
Abstract
Cyclic vomiting syndrome (CVS) is an underdiagnosed disorder of the gut–brain interaction. Our understanding of the pathophysiology of CVS is evolving. Here, we tested the hypotheses that: (1) the levels of endocannabinoids and related lipids are altered in CVS, and (2) cephalic–vagal stimulation drive changes in endolipid levels. Ten adult patients with CVS and eight healthy controls were included. Indirect measurements of parasympathetic (RFa) functions were performed with spectral analysis of heart rate variability and respiratory activity. Plasma levels of endocannabinoids and related lipids were measured at baseline and during a sham feeding. Values are reported as mean ± standard error of the mean and compared using t-test or ANOVA. CVS patients had a lower parasympathetic tone and response to the Valsalva maneuver and deep breathing than the controls. The baseline 2-Arachidonoylglycerol (2-AG) had a significantly higher concentration in CVS (5.9e-008 ± 3.7e-008 mol/L) than control (3.7e-008 ± 1.3e-008 mol/; p < 0.05). Sham feeding did not change the concentration of 2-AG. 2-oleoylglycerol (2-OG) was significantly higher in CVS than control and did not change with sham feeding. Levels of N-acylethanolamines, including anandamide (AEA), were not different in CVS vs control. After sham feeding, AEA showed a trend toward increasing (p = 0.08) in CVS, but not in control. With sham feeding, palmitoyl ethanolamine significantly increased in both CVS and control groups; oleoyl ethanolamine in CVS only, and stearoyl ethanolamine in the control group. Levels of endocannabinoids and related lipids are altered in CVS patients. Sham feeding affects endogenous signaling lipids in a disease and time-dependent manner.
Keywords
Our understanding of the pathophysiology of cyclic vomiting syndrome (CVS) is evolving. Studies on the roles of endocannabinoids in CVS are limited.
Levels of endocannabinoids and related lipids are altered in CVS patients. Sham feeding affects endogenous signaling lipids in a disease and time-dependent manner.
This study shows endocannabinoids have some potential roles in the pathophysiology of CVS. Targeting the endocannabinoid system may be therapeutic in CVS.
Introduction
Cyclic vomiting syndrome (CVS) is a disorder of gut–brain interaction (DGBI) characterized by stereotypical episodes of debilitating nausea and vomiting associated with abdominal pain, followed by asymptomatic periods.1,2 CVS often necessitate emergency department visits or hospitalizations and it has a worldwide prevalence of 1.2%.1,3 In the United States, the reported prevalence of CVS is 2%, in Canada 0.7%, and in the United Kingdom 1%. CVS patients suffer from significant health impairment, which is associated with a high rate of healthcare utilization. 4 A sub-category of DGBI that mimics features of CVS is cannabinoid hyperemesis syndrome (CHS). A history of regular cannabis use is essential in diagnosing this subtype.5,6 The role of cannabis and its derivatives in the development of CHS suggests the importance of the endocannabinoid (eCB) system in the pathophysiology of CHS and drives the hypothesis that the eCB system is involved in CVS in general.
Autonomic nervous system signaling is dysregulated in patients with CVS.7,8 In a cohort of 21 subjects with CVS, autonomic nerve dysfunction was observed in 43%. 7 An additional study with 20 CVS subjects showed that 90% had some impairment of the sympathetic nervous system with postural tachycardia and/or sudomotor dysfunction. 8
The eCB system is a modulator of physiological functions not only in the central nervous system (CNS) but also in the autonomic nervous system, which is dysregulated in CVS.9,10 Broadly, the canonical eCB system consists of G protein-coupled receptors (GPCRs) cannabinoid-1 (CB1) and cannabinoid-2 (CB2), and eCB ligands anandamide (AEA), and 2-arachidonoyl glycerol (2-AG). There are additional receptors and channels as well as endogenous lipids that are associated with the eCB system; however, a key overlap of these are the biosynthetic and metabolic enzymes for the eCB ligands.11,12
These enzymes are also involved in the processing of a wide range of endogenous lipids with cannabimimetic-like properties; however, there are limited available data on their physiological roles in the development and progression of human disorders, including DGBI. 13
Given the importance of the eCB system in the pathophysiology of CVS and their role as a modulator of the autonomic nervous system, which is dysregulated in CVS, we tested the hypothesis that CVS is associated with changes in circulating eCB and related lipid signaling molecules, and these changes are potentiated by the cephalic–vagal stimulation during sham feeding.
