
Abstract
Our objectives were to investigate the chemical composition, nutritive value and antioxidant activity potentials of Terfezia boudieri Chatin collected from different areas from Southeast of Turkey. The amounts of energy, protein, fat, carbohydrate, crude fibre, moisture and ash were 34.00–65.00 (kcal/100 g), 1.40–2.73, 0.86–1.71, 4.84–11.60, 80.86–90.33 (g/100 g, f.w.) and 1.02–1.98 g/100 g dry mass, respectively. Fatty acids and amino acids analyses revealed that linoleic acid and glutamic acid were, respectively the most abundant. Among the essential amino acids leucine was the highest amount and this was followed by threonine. Magnesium was the most abundant element with a concentration value of 182.30 mg/100 g dry mass. T. boudieri exhibited excellent antioxidant activity when compared with standard compounds. In total antioxidant and chelating effect on ferrous ions activity T. boudieri showed the highest activity against the standards. Additionally, at all studied tests antioxidant activity of the mushroom increased with increasing concentration. The total phenolic content of T. boudieri was found as 8.45 mg/g.
Introduction
Desert truffles are a type of an obligate hypogeous ascomycetes ectomycorrhizal fungi formed in association with host roots of Helianthemum spp. and the soil inhabiting fungi Terfezia or Tirmania spp. (Al-Laith, 2007). The type of mycorrhizal fungus ramifies through the soil, absorbing nitrogen and other minerals, which is transported back to the host plant (Ewaze and Al-Naama, 1989). Desert truffles are seasonal and socio-economically important fungi. These truffles are edible and grow wild in the southeast part of Turkey.
The truffles usually appear in the deserts following the rainy season between February and April. Truffles are grown naturally in large quantites in virgin lands in the Middle East and North Africa (Morocco, Tunisia, Algeria and Egypt), Southern Europe (Spain, France, Italy and Greece) and other Mediterranean bordering countries (Libya, Tunisia, Syria) as well as in Iran, Iraq, Kuwait and Turkey (Al-Delaimy and Ali, 1970; Al-Delaimy, 1977; Ahmed et al., 1981a; Paul, 1983) during the autumn rain and thunder storms (Gücin and Dülger, 1997). In these countries, no data is available on total production and consumption.
The truffle is a type of tuber highly prized for its unique musky aroma and flavour. The popularity of truffles is believed to be due to their nutritional value and delicious taste. Wild edible fungi not only add flavor to bland staple foods but they are also valuable foods in their own right. As such, desert truffles are a rich source of protein, amino acids, fatty acids, minerals and carbohydrates (Bokhary and Parvez, 1993). Similar to other fungi, desert truffles comprise a vast and yet largely unexploited source of new pharmaceutical products. In search for new therapeutic alternatives, and most importantly for modern medicine, truffles represent an unlimited source of therapeutic compounds with anti-inflammatory, immunosuppressor, antimutagenic and anticarcinogenic properties (Hannan et al., 1989), antioxidant properties (Murcia et al., 2002), antimicrobial (Janakat et al., 2004) and steroidal glucoside with polyhydroxy ergosterol nucleus (tuberoside) (Gao et al., 2001).
Specific and peculiar attributes of desert truffles like hypogeous, mycorrhizal and microaerophilic make their metabolism interesting model to study, especially, under stressful desert arid environment conditions. Truffles that live under microaerobic habitats are expected to show morphological, physiological and biochemical adaptations to an environment poor in oxygen as found under hypogeous conditions. As mycorrhizal symbiotic fungi of several species of plants, they exchange metabolites, mineral salts and ions. These adaptations are largely controlled by several environmental factors like the rainy season with its amount and timing, soil types and characteristics, water availability and climatic conditions (Bokhary and Parvez, 1993).
Very little information is available relating to truffle metabolism and, in particular, oxidative metabolism (Pacioni et al., 1995). Oxidation is necessary to many organisms for the production of energy to support metabolism of aerobic cell. However, in some instances, the production of oxygen through uncontrolled metabolic pathways results in free radicals. This imbalance can result in damage to molecules, including lipids, DNA, carbohydrates and proteins (Dubost et al., 2007). Moreover, these free radicals are potentially involved in the onset of many diseases such as cancer, rheumatoid arthritis and atherosclerosis as well as in degenerative processes associated with aging (Halliwell and Gutteridge, 1984). Exogenous chemical and endogenous metabolic processes in the human body or in the food system might produce highly reactive free radicals, especially reactive oxygen species (ROS) capable of oxidising biomolecules, resulting in cell death and tissue damage (Halliwell and Gutteridge, 2003). On a cellular basis, damage provoked by these free radical formations is usually protected by oxidative enzymes as well as compounds such ascorbic acid, tocopherols and phenolics. When the mechanism of antioxidant protection becomes unbalanced by factors such as aging, deterioration of physiological functions may occur resulting in diseases and accelerated aging. However, the antioxidant components present in truffles are of great interest as possible protective agents to help reduce oxidative damage. Previous reports had shown that the 120 expressions of some antioxidant enzymes were correlated with the microaerobic metabolism, growth rate and mycorrhizal symbiosis of truffles (Amicarelli et al., 1999). Wild and cultivated mushrooms are well known to contain various polyphenolic compounds which are recognised as an excellent antioxidant due to their ability to scavenge free radicals by acting as reducing agents, hydrogen donating antioxidants and singlet oxygen quenchers (Barros et al., 2007).
