
Abstract
Parathyroid hormone (PTH) interacts with components of the gut microbiota to exert its bone-regulating effects. This study aimed to investigate the gut microbial composition in patients with primary hyperparathyroidism (PHPT). Nine patients with PHPT and nine age–sex and body mass index-matched healthy controls were included. Gut microbial composition was assessed using 16S rRNA gene amplicon sequencing in both groups at baseline and 1 month after parathyroidectomy in the PHPT group. Data were imported into QIIME-2 and both QIIME-2 and R packages were used for microbiome analysis. Alpha and beta diversities were similar between the groups and remained unchanged after parathyroidectomy. The relative abundance of Subdoligranulum was significantly higher, whereas Ruminococcus, Alloprevotella, Phascolarctobacterium, and Clostridium sensu stricto_1 were significantly lower in PHPT than in controls (p < 0.001). After parathyroidectomy, the relative abundance of Subdoligranulum decreased, and Ruminococcus and Alloprevotella increased (p < 0.001). The PHPT group had lower total femoral and lumbar bone mineral density (BMD) than the controls (p < 0.05). At baseline, Alloprevotella abundance was positively correlated with serum phosphorus and Subdoligranulum was positively correlated with total lumbar BMD. Clostridium sensu stricto_1 was negatively correlated with serum calcium and positively correlated with femoral neck BMD. Postoperatively, Alloprevotella was positively correlated with baseline serum phosphorus and Phascolarctobacterium was positively correlated with distal radius BMD. This study demonstrated that the diversity of the gut microbiome was altered, possibly in response to electrolyte changes in PHPT, both before and after parathyroidectomy.
Introduction
Primary hyperparathyroidism (PHPT) is an endocrine disorder characterized by excessive secretion of parathyroid hormone (PTH) by one or more parathyroid glands, resulting in dysregulation of calcium and phosphorus metabolism. 1 Abnormal overproduction of PTH leads to hypercalcemia, hypophosphatemia, low bone mass, and osteoporosis. Clinical manifestations may include fatigue, bone pain, nephrolithiasis, neuropsychiatric symptoms, and gastrointestinal disturbances. 2 Diagnosis is established through biochemical assessment of calcium, phosphorus, urinary calcium, PTH levels, and imaging studies to identify the underlying pathology. Parathyroidectomy is the definitive treatment aimed at restoring calcium and phosphorus homeostasis. 1
Primary hyperparathyroidism (PHPT) is an endocrine disorder characterized by excessive secretion of parathyroid hormone (PTH) leading to hypercalcemia, hypophosphatemia, low bone mass, and osteoporosis.
The gut microbiota influences healthy bone turnover, and changes in microbial composition may contribute to the development of osteoporosis.
Short-chain fatty acid (SCFA)-producing bacteria modulate the effects of the gut microbiota on bone mass. Microbiota-dependent butyrate production is required for the bone anabolic activity of intermittent PTH release.
The relative abundance of Subdoligranulum was significantly higher in PHPT, whereas Ruminococcus, Alloprevotella, Phascolarctobacterium, and Clostridium sensu stricto_1 were significantly lower than in controls.
The relative abundances of the SCFA-producing bacteria Subdoligranulum, Ruminococcus, and Alloprevotella changed after parathyroidectomy.
Altered bacterial abundance in PHPT was correlated with hypercalcemia, hypophosphatemia, and decreased bone density.
Our findings might be helpful for better understanding bone loss in PHPT, characterized by sustained PTH release.
Further research on the influence of interventions in the gut microbiota on mitigating bone loss in patients with PHPT is needed.
The gut microbiome is a diverse ecosystem of microorganisms in the gastrointestinal tract that plays a vital role in human health. It consists of trillions of microorganisms, including bacteria, viruses, fungi, and other microbes. 3 Recent advances in sequencing technology have revealed the composition and functions of the gut microbiome. The gut microbiome influences digestion, produces beneficial metabolites, modulates immune responses, and protects against pathogens. Factors such as diet, genetics, some diseases, and lifestyle affect the gut microbiome. 4 Firmicutes and Bacteroidetes are the dominant bacterial phyla. 5
Hypercalcemia may affect bowel function as excessive calcium levels can reduce intestinal motility and impair peristaltic movements, leading to symptoms of constipation. 6 In patients with PHPT, hypercalcemia leading to slow transit constipation may also cause changes in the gut microbiota. 7 The gut microbiota influences healthy bone turnover, and changes in microbial composition may contribute to the development of osteoporosis. 8 Short-chain fatty acid (SCFA)-producing bacteria play a role in bone remodeling, 9 and butyrate-producing bacteria in particular have an anabolic effect on bone. 10 The microbiota-dependent production of butyrate is required for the bone anabolic activity of intermittent PTH release. 11
As far as we know, no study in the literature has investigated changes in the gut microbiome of patients with PHPT. This study aimed to investigate the alterations in gut microbial composition before and after parathyroidectomy for PHPT, as well as the effect of possible changes in microbiota composition on bone mineral density (BMD) in these patients.
Materials and methods
Identification of participants
This study was planned as a single-center, multidisciplinary, proof-of-concept pilot study. The sample size was determined using the G*Power 3.1 program based on studies with similar incidence. 12 Ten patients with PHPT, in addition to 10 healthy controls matched in terms of sex, age, and body mass index (BMI), were included in the study. Since sufficient gut microbial DNA could not be obtained after DNA extraction in one preoperative sample from the patient group, one postoperative sample result of this patient and one control counterpart of this patient were excluded from the study. All results are presented for nine patients with PHPT and nine controls.
The diagnosis of PHPT was made following the current guideline, with elevated serum calcium adjusted for albumin in the presence of a high or inappropriately normal intact PTH on two occasions at least 2 weeks apart. 13 Parathyroidectomy indications included asymptomatic patients who meet one or more of the following criteria: Age < 50 years, serum calcium concentration of ≥1 mg/dL above the upper normal limit, T-score at or below −2.5 at the lumbar spine, femoral neck, total hip or distal one-third of the radius, or the presence of vertebral fracture by X-ray, hypercalciuria (>250 mg/day in women; >300 mg/day in men), creatinine clearance of <60 mL/min, and kidney stones or nephrocalcinosis detected by abdominal imaging. 13
BMD measured using the Hologic Horizon-WI densitometer, serum calcium, phosphorus, albumin, PTH, urinary calcium, 25OH-vitamin D, and history of nephrocalcinosis or nephrolithiasis, and history of fracture were recorded. Patients who are diagnosed as PHPT in the thyroid/parathyroid outpatient clinic in our University Hospital and need parathyroidectomy are included. Exclusion criteria included the use of proton pump inhibitors, probiotics, or antibiotics that may change the stool microbiota in the last 15 days; having a primary bowel disease such as inflammatory bowel disease, irritable bowel disease, or gastrointestinal tract tumor; and having undergone any surgery that changes microbiota, such as systemic surgery or bariatric surgery, and had a history of smoking.
