
Abstract
Asthma is a complex respiratory disease, which is controlled by genetic and environmental factors. Type 2-dominant immune response is responsible for asthma. Decorin (Dcn) and stem cells have modulatory effect on immune system and may control tissue remodeling and asthma pathophysiology. In this study, immunomodulatory effect of transduced induced pluripotent stem cells (iPSCs) with expression of Dcn gene on allergic asthma pathophysiology was evaluated. After transduction of iPSCs with Dcn gene, allergic asthma mice were treated with iPSCs and transduced iPSCs via intrabronchial. Then, airway hyperresponsiveness (AHR), levels of interleukin (IL)-4, IL-5, IL-13, IL-33, total IgE, leukotrienes (LTs) B4, C4, hydroxyproline (HP) content, and transforming growth factor-beta (TGF-β) were measured. Also, lung histopathology study was done. AHR, levels of IL-4, IL-5, IL-13, IL-33, total IgE, LTs B4, C4, TGF-β, HP content, mucus secretion, goblet cell hyperplasia, and eosinophilic inflammation were controlled by iPSCs and transduced iPSCs treatment. Therapeutic effect of iPSCs could control main allergic asthma symptoms and related pathophysiologic mechanisms and the effect can be increased when applied with Dcn expression gene.
Asthma is a chronic respiratory disease and may lead to lung remodeling.
Transduced induced pluripotent stem cells (iPSCs) with Decorin (Dcn) could control main allergic asthma symptoms and related pathophysiologic mechanisms.
Transduced iPSCs with expression Dcn gene could control asthma pathophysiology and decrease remodeling factors and pulmonary fibrosis.
Introduction
Asthma is one of the main complex disease of the respiratory system, which is characterized by recurrent coughing, dyspnea, wheezing, airway narrowing, increased bronchial responsiveness to stimuli, eosinophilic inflammation, mucus hypersecretion, and smooth muscle spasm.1–3 Asthma is controlled by genetic and environmental factors. Type 2 (Th2)-dominant immune response is responsible for asthma, the main respiratory disease, and asthma leads to remodeling of lung in long time. In allergic asthma, Th2 cytokines lead to further heightening of airways’ inflammation and bronchoconstriction.4–6 Currently, available drugs cannot completely cure asthma and immunomodulation can help to control pathophysiology of asthma.7–9 Decorin (Dcn) and stem cells have modulatory effect on immune system and may have ability to control remodeling and treatment of asthma.
To repair the lung and bronchiole epithelium, the presence of stem cell populations in distinct niches is necessary. Induced pluripotent stem cells (iPSCs) are introduced by Yamanaka factors (octamer-binding transcription factor 4 (OCT4), SRY-box 2 (SOX2), Krüppel-like factor 4 (KLF4), transient transcription factor (cMYC: OSKM)), which display self-renewal properties and pluripotency. These cells can reduce injury, promote repair, and improve lung function. Pluripotent stem cells have plasticity and immunomodulatory effect in tissues with inflammation.10–12
TGF-β is considered as the principal regulatory factor of airway remodeling in asthma and directly influences deposition of collagen-promoting fibrosis formation in the wall of the airway. TGF-β/Smad signal pathway is one of the important mechanisms for signal conduction in airway remodeling. Therefore, it would be very helpful for control of airway remodeling in asthma to block the intracellular TGF-β signal transduction for fibroblast proliferation and collagen synthesis.7,13
The small leucine-rich proteoglycan, Dcn, plays a key role in the development of several tissues and is involved in the formation and stability of collagen fibers. Dcn is an extracellular matrix proteoglycan and affects airway-parenchymal interdependence, airway smooth muscle (ASM) proliferation and apoptosis, mechanics, and TGF-β bioavailability. It was demonstrated that Dcn reversibly binds with the core protein of the TGF-β active form, resulting in the reduction of its bioavailability. Lack of Dcn resulted in abnormal lung collagen deposition, and airway-parenchymal interdependence. Furthermore, Dcn effects on TGF-β bioavailability that may influence remodeling and inflammation indirectly.14–16 Therefore, we evaluated the immunomodulatory effect of transduced iPSCs with expression Dcn gene on allergic asthma pathophysiology and remodeling bio-factors in ovalbumin (OVA) induced allergic asthma lung of mice.
Material and methods
Dcn cells
The production of transduced iPSCs with Dcn gene was carried out according to the previous studies.11,17 Briefly, after preparation of expression vector containing a fully sequenced open reading frame, expression vector of Dcn was transduced to the iPSC. The pAAV-CMV-Dcn plasmid as expression vector contains expressible mouse Dcn gene.
