Cortisol, 8-14 μg/dL; urinary Cortisol, 60-100 μg/d; plasma ACTH, 22-32 pg/mL). Normal HACs were obtained from six adult patients undergoing unilateral nephrectomy-adrenalectomy for kidney cancer, whose recruitment criteria were described previously. 11 Their urinary Cortisol level ranged from 70-110 μg/d, and their plasma levels of Cortisol and ACTH were 10-15 μg/dL and 15-35 pg/mL, respectively. Fragments of adenomas and adrenal tails, which do not contain adrenal medulla, 12 were collected immediately after excision in the operating room, placed in a Krebs-Ringer bicarbonate buffer with 2% glucose at 4°C, and immediately carried to our cell biology laboratory. The local Ethics Committee for Human Studies approved the study protocol.
11βHSD Isoenzyme Expression
Part of each tissue fragment was immediately frozen in liquid nitrogen and stored at — 80°C. Total RNA was extracted using the guanidium isothiocyanate method, and the reverse transcription-polymerase chain reaction (RT-PCR) procedures used were described in an earlier study.
6
The primer sequence and the optimal thermal profile were selected with Primer3 software (Whitehead Institute for Biomedical Research, Cambridge, Mass). In a thermal cycler (PCR Sprint; Hybaid, Ashford, UK) after initial denaturation at 95°C for 10 min, we used a denaturation step at 95°C for 45 sec, an annealing step at 60°C for 45 sec, and an extension step at 72°C for 45 sec, for a total of 38 cycles. An additional extension step at 72°C for 7 min was then performed. Primer sequences and the predicted size of PCR products were as follows:
11βHSD1-161-5', 5'-GCA ACC GAG GAA TTC AGA CC-3', and 11βHSD1-559-3', 5'-CAA GGC AGC TAC AGT CAG GA-3'; 339 bp 11βHSD2-384-5', 5'-TCT GGT TTT GGT AAG GAG AC-3', and 11βHSD2-983-3', 5'-CAG CTC TTG AGG CAG CTT G-3'; 600 bp GAPDH-130-5', CCC TTC ATT GAC CTC AAC TA-3', and GAPDH-695-3', 5'-GCC AGT GAG CTT CCC GTT CA-3'; 585 bp
To rule out the possibility of amplifying genomic DNA, one PCR was performed without prior RT of the RNA. Detection of the PCR amplification products was first performed by size fractionation on 2% agarose gel electrophoresis. Then, after purification using the QIA quick purification kit (Qiagen, Hilden, Germany), PCR products were identified by sequencing on an Alf sequencer (Pharmacia Biotech, Freiburg, Germany).
Further experiments were performed to provide a semiquantitative evaluation of the expression of 11βHSD1 and 11βHSD2 mRNA. Amplification was performed for 22, 30, or 38 PCR cycles, gels were photographed with Polaroid film, and amplicon-band densities were measured with a digital image system (Leitz-Wetzlar, Wetzlar, Germany). The level of expression of the target genes was estimated by the ratio, at each number of PCR cycles, of their amplicon densities to those of the housekeeping gene GAPDH (whose expression was assumed equal to 1).
11βHSD Isoenzyme Activity
Fresh tissue fragments were homogenized at 4°C in 4 v/v 100 mM potassium phosphate buffer (pH 7.4) containing 250 mM sucrose. Microsomes were prepared according to the method of Mackinnon et al., 13 and 11βHSD activity was assayed according to the method of Monder et al., 14 as detailed previously. 9 Briefly, 300 nCi of [ 3 H]cor-tisone or 100 nCi of [ 3 H]cortisol (specific activity, 50-80 Ci/nmol; Amersham Pharmacia Biotech, Milan, Italy) were added to 50 μg of microsomal protein in the presence of 0.25 mM NADPH (11βHSD1 assay) or NAD (11βHSD2 assay), respectively, and 100 mM Tris (pH 8.3). Samples were incubated at 37°C for 60 min, the reaction was stopped, and steroids were extracted with dichloromethane at 4°C. Extracts were evaporated to dryness and redissolved in 20 μL methanol. Cortisol and cortisone were separated and purified by high-performance liquid chromatography, 15 transferred into scintillation vials, and counted with a Wallac MicroBeta liquid scintillation counter (PerkinElmer, Boston, Mass). Results were expressed as cycles per minute of [ 3 H]cortisol or [ 3 H]cortisone formed per mg protein.