Methods and material
Patient recruitment and study design: This case–control study was approved by IRB at Texas Tech University Health Sciences Center El Paso (E14068). Ten adult (18–65 years old, 5 males) CVS patients diagnosed based on Rome IV criteria, and 8 adult (3 males) healthy controls were included. According to Rome IV, CVS patients present with acute stereotypical episodes of vomiting (less than 1 week), with three or more discrete episodes in the prior year and two episodes in the past 6 months, occurring at least 1 week apart. 14 All of the CVS cases were in the remission phase with no CVS attack for the past 3 months.
Patients with documented organic or intestinal pseudo-obstruction, primary eating or swallowing disorders, rumination syndrome, psychogenic vomiting, gastroparesis, systemic sclerosis, cancer, peritoneal dialysis, pregnancy, breastfeeding, history of thoracic or abdominal surgeries that could potentially damage the vagus nerve, any abdominal or thoracic surgery within the past 3 months, Parkinson’s disease, amyloidosis, multiple sclerosis, chemical dependency, recreational or medical cannabinoid use, and significant malnutrition or low body weight were excluded from the study. Any anti-emetic, antidepressant, anxiolytic, or narcotic medication was asked to be stopped 2 weeks prior to participation in the study.
Autonomic nervous system (ANS) testing: On the test day, patients arrived at 8:00 a.m. after 12 h fast. Autonomic function testing was performed at baseline using the ANX 3.0 autonomic monitoring system (ANX3.0; ANSAR Medical Technologies INC., Philadelphia, PA, USA), which measures both branches of the cardiovagal ANS using simultaneous spectral analysis of heart rate variability and respiratory activity during a 15-min examination. 15 As described previously, 15 measurements were performed at rest and following challenges to the sympathetic and parasympathetic system: 5 min of rest, 1 min of deep breathing (parasympathetic challenge), 1 min of short Valsalva maneuvers (sympathetic challenge), followed by a rapid stand and 5 min of standing quietly (sympathetic and parasympathetic challenge). Each challenge was separated by 1 min period of return to baseline. Computed autonomic parameters included parasympathetic activity Respiratory frequency area (RFa) during different maneuvers. Immediately after the ANS testing and on the same day, the circulating eCB and related lipid were measured before and after cephalic–vagal stimulation as described below.
Cephalic–vagal stimulation: The goal of this study was to measure endocannabinoids and related lipids during the cephalic–vagal stimulation. As described previously, the vagal nerve modulation of digestion includes both cephalic and gastric phases. The cephalic phase of digestion is the CNS-driven increase in gastric/pancreatic activity through the sensory stimulation by food. The gastric phase is activated by food in the stomach. By chewing, the vagus nerve is centrally stimulated. However, by spitting the gastric phase is bypassed.16,17 A modified sham food, which consisted of a standardized turkey sandwich, was used for the vagal stimulation. An intravenous line was placed into an antecubital vein, and the plasma samples were collected and immediately frozen on dry ice at baseline immediately prior to receiving the sandwich (0 min), as well as at 10 min, and 30 min while chewing and spitting a standardized turkey sandwich.
Lipid extraction and HPLC/MS/MS analysis: As described previously, 13 plasma samples were aliquoted into 100 μL volumes and then added to 2 mL of (HPLC)-grade methanol (Thermo Fisher Scientific). Solutions were spiked with 500 pmol deuterium-labeled N-arachidonoyl ethanolamine (d8AEA; Cayman Chemical; Ann Arbor, Michigan, USA) to determine extraction efficiency and centrifuged at 19,000 × g for 20 min at 24°C. Supernatants were diluted with HPLC water (purified in-house) to make a 15% supernatant solution. Lipid extractions were performed using C18 solid-phase extraction columns (Agilent). Briefly, columns were conditioned with 5 mL HPLC methanol followed by 2.5 mL HPLC water. Then, the supernatant/water solution was loaded onto the column. Impurities were washed off with 2.5 mL HPLC water. A series of five elutions with 1.5 mL 60%, 75%, and 100% methanol were collected.