Dcn approval
For Dcn gene expression, after isolation of total RNA from transduced iPSCs and cDNA synthesis, RT-PCR was done with specific primers of Dcn gene.
Mice treatment
The BALB/c mice (N = 36 male mice, 6- to 7-week old) were kept in animal laboratory house under standard conditions (22°C–24°C temperature, 55%–65% humidity, 12 h light/12 h dark cycle). The mice were divided into three groups (n = 12), that include: asthma group (A), asthma group that was treated with iPSCs (AP) intrabronchial on Day 25, and asthma group that was treated with Dcn gene transduced iPSCs (APD) intrabronchial on Day 25. The allergic asthma mouse model was produced according to the previously described method2,5,6 by OVA sensitization and challenging. In Day 30, airway hyperresponsiveness (AHR) by methacholine challenge test was done and on Day 31, samples of blood, bronchoalveolar lavage fluid (BALf), and lung tissue were taken.2,5,6 BALf was collected via lung intubation and from each mice, 0.7 ml BALf was sampled.
AHR measurement
Methacholine (MCh) challenge test was used to determine the Penh value and assess AHR. 7 In each group, six mice were initially exposed to PBS (phosphate buffered saline) aerosol and MCh concentrations (5, 10, 20, 40, and 80 mg/ml).
Cytokines measurement
The levels of IL-4, IL-5, IL-13, and IL-33 in BALf were measured using ELISA method with specific kits.
IgE measurement
Total IgE level was measured in blood of the mice after separation of serum via anti-Mouse IgE ELISA kit.
Leukotrienes measurement
The levels of leukotriene (LT) B4 and LTC4 were measured in BALf of the mice using specific ELISA kits.
Remodeling factors measurement
The hydroxyproline (HP) content, as an important index of the deposition of collagen fibers in the lung tissue, was measured by a colorimetric modified method and also TGF-β was measured in supernatants of lung tissue homogenate described before. 7
Histopathology
Lung tissues of mice were fixed and after preparation of slides, the sections were stained with Alcian Blue (AB), hematoxylin and eosin, Periodic acid-Schiff (PAS) stain, and AB–PAS. Afterwards, mucus hypersecretion, goblet cell hyperplasia, in the airways, peribronchiolar and perivascular inflammation were evaluated.
Statistical analysis
All experimental tests were repeated three times and the results were reported as a mean of three times with standard deviations. The SPSS version 21 was used for statistical analyses and the p value less than 0.05 was supposed to be significant. The analyses were performed using version 5.0 of the GraphPad Prism. Correlation analysis was carried out using Pearson’s method and to analyze data, the paired t-test was applied.
Results
Airway hyperresponsiveness
Increased Penh values at all concentrations of MCh in A group (at 20 mg/ml = 29.0 ± 1.7) were significantly (p < 0.05) decreased in AP and APD groups (at 20 mg/ml = 19.0 ± 1.4 and 8.9 ± 0.7, respectively). Also, Penh values were significantly (p < 0.05) decreased in APD group compared to AP group (Figure 1).
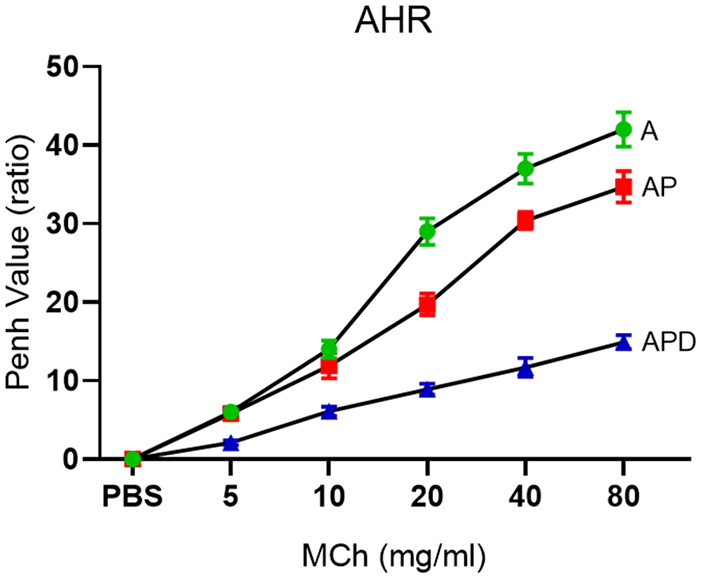
Airway hyperresponsiveness. After the last OVA challenge on Day 30, MCh challenge test was done with five concentrations of MCh for three groups. AHR could be controlled in AP and APD groups.