The healthy control group was selected from healthy volunteers who underwent routine endocrinology control. Age and BMI are the factors that affect the gut microbiome, 4 so the controls were chosen to be individuals similar in terms of age, gender, and BMI; they had no acute or chronic disease, no history of osteoporosis or gastrointestinal surgery, no smoking history, and no medication use. All female participants in the study were postmenopausal. Before taking stool samples, both the patient and control groups were given a 3-day standard diet list containing 20–25 g of fiber along with balanced carbohydrates, protein, and fat to eliminate the effect of diet on gut microbiota, and they were also asked to keep a nutritional record. In the PHPT group, medical follow-up was performed without any changes in medication.
All stool samples were collected at the outpatient clinic for both patients with PHPT and the controls. The first stool samples were obtained from the patient group before starting any PHPT-related therapy, and this group defined the PHPT group. The second stool samples of the patient group were obtained 1 month after parathyroidectomy when the serum PTH, calcium, and phosphorus had returned to normal, and this group defined PHPT after parathyroidectomy. As parathyroidectomy is a clean surgery, PHPT patients did not receive prophylactic antibiotics during surgery. No patient needed calcium replacement following parathyroidectomy. The patients were informed that they should not alter their medication and not use antibiotics or probiotics in the preoperative and postoperative periods. The stool samples of at least 5 mL were taken in 50 cc Falcon tubes and then immediately frozen. The samples were stored at −80°C without treatment to preserve their integrity. All samples were delivered to the laboratory, where the DNA analysis was conducted in accordance with cold chain requirements.
DNA extraction from stool samples
DNA extraction was performed using the QuickGene (Kurabo, Japan) extraction instrument and Tissue DNA Extraction kit (DT-S, Japan). First, 25 mg of stool sample was transferred to a homogenization tube with 250 µL Tissue Lysis Buffer (MDT) solution and 15 mg of 0.1 mmø zirconium beads were added to homogenize the samples. The tubes were then applied to 5000 rpm for 2 × 120 s. After homogenization, 25 µL Proteinase K (EDT) solution was added to the tube and incubated at 56°C for 60 min. Subsequently, the samples were centrifuged at 15,000g at room temperature for 10 min. 200 µL of the supernatant was then transferred to a sterile 1.5 mL microcentrifuge tube. After adding 180 µL LDT solution, the tube was vortexed for 15 s and incubated at 70°C for 10 min. 240 µL of 99% cold ethanol was then added to the tube and vortexed for 15 s. The whole volume inside the microcentrifuge tube was transferred to QuickGene (Kurabo) columns and three times washed with 750 µL WDT solution. Finally, 50–60 ng genomic DNA was collected to a new sterile 1.5 mL microcentrifuge tube by adding 200 µL Elution Buffer (CDT) solution to QuickGene (Kurabo) columns. The double-stranded DNA (dsDNA) quantification process was completed with Qubit™ dsDNA HS Kit (Thermo Fisher, PicoGreen® Dye) and Qubit Fluorometer-2 (Thermo Fisher Scientific, Waltham, MA, USA) device.
16S ribosomal RNA gene (rRNA) amplification and sequencing
Based on the extraction process results, bacterial 16S rRNA gene target sequencing was performed on the materials obtained in the study. The resulting genomic DNA was amplified using 16S V3-V4 314F-860R primer sets. 14 Library preparation was conducted using the Nextera XT Index and Nextera DNA Library Prep kits (Illumina Inc., San Diego, CA). The pooled libraries were cleaned up with size selection following the manufacturer’s protocol (AMPure XP, Beckman Coulter). After library preparation, sequencing runs were performed on an Illumina MiSeq sequencer (Illumina Inc., San Diego, CA, USA). Bacterial evenness and abundance data were obtained after sequencing and data handling.
Data handling and bioinformatics analysis
Pair-end Illumina reads (2 × 250) were imported to the Quantitative Insights into Microbial Ecology-2 (QIIME-2) environment. 15 All samples except one had more than 100X sequence depth. As mentioned before, the poor sample of the PHPT patient and its post-parathyroidectomy sample and healthy control counterpart were removed from the study. Divisive Amplicon Denoising Algorithm (DADA) introduced a model-based approach for correcting amplicon errors without constructing Operational Taxonomic Units. DADA2 was an open-source R package (https://github.com/benjjneb/dada2) that extends and improves the DADA algorithm. Quality clipping, chimera detection, and cleaning of reads were implemented through the QIIME2 DADA-2 pipeline (viaQ2 DADA-2). 16 Bases with low phred scores (https://www.arb-silva.de/documentation/release-138/) database. 17 The Phyloseq object was created from QIIME-2 artifact files in the R 4.1 environment. 18 Alpha diversity assessment was used to evaluate the diversity of related taxonomic units in a sample and was interpreted using three different indices, including Chao1, Shannon, and Simpson. P values between groups were calculated with the Kruskal–Wallis test. Principal component analysis, principal coordinate analysis (PCoA), and nonmetric multidimensional scaling (NMDS) were performed with R software. Beta diversity analysis, used to assess individual taxonomic differences, was calculated based on Jaccard, Bray–Curtis, weighted, and unweighted Unifrac. 19 Beta diversity statistical significance between groups was calculated using a PERMANOVA test, and intergroup p values were calculated using the Kruskal–Wallis test. Linear discriminant analysis Effect Size (LEFSe) analysis was conducted between groups to show statistically significant taxonomies. 20 Specific differences between groups were determined by differential abundance analysis using the Deseq2 R package. 21
Statistical analysis
IBM SPSS Statistics 22.0 was used for statistical analyses and the R package program version 4.1 for graphical representation and bioinformatics analysis. The conformity of continuous variables to normal distribution was evaluated using the Shapiro-Wilk test, while the homogeneity of variance was evaluated using Levene’s test.
Continuous variables with normal distribution were presented as the mean ± standard deviation). Continuous variables that were not normally distributed were presented as the median (min-max). Pearson’s χ2 (chi-square) test was used for categorical data and Student’s t-test for normally distributed continuous data. Preoperative and postoperative calcium and phosphorus values were compared using the Student’s paired t-test. Pearson correlation analysis was used for correlation analysis between clinical parameters and relative bacterial abundance. The p values were considered statistically significant when p < 0.05.
Results
There was no difference between PHPT at baseline and controls in terms of gender (p = 1.000), age (p = 0.404), BMI (p = 0.823), creatinine (p = 0.865), and vitamin D levels (p = 0.764). The PTH, calcium, phosphorus, urinary calcium levels and the distal radius, femoral total, femoral neck, and lumbar BMD values are presented in Table 1. After parathyroidectomy, calcium levels of PHPT were lower (9.51 ± 0.39 mg/dL) than baseline calcium levels (p < 0.001) and phosphorus levels were higher (3.20 ± 0.33 mg/dL) than baseline phosphorus levels (p = 0.021).