Extracts were analyzed using an Applied Biosystems API 3000 triple quadrupole mass spectrometer (Foster City, CA, USA). About 20 μL from each elution were chromatographed using a 2.1 mm × 50 mm XDB-C18 reversed-phase HPLC analytical column with a 3.5-micron particle size (Agilent) using optimized mobile phase ingredients (mobile phase A: 80% water, 20% methanol with 1 mmol/L ammonium acetate; mobile phase B: 100% methanol with 1 mmol/L ammonium acetate). Two Shimadzu 10ADvp pumps provided pressure for gradient elution. Analysis of the HPLC/MS/MS data was performed using Analyst software (Applied Biosystems). Chromatograms displaying the retention time of analytes matching programmed parent and fragment ion masses were generated by running each sample using a multiple reaction monitoring method. Retention times were then compared with those from standards for the suspected compound. If retention times matched, then concentrations were determined by calculating the area under the curve for the unknown and comparing it to the calibration curve obtained from the standards. Extraction efficiency was calculated using the recovery vial spiked with 500 pmol d8NAGly as a standard, and analyte levels were adjusted for extraction efficiency.
Data analysis: Data were analyzed using GraphPad Prism 6.1 software (GraphPad Software, Inc; Boston, MA, USA) and presented as mean ± standard error of the mean (SEM). Normality was tested based on the D’Agostino-Pearson omnibus test. Values are reported as mean ± SEM and compared using t-test or ANOVA (LSD post hoc).