Cytokines
The levels of IL-4, IL-5, IL-13, and IL-33 in asthma group (114.2 ± 4.8, 101.9 ± 6.7, 158.9 ± 10.4, and 314.5 ± 23.4 pg/ml, respectively) were significantly (p < 0.05) decreased by receiving iPSCs (90.5 ± 4.2, 70.1 ± 2.4, 114.8 ± 7.8, and 210.7 ± 19.8 pg/ml, respectively) and Dcn gene transduced iPSCs (71.9 ± 8.1, 54.9 ± 4.1, 90.2 ± 4.9, and 176.4 ± 27.9 pg/ml, respectively; Figure 2).
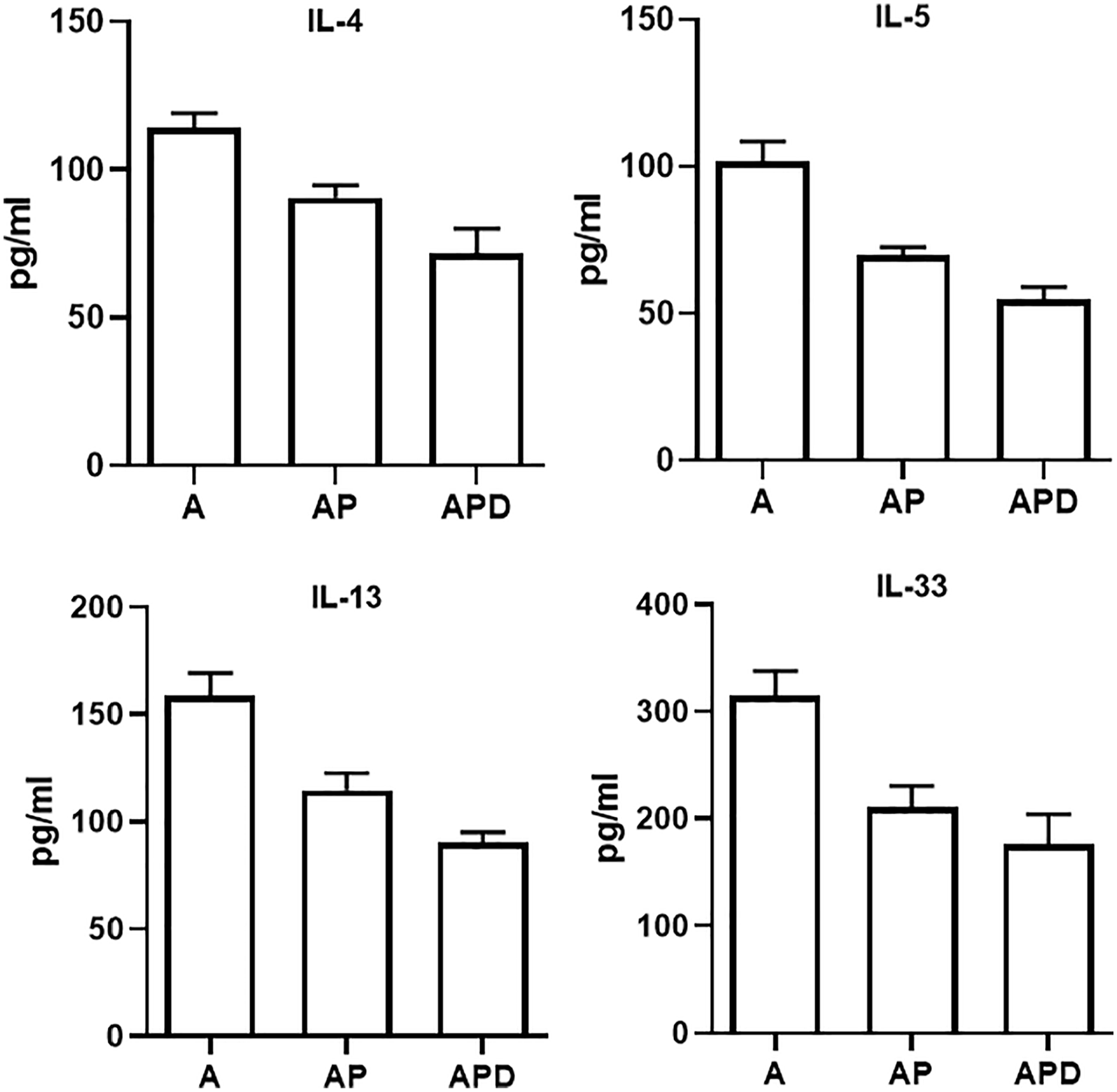
Cytokines levels. BALf samples were taken and used for cytokine measurement. The levels of IL-4, 5, 13, and 33 were measured in BALf of the all groups.