Cortisol Secretion
HAC and CSA slices were placed in medium 199 (Difco, Detroit, Mich) and Krebs-Ringer bicarbonate buffer with 0.2% glucose containing 5 mg/mL bovine serum albumin and incubated (8-10 mg/mL, in replicates of three each) in the presence or absence of 10-5 M aminoglutethimide (Elipten Ciba; Origgio, Italy), an inhibitor of cholesterol side-chain-cleaving enzyme. 16 The slices were incubated in a shaking bath at 37°C for 60 min in an atmosphere of 95% O2/5% CO2. Cortisol was extracted and purified, 15 and its concentration was measured by radioimmunoassay using a commercial kit (IRE-Sorin, Vercelli, Italy). The findings were sensitivity, 30 pg/mL; cross-reactivity: Cortisol, 100%; 11-deoxycortisol, 4.8%; cortisone, 3%; progesterone, 0.5%; 11-deoxycorticosterone, 0.02%; other steroids, <0.01%). Intra- and interas-say coefficients of variation were 6.5 and 7.9%, respectively. The protein concentration of the incubated slices was determined by the Lowry method, and Cortisol production was expressed in picomoles per milligram of protein.
[3H]Cortisol or [ 3 H]Cortisone Production
Other HAC and CSA slices were incubated as described above with 30 nCi/mg of [ 3 H]cortisone or 10 nCi of [ 3 H]cortisol in the presence or absence of 10-5 M aminoglutethimide. Cortisol and cortisone were extracted from the incubation media and purified, 15 transferred into scintillation vials, and counted in a Wallac MicroBeta counter. The [ 3 H]cortisol or [ 3 H]cortisone that was formed was expressed in cycles per minute per milligram of protein.
Statistics
Data were expressed as mean ± SEM of six separate experiments (six HACs and six CSAs), and statistical comparison was performed by analysis of variance followed by the multiple range test of Duncan.
Results
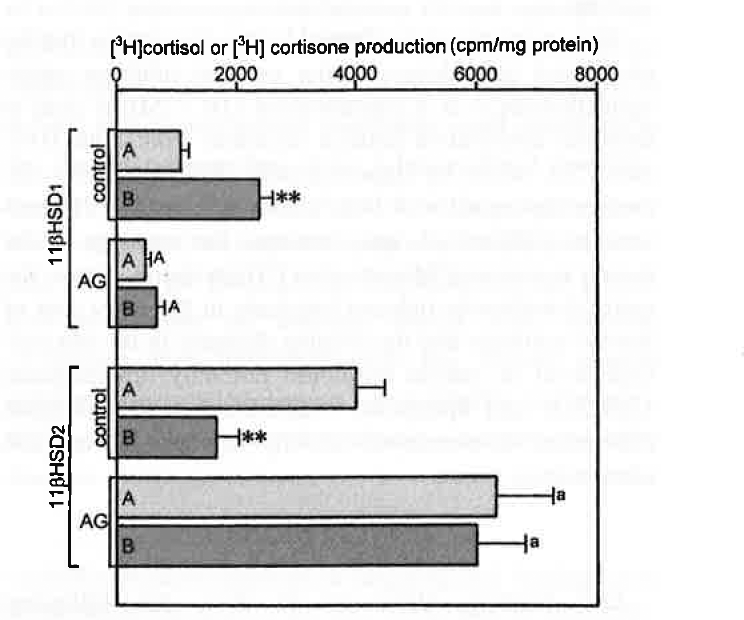
RT-PCR allowed the detection of 11βHSD1 and 11βHSD2 mRNA in all HACs and CSAs (Figure 1). Semiquantitative PCR, however, showed that 11βHSD1 expression was approximately 70% higher and 11βHSD2 expression was approximately 40% lower in adenomas than in normal cortex (Figure 2). The microsomal fraction of both HACs and CSAs displayed sizable 11βHSD1 and 11βHSD2 activity. Basal 11βHSD1 activity was approximately 2.3-fold higher and 11βHSD2 activity was approximately 45% lower in adenomas than in normal cortex (Figure 3). Basal Cortisol secretion from CSA slices was approximately 65% higher than that from HAC slices, and aminoglutethimide (10-1 M) reduced it (by approximately 70%) in both tissues (Figure 4). Under basal conditions, adenoma slices produced higher amounts of [ 3 H] Cortisol and lower amounts of [ 3 H]cortisone than did HAC slices (125 and —66%, respectively). These differences were annulled by aminoglutethimide (10-5 M), which also markedly decreased [ 3 H]eorlisol production and increased [ 3 H]cortisone production by both tissues (Figure 5).