Demographic and clinical findings of the study groups.
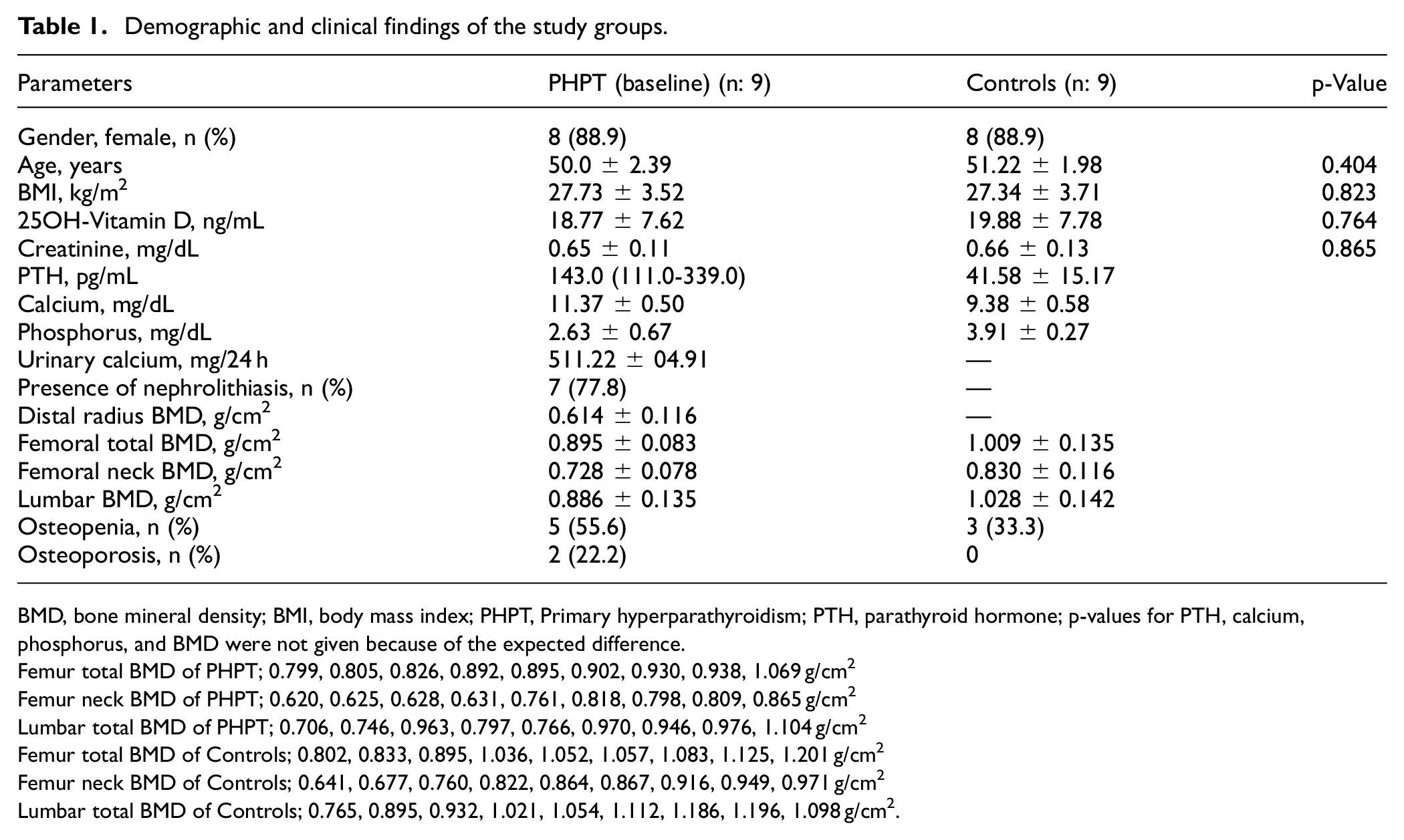
BMD, bone mineral density; BMI, body mass index; PHPT, Primary hyperparathyroidism; PTH, parathyroid hormone; p-values for PTH, calcium, phosphorus, and BMD were not given because of the expected difference.
Femur total BMD of PHPT; 0.799, 0.805, 0.826, 0.892, 0.895, 0.902, 0.930, 0.938, 1.069 g/cm2
Femur neck BMD of PHPT; 0.620, 0.625, 0.628, 0.631, 0.761, 0.818, 0.798, 0.809, 0.865 g/cm2
Lumbar total BMD of PHPT; 0.706, 0.746, 0.963, 0.797, 0.766, 0.970, 0.946, 0.976, 1.104 g/cm2
Femur total BMD of Controls; 0.802, 0.833, 0.895, 1.036, 1.052, 1.057, 1.083, 1.125, 1.201 g/cm2
Femur neck BMD of Controls; 0.641, 0.677, 0.760, 0.822, 0.864, 0.867, 0.916, 0.949, 0.971 g/cm2
Lumbar total BMD of Controls; 0.765, 0.895, 0.932, 1.021, 1.054, 1.112, 1.186, 1.196, 1.098 g/cm2.
Alpha and beta diversities
The Chao1 richness estimator and ASVs were used to estimate the community richness in the different samples. The larger the Chao1 and ASV indices are, the higher the richness of the community is. Although it was thought that ASVs and Chao1 narrowed in the PHPT at baseline, there was no statistically significant difference between the controls, PHPT at baseline, and PHPT group after parathyroidectomy (p > 0.05; Figure 1). The Simpson and Shannon indexes are commonly used to characterize species diversity in a community. They indicate both the richness and evenness of the species present. Although the diversity of the PHPT at baseline seemed to be higher for alpha diversity according to the Shannon index, there was no statistical difference in Simpson and Shannon index between the controls, PHPT at baseline, and PHPT group after parathyroidectomy (p > 0.05) (Figure 1).