IgE
Total IgE level in serum of asthmatic mice was 3211.9 ± 218.9 ng/ml that significantly (p < 0.05) decreased in AP (1914.5 ± 201.8 ng/ml) and APD (1125.6 ± 194.6 ng/ml) groups. Decreasing in total IgE level in serum of APD was significant (p < 0.05) compared to AP mice.
LTs
The levels of LTB4 and LTC4 were decreased significantly (p < 0.05) in two AP (105.6 ± 8.7 and 202.4 ± 38.4 ng/ml, respectively) and APD (89.4 ± 10.6 and 174.5 ± 30.7 ng/ml, respectively) groups compared to asthma (139.7 ± 16.4 and 297.4 ± 41.3 ng/ml, respectively) group (Figure 3).
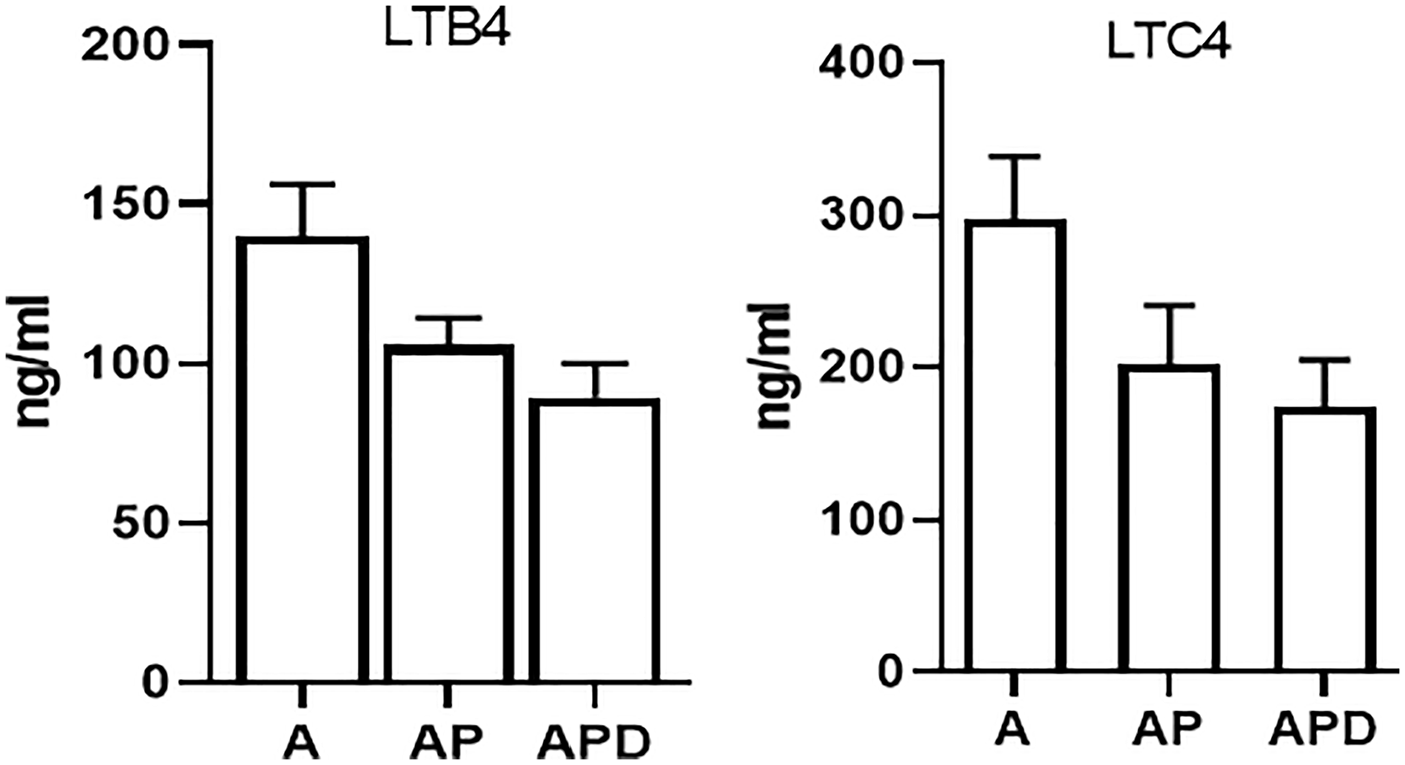
Levels of LTs. The levels of LTB4 and LTC4 were measured and they were found to be decreased in treated groups compared to non-treated asthma group.