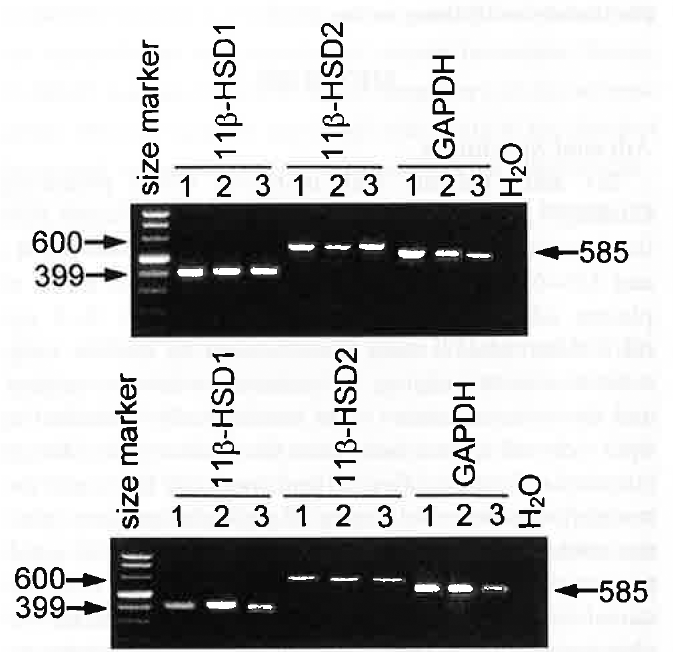
Ethidium bromide-stained 2% agarose gel showing cDNA amplified with human specific 11βHSD1, 11βHSD2 and GAPDH primers from three exemplary HACs (upper panel) and CSAs (lower panel). Thirty-eight PCR cycles were used, as well as additional extension step at 72°C for 7 min; Lanes 1 were loaded with 200 ng of a size marker (Marker VIII; Roche Molecular Biochemicals, Indianapolis, IN). No amplification with water instead of RNA is shown as negative control.
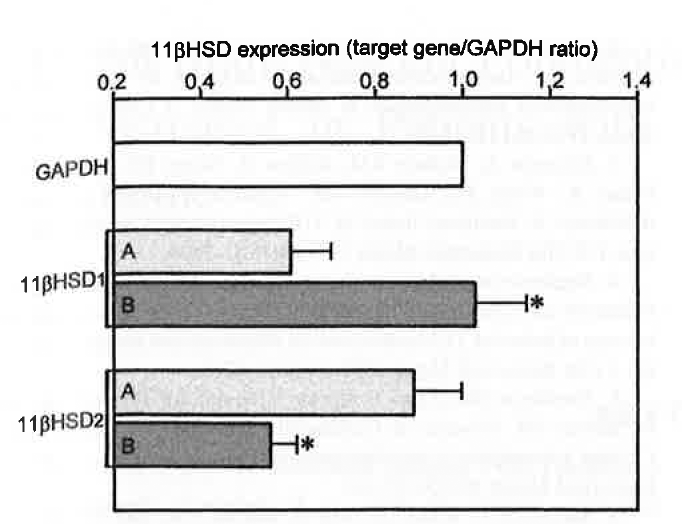
Semiquantitative PCR of 11βHSD1 and 11βHSD2 expression in HACs (A) and CSAs (B). Expression is indicated by the ratio of amplicon density of target genes over that of the housekeeping gene GAPDH. Values are means ± SEM of six separate experiments.
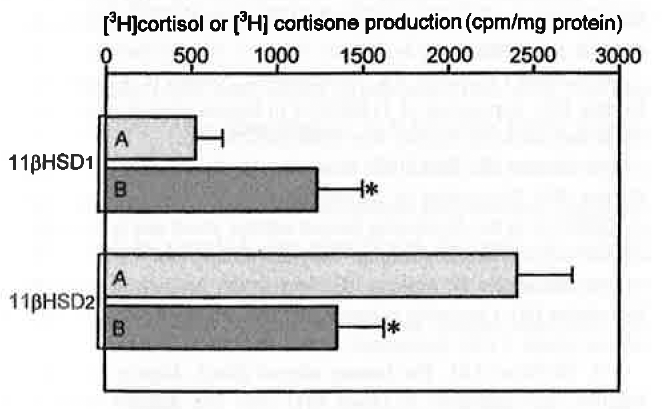
Comparison of basal 11βHSD1 and 11βHSD2 activity of microsomal fraction preparations from HACs (A) and CSAs (B). Values are means ± SEM of six separate experiments.
