
Abstract

Introduction
The term, shortening deactivation, sometimes referred to as uncoupling, refers to the difference in the tension developed by an isometrically contracting muscle at a given length and a muscle permitted to shorten to that same length from a longer length during contraction.
The objectives of this review are to: 1) discuss proposed mechanisms of shortening deactivation of cardiac muscle; 2) describe what role shortening deactivation may play in normal and abnormal cardiac function; and 3) illustrate how shortening deactivation can be modified by both pharmacological and physiological interventions.
We will discuss work carried out both by ourselves and others on various types of skeletal and cardiac muscle preparations, whole heart preparations, and single cells. We will discuss the effects of pharmacologic and chemical interventions on shortening deactivation and will present the results of preliminary studies on chemically skinned papillary muscle.
Shortening deactivation appears to be involved in autoregulation of cardiac contractility. In the intact heart, it would be expected to influence diastolic filling and ventricular ejection, thus influencing myocardial oxygen consumption. It is hoped that the discussion will increase the understanding of the basic mechanisms by which active muscle shortening and ventricular contraction affect pressure development and stroke volume, and lead to a more complete understanding of the mechanisms by which ventricular function may be modified at a clinical level.
Overview
Studies of the interrelationships among cardiac muscle length, contractile state, and speed of shortening indicate that the developed tension is related to sarcomere length, myofilament overlap, the degree of activation, and the mode of contraction. There is substantial evidence that increased tension development associated with increased length of isometrically contracting muscle, as described by the Frank-Starling relationship, is because of a true increase in sarcomere activation as well as a more optimum overlap of the myofilaments.
When initial muscle length is plotted against actively developed tension, the classic length/tension (“Starling”) curve results, ie, the initial increase in length is followed by an abrupt and later, a more gradual increase in tension development. The increase in contractile tension noted immediately after the length change is not associated with an increase in the Ca2+ transient, as measured by aequorin, but appears because of an increased affinity of the myofilaments for Ca2+. The subsequent, more gradual increase is associated with an increased Ca2+ transient attributed to the release of more Ca2+ into the myoplasm.1,2
In addition to sarcomere length, myofilament overlap and length-dependent activation force development depends on the mode of contraction. Force developed at a given length is different if the muscle is fixed in length (isometric contraction) or if it is permitted to shorten from a longer length (isotonic or auxotonic contraction). If the muscle is permitted to shorten during the contraction, the force developed is less than predicted because of “deactivation” or “uncoupling” of the contractile system. Proposed mechanisms of deactivation have included: 1) reduced affinity of myofilament binding sites for Ca2+, leading to the release of Ca2+ from troponin and rapid uptake by the sarcoplasmic reticulum3,4; 2) increased passive resistance to sliding of myofilaments because of crossbridge distortion 5 ; and 3) fewer number of myosin heads available for binding with actin. 5
Previous Studies in Skeletal and Cardiac Muscle
Jewell and Wilkie 6 using frog skeletal muscle demonstrated that after lifting a load, the muscle became completely relaxed after shortening at a time when, had it contracted isometrically, it could have still developed force greater than the current load. In other words, the muscle could bear tension in an isometric state at a time when it could no longer actively shorten in an isotonic contraction, as though shortening increased the rate of exhaustion of energy stores. Hill 7 showed that if the tension of an isometric twitch was suddenly reduced to zero by a short, quick release near the peak, heat production stopped. This observation suggested that the persistence of isometric tension tends to maintain metabolic activity related to contraction, ie, the ability of the muscle to do work. Edman 8 showed that active shortening of skeletal muscle fibers caused depression of contractile force without a change in the total duration of contraction and suggested that the depressant effect derived from a structural change in the myofilament system. Huxley, 9 referring to Edman's work, speculated that during shortening, detached myosin heads with bound ADP would be unable to bind to actin sites; thus, the number of myosin heads capable of force generation would be reduced.
Qualitatively, similar effects of displacement on contractile force have been noted in cardiac muscle. Some of the earliest studies were carried out by Brady 10 who applied abrupt length changes to isometrically contracting rabbit papillary muscles. If the length changes were applied before or shortly after the stimulus, the subsequently developed tension was appropriate for the new length according to the previously determined isometric Frank-Starling relationship. However, if the length changes were applied beyond the midpoint of the rising phase of the contraction, the resultant developed tension was less than expected for the new length, suggesting that there was a “deactivating” or “uncoupling” effect on the force-generating system.