Results
Ten adult (5 males) CVS patients and 8 adult (3 males) healthy controls were included in this study. The mean age of the patients (47.3 ± 4.9 years old) and healthy controls (42.1 ± 3.5 years old) were similar.
Figure 1 illustrates data from the autonomic nervous system analysis showing that CVS patients had a significantly lower parasympathetic tone during rest compared with healthy controls. Parasympathetic response after deep breathing and the Valsalva maneuver were significantly lower in CVS vs. healthy controls (Figure 1).
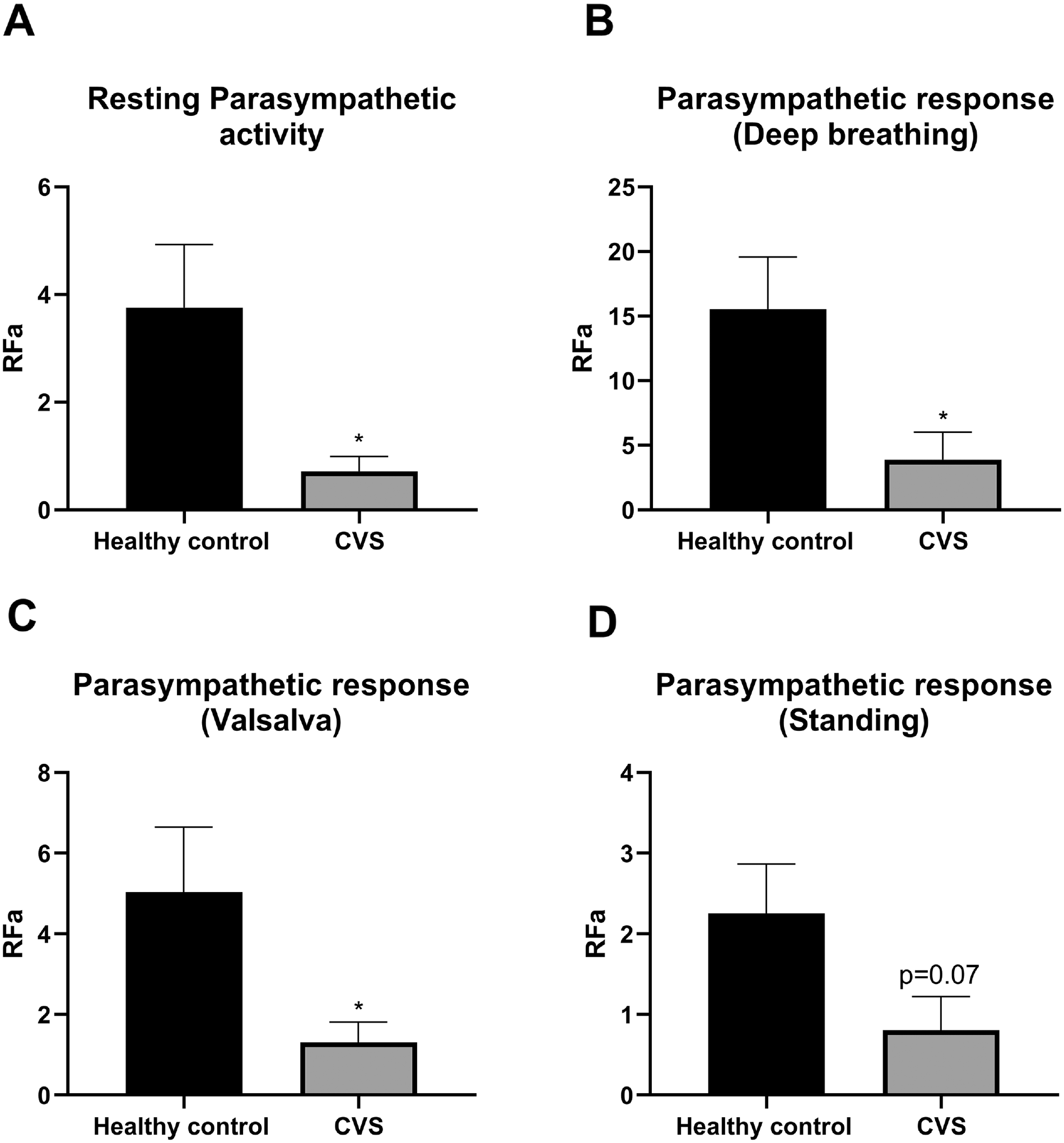
Parasympathetic tone and activity in cyclic vomiting syndrome (CVS) subjects versus controls. Values are mean ± SEM (ANOVA; LSD post hoc). p-values compare CVS and control (*p < 0.05).
The eCB, 2-AG, and 2-acyl glycerols, 2-LG, and 2-OG had higher overall concentrations in CVS patients compared with healthy controls, with no effect of sham feeding. 2-palmitoyl glycerol (2-PG) had similar overall concentrations in CVS and healthy controls (Figure 2).
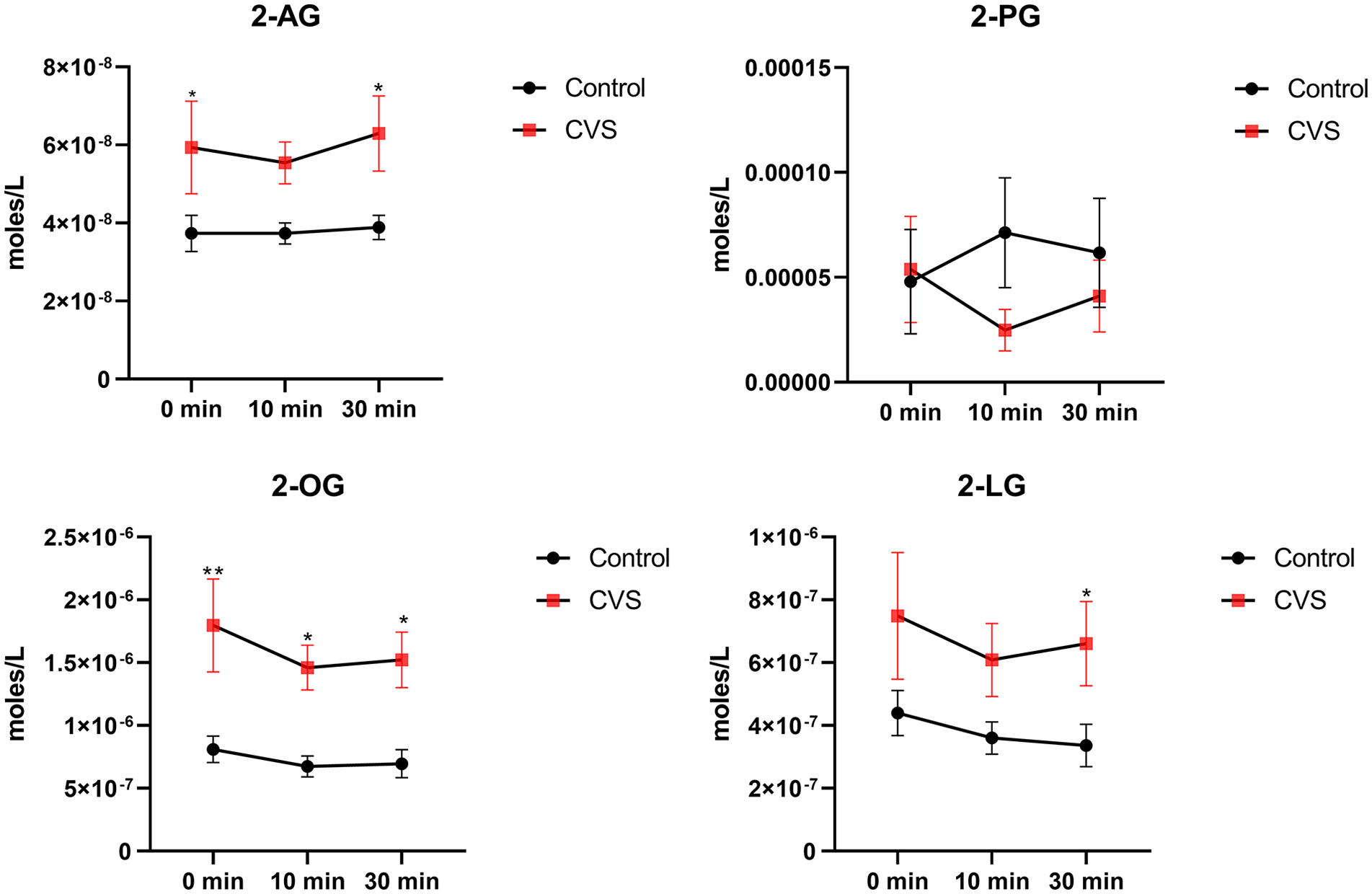
Plasma 2-acyl glycerols in cyclic vomiting syndrome (CVS) vs healthy controls.