Fibrotic factors
TGF-β and HP, as fibrotic and remodeling factors, were decreased significantly (p < 0.05) in AP (189.9 ± 9.2 pg/ml and 2.2 ± 0.1 mg/g, respectively) and APD (65.4 ± 11.5 pg/ml and 0.8 ± 0.2 mg/g, respectively) groups compared to asthma group (206.8 ± 17.4 pg/ml and 2.7 ± 0.2 mg/g, respectively). TGF-β and HP were decreased significantly (p < 0.05) in APD compared to AP group (Figure 4).
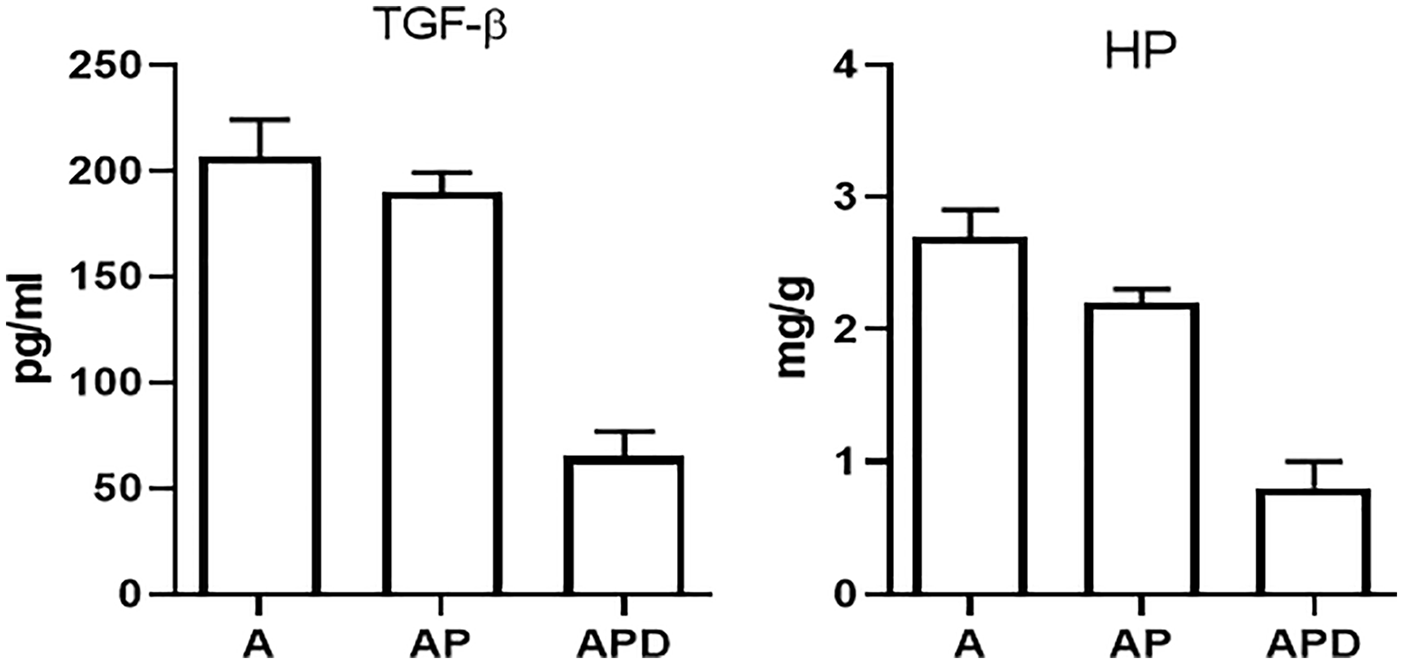
Levels of remodeling factors. The levels of HP and TGF-β as remodeling factors were measured. The levels of HP and TGF-β in AP and APD groups were decreased compared to asthma group. HP and TGF-β decreasing in the APD group was significant in comparison to the AP group.
Histopathology
Mucus hypersecretion, goblet cell hyperplasia, eosinophilic inflammation around bronchi, and vessels induced in the asthma mice (Figure 5). Increased inflammatory cells in the lungs of asthmatic mice were controlled in AP and APD groups. Also, mucus production and goblet cell hyperplasia were controlled in AP and APD compared to A group. Controlling of mucus production, goblet cell hyperplasia, and eosinophilic inflammation were significant in APD group compared to AP group.