Kauffman et al 3 applied quick-release, release-stretch, and stretch-release perturbations to isometrically contracting papillary muscles from the right ventricles of cats. These investigators demonstrated that the magnitude of the uncoupling effect was directly related to both the stimulus intervention interval and the amplitude of the length change and was reduced if myofibrillar Ca2+ was increased by increasing Ca2+ in the bath. They also proposed that during the abrupt length change, Ca2+ is released from the crossbridges and taken up by the sarcoplasmic reticulum and is therefore unavailable for reformation of crossbridges. Thus, the duration and extent of activation would be diminished for the remainder of that contraction.
Brutsaert et al 10 described a technique referred to as “velocity clamping” to study the effects of a constant velocity of shortening on the interrelationship of force, velocity, length, and time on the duration of the active state in cat papillary muscle. They found that the time course of the active state could be prolonged if there was less shortening and more developed tension. They referred to the uncoupling effect of quick stretches and releases on the active state and speculated that the uncoupling effect was related more to the amount of shortening than to the velocity of shortening.
Edman 4 briefly summarized results of studies on skeletal and cardiac muscle carried out by other investigators and reported results of his own studies in both nonskinned and skinned frog skeletal muscle. He showed that deactivation was more pronounced in a single twitch than in a fused tetanus and that it was reduced after addition of caffeine to the muscle bath. He interpreted his data as indicating that the magnitude of deactivation is directly related to the state of activation, ie, the concentration of myoplasmic Ca2+. He concluded that active shortening caused a true deactivation of the contractile system, as would occur if Ca2+ binding to the myofilaments was reduced. Subsequently, Lab et al, 11 in studies of cat and ferret papillary muscles, found that action potentials as well as Ca2+ transients measured with aequorin were greater during active shortening than if the muscle were contracting isometrically. This study also suggested that shortening reduced the binding of Ca2+ to contractile proteins.
Depression of contractility during changes in length has also been noted in single cells. Tarr et al 12 found that stretch and release imposed on contracting isolated frog atrial cells resulted in depressed force and rate of force development. In a recent review, Brady 13 noted that during shortening of unattached myocytes, Ca2+ binding to troponin C is likely to be reduced compared to the isometric state. Therefore, developed tension would also be expected to be less.
Papillary Muscle Studies from Our Laboratory
Figures 1–6 illustrate the effects of rapid changes in length applied by a programmed ergometer during isometric contractions in isolated, superfused rabbit papillary muscles. Quick release (QR) and quick release-quick stretch (QR-QS) perturbations at a speed of 20 mm/sec were applied at various times during the rising phase of the contraction, and the subsequently developed force was compared to that of the control, unperturbed contraction.
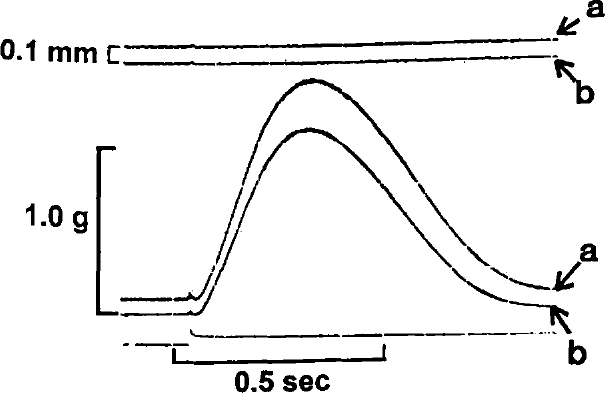
Superimposed isometric contractions of one rabbit papillary muscle are shown at resting lengths of 3.0 (b) and 3.1 mm (a). Following the contraction of muscle from an initial length of 3.1 mm, the muscle was shortened to 3.0 mm and a second contraction was recorded.
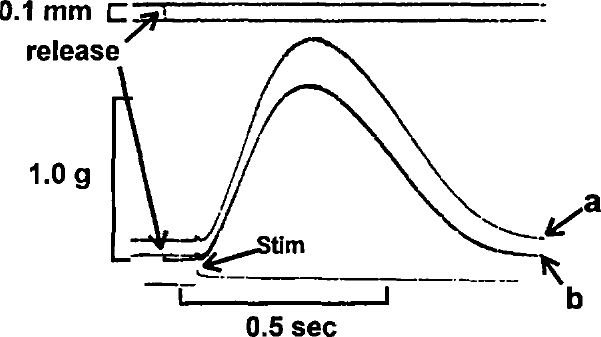
Superimposed isometric contractions of same muscle as shown in Figure 1. The muscle was quickly released @ 20 mm/sec, from 3.1 mm (a) to 3.0 mm (b) just prior to stimulus.