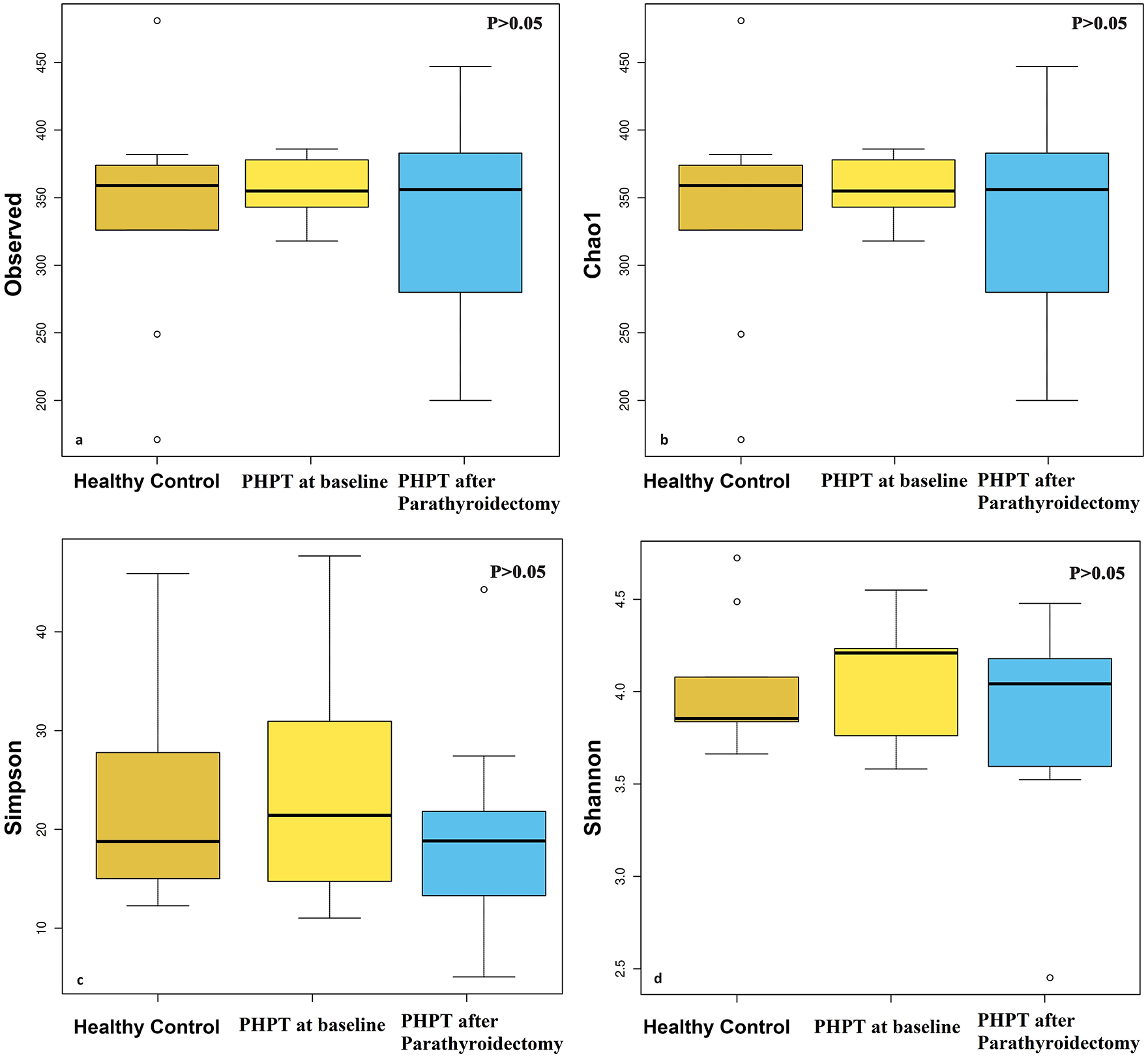
Alpha diversity plots to visualize the difference in microbiota structure between controls, PHPT at baseline, and PHPT group after parathyroidectomy. There was no significant difference in stool sample data between the groups in the study with ASVs (a), Chao1(b), Simpson (c), and Shannon indexes (d) (p > 0.05).
When beta diversity was evaluated, no difference was found between the groups with Bray Curtis, Jaggard, Unweighted Unifrac, and Weighted Unifrac analysis in both NMDS (Figure 2) and PCoA (data not shown) method (p > 0.05). Although a difference was observed between PHPT at baseline and controls in the Multiple Response Permutation Procedure (MRPP) analysis, no significant difference was found in terms of beta diversity in both MRPP (p > 0.05) and Adonis (p > 0.05).
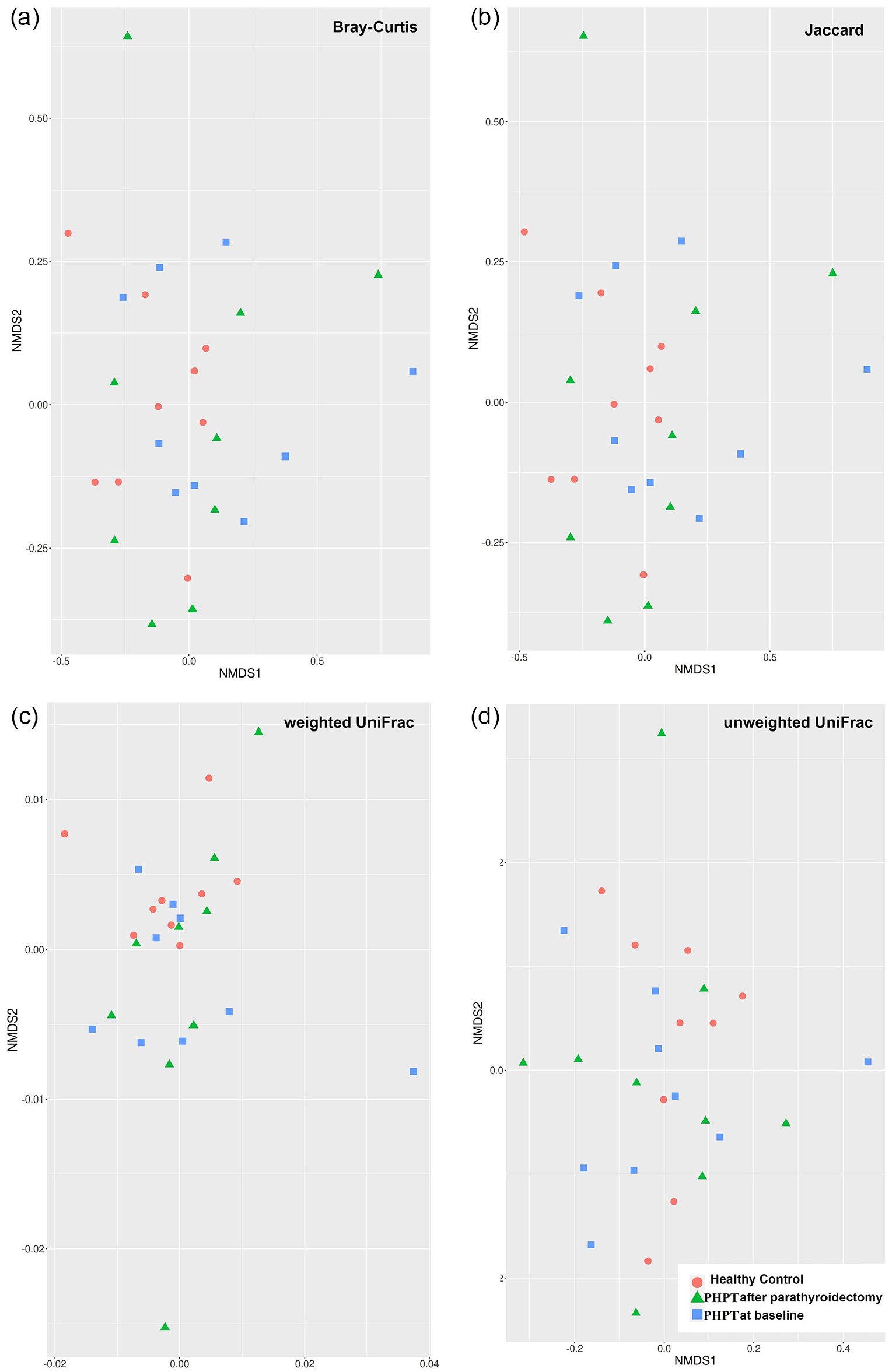
NMDS 2D plots of beta diversity analysis of the controls, PHPT at baseline, and PHPT group after parathyroidectomy. Each dot represents a stool sample. The red circle, blue square, and green triangle represent the controls, PHPT at baseline, and PHPT group after parathyroidectomy, respectively. Sample dissimilarities were measured by Bray–Curtis distances (a), Jaccard distance (b), weighted UniFrac distances (c), and unweighted UniFrac distances (d). There was no significant difference in stool sample data between the groups in the study (p > 0.05).