Some common edible mushrooms have been found to possess antioxidant activity which is well correlated with their total phenolic contents (Lo and Cheung, 2005). According to our litearature survey data about the antioxidant/antiradical activity and the antioxidant compounds of Terfezia boudieri are lacking all over the world especially we did not find any data the ones grow southeast of Turkey. Thus, there is an increasing demand to assess these attributes on truffles and other wild fungi. One of the aim of this study was to investigate the antioxidant activitiy of T. boudieri. For this purpose mushroom sample was analysed in different test systems namely β-carotene–linoleic acid, DPPH, reducing power, chelating effect on ferrous ions and superoxide anion radical scavenging activity. The chemical composition and nutritional value of the mushrooms were determined, with reference to the contents of energy, protein, fat, carbohydrate, dietary fibre, moisture, ash, amino acids, fatty acid and minerals matter.
MaterIals and methods
Materials
Mushroom samples and chemicals
In the period of April–May 2006 at four different areas (Hilvan-Sanliurfa, Yenice/Ceylanpinar/Sanliurfa, Polatlı/Ceylanpinar/Sanliurfa, Kiziltepe-Mardin and Malatya) from Southeast of Turkey ascocarps of T. boudieri were collected. Mushroom samples were carried into the laboratory in an ice bath and stored deep-frozen at −34 °C until used. All other biochemical studies were completed in 10 days.
1,1-Diphenyl-2-picryl-hydrazyl (DPPH), ferrous chloride, polyoxyethylenesorbitan monolaurate (Tween-20), α-tocopherol, 3-(2-pyridyl)-5,6-bis (4-phenyl-sulfonic acid)-1,2,4-triazine (Ferrozine), butylated hydroxyanisole (BHA) and trichloraceticacid (TCA) were purchased from Sigma (Sigma-Aldrich GmbH, Sternheim, Germany). Ammonium thiocyanate and butylated hydroxytoluene (BHT) were purchased from E. Merck. All other chemicals were analytical grade and obtained from either Sigma-Aldrich or Merck.
Methods
Sample preparation
Fresh samples of mushrooms, after removal of external material such as mud, bush, soil, plant, etc. by washing with demineralised water, were airdried between filter papers. Approximately 5 g of each sample were taken immediately for determination of moisture. Remaining samples were stored in deep-frozen until analysis.
Chemical analysis
Samples of mushrooms were analysed for chemical composition (moisture, protein, fat, carbohydrates, crude fibre and ash) using the AOAC procedures (1995). The crude protein content (N × 4.38) of the samples was estimated by the macro Kjeldahl method according to Léon-Guzmán et al. (1997); the crude fat was determined by extracting a known weight of powdered mushroom sample with petroleum ether, using a Soxhlet apparatus; the ash content was determined by incineration at 600 ± 15 °C. Total carbohydrates were calculated by difference method (100 − total moisture + total ash + total protein + total fat + total fibres) (AOAC, 1995). Total energy was calculated according to the following equations (Manzi et al., 1999)
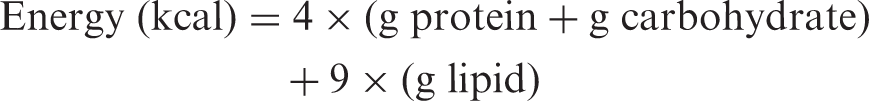
Trace element analysis
Fresh mushrooms, after removal of external material, were dried in an oven at 105 °C for 24 h after airdried for several days. Dried samples were homogenised, using an agate homogeniser, and stored in pre-cleaned polyethylene bottles until analysis. 1 g of sample was placed in a porcelain crucible and ashed at 450 °C for 20 h; then the ash was dissolved in 1 mL concentrated HNO3, evaporated to dryness, heated again at 450 °C for 4 h, treated with 1 mL concentrated H2SO4, 1 mL HNO3 and 1 mL H2O2, and then diluted with double deionised water up to a volume of 10 mL. The blank samples were treated in the same way (Colak et al., 2007). For the determination of metal contents, an ATI Unicam 929 model Atomic Absorption Spectrometer (AAS) was used. The determination of all metal contents was carried out in an air/acetylene flame. The maximum absorbance was obtained by adjusting the hallow cathode lamps at the operation conditions. All the experimental results were means ± SD of five parallel measurements.
Determination of fatty acid composition
Fatty acids were determined by gas–liquid chromatography with flame ionisation detection (GLC–FID)/capillary column based on the ISO 5509 (2000) trans-esterification method. The fatty acid profile was analysed with a Chrompack CP 9001 chromatograph (Chrompack, Middelburg, Netherlands) equipped with a split–splitless injector, a FID, and a Chrompack CP-9050 autosampler. The temperatures of the injector and detector were 250 °C. Separation was achieved on a 50 m × 0.25 mm i.d. fused silica capillary column coated with a 0.19 lm film of CP-Sil 88 (Chrompack). Helium was used as carrier gas at an internal pressure of 120 kPa. The column temperature was 140 °C, for a 5-min hold, and then programmed to increase to 220 °C at a rate of 4 °C/min and then held for 10 min The split ratio was 1:50, and the injected volume was 1.2 µL. The results are expressed in relative percentage of each fatty acid, calculated by internal normalisation of the chromatographic peak area. Fatty acid identification was made by comparing the relative retention times of FAME peaks from samples with standards. A Supelco (Bellefonte, PA) mixture of 37 FAMEs (standard 47885-U) was used. Some fatty acid isomers were identified with individual standardsalso purchased from Supelco.