In Figure 1, two superimposed control contractions are shown at initial muscle lengths of 3.1 (a) and 3.0 (b) mm. Figure 2 illustrates the effects of quickly releasing (QR) the muscle from 3.1 (a) to 3.0 (a) mm just prior to the stimulus. It is clear that the tension developed following release to 3.0 mm is identical to that measured in Figure 1 for the same initial length. In Figure 3, the muscle is quickly released from 3.1 to 3.0 mm prior to the stimulus (b), at the time of the stimulus (b1), and 100 ms after the stimulus (b2). It can be seen that if the QR takes place significantly after the stimulus (b2), developed tension is less than that which would be predicted based on the control contraction from an initial length of 3.0 mm. In Figure 4, the muscle is released (QR) from 3.1 to 3.0 mm just beyond the midpoint of tension development (b1). In b2, with the initial muscle length set to 3.0 mm, a 0.1 mm quick release followed by a quick stretch (QR/QS) back to 3.0 mm is applied at the same time during tension development. Again, the tension developed following either perturbation is less than expected at the new length because of shortening deactivation.
Figure 5 illustrates the effects of QR-QS perturbations in another rabbit papillary muscle. The muscle was rapidly shortened 0.1 mm and returned to control length shortly after the stimulus (0) and at 100 ms intervals (1-4) until peak tension was reached. The tension following the initial QR-QS (0) was appropriate for the length but, beyond this point, the developed tension following QR-QS was clearly less than expected at intervals 1 through 4. In addition, the later in the contraction that the QR-QS was applied, the less the redeveloped tension; ie, the greater the deactivation.
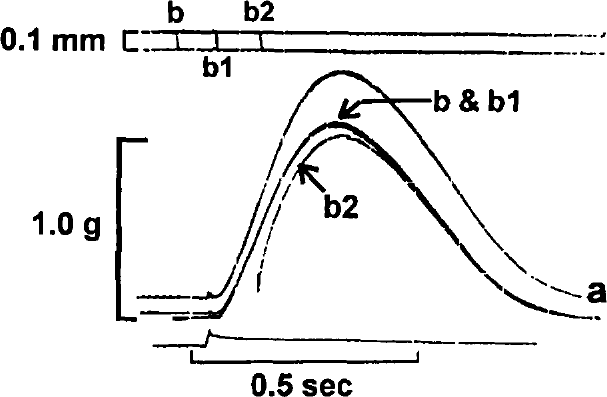
Superimposed contractions of the same muscle shown in Figure 1. The muscle was quickly released @ 20 mm/sec from 3.1 to 3.0 mm, prior to stimulus (b), at the time of the stimulus (b1), and 100 ms after the stimulus (b2).
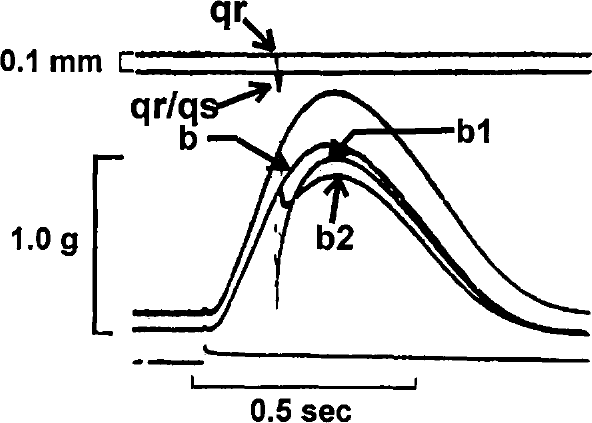
Superimposed contractions of the same muscle as shown in Figure 1. Control contractions were recorded at 3.1 mm (a) and 3.0 mm (b). A quick release (qr) @ 20 mm/sec, from 3.1 to 3.0 mm was applied just beyond the midpoint of tension development (b1). A quick release-quick stretch (qr-qs) of 0.1 mm was then done at the same time with the muscle set at a resting length of 3.0 mm (b2).