Biomarker taxa for each group
LEfSe was used to predict biomarker taxa in each group (LEfSe p < 0.05 and Linear discriminant analysis (LDA) score>2 for all taxa listed as enriched). The cladogram illustrates the taxa with different abundances between the groups. Both the PHPT at baseline and PHPT group after parathyroidectomy had less abundance of Clostridium sensu stricto_1 (LDA = 4.04, p = 0.036) and Phascolarctobacterium (LDA = 4.14, p = 0.038) at the genus level, and Clostridium_sensu_stricto_1_s (LDA = 4.04, p = 0.036) and Phascolarctobacterium_s (LDA = 4.09, p = 0.029) at the species level compared to the controls (Figure 3).
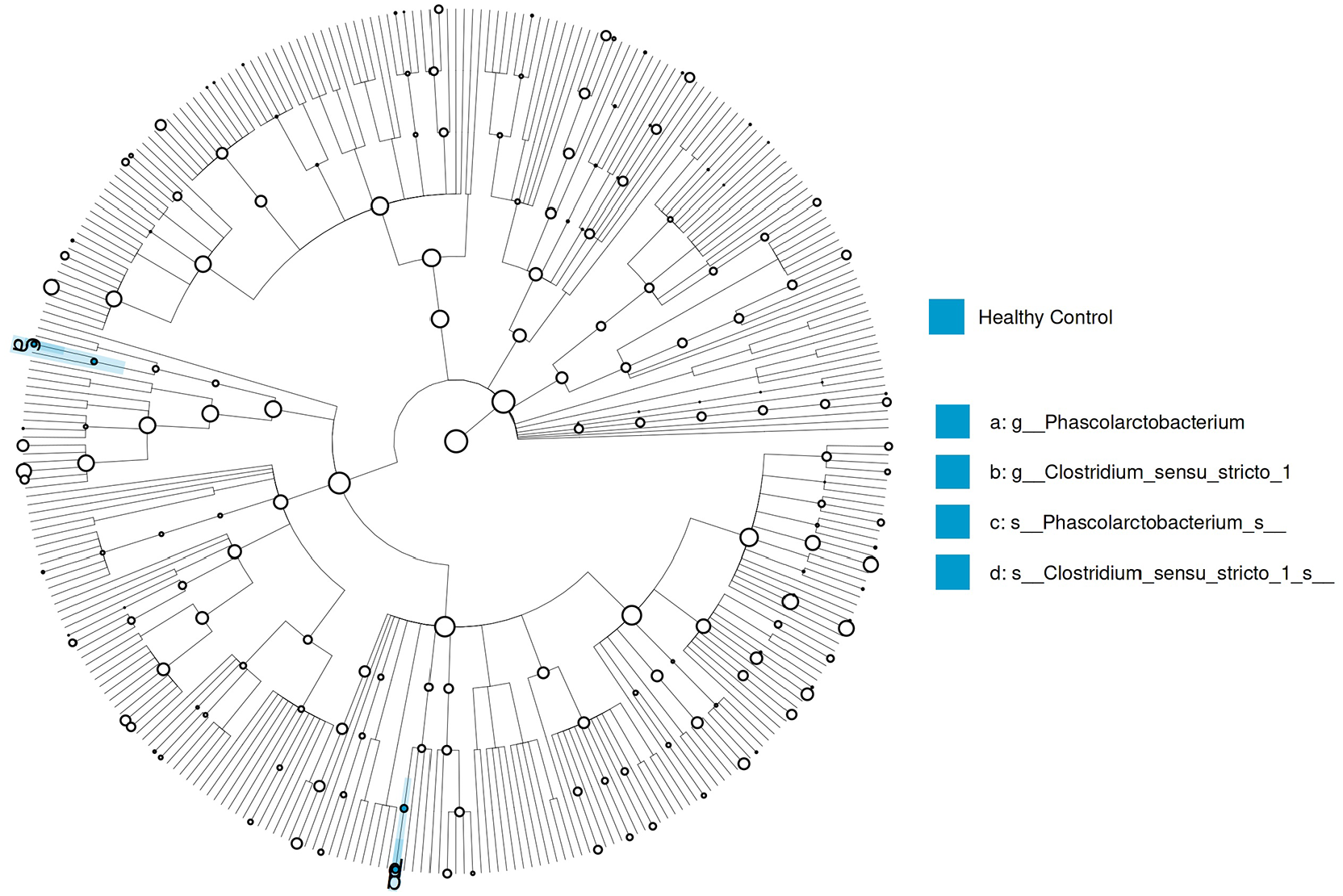
LEfSe analysis graphics showing bacterial taxa that were significantly different in abundance between the study groups. LefSe cladogram was performed to identify differentially abundant taxa. Only species and functional modules with LDA effect size>2 and p-value < 0.05 were plotted.
Differential abundance analysis
The PHPT at baseline showed the following phylum levels: Firmicutes (70%), Bacteroidetes (18%), and Proteobacteria (7%). In the PHPT group after parathyroidectomy, they were as follows: Firmicutes (77%), Bacteroidetes (13%), and Proteobacteria (5%). In the controls, the levels were as follows: Firmicutes (73%), Bacteroidetes (20%), and Proteobacteria (3%). The ratio of Firmicutes–Bacteroides was 3.65 in the controls, 3.88 in the PHPT at baseline, and 5.92 in the PHPT group after parathyroidectomy; regarding the presence of Bacteroidetes, no difference was observed between the groups (p > 0.05).