Determination of amino acid composition
Amino acid contents in the four types of mushrooms were determined using high performance liquid chromatography (HPLC) under the following conditions (FAO, 1991):
The mobile phase consisted of three solvents:

A flow rate of 0.92 mL/min, a wave length of 266 nm, UV and a temperature of 30.2 °C were used in the analysis of amino acids. The standards were prepared as follows: 20 µL of standard amino acid solution (Sigma, AA-S-18, USA) were dissolved in 230 µL 0.1 N HCl, 750 µL Buffer, 1 mL 9-luorenyl methyl chloroformate and 2 mL n-pentane, then diluted to the concentration needed. However, a known weight of DL-methionine and L-cystine (Merck,Germany) were treated with a mixture of formic acid and hydrobromic acid to oxidate bound and free cysteine and methionine into cysteic acid and methionine sulphonic acid (Pellett and Young, 1980; Alofe, 1991). The samples were prepared by hydrolysing 500 mg in 100 mL 6.0 N HCl for 22–24 h at 110 °C according to the method of Henderson et al. (2000).
Determination of antioxidant properties of T. boudieri
Mushroom Extraction
For ethanol extraction, 5 g of air dried of T. boudieri sample (Yenice/Ceylanpinar/Sanliurfa) was weighed and ground into a fine powder in a mill, then mixed with 100 mL ethanol at room temperature at 150 r/min for 24 h. The residue was re-extracted under the same conditions until the extraction solvents became colourless. The extract obtained was filtered over Whatman no. 1 paper and the filtrate was collected, then ethanol was removed using a rotary evaporator (Laborata 4001, Heidolph WB, Germany, and Serial no. 069800367) at 40 °C to obtain dry extract. The extract was placed in a plastic bottle and then stored at 4 °C to prevent oxidative damage until analysis for nearly a week.
Total antioxidant activitiy by the β-carotene–linoleic acid method
In this assay, antioxidant capacity is determined by measuring the inhibition of the volatile organic compounds and the conjugated diene hydroperoxides arising from linoleic acid oxidation (Dapkevicius et al., 1998). A stock solution of β-carotene–linoleic acid mixture was prepared as following: 0.5 mg β-carotene was dissolved in 1 mL of chloroform (HPLC grade). Twenty-five microliters of linoleic acid and 200 mg Tween 40 were added. Chloroform was completely evaporated using a vacuum evaporator. Then 100 mL of oxygenated distilled water was added with vigorous shaking; 4.6 mL of this reaction mixture was dispersed to test tubes and 0.4 mL of various concentrations (0.5, 2.5, 5.0 and 10.0 mg/mL) of the extracts in ethanol were added and the emulsion system was incubated for up to 2 h at 50 °C. The same procedure was repeated with the positive control BHT, BHA, α-tocopherol, trolox and for blank sample. After this incubation period, absorbance of the mixtures was measured at 490 nm. Measurement of absorbance was continued until the color of β-carotene disappeared. The bleaching rate (R) of β-carotene was calculated according to the following equation
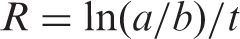
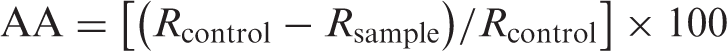
Antioxidative activities of the ethanolic extracts of T. boudieri were compared with BHT, BHA, α-tocopherol and trolox.
Determination of the reducing power
The reducing power of ethanolic extract of mushroom species was determined according to the method of Oyaizu (1986). Various concentrations of mushroom species extract (1, 2, 5, 10 mg/mL) in 1 mL of ethanol were mixed with phosphate buffer (2.5 mL, 0.2 M, pH 6.6) and 2.5 mL of 1% potassium ferricyanide [K3Fe(CN)6]. The mixture was incubated at 50 °C for 20 min, then 2.5 mL of trichloroacetic acid (10%) was added to the mixture, which was then centrifugated for 10 min at 1000 g (MSE Mistral 2000, UK, Serial no: S693/02/444). The upper layer of solution (2.5 mL) was mixed with distilled water (2.5 mL) and FeCl3 (0.5 mL, 0.1%), and the absorbance was measured at 700 nm. A higher absorbance indicates a higher reductive capability. BHT, BHA and α-tocopherol were used for comparison.