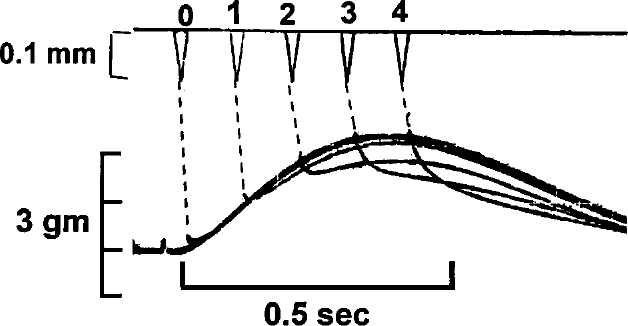
A control isometric contraction of another rabbit papillary muscle is shown. This is followed by a sequence of 0.1 mm quick release-quick stretch (qr-qs) perturbations applied at the time of stimulus (0) and at 100 ms intervals following the stimulus.(1-4).
In Figure 6, QR-QS perturbations of different magnitude (.05 mm, b1, and 0.10 mm, b2) are applied near the peak of the contraction. It is apparent that the greater the increment of the QR/QS that is applied, the greater the amount of deactivation.
Figures 2–6 indicate that the magnitude of deactivation is directly related both to the time at which the QR-QS is applied as well as to the size of the length change.
Because rapid length changes applied to an isometric contraction result in reduced tension via shortening deactivation, it would be expected that active muscle shortening at physiological speeds would also result in reduced force-generating capacity as the muscle length decreases. We therefore examined the effects of active shortening on developed tension in rabbit papillary muscle. 14 Tension during active shortening was compared to tension measured by the classic isometric length-tension relationship derived from individual contractions at the same muscle lengths. Figure 7 illustrates results from this study. All QRs were applied at the same time during the contraction and all were released the same increment of 0.1 mm, only the rate of change (dL/dt) of shortening was varied. With timing and magnitude of shortening held constant, both predicted and measured tensions were inversely related to the velocity of shortening. However, for any given length and velocity, tension developed by an actively shortening muscle was significantly less than that developed during isometric contraction at the same length. The fall in tension during shortening was time-dependent, suggesting that it is related to both the Hill force-velocity relationship 15 as well as to shortening deactivation.
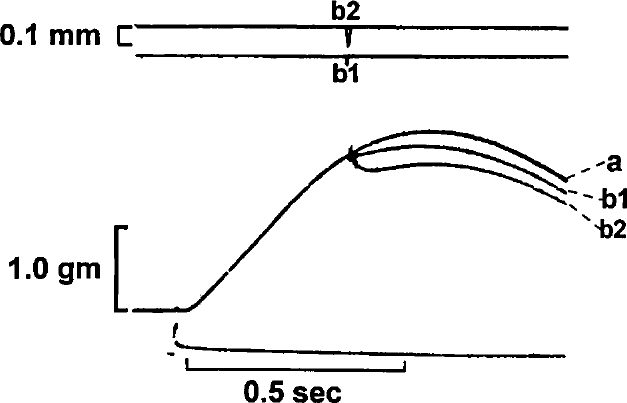
An isometric contraction (a) of a different rabbit papillary muscle set at a resting length of 3.5 mm is shown. This is followed by two quick release-quick stretch (qr-qs) perturbations of .05 mm (b1), and 0.1 mm (b2). The amount of shortening deactivation increases with the magnitude of the qr-qs.
Whole Heart Preparations
Studies carried out in whole heart preparations have demonstrated both similarities and differences between isolated papillary muscles and the whole ventricle. Suga and Yamakoshi 16 studied the relationship between stroke volume and isovolumic relaxation in an isolated heart preparation. Increasing stroke volume resulted in decreased duration of isovolumic relaxation. They theorized that this might be because of the uncoupling effect of muscle shortening, which would result in earlier opening of the mitral valve allowing greater time for ventricular filling.
Piene and Covell, 17 in a study of force-velocity relationships in left ventricular wall segments of dogs, found that developed force was not as dependent on velocity of shortening as reported in the papillary muscle studies of Leach et al 14 and suggested that this might indicate a functional difference between papillary muscle and ventricular myocardium.
Okada 18 determined the end-diastolic volume/stroke volume relation in a cross-circulated canine heart-lung preparation. He found that at a given afterload impedance, the stroke volume increased linearly with increasing end-diastolic volume. However, the increase in stroke volume was proportionately less than the increase in end-diastolic volume, indicating that ejection itself seemed to decrease contractility, presumably by increasing shortening deactivation. He found a similar linearity between the initial length and the amount of shortening in isolated frog ventricular muscle. He suggested that this deactivation was not because of reduced active-state duration but was more likely the result of a negative effect on Ca2+ utilization by the contractile system during the myofilament sliding phase of contraction, ie, shortening deactivation.