Microbiota composition is shown according to genus levels in Figure 4. The dominant genus in the controls was Prevotella (16%), Faecalibacterium (13%), and Dialister (11%), while the percentages of Prevotella (14%), Faecalibacterium (14%), and Dialister (14%) in the PHPT at baseline was equal. In the PHPT group after parathyroidectomy, Dialister (20%), Subdoligranulum (11%), and Prevotella (10%) have become the dominant group.
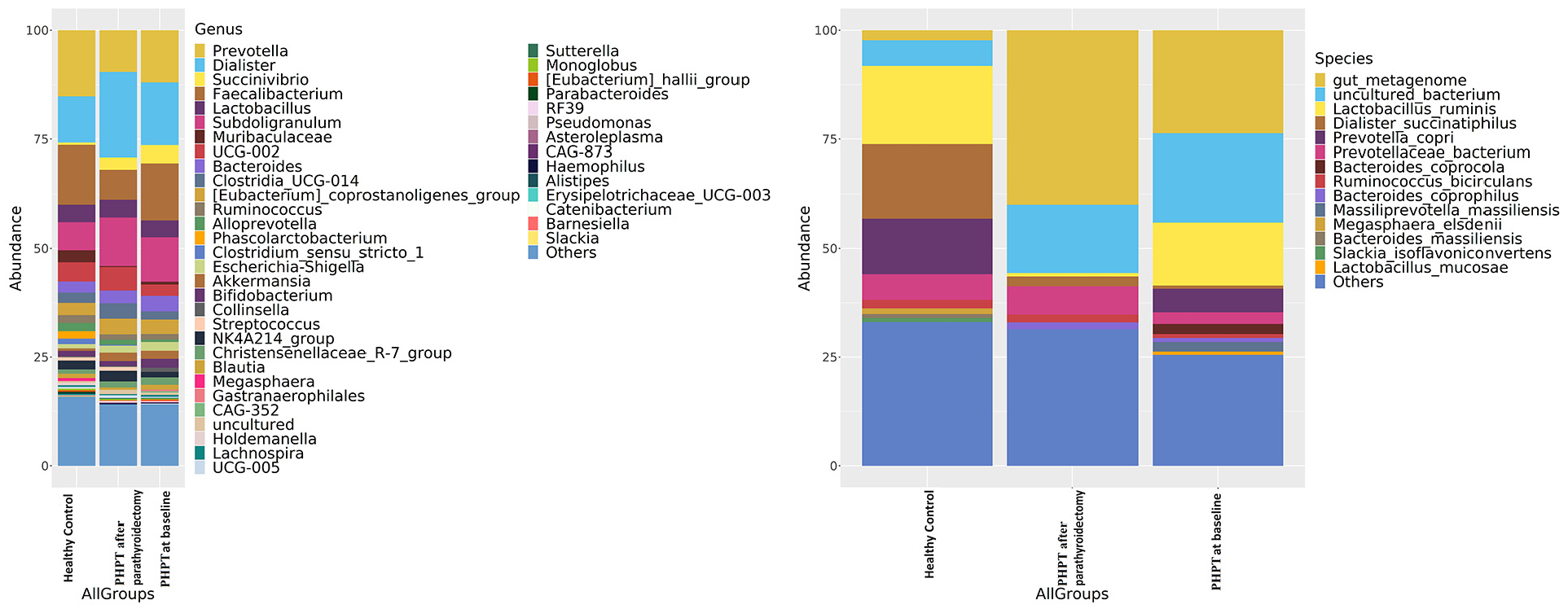
Intestinal microbiota composition of the study groups at species and genus levels. Bacterial community relative abundance analysis at the genus and species levels (relative abundance >1%; bacteria with relative abundances <1% were pooled in the “others” category and sorted by total concentration).
As shown in Figure 5, according to the Deseq2 differential abundance analysis, Ruminococcus (p < 0.001) and Alloprevotella (p < 0.001) were significantly less abundant, and Subdoligranulum (p < 0.001) was more abundant in PHPT at baseline than the controls at the genus level. After parathyroidectomy, Ruminococcus (p < 0.001) and Alloprevotella (p < 0.001) increased, and Subdoligranulum (p < 0.001) decreased compared to the PHPT group at baseline.
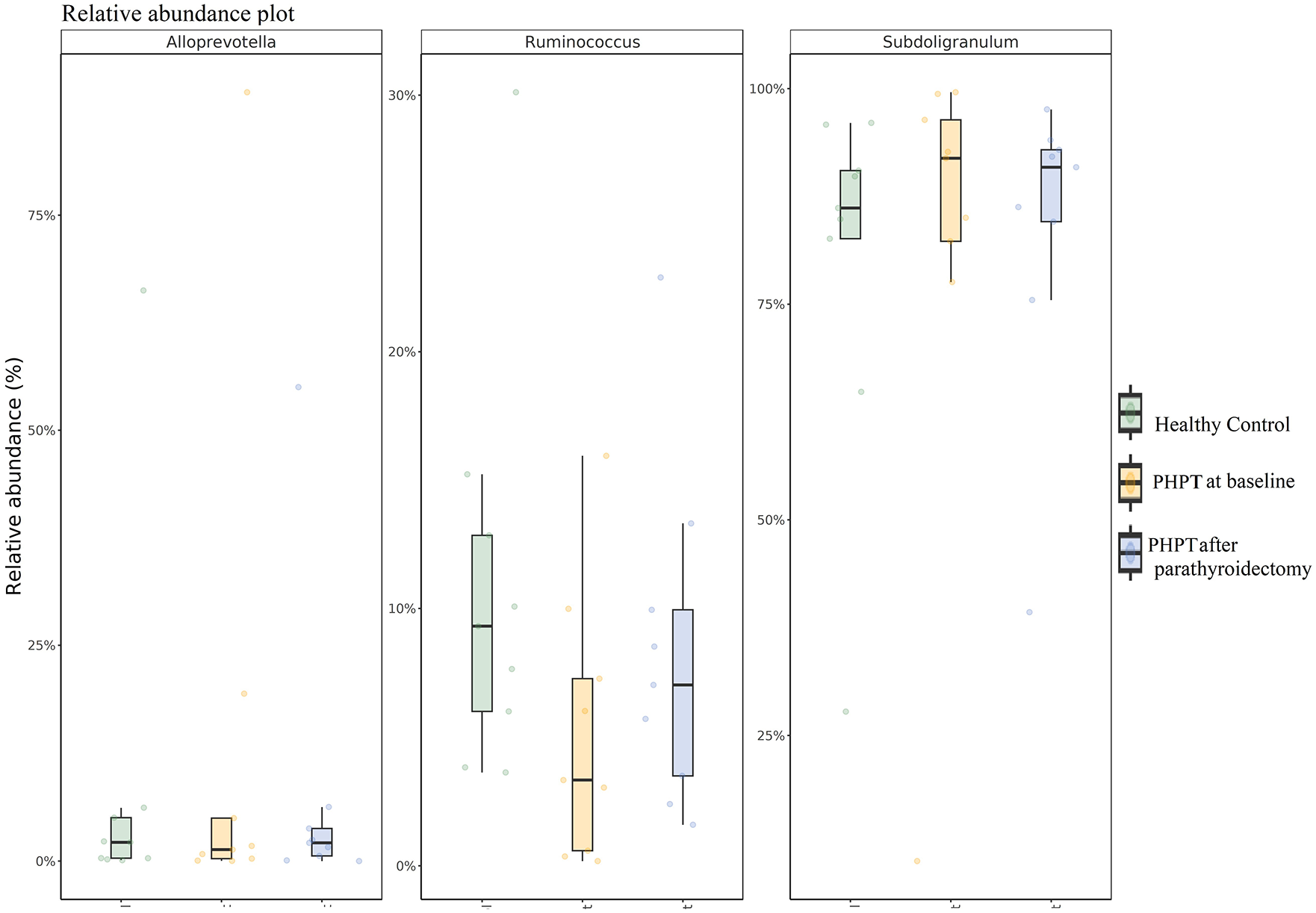
Relative abundance of Alloprevotella, Ruminococcus, and Subdoligranulum.