Superoxide anion radical scavenging activity
The superoxide anion radicals scavenging activity of ethanolic extract of T. boudieri was determined according to the method of Zhishen et al. (1999). All solutions were in 0.05M phosphate buffer (pH 7.8). The photo-induced reactions were performed in an aluminium foil-lined box with two 20W fluorescent lamps. The distance between reactant and lamp was adjusted until the intensity of illumination reached 4000 lx. The total volume of reactant was 5 mL and the concentrations of riboflavin, methionine and nitro blue tetrazolium (NBT) were 3 × 10−6, 1 × 10−2 and 1 × 10−4 M, respectively. The reactant was illuminated at 25 °C for 25 min. The photochemically reduced riboflavins generate superoxide anion radicals, thus reducing NBT to form blue formazan. The un-illuminated reaction mixture was used as a blank. Absorbance (A) was measured at 560 nm. Ethanolic extracts of mushroom and standards were added to the reaction mixture, in which superoxide anion radicals was scavenged, thereby inhibiting the NBT reduction. Absorbance (A1) was measured and the decrease in superoxide anion radicals was represented by A–A1. The degree of scavenging was calculated by the following equation:
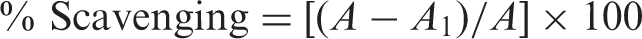
Free radical scavenging activity
Radical-scavenging ability was determined by a spectrophotometric method based on the reduction of a ethanol solution of DPPH using the method of Blois (1958). Each extract (0.1–10 mg/mL) in ethanol (1 mL) was mixed with 4 mL of ethanol solution containing DPPH radical. The mixture was shaken vigorously and left standing for 30 min in the dark. The absorbance was then measured at 517 nm against a blank by a spectrophotometer. Lower absorbance of the reaction mixture would indicate higher free radical scavenging activity. Inhibition of free radical, DPPH, in percent (%) was calculated according to formula
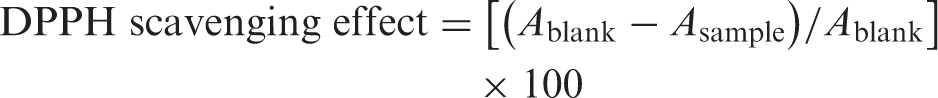
Chelating effect on ferrous ions
Chelating ability of ethanolic extracts from T. boudieri was determined according to the method of Dinis et al. (1994). Each extract (0.1–10 mg/mL) in ethanol (1 mL) was mixed with 3.7 mL of ethanol and 0.1 mL of 2 mmol/L ferrous chloride. The reaction was initiated by the addition of 0.2 mL of 5 mmol/L ferrozine. After 10 min at room temperature, the absorbance of the mixture was determined at 562 nm against blank. The results were expressed as percentage of inhibition of the ferrozine–Fe2+ complex formation. The percentage of inhibition of the ferrozine–Fe2+ complex formation was calculated using the formula given below
Determination of total phenolics
Total phenolic compounds in the ethanolic extract of T. boudieri were determined with Folin–Ciocalteau reagent according to the method of Slinkard and Singleton (1977) using gallic acid as a standard phenolic compound. Briefly, 1 mL of extract solution (containing 1 mg extracts) was transferred into a volumetric flask diluted with distilled water (46 mL); 1 mL of Folin–Ciocalteau reagent was added and the content of the flask mixed thoroughly. After 3 min, 3 mL of Na2CO3 (2%) was added, the mixture then was allowed to stand for 2 h with intermittent shaking. The absorbance was measured at 760 nm. The concentration of total phenolic compounds in the diluted extract determined as microgram of gallic acid equivalent by using an equation that was obtained from standard gallic acid graph (R2 = 0:995).
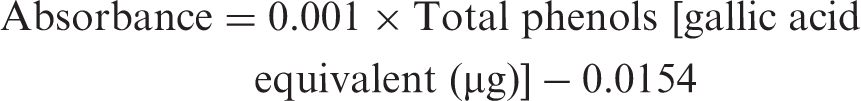
Statistical analysis
The data obtained were analysed by ANOVA and tests of significance were caried out using Duncan’s multiple range tests.
Results and discussion
Proximate composition and energetic value of T. boudieri
Chemical composition and energetic value of T. boudieri collected in different areas in Turkey
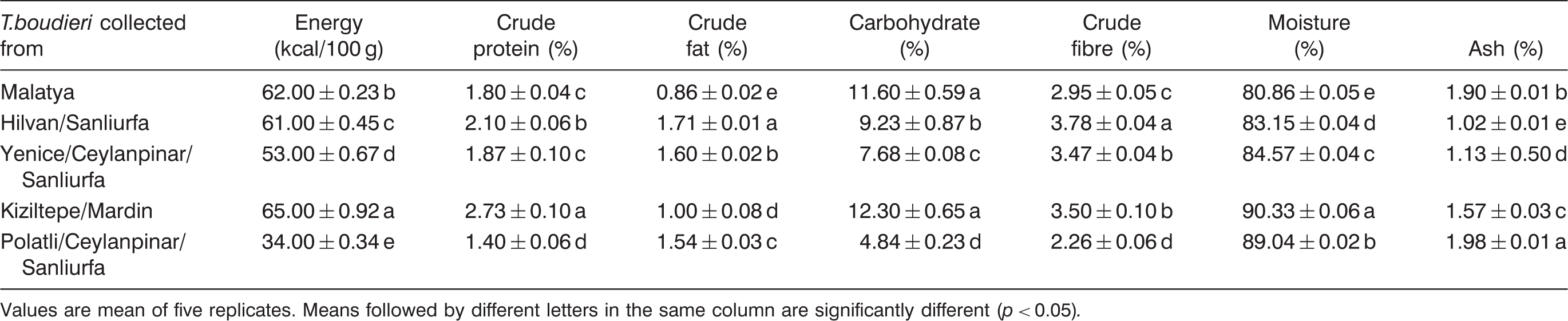
Values are mean of five replicates. Means followed by different letters in the same column are significantly different (p < 0.05).