Nwasokwa et al 19 studied the effects of varying intracycle length or tension on force-length-time relations in a blood-perfused canine right ventricular papillary muscle preparation. Shortening resulted in a negative inotropic effect, which they attributed to a dissociation of Ca2+ from troponin and its uptake by the SR, making less Ca2+ available for subsequent force development.
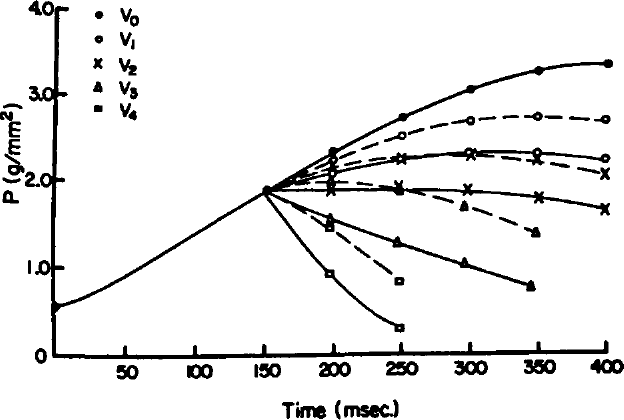
Isometric developed tension (P) was recorded from a rabbit papillary muscle at a resting length of 3.9 mm (V0, solid lines and closed circles). The muscle was then allowed to shorten the same increment at the same point in the contraction but at increasing velocities (dL/dt) of 0.4, 0.8, 1.2, and 4.0 mm/sec (V1-V4). Predicted tension is shown with broken lines and shortening tension with solid lines for the four shortening velocities shown in the Figure. (Reprinted from Am J Physiol 1980;238:H8-H13, with permission. 14 )
Physiological Model
More recently, Landesberg and Sideman 20 proposed a dynamic model of mechanical regulation of cardiac muscle in which they described intracellular control of contraction by relating the binding of calcium to troponin with cross-bridge cycling and force development. This model describes shortening deactivation and a calculated force deficit consistent with the studies of Leach et al 14 and Sagawa et al 21 and presumes that the force deficit depends upon: 1) the velocity of shortening, 2) the duration of shortening, and 3) the magnitude of change in developed force.
Chemically Skinned Cardiac Muscle Preparations
Studies from other Laboratories
Although shortening deactivation has been demonstrated in nonskinned cardiac and skeletal muscle preparations from many species, in skinned skeletal muscle fibers and in several types of isolated heart preparations, it has not yet been systematically studied in skinned cardiac muscle. Chemical skinning has been shown to make the sarcolemma permeable so that the muscle cannot respond to electrical stimulation but will contract when submicromolar concentrations of Ca2+ are added to the bathing solution. Depending on the concentration of a detergent such as Triton or saponin in the skinning solution, the sarcoplasmic reticulum (SR) can either remain functional or become nonfunctional.22,23 Thus, the effects of selected interventions on contractile function can be determined with known concentrations of Ca2+ in the myoplasm, with or without the influence of Ca2+ release from or uptake by the SR. Although chemical skinning has been shown to reduce Ca2+ sensitivity of the myofilaments, qualitative changes induced by many different interventions remain similar to those produced in nonskinned muscle. 24
Studies from Our Laboratory
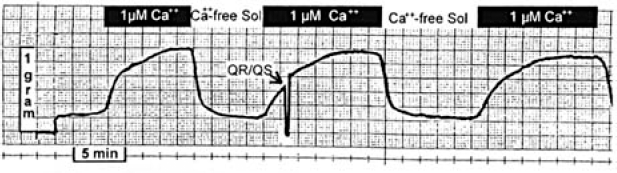
Figure 8 shows an isometric contraction of a nonskinned rat papillary muscle (Control) followed by a QR-QS of 5% of muscle length. Tension developed following the QR-QS is substantially less than in the isometric contraction because of shortening deactivation. Note that both contractions take place from the same initial length. The muscle was then superfused with Ca2+-free Tyrode's solution (Figure 9). This was followed by superfusion with a skinning solution containing 1.0% Triton. In Figure 10, contraction of the skinned muscle is induced by superfusion with a 1.0 µmol/L free Ca2+ solution followed by superfusion with a Ca2+-free relaxing solution. During the second contraction, a QR-QS was applied and the muscle was again relaxed. A third control contraction was then obtained. As can be seen, there was no apparent reduction in peak tension following the QR-QS, suggesting that deactivation did not occur.