Microbiota composition is shown according to species levels in Figure 4. The dominant species Lactobacillus ruminis (22%), Dialister_succinatiphilus (18%), and Prevotella copri (14%) in the controls; Lactobacillus ruminis (16%), Prevotella copri (8%), and Prevotellaceae bacterium (4%) in the PHPT at baseline; and Prevotellaceae bacterium (7%), Lactobacillus_ruminis (4%), and Dialister_succinatiphilus (4%) in PHPT group after parathyroidectomy. According to the Deseq2 analysis, the Prevotellaceae bacterium was less abundance in the PHPT at baseline than the controls (p < 0.001), while it increased in the PHPT group after parathyroidectomy compared to both the PHPT at baseline (p < 0.001) and the controls (p < 0.001).
Correlations of clinical and laboratory parameters with gut microbiota metrics
The relationships between bacterial relative abundance in patients with serum calcium, phosphorus, PTH, vitamin D, urinary calcium, and BMD were examined in PHPT at baseline and after parathyroidectomy. At baseline, relative abundance of Alloprovetella (r = 0.934, p < 0.001) and Prevotellaceae bacterium (r = 0.944, p < 0.001) was positively correlated with serum phosphorus, and Clostridium sensu stricto_1 was negatively correlated with serum calcium levels (r = −0.848, p = 0.004) and positively correlated with femoral neck BMD (r = 0.800, p = 0.031). In addition, the relative abundance of the Subdoligranulum was positively correlated with lumbar total BMD (r = 0.884, p = 0.008). After parathyroidectomy, the relative abundance of Alloprovetella was positively correlated with baseline serum phosphorus level (r = 0.934, p < 0.001), Lactobacillus was negatively correlated with baseline urinary calcium level (r = −0.698, p = 0.037), and Phascolarctobacterium was positively correlated with distal radius BMD (r = 0.819, p = 0.046).
Discussion
In this study, we found that alpha and beta diversities of the gut microbiome were similar in patients with PHPT at baseline and after parathyroidectomy compared to age- and BMI-matched healthy controls. However, our results indicated that while Subdoligranulum was relatively more abundant, Ruminococcus, Alloprevotella, Phascolarctobacterium, and Clostridium sensu stricto_1 were less abundant in patients with PHPT. The relative abundance of Alloprevotella at baseline and after parathyroidectomy appeared to be associated with baseline phosphorus and Clostridium sensu stricto_1 at baseline also appeared to be negatively associated with baseline calcium. Furthermore, the decreased relative abundance of Clostridium sensu stricto_1 and Phascolarctobacterium appeared to be associated with the decrease in BMD. Moreover, the abundances of Ruminococcus, Subdoligranulum, and Alloprevotella in PHPT patients showed significant changes in the short term after parathyroidectomy, with the normalization of calcium and phosphorus.
Our study showed that in the evaluation of relative abundance, the dominant bacterial abundance at the phylum level belonged to the Bacteroidetes and Firmicutes phyla, similar to the normal population. Furthermore, there was no difference between the study groups in terms of alpha and beta diversities. Alpha diversity evaluates the diversity within a sample, including richness and evenness measurements, 22 while beta diversity evaluates differences in the microbiome among samples and is combined with dimensional reduction methods. 22 Bacteroidetes and Firmicutes were the prominent phyla of the gut microbiome, with the remaining bacteria representing less than 10% of the total relative abundance of bacterial phyla. 5 The lack of change in these parameters of our study group can be explained by the absence of a serious infectious or inflammatory condition in PHPT.
The most striking data in our study is that all of the bacteria whose abundance varies between groups are SCFA-producing bacteria. SCFAs are a group of fatty acids and also a major class of metabolites produced by the gut microbiota following the fermentation of indigestible polysaccharides. The most well-known SCFAs are acetate, propionate, and butyrate. 23 In our study, Subdoligranulum and Clostridium sensu stricto_1 produce butyrate,24,25Alloprevotella produce acetate, and Phascolarctobacterium produce propionate; moreover, the different species of Ruminococcus have been reported to produce either butyrate or acetate. 26 SCFAs are key mediators of cell function in a range of local, intermediary, and peripheral tissues. It is thought that calcium–phosphorus complexes decrease as SCFAs decrease local pH values in the intestines, 27 thus increasing calcium absorption in the presence of SCFA, especially acetate and propionate. 28 In our study, except for Subdoligranulum, SCFA-producing bacteria such as Ruminococcus, Alloprevotella, Clostridium sensu stricto_1, and Phascolarctobacterium were decreased in PHPT at baseline compared to the control group. Considering that SCFAs increase intestinal calcium absorption, it can be thought that the relative abundance decrease in these bacteria was an adaptive mechanism. The increase in abundance of Alloprevotella and Ruminococcus with the normalization of electrolyte levels after parathyroidectomy further supported our theory.
Hypercalcemia is a well-known cause of slow transit constipation. 7 Tian et al. found a high abundance of Bifidobacterium, Oscillospira, Parabacteroides, Prevotella, and Roseburia in patients with slow-transit constipation. This study also observed a decrease in acetate and propionate, two SCFAs, in patients with slow-transit constipation compared to healthy controls. 29 As previously mentioned, acetate and propionate play a role in calcium absorption. Constipation caused by hypercalcemia in PHPT may lead to a decline in calcium absorption due to an adaptive mechanism. This reduction can be linked to a decrease in the abundance of Alloprevotella and Phascolarctobacterium, which are bacteria known to produce SCFAs.