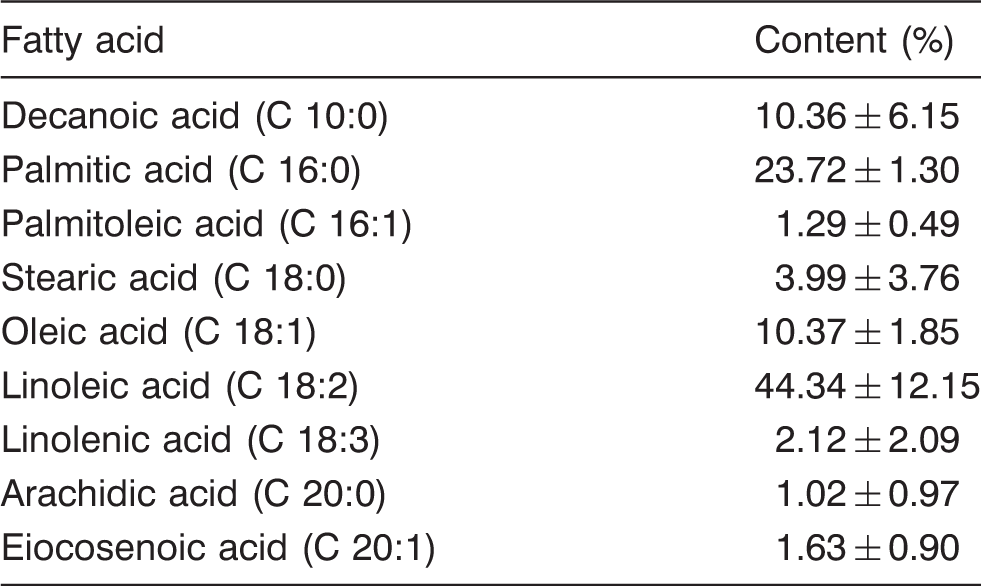
Fatty acid and mineral element content of T. boudieri
Fatty acids composition of T. boudieri from Kiziltepe/Mardin area (mean ± SD, n = 5)
Content of micro and macro elements of from Kiziltepe/Mardin area

Each value is expressed as mean ± SD of five replicate analyses.
Amino acid composition of T. boudieri
Amino acid content of T. boudieri collected in different areas from Turkey
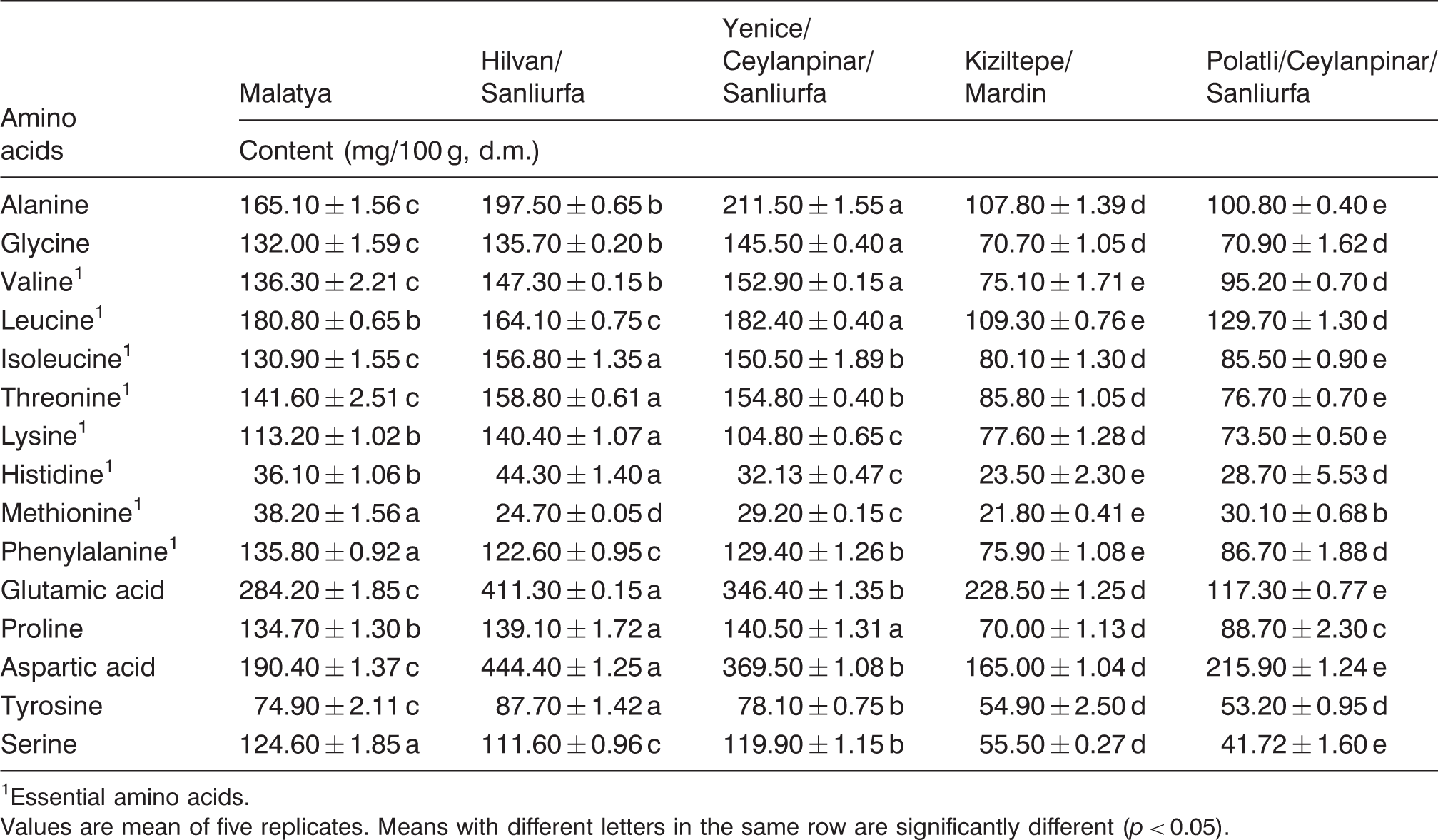
Essential amino acids.