An unperturbed isometric contraction of a nonskinned rat papillary muscle is shown (Control) followed by a contraction that is interrupted by a quick release-quick stretch (QR-QS) of 5% of muscle length.
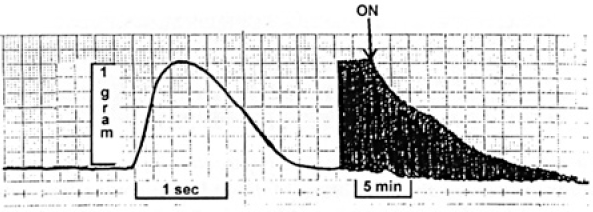
The same muscle as shown in Figure 8. Isometric contractions are recorded both for a control period and following the onset of superfusion with Ca2+-free Tyrode's solution at the arrow.
Since the muscle was skinned with 1.0% Triton as a detergent, it suggests that absence of a functional SR prevented the development of shortening deactivation. If shortening deactivation can be demonstrated with concentrations of saponin known to preserve SR function, it will further support the concept of the requirement of a functional SR in order for shortening deactivation to take place. These studies are planned for the future.
Putative Mechanism of Shortening Deactivation
Increased muscle length has been shown to result in increased myofilament Ca2+ affinity and binding resulting in increased crossbridge attachment. 1 Therefore, it is expected that during active muscle shortening, myofilament Ca2+ affinity and binding would be reduced and the free Ca2+ released by this mechanism would be taken up by the SR. Studies in both nonskinned and skinned ventricular muscle preparations have shown that a sudden reduction in length of a contracting muscle results in a rapid increase in myoplasmic Ca2+.11,25 This has been attributed to detachment of crossbridges during shortening, leading to decreased Ca2+-troponin affinity and the release of Ca2+ into the myoplasm. 25 The released Ca2+ is then taken up by the sarcoplasmic reticulum, leaving less Ca2+ available for subsequent force development.
Although attachment of crossbridges and increased tension have been shown to increase myofilament Ca2+ affinity and binding, the magnitude of shortening deactivation increases the later that it occurs in the rising phase of the contraction. This reflects the fact that the Ca2+ transient peaks early in the contraction, then subsides as free Ca2+ becomes attached to troponin C or is taken up by the SR. When the muscle is re-extended to its original length, less Ca2+ is now available to bind to troponin C and the redeveloped tension is decreased relative to that seen before the release. Thus, the later in the contraction that the muscle is permitted to shorten, the less the available Ca2+ and the greater the deactivation. So, even though myofilament Ca2+ affinity is increased by tension development, less Ca2+ is available to re-attach to troponin C if an abrupt length decrease occurs later in the cycle. 25
If active muscle shortening is associated with reduced Ca2+ affinity and binding, is it the change in length or the change in tension that is responsible? Most of the available evidence suggests that myofilament Ca2+ affinity or sensitivity is determined primarily by the number of attached crossbridges, supporting the view that the tension change is responsible for the Ca2+ release.1,26 Allen and Kentish 1 reported that the affinity of troponin C for Ca2+ correlated more closely with developed tension than with muscle length. Saeki et al 27 studied tension and intracellular Ca2+ transients following rapid length changes in ferret papillary muscles during ouabain contracture and ryanodine tetanic contraction. They found that rapid release or stretch of ouabain-activated muscle resulted in oscillatory responses of tension and the aequorin light signal. The time course of the aequorin light signal corresponded to the time course of the tension transients, ie, the increase in aequorin light corresponded to the decrease in tension, and the decrease in aequorin light corresponded to the increase in tension. This indicated to them that the aequorin light changes were driven by the tension changes and suggested that the affinity of troponin C for Ca2+ was directly related to the muscle tension (ie, cross-bridge attachment) and that the released Ca2+ was taken up by the SR.
Isometric contractions of a the same rat papillary muscle as in Figure 9. Contractions after superfusion with Tyrode's solution containing 0.5 µmol/L Ca2+ followed by superfusion with a “skinning” solution containing 1% Triton. Contractions are then produced by superfusion with Tyrode's solution containing 1.0 µmol/L Ca2+ followed by Ca2+-free Tyrode's to produce relaxation. The first and last contractions are controls while the middle contraction shows the effect of a QR-QS.