Overall, levels of N-acylethanolamines (NAEs) including the endocannabinoid AEA were not different in CVS vs. healthy controls. However, after sham feeding, AEA showed a trend toward increasing in CVS, but not in the healthy control group. With sham feeding, the levels of palmitoylethanolamide (PEA) significantly increased in both CVS and healthy controls. Oleoylethanolamide (OEA) and docosahexaenoyl ethanolamine (DHEA) significantly increased in CVS only, and stearoylethanolamide (SEA) increased only in the healthy control group (Figure 3).
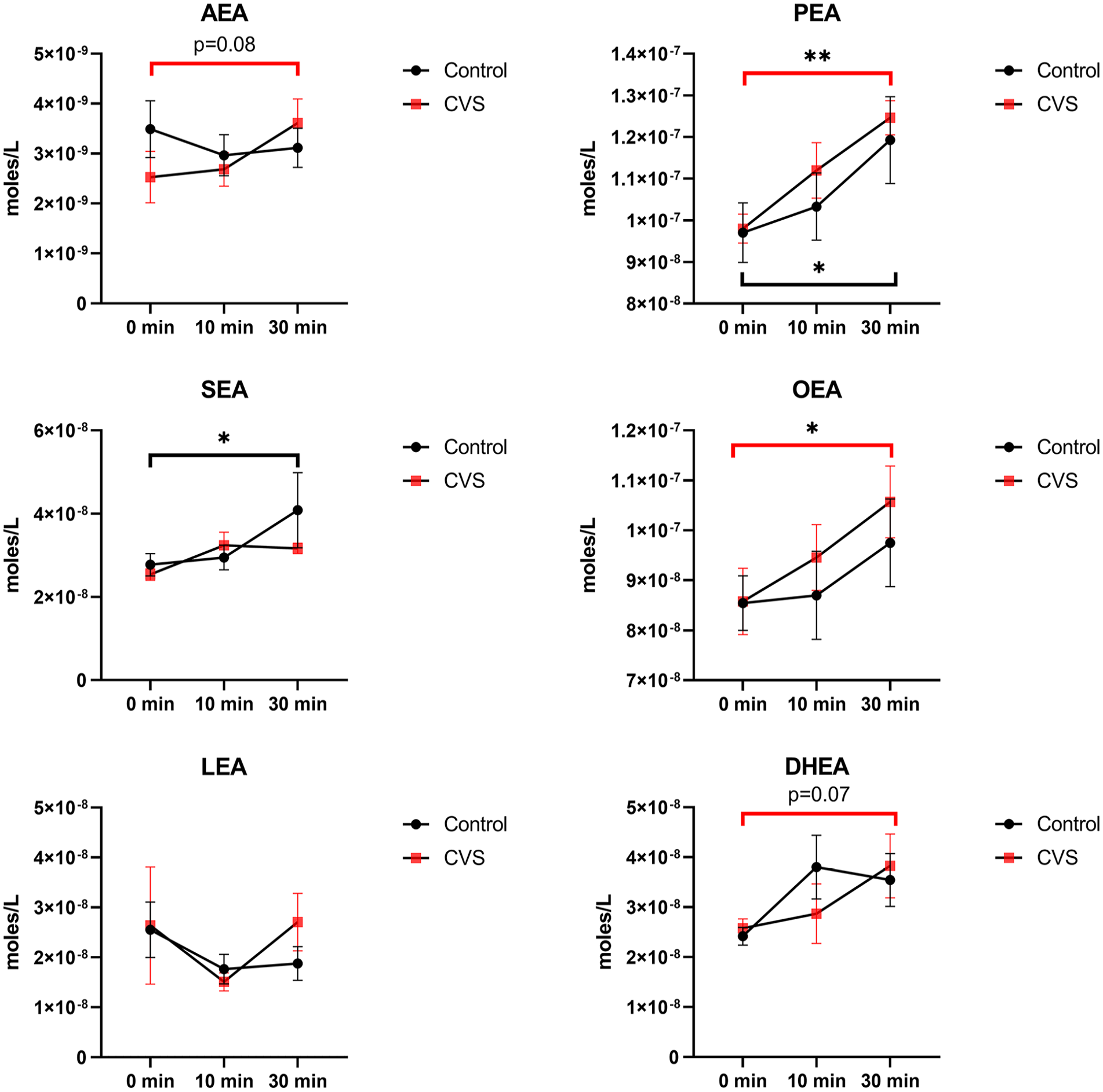
N-acylethanolamines (NAEs) in the plasma of cyclic vomiting syndrome (CVS) subjects vs. healthy controls. Values are mean ± SEM (ANOVA; LSD post hoc). p-values compare changes in NAEs by time in each study group (red line for CVS and black line for control; *p < 0.05, **p < 0.01). The levels of NAEs were not statistically different in CVS compared with controls.
The ω-6 linoleic acid (LA) had a significantly higher concentration in healthy controls. Arachidonic acid and oleic acid had a similar concentration in both groups. The ω-3 eicosapentaenoic acid (EPA) tended to increase in the CVS group only after sham feeding (Figure 4).
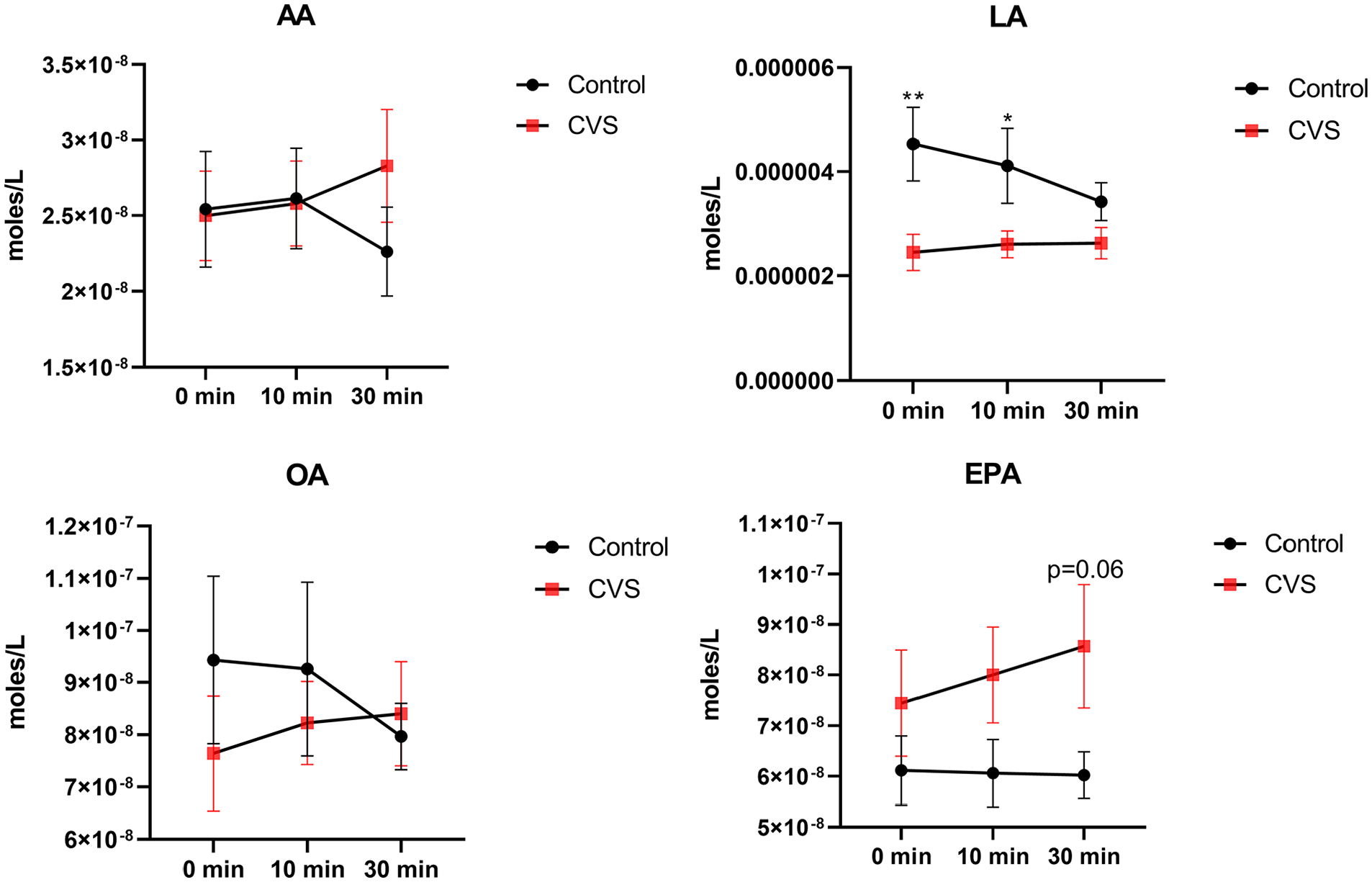
Free fatty acids in the plasma of cyclic vomiting syndrome (CVS) subjects vs. healthy controls. Values are mean ± SEM (ANOVA; LSD post hoc). p-values compare CVS and control at each time point (*p < 0.05; **p < 0.01).
Discussion
CVS is a complex functional gastrointestinal disorder.1,14 Regular cannabis use may lead to CHS, which mimics the clinical features of CVS. 5 Whether the endocannabinoid system is imbalanced in CVS has not been sufficiently explored.