Values are mean of five replicates. Means with different letters in the same row are significantly different (p < 0.05).
Total antioxidant activity of T. boudieri ethanolic extracts
Total antioxidant activity of the ethanolic extracts of T. boudieri and standards in β-carotene–linoleic acid test system
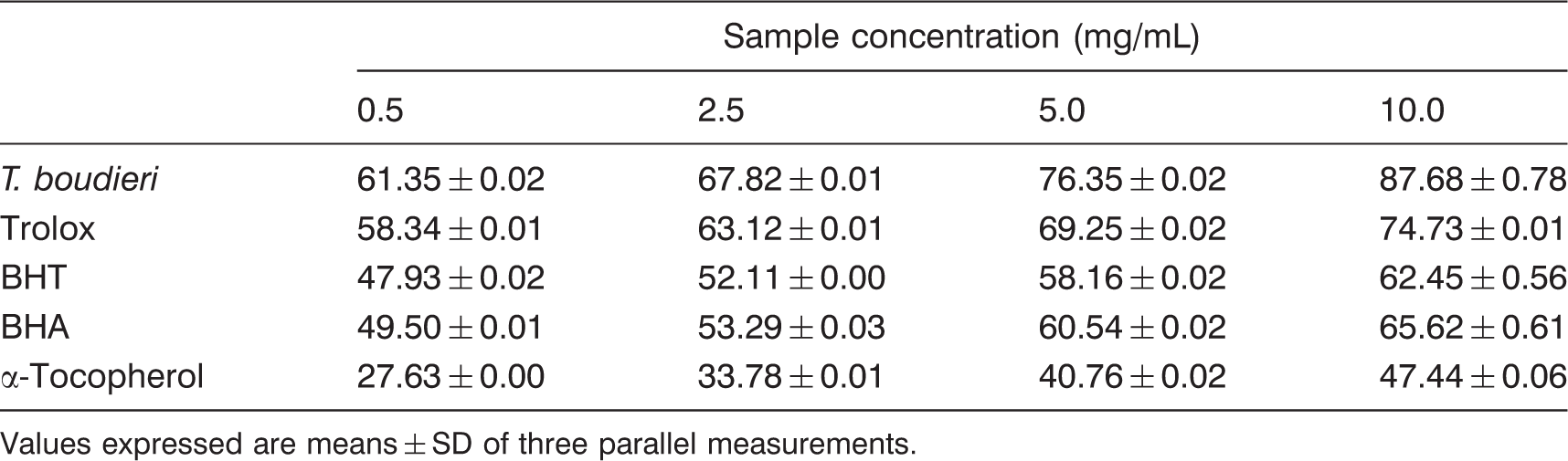
Values expressed are means ± SD of three parallel measurements.