Effects of Pharmacological and Physiological Interventions
Studies from Our Laboratory
Recently completed studies in our laboratory illustrate the effects of caffeine, acidosis, and strontium on shortening deactivation. Quick release-quick stretch perturbations were applied to isometrically contracting rat papillary muscles at 10 ms intervals during the rising phase of the contraction during superfusion with control Tyrode's solution and during superfusion with 10 mmol/L caffeine, during acidosis produced by gassing the superfusate with 70% O2, 30% CO2, and after addition of 5.0 and 10 mmol/L strontium to the superfusate. Percent deactivation was calculated by dividing the peak tension that developed following the QR-QS by the peak tension of the control contraction to normalize the responses. Shortening deactivation was reduced by 10 mmol/L caffeine (Figures 11,12), increased during acidosis (Figure 13), and essentially abolished by 10 mmol/L strontium (Figure 14).
Caffeine (10 mmol/L) has been shown to increase myofilament Ca2+ responsiveness in cardiac muscle from many species, 28 while acidosis reduces myofilament responsiveness to Ca2+ by decreasing Ca2+ binding to troponin C.29,30 Thus, one would predict that Ca2+ responsiveness would be inversely related to shortening deactivation.
Finally, strontium, which shows the same affinity as Ca2+ for the contractile system, has been shown to activate sarcomere contraction. 31 However, it is generally believed that strontium is neither taken up by nor released from the sarcoplasmic reticulum but that sarcomeres are activated by strontium entering from the extracellular space. In our studies, increasing concentrations of strontium reduced and essentially abolished shortening deactivation (Figure 14).
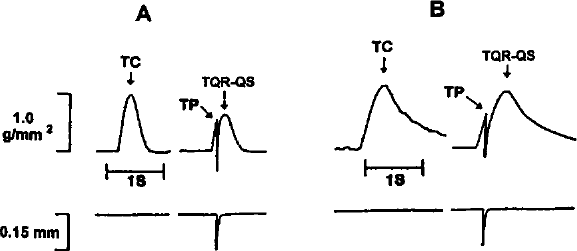
Quick release-quick stretch of a rat papillary muscle is shown before (A), and during (B) superfusion with Tyrode's solution containing 10 mmol/L caffeine. Abbreviations: TC = tension of control contraction; TP = tension at time of QR-QS perturbation; TQR-QS = tension developed following QR-QS.
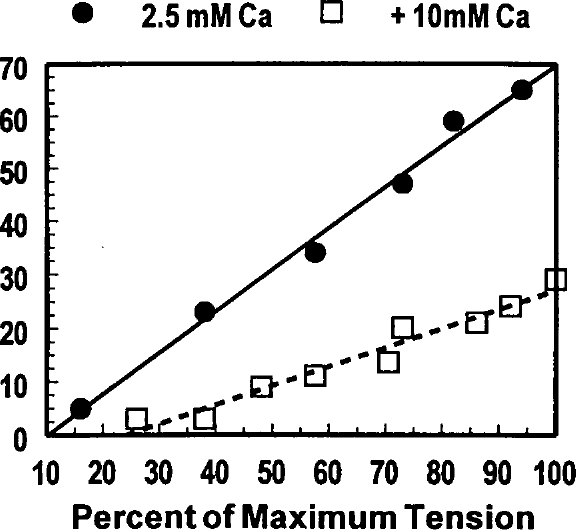
Effects of 10 mmol/L caffeine on shortening deactivation are illustrated. Percent deactivation is plotted against the percent of maximum tension developed following QR-QS during superfusion with control Tyrode's solution (closed circles and solid line) and Tyrode's solution containing 10 mmol/L caffeine (open squares and broken line). Maximum tension refers to the highest developed tension at which the QR-QS was applied.
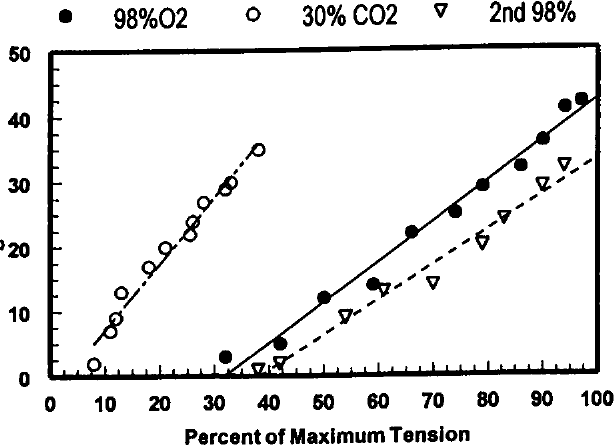
The graph shows the effects of acidosis on shortening deactivation. Again, percent deactivation is plotted against the percent of maximum tension developed following QR-QS. Under control conditions, the muscle was superfused with Tyrode's solution gassed with 98% O2 + 2% CO2 (pH 7.2-7.4). Acidosis was induced by superfusion with Tyrode's solution gassed with 70% O2 + 30% CO2 (pH 6.4-6.6). Clearly, acidosis significantly and reversibly inhibits shortening deactivation. See Figure for symbol legend.