The current study showed that levels of 2-acyl glycerols are elevated in CVS patients and do not change after cephalic–vagal stimulation through modified sham feeding. Levels of NAEs are not different in CVS and healthy controls. However, some of them do change after modified sham feeding in a disease-dependent fashion. Levels of the free fatty acids (LA and EPA) show both overall differences between CVS and healthy controls.
A previous study on 22 CVS patients in the well and sick phases and 12 matched controls showed that serum concentrations of the endocannabinoids AEA and 2-AG were not different among the study groups. However, serum concentrations of OEA and PEA were significantly higher during the sick than the well phase in CVS patients. 18 The actual data show that the average levels are almost double for 2-AG with a high level of variability for this study, 18 suggesting different study powers or extraction/analysis differences compared with our study.
Changes in the endocannabinoid system in CVS have been also reflected in genetic variations. The role of cannabinoid receptor (CNR) 1 gene polymorphisms has been reported in a case–control study on 65 patients with CVS and 1092 healthy controls, with an increased risk of CVS among individuals with AG and GG genotypes of CNR1 rs806380, and a decreased risk of CVS in subjects with CC genotype of CNR1 rs806368. CT and CC genotypes of CNR1 rs806368 were associated with a family history of migraines, and CT and CC of CNR1 rs2023239 with a positive response to treatment. 19
The importance of the endocannabinoid system in CVS is more potentiated when considering some CVS patients who use cannabis to alleviate their symptoms. In the setting of endocannabinoid imbalance, exogenous cannabis and its derivatives may modulate the endocannabinoid signaling system. On the other hand, cannabis use was more common (53%) in CVS patients who were not responding to medications compared with medication responders (22%). 20 Among 140 patients with CVS, with only one patient with diagnosed with CHS, though 41% were current cannabis users, and 21% were regular cannabis users. Most of the cannabis users reported that cannabis helps with their CVS symptoms. 21
The overall increase in all of the 2-acyl glycerols in CVS patient plasma observed in this study is likely because these fatty acids are synthesized and degraded through shared pathways. Diacylglycerol lipase (DAGL) is the primary biosynthetic enzyme of 2-acyl glycerols from the cell-membrane lipids, and monoacylglycerol lipase (MAGL) catalyzes their hydrolysis. 22 Therefore, changes in enzyme activities like DAGL and MAGL during the disease may be what alter the levels of 2-acyl glycerols in the same direction. In other words, enhanced DAGL activity and/or inhibition of MAGL could cause an increase in 2-acyl glycerols levels. This hypothesis needs further investigation.