Determination of reducing power of T. boudieri ethanolic extracts
Reducing power of T. boudieri and standards
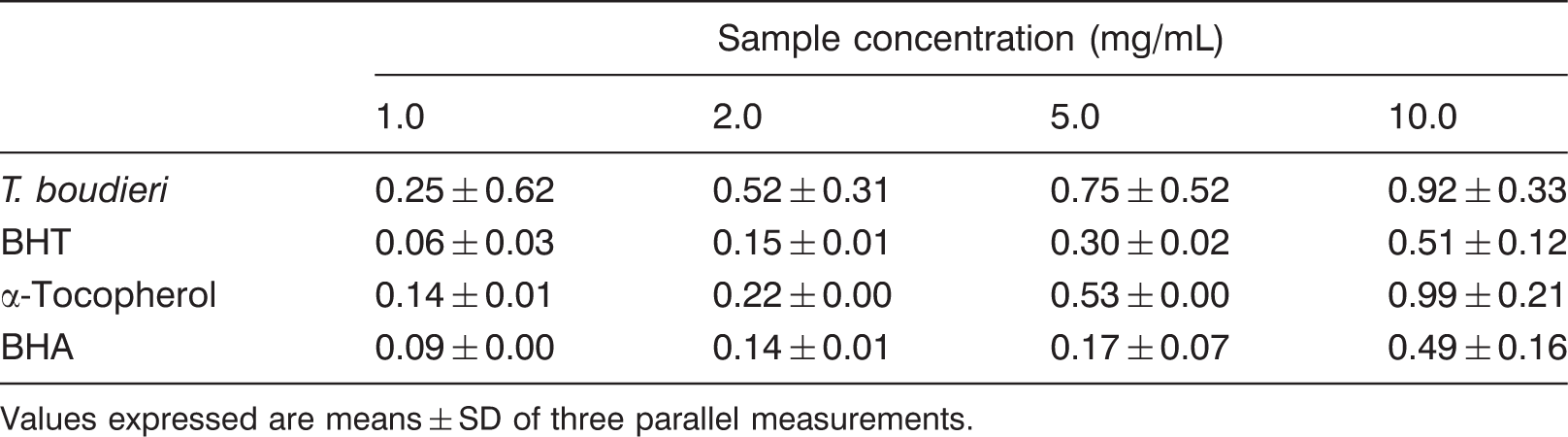
Values expressed are means ± SD of three parallel measurements.
Scavenging effect of T. boudieri ethanolic extracts on superoxide anion radical
Superoxide anion radical scavenging effect (%) of T. boudieri and standards
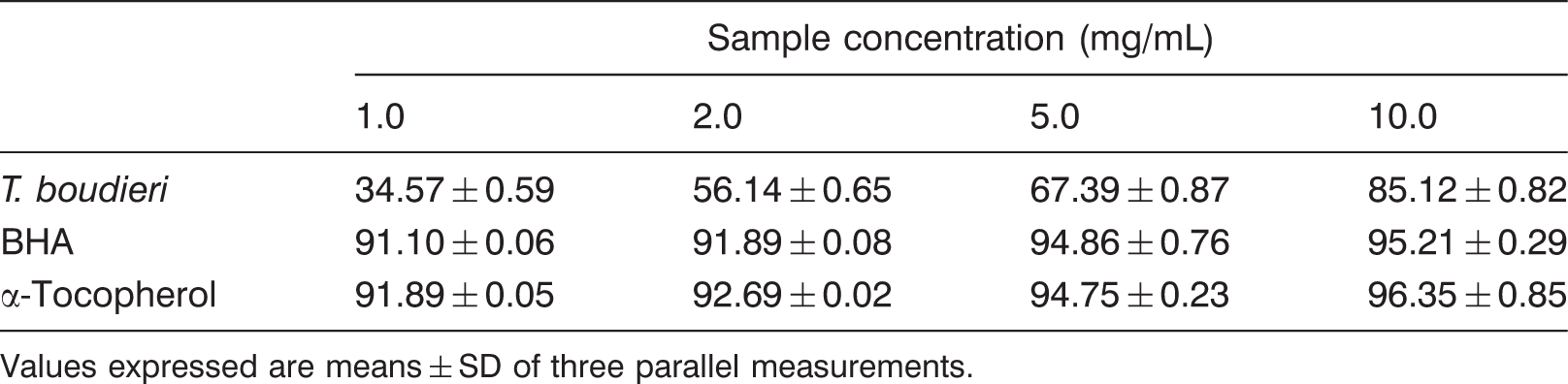
Values expressed are means ± SD of three parallel measurements.
DPPH scavenging activity of T. boudieri ethanolic extracts
Scavenging effect (%) of T. boudieri and standards on DPPH
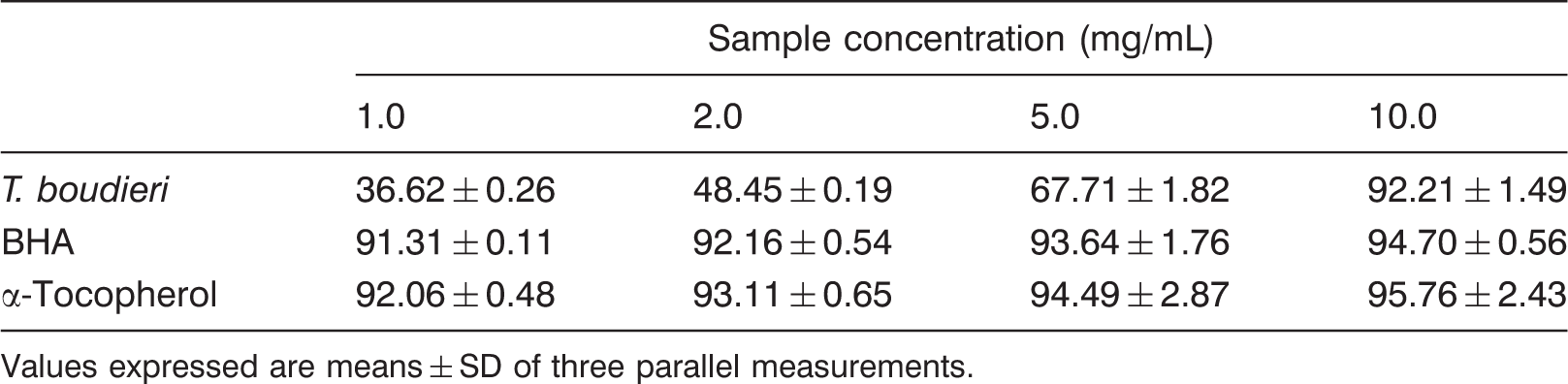
Values expressed are means ± SD of three parallel measurements.