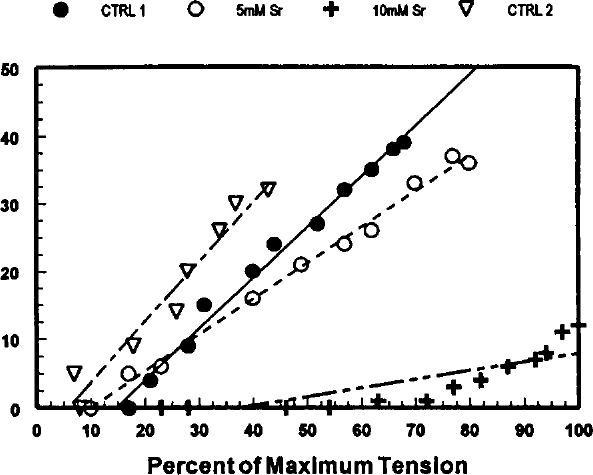
Effects of strontium (Sr2+) on shortening deactivation. The data are presented in Figures 12 and 13. Control data were recorded during superfusion with normal Tyrode's solution (CTRL 1 … CTRL 2). In between these two controls, the muscle was superfused with Tyrode's solution containing 5 and 10 mmol/L Sr2+. See Figure for symbol legend.
Thus, the effects of caffeine, acidosis, and strontium on shortening deactivation lend further support to the hypothesis that the reduced tension associated with active muscle shortening is largely the result of release of Ca2+ from troponin and its subsequent uptake by the sarcoplasmic reticulum.
Summary and Conclusion
Physiological Mechanism
A rapid change of length applied during isometric contraction of skeletal or cardiac muscle may result in redeveloped tension less than appropriate for the new length because of “deactivation” of the contractile system. The amount of shortening deactivation is directly related to both the time during the contraction when the length change occurs and to the extent of muscle shortening. If the muscle is permitted to shorten early in the contraction, the redeveloped tension will be appropriate to the new length as predicted from the classic Frank-Starling relationship. However, the same length change, which is imposed later in the contraction, results in a redeveloped tension that is less than predicted. Furthermore, a greater change in length results in less tension being redeveloped than if a smaller length decrement is applied at the same time during the contraction.
It has been demonstrated that the reduced tension during active muscle shortening is associated with reduced affinity of troponin C for Ca2+. The free Ca2+ is then picked up by the SR, with less Ca2+ available for tension development until the subsequent contraction.
Clinical Significance
Although the clinical significance of shortening deactivation remains speculative, it seems likely that in the intact heart deactivation would affect myocardial O2 consumption. The decreased efficiency with which the heart maintains a given stroke work against a high afterload might be related to the lesser degree of fiber shortening and, therefore, less shortening deactivation. Conversely, it is well-known that the same level of stroke work accomplished by an increase in end-diastolic volume requires much less O2. This may be related, at least in part, to the greater degree of shortening with an accompanying increase in deactivation under the latter conditions. For example, in congestive heart failure where ejection fraction and fiber shortening are minimal, the maintenance of the longer fiber lengths could significantly increase the MVO2. Ford 32 has suggested that the deactivating effect of shortening produced by afterload reduction would limit energy expenditure, therefore, exerting a favorable effect on the failing myocardium. It would also seem that an inotropic agent that increased shortening deactivation might compensate for the increased MVO2 caused by the inotrope and have a favorable effect on cardiac work.
From most of the studies we have reviewed, it appears likely that shortening deactivation acts as a physiological “feedback” mechanism that affects afterload and in turn, myocardial oxygen consumption. Pathological situations such as acidosis and ischemia have been associated with reduced myofilament Ca2+ sensitivity or affinity and depressed cardiac contractility.29,30 Is it then possible that interventions that increase Ca2+ sensitivity might favorably alter ventricular pressure-volume relations during ejection and improve myocardial function by reducing the magnitude of shortening deactivation?
Whatever the mechanism and clinical significance, future investigations will help to define the role of shortening deactivation in modifying ventricular function.