The effects of MAGL inhibition and increased 2-AG in controlling nausea have been shown in animal models.23–25 In line with these studies, increased 2-AG in CVS should protect against nausea and vomiting, and it is important to state that the CVS patients in this study were in the remission phase and were asymptomatic; therefore, this elevation may be part of what is driving this remission. In a recent study, we showed that plasma 2-AG, 2-PG, 2-OG, and 2-LG levels were significantly lower in women with diabetic gastroparesis compared with controls. 13 Gastroparetic patients suffer from sustained nausea throughout the day. This might be due to decreased endocannabinoid levels in these patients. increased 2-AG levels in CVS patients in the remission phase may have a protective role against the development of CVS vomiting attacks. What happens during the attack phase needs to be further investigated.
Cannabis derivatives, mainly tetrahydrocannabinol, have anti-nausea/-emetic effects. This indirectly shows the importance of endogenous cannabinoids and cannabimimetic compounds, which target similar receptors in the body including CB1. On the other hand, human data on the anti-nausea/-emetic effects of endogenous cannabinoids are limited. In animal models, blocking the degradation of NAEs with a fatty acid amide hydrolase (FAAH) inhibitor reduced acute nausea-mediated conditioned gaping through a peroxisome proliferator-activated receptor alpha receptor antagonist, but not a CB1 receptor antagonist, 26 suggesting these therapeutic effects are potentially through increases in OEA and PEA levels. In other animal studies, inhibiting degradation of NAEs reduced contextually elicited conditioned gaping; unlike acute nausea, the anti-nausea effect of FAAH inhibition was reversed by a CB1 receptor antagonist, 27 likely through increased AEA. The regulatory roles of endocannabinoids on nausea and vomiting pathways are extensively reviewed elsewhere.28–30
In the current study, the levels of NAEs including endocannabinoid AEA were not different in CVS and healthy controls; however, cephalic–vagal stimulation increased the levels of most NAEs in CVS and/or healthy controls. Changes in plasma eCBs have been demonstrated in a previous study of sham feeding in humans. 31 Using a cross-over study design, healthy volunteers had three puddings that were rated as “sweet,”“neutral,” or “bitter” that subjects would palpate and expectorate in a similar design as the current study. Overall levels of 2-AG increased during the 20 min of monitoring only with the “sweet” pudding and decreased with the “bitter” pudding. NAEs had some modest increases with the “sweet” pudding as specific time points, but overall effects were not significant. What is important to note between the studies is that eCBs and related lipids show dynamic changes with sham feeding, which is independent of food nutrient value. An important question that remains unanswered is the origin of the plasma lipids given that they should not be the direct result of food digestion. In both the current and the Mennella et al. 31 studies, the effects are hypothesized to be largely driven by the gut–brain interactions for anticipated digestion, that is, at least in part, by vagal activity.
In our study, CVS patients had a lower parasympathetic tone and activity compared with healthy controls based on ANS testing. Whether disrupted vagal activity in these patients is directly linked with changes in baseline plasma lipid levels as well as after cephalic–vagal stimulation needs to be investigated.
This current study was limited; in that, it originated from a single-healthcare center and had a relatively small sample size, though with enough statistical power to determine overall results. The sample size did limit the ability to analyze the data as a function of genetics, sex, or gender.
In conclusion, the current study shows that plasma levels of endocannabinoids and related lipid molecules are significantly modified in CVS patients. This modulation implies likely changes in their rates of biosynthesis and degradation. Endocannabinoid signaling is affected by cephalic–vagal stimulation in both CVS and healthy controls, and parasympathetic activity is distorted in CVS subjects. Whether targeting endocannabinoid biosynthesis and degradation can correct vagal signaling in these patients needs to be explored.
Footnotes
Author contributions
MB, HBB, and IS designed the study. MB, JS, RWM, and IS collected the samples. HBB and CTJ measured the endocannabinoid levels. MB, HBB, and CTJ analyzed the data. MB and HBB drafted the manuscript. All authors were involved in the interpretation of the data, editing, and finalizing of the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Supported by TTUHSC EL Paso SARP grant to Irene Sarosiek.