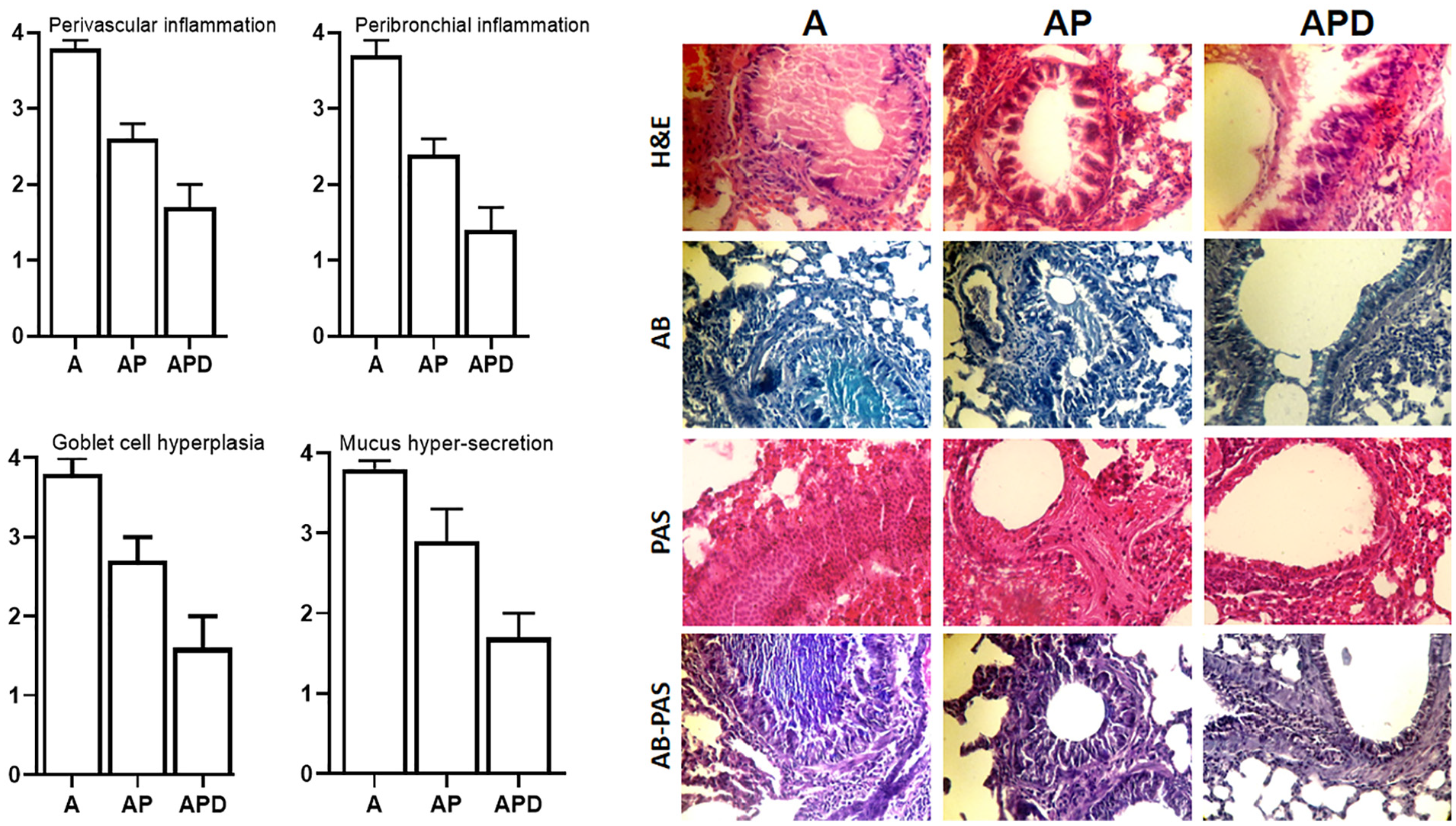
Histopathology. The lung tissues of all groups were stained with H&E, PAS, AB, and AB–PAS. The perivascular inflammation, peribronchial inflammation, the goblet cell hyperplasia, and mucus hypersecretion were evaluated.
Discussion
Dcn absence allergen-induced asthma C57Bl/6 mice model was developed in the study by Marchica et al. 14 They described that Dcn deficiency would lead to enhanced allergen-induced AHR, collagen deposition, and airway remodeling. Lung mechanics are influenced by Dcn and Dcn deficiency affects airway and lung compliance, and also, airway-parenchymal interdependence under baseline conditions, which plays significant role in airway responsiveness. 18 Mucus hypersecretion plays notable role in airway obstruction and increases in respiratory resistance. The consequence of inflammatory exudate within the peribronchial has important implications in the mechanical activity of the airways themselves. ASM cell shortening leads to constriction of the airways, which is critical determinants of airway narrowing.