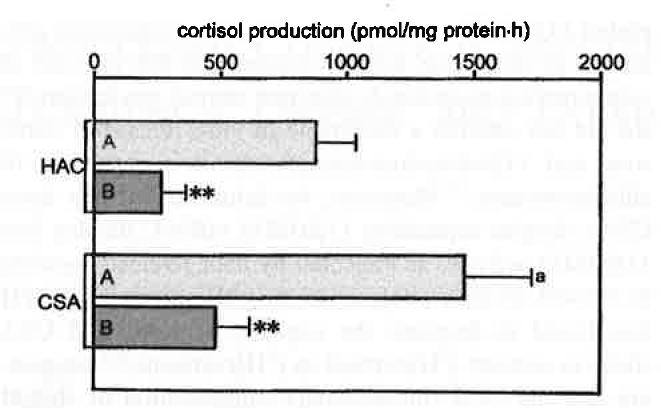
Effect of aminoglutethimide (AG) (10-5 M) on basal Cortisol secretion from HACs and CSAs. A, control; B, AG. Values are means ± SEM of six separate experiments.
Discussion
Our present findings clearly show that HACs and CSAs, in addition to expressing 11βHSD2 gene, also express 11βHSD1 as mRNA and protein, as indicated by the sizable 11βHSD1 activity displayed in vitro by both normal and neoplastic tissue. Moreover, our study provides the first evidence that marked differences in the level of expression of the two 11βHSD isoenzymes occur between normal cortex and adenomas.
Semiquantitative PCR and enzymatic assays indicate that 11βHSD1 mRNA and activity are significantly higher and 11βHSD2 mRNA and activity significantly lower in CSAs than in HACs. Although early investigators interpreted 11βHSD2 expression in aldosterone-secreting adenomas as indicating the involvement of this enzyme in counteracting exceedingly elevated steroid production,9,10 we did not observe a detectable in vitro release of cortisone and 11β-dehydrocorticosterone in a series of 10 aldosteronomas. 17 Moreover, we demonstrated that three CSAs, despite expressing 11βHSD2 mRNA, display low 11βHSD2 activity, as indicated by their reduced capacity to convert in vitro [ 3 H]cortisol to [ 3 H]cortisone. 7 ACTH was found to decrease the capacity of HAC and CSA slices to convert [ 3 H]cortisol to [ 3 H]cortisone,6,7 suggesting that the local (intracellular) concentration of steroid hormones negatively modulates 11βHSD2 activity. Our results indicate that the high tissular level of steroid hormone intermediates in CSAs not only downregulates 11βHSD2 but also upregulates 11βHSD1 expression and activity.
This contention is confirmed by the observation that the cholesterol side-chain-cleaving enzyme inhibitor amin-oglutethimide, 16 at a concentration (10-5 M), is able to decrease constitutive Cortisol secretion from both HAC and CSA slices by approximately 70%, markedly decreases the capacity of both tissues to convert [ 3 H]corti-sone to [ 3 H]cortisol, and increases the capacity of the tissues to convert [ 3 H]cortisol to [ 3 H]cortisone. Hence, the aminoglutethimide-induced blockade in the early step of steroid synthesis and the ensuing decrease in the intracellular level of steroid hormones not only downregulate 11βHSD1 and upregulate 11βHSD2 but also abrogate differences in isoenzyme activity between normal and adenomatous tissue.
Conclusions
Our findings allow us to draw the following conclusions:
11βHSD1 and 11βHSD2 are expressed as mRNAs and proteins in both HACs and CSAs.
11βHSD1 is upregulated and 11βHSD2 is downregu-lated in CSAs.
11βHSD1 and 11βHSD2 activities are positively and negatively correlated with the intracellular concentration of steroid hormones.
11βHSD isoenzymes could act as amplifiers of the secretagogue effect of agonists, and could contribute to the elevated hormonal secretion of CSAs.