In ovariectomized osteoporotic rats, intermittent PTH treatment increased the relative abundance of Lactobacillus_reuteri, Muribaculaceae, Ruminococcaceae, and Clostridia in the gut microbiota and decreased Rikenellaceae. 11 In our study, we observed a decrease in the relative abundance of Ruminococcus and Clostridium sensu stricto_1, but no differences in abundance were observed between the groups in other bacterial genera. We believe that this decrease may be attributed to the continuous release of PTH in PHPT, as opposed to the intermittent release. In our study, the relative abundance of Alloprevotella and serum phosphorus showed a positive correlation and the relative abundance of Clostridium sensu stricto_1 and calcium also showed a negative correlation in patients with PHPT. Alloprevotella are obligate anaerobic, non-motile, gram-negative bacilli, which are weak to moderately saccharolytic and produce acetate and succinate as the end products of fermentation. 30 Clostridium sensu stricto_1 are anaerobic, fermenting bacteria, and butyric acid is a genus-specific product of fermentation. 25 Studies in the literature have reported changes in the relative abundance of bacteria in chronic renal failure, which causes an increase in serum phosphorus and a decrease in serum calcium, unlike PHPT. Li et al. showed that, although alpha and beta diversities were reduced in chronic renal failure, the abundance of Lactobacillus, Clostridium IV, Paraprevotella, Clostridium sensu stricto, Desulfovibrio, and Alloprevotella was enriched. The authors did not mention the relationship between microbiome analysis and electrolyte levels. 31 In addition, when the effects of dietary phosphorus restriction on microbial composition were examined in healthy adult humans, the relative abundances of Bacteroidetes, Ruminococcaceae, and Lachnospiraceae increased shortly after phosphorus restriction. 32 Furthermore, it was reported that calcium supplementation decreased the abundance of Provetella while a low-calcium diet increased the abundance of Prevotella and Subdoligranulum.33,34 Consistently, in our study, the abundance of Alloprovetella, Ruminococcus, Clostridium sensu stricto_1, and Phascolarctobacterium decreased in the presence of hypercalcemia and hypophosphatemia and after parathyroidectomy their abundance increased. On the other hand, the abundance of Subdoligranulum, which was initially high, decreased. In the study of Silva et al., a positive correlation was observed between the abundance of Lactobacillus and serum calcium in healthy premenopausal women, but this relationship was not found in postmenopausal women. In this study, the positive effect of Lactobacillus on bone density was also mentioned. 35 In our study, however, no relationship was found between the relative abundance of Lactobacillus and serum calcium. This result may be partly because our female participants were postmenopausal. Nevertheless, the inverse relationship between increased urinary calcium excretion and Lactobacillus abundance suggests the possible effect of microbiota on bone loss due to electrolyte changes in PHPT.
Osteoporosis and increased fracture risk occur as a result of deterioration in bone microarchitecture due to the continuous release of PTH in PHPT. It is stated that there is primarily a loss of cortical bone in PHPT; however, loss of trabecular bone also occurs particularly in post-menopausal women. 13 In our study, we found that both lumbar and total femoral BMDs were significantly lower in PHPT patients compared to the control group. In addition, femoral neck BMD was lower in PHPT patients, but no difference was detected between the groups. It is thought that the intestinal microbiota plays an important role in regulating bone turnover by modulating the immune system, controlling inflammation, and affecting calcium absorption. Furthermore, it is known that the intestine exerts some of its effects on bone through SCFAs. 36 SCFAs are regulators of osteoclast metabolism and bone mass. Treatment with SCFA increases bone mass and prevents postmenopausal and inflammation-induced bone loss. The protective effects of SCFA on bone mass are associated with the inhibition of osteoclast differentiation and bone resorption in vitro and in vivo. 37 Especially, butyrate suppresses osteoclast differentiation by increasing the production of osteoprotegerin in human osteoblasts. 38 Intermittent, physiological PTH release in the presence of butyrate in the gut expands Treg cells in the bone marrow and enhances bone formation in a Wnt-dependent way. 10 However, continuous PTH release and colonization with segmented filamentous bacteria in the gut lead to Th17 recruitment in the bone marrow resulting in RANKL-dependent bone loss. 39 Segmented filamentous bacteria, also known as Candidatus Arthromitus, are Gram-positive, non-culturable, spore-forming and not pathogenic members of gut microbiota. 40 In our study, the abundance of butyrate-producing Ruminoccoccus and Clostridium sensu stricto_1 and propionate-producing Phascolarctobacterium in PHPT patients decreased as well as the decreased relative abundance of Clostridium sensu stricto_1, and Phascolarctobacterium was found to be associated with lower BMD. These results suggest that PHPT may contribute to bone loss due to a change in the microbiome.
To the best of our knowledge, this is the first clinical study investigating the effect of both the disease and parathyroidectomy on changes in the gut microbiota of patients with PHPT. Our main limitation was the relatively small sample size. Although there are similar-sized gut microbiome studies in the literature, our observations need to be confirmed by larger sample studies. The second limitation of our study is that it is observational, so further research is needed to demonstrate causality.
Conclusion
This study demonstrated that PHPT caused changes in the diversity of the intestinal microbiome. The relative abundance of Subdoligranulum was higher, while Alloprevotella, Ruminococcus, Phascolarctobacterium, and Clostridium sensu stricto_1 were lower in PHPT than in the control group. Furthermore, after parathyroidectomy, there was a rapid reversal in the microbiome analysis during the early period; Subdoligranulum increased, while Alloprevotella and Ruminococcus decreased.
Footnotes
Acknowledgements
The authors thank all the study participants. We are grateful to Selma Avcu, RN, for collecting stool samples for the study group. In this study, DNA sequence analysis, microbiome analysis, and bioinformatics analysis were conducted in Diagen Biotechnology Laboratories.
Author contributions
All authors contributed to the study conception and design. Meric Coskun, Afruz Babayeva, and Tugba Barlas: Conceptualization, Methodology, and Data curation. Meric Coskun: Writing—Original draft preparation. Mehmet Muhittin Yalcin, Mujde Akturk, Fusun Balos Toruner, and Mehmet Ayhan Karakoc: Visualization and Investigation. Tarkan Karakan, Mehmet Cindoruk, and Ilhan Yetkin: Software and Validation. Alev Eroglu Altinova: Writing—Reviewing and Editing. All authors read and approved the final manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Endocrinology and Metabolism Training Coordination Association.
Ethical approval
This research involved human participants. This study was performed in line with the principles of the Declaration of Helsinki. Approval was granted by the local ethical committee (05.12.2022-893).
Consent to participate
Informed consent was obtained from all individual participants included in the study.
Data availability statement
The data are available from the corresponding author on reasonable request.