Dcn effects on cell migration, smooth muscle contractility, immune system responsiveness, and context of allergic asthma.19,20 Dcn modulates several essential biological processes and related bio-factors including collagen fibrillogenesis and has interaction with a wide variety of bioactive molecules. 21 It was demonstrated that collagen bound Dcn is still able to interact with TGF-β and also, Dcn is able to bind to other components of the extracellular matrix, such as fibronectin and thrombospondin. TGF-β in fibrotic diseases leads to the remodeling and fibrosis that can be inhibited by Dcn, and administration of Dcn inhibits accumulation of matrix proteins. Therefore, Dcn was considered as a natural TGF-β antagonist, which can bind quantitatively to the reconstituted collagen fibrils and is retained in the collagen lattice.22–24 Lack of Dcn results in more bioavailable TGF-β, a factor that was demonstrated to function as both an anti-inflammatory and pro-remodeling molecule, depending on the setting. 25 In this study, TGF-β and HP, as main fibrotic and remodeling factors, were reduced in AP and APD groups compared to asthma group. TGF-β and HP were also decreased significantly in APD compared to AP. Therefore, the expression of Dcn could control significantly remodeling factors and can prevent remodeling process and lung fibrosis.
Disproportionate deposition of extracellular matrix in response to trauma and persistent inflammation lead to fibrosis and consequent loss of tissue integrity, intercepting fibrillar regeneration, inelastic scars, and interrupted functional tenogenesis. TGF-β is the key mediator in driving fibrosis through Smad (2 and 3) and mitogen-activated protein kinase (MAPK) pathways.26–28 Mesenchymal progenitors grown in the presence of TGF-β acquired a more fibroblastic phenotype, with increased expression of collagen type I, III, fibronectin, laminin, elastin, and proteoglycans, which lead to pro-fibrotic tissue reactions.29–31 Moreover, fibromodulin gene therapy can represent a more robust control of TGF-β-mediated fibroplasia. Thus, Dcn is an anti-TGF-β efficacy and can modulate fibrotic reactions.
Dcn is an effective moiety for reducing TGF-β bioavailability as it neutralizes and effectively impedes the TGF-β RI/II activation followed by signaling via Smad2, 3, and the extracellular-signal regulated kinase (Erk) proteins, which also diminishes the inflammatory cytokine signaling by the MAPK modulation and nuclear factor kappa light chain enhancer of activated B cells activity. Moreover, Dcn binds to the endogenous ligands of toll-like receptor 2/4 (TLR2/4) to repress the inflammation.32–34 Dcn downregulates TGF-β and enhances phosphorylation of glycogen synthase kinase 3 beta. Binding of Dcn molecule to the TLR2 and TLR4 leads to escalated generation of the pro-inflammatory protein programmed cell death 4 and hence, simultaneously fostering decrement in the anti-inflammatory cytokine (IL-10) synthesis.35,36 Dcn decreases the ErbB2 and ErbB4 receptors’ activity via degradation and in addition, negatively regulates the hepatocyte growth factor receptor Met, vascular endothelial growth factor receptor 2, and platelet-derived growth factor receptor.37–40 The levels of IL-4, IL-5, IL-13, and IL-33 as Type 2 allergic cytokines in asthma group were decreased by receiving iPSCs and Dcn gene transduced iPSCs. IL-33 is upper hand of main allergic asthma cytokines (IL-4, IL-5, and IL-13) and has strong effect in related signaling pathways of these cytokines modulation. Dcn, via effect on the IL-33, can have effect on signaling pathways of cytokines network. Also, with control of IL-4, can control IgE level as main allergic immunoglobulin. We observed that total IgE level in serum of asthmatic mice was decreased in AP and APD groups and this decreasing significant was in APD compared to AP mice.
A study in 2010 showed that Dcn transfection inhibits TGF-β-driven elevated expression of profibrogenic genes, myofibroblasts formation, and fibrosis. Dcn downregulates the expression of the growth factors, such as PGDF-β, TGF-β, extracellular matrix components such as tenascin, fibronectin, and collagens (type I and IV). 41 Dcn as an extracellular matrix is extensively present in the body. Principally, Dcn was identified as a collagen-binding partner and key biomechanical factor regulators of tissue architecture in tissues such as tendon, muscle, skin, lungs, and cornea. The Dcn involvement is not restricted to matrix structural proteins and also it affects a diverse variety of biological functions such as cell growth, proliferation, differentiation, migration, and adhesion. In addition, it can modulate inflammation and fibrillogenesis process.42–44 Mucus hypersecretion, goblet cell hyperplasia, and eosinophilic inflammation around bronchi and vessels were induced in the asthma and in our study, in histopathological sections of lung, increased inflammatory cells in the lungs of asthmatic mice, mucus production, and goblet cell hyperplasia were controlled in AP and APD compared to A group. Controlling of mucus production, goblet cell hyperplasia, and eosinophilic inflammation were significant in APD group compared to AP group and these results showed that Dcn could control notably inflammation, mucus secretion, goblet cell hyperplasia and therefore, Dcn is a strong anti-asthma therapeutic agent.