Metal chelating activity of T. boudieri
Chelating effect (%) of T. boudieri and standards
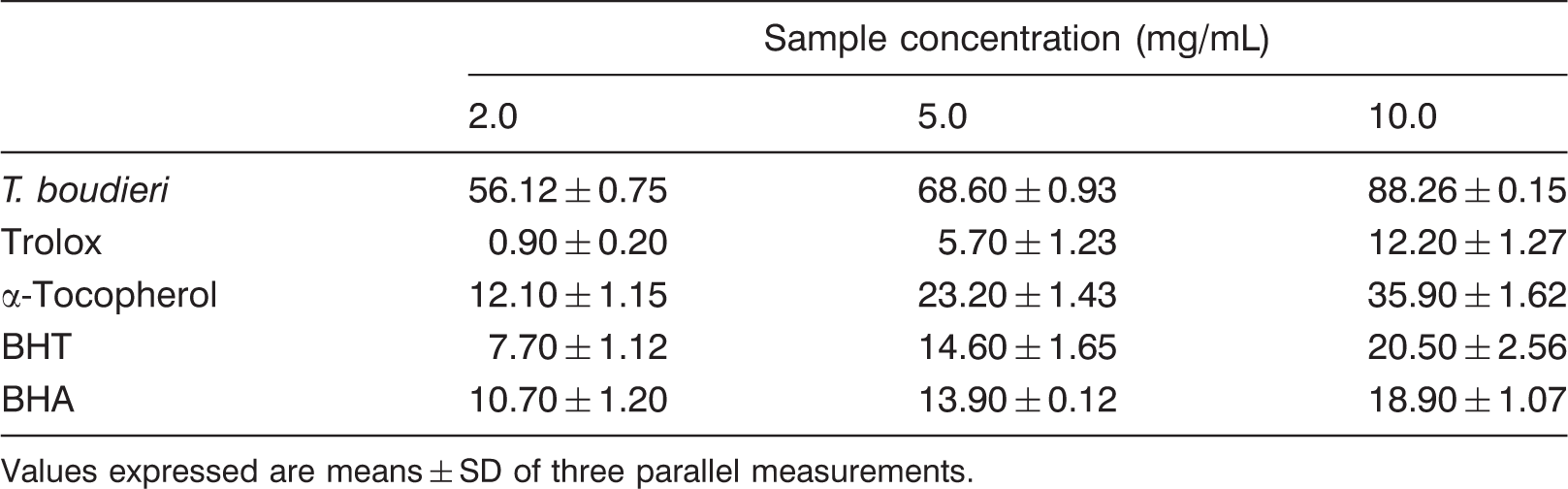
Values expressed are means ± SD of three parallel measurements.
Determination of total phenolic compounds of T. boudieri
The major natural antioxidant components found in the ethanolic extracts of T. boudieri shown was 8.45 ± 1.63 µg/mL (gallic acid equivalent/mL of extract). Phenols are important plant constituents because of their scavenging ability due to their hydroxyl groups (Hatano et al., 1989). The phenolic compounds may contribute directly to the antioxidative action (Duh et al., 1999). In addition, it was reported that phenolic compounds were associated with antioxidant activity and play an important role in stabilising lipid peroxidation (Yen et al., 1993). Polyphenols such as BHT and gallate were known to be effective antioxidants (Halliwell and Gutteridge, 2003). Numerous studies have conclusively showed that consumption of foods high in phenolic content can reduce the risk of heart disease by slowing the progression of atherosclerosis, because they act as antioxidants (Halliwell and Gutteridge, 2003; Kaur and Kapoor, 2002; Kahkonen et al., 1999). From the study we conclude that T. boudieri showed effective antioxidant activity when compared with standard antioxidant components.
Conclusions
In the present study, the antioxidant properties of ethanol extracts from T. boudieri were demonstrated by using a range of testing systems in vitro. According to the results of this study, it is clearly indicated that the ethanolic extracts of mushroom have significant antioxidant activity against various antioxidant systems. Moreover the mushroom can be used as an easily accessible source of natural antioxidants and as a possible food supplement or can be used in the pharmaceutical industry. On the other hand, according to the results obtained from the study the protein content in T. boudieri is significantly higher than most vegetables and fungi. Also the fruit body of mushroom is rich in fatty acid variety, diatery fibre with low fat contents. This situation provied to be excellent food that could be used as well-balanced diets.
Footnotes
Funding
This study was financially supported by TUBITAK/Turkey, (Project No: TOVAG-104O108) and Scientific Research Commission of Dicle University/Turkey, (Project no: 08-FF-05).