Dcn interacts with collagen and modulates collagen fibril organization, cross-linking, and degradation. Dcn gene transfer attenuate fibrosis in many non-ocular tissues such as heart and lungs. Dcn modulates signaling of EGFRs and insulin-like growth factor-I and its receptor and binds to three isoforms of TGF-β with equal affinity.45–47 In addition, Dcn inhibits angiogenesis in tumor xenograft and can inhibit vascular endothelial growth factor (VEGF)-induced corneal angiogenesis. VEGF is an initiator and promoter of angiogenesis, and induces robust neovascularization by stimulating new blood vessels growth. Furthermore, Dcn inhibits angiogenesis by suppressing VEGF (vascular endothelial growth factor) and FGF (fibroblast growth factor) expression.48–50 In our evaluation, Penh values in AHR and also the levels of LTB4, LTC4 were decreased significantly in two AP and APD groups compared to asthma group. Penh values were significantly decreased in APD group compared to AP group.
Retinoic acid dependently increases the Dcn mRNA concentration. 51 Dcn is a natural product and there is no any immune response to it. 52 Giri et al. 53 presented that bleomycin-induced lung toxicity is usually accompanied by an increased lipid peroxidation (measured by malondialdehyde content), collagen (measured by HP content), increased pulmonary vascular permeability (measured by the degree of leakage of plasma proteins in the alveoli), an enzyme responsible for post-translational modification of collagen before its maturation, and prolyl hydroxylase activity that leads to pulmonary fibrosis and can be controlled by Dcn.
Thus, Dcn could be used as a therapeutic modality agent to control of remodeling 54 that makes Dcn gene therapy as an attractive mode of intervention for numerous forms of lung inflammation. The potential of Dcn gene therapy to inhibit TGF-β-driven fibrosis has been demonstrated in inflamed tissues that can be used in clinics but deserves further investigation. Dcn could be a potent therapeutic agent against pulmonary fibrosis and systemic inflammation. Aerosol-based delivery of Dcn specifically to the lungs could provide better effect and Dcn-based formulation can achieve 100% bioavailability. Also, Dcn could be strong anti-asthma treatment and via therapeutic effect of Dcn, main allergic asthma symptoms and related pathophysiologic mechanisms can be harnessed.
IL-33 belongs to IL-1 cytokine family that plays important roles in Type 2 allergic immunity via activation of eosinophils, basophils, mast cells, macrophages, and Th2. Its gene and receptors have been identified as susceptibility loci in asthma. IL-33 is a Th2-oriented cytokine that enhances of the Th2 cytokines production, particularly IL-5 and IL-13. In addition, IL-33 is also a chemoattractant for Th2 cells and cells mobilization. IL-33 significantly promotes eosinophil survival.55,56 IL-33 goes on to perpetuate the Type 2 inflammatory lung profile through activating resident immune populations’ cells.57,58 IL-33 acts as upper hand of Type 2 cytokines and can orchestrate allergic-reaction-related cytokines. It has an important role in pathophysiology of allergy and allergic asthma. Therefore, controlling of IL-33 by treatment can be effective on control of asthma and it was observed in this study by Dcn/iPSC therapy.
We had some limitations in this study: the effect of iPSC and iPSC–Dcn therapy in different times on the allergic factors was not studied; and also, changes in gene expression via treatment were not studied. We were unable to study the effect of iPSC and iPSC–Dcn on chronic asthma and remodeling factors.
Footnotes
Authors’ contributions
RZ, ZY, JP, CZ, SSA, and LD contributed to data gathering, writing and scientific revising the manuscript, designing, laboratory study, and data analysis. All the authors reviewed and confirmed the final manuscript before submission. LD contributed to the correspondence, and revising of the final manuscript before submission.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Ethics approval and consent to participate
All methods and animal model studies were approved by the ethic committee of animal house of ix.med.vet.dep, 2022 (No. IX.MED.VET.DEP.REC.2022.4600011.4).
Availability of data and materials
Data are available in request from corresponding author.