
Abstract
In this study, the effects of oligochitosan treatment on controlling postharvest diseases in Dongxue peach (Prunus Persica L. Batsch, cv Dongxuemi) were examined and the possible underlying mechanisms were discussed. Results showed that the disease incidence and lesion area in peach fruit inoculated with Monilinia fructicola and Penicillium expansum were all remarkably reduced by oligochitosan treatment. Oligochitosan treatment inhibited spore germination and mycelial growth of the two fungi in vitro. Oligochitosan treatment also induced upregulation of the salicylic acid signalling pathway-related genes (NPR1, PR1 and phenylalanine ammonia lyase) and enhanced the levels of total phenolics, flavonoids and lignin in peach. Meanwhile, enzymatic activities of superoxide dismutase, catalase, polyphenoloxidase, ascorbate peroxidase and phenylalanine ammonia lyase also increased. These findings suggest that the effects of oligochitosan on the disease control of peach fruit may be associated with its direct antimicrobial effects as well as increasing antioxidant, phenylpropanoid metabolism and accumulating antifungal compounds by activating the salicylic acid-dependent pathway.
Introduction
Dongxue peach (Prunus Persica L. Batsch, cv Dongxuemi) fruit is one of famous late-maturing peach cultivars in north areas of China. It is favoured for its high nutritional value and good taste. However, the fruit has a short postharvest life at room temperature due to its high susceptibility to pathogen infection. Brown rot and blue mould rot caused by Monilinia fructicola (G. Wint.) Honey and Penicillium expansum Link., respectively, are two major diseases on peach fruit (Yang et al., 2011; Zhou et al., 2008).Traditionally, synthetic fungicides are primarily used to control these diseases. However, to fulfil consumer’s demand on lesser use of chemicals on fruits, studies must focus on the development of natural compounds that can be utilised as alternative antimicrobials and preservatives (Zhang et al., 2015).
Inducing fruit diseases resistance with natural elicitors is an effective alternative. Oligochitosan, which is prepared by enzymatic hydrolysis of deacetylated chitosan polymers, has been confirmed to be natural elicitors. As it is evidenced that oligochitosan can elicit multiple plant defence responses against infection by pathogens or viruses in several growing crops (Yin et al., 2010). Besides, oligochitosans as fungicides are effective in inhibiting spore germination, germ tube elongation and mycelial growth of fungal phytopathogens, such as Alternaria alternata (Yan et al., 2011), M. fructicola (Yang et al., 2010, 2012), Alternaria kikuchiana Tanaka, Physalospora piricola Nose (Meng et al., 2010), Botrytis cinerea Pers (Badawy and Rabea, 2009). In recent years, many studies have shown that postharvest oligochitosan treatment (OT) can suppress multiple postharvest diseases affecting citrus (Chien et al., 2007; Deng et al., 2015), tomatoes (Badawy and Rabea, 2009), pears (Meng et al., 2010), apples (Yang et al., 2010), peaches (Ma et al., 2013) and jujubes (Yan et al., 2011) and successfully extend their storage life by inducing defence response. Therefore, oligochitosan, with its renewable, nontoxic, biocompatible and biodegradable characteristics, might provide a promising alternative to synthetic fungicides for controlling postharvest diseases of fruits.
With regard to the mechanism of oligochitosan-induced resistance in postharvest fruits, several studies have indicated that oligochitosan can enhance antioxidant and defence-related enzymes, such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), phenylalanine ammonia lyase (PAL), chitinase, β-1, 3-glucanase, as well as defence-related compounds, such as lignin, phytoalexins and phenolic secondary metabolites (Bautista-Baños et al., 2006; Deng et al., 2015; Ma et al., 2013; Meng et al., 2010; Yan et al., 2011). In addition, during the defence response, Ca2+, reactive oxygen species (ROS), jasmonic acid, salicylic acid (SA) and nitric oxide are involved in the oligochitosan-activated signalling pathway (Li et al., 2009; Lu et al., 2010; Obara et al., 2002; Zhang et al., 2007). However, the detailed defence responses involved in fruit resistance system still need further investigation.
The objectives of this study were (1) to investigate the effects of postharvest treatment with oligochitosan on controlling of brown rot caused by M. fructicola and blue mould rot caused by P. expansum in Dongxue peach fruit during storage at 25 ℃, (2) to evaluate the effects of oligochitosan on phenylpropanoid metabolism and antioxidant metabolism of Dongxue peach fruit, (3) to determine the differential expression of SA signalling pathway-related genes (NPR1, PAL, PR1) in oligochitosan treated and untreated peach fruit.
Materials and methods
Materials
Dongxue peach (Prunus Persica L. Batsch, cv Dongxuemi) fruit were harvested at the mature stage and sorted based on the size and the absence of physical injuries or disease infection. Before treatments, fruit were disinfected on the surface with 2% (v/v) sodium hypochlorite for 2 min, then rinsed with tap water and air-dried (Liu et al., 2007).
Oligochitosan (minimum 90% deacetylated, molecular weight < 1 kDa) was obtained from Dalian Institute of Chemical Physics, Chinese Academy of Sciences (Dalian, China). It was dissolved in sterile distilled water at 5 g l−1.
Pathogen inoculum preparation
M. fructicola and P. expansum were isolated from infected peach fruit and maintained on potato dextrose agar (PDA) at 4 ℃. In order to reactivate the culture and verify its ability to cause decay, M. fructicola and P. expansum were separately inoculated into wounds of peach fruit and re-isolated onto PDA once an infection was established. Spore suspension was obtained by flooding two-week-old cultures with sterile distilled water containing 0.5 ml l−1 Tween80. The number of spores was calculated with a haemocytometer and the desired spore concentration was adjusted with sterile distilled water.
Spore germination and mycelial growth of M. fructicola and P. expansum
The effect of oligochitosan on spore germination and mycelial growth of M. fructicola and P. expansum were assayed according to Yang et al. (2012). Aliquots of 50 ml conidial suspensions at 1 × 106 spores ml−1 were plated on Petri dishes (60 mm in diameter) with 10 ml PDA which contained oligochitosan (final concentration of 5 g l−1) and the Petri dishes incubated at 25 ℃ for 12 h. Approximately 200 spores of each pathogen were measured for germination rate. The mycelial disks (7 mm in diameter) from two-week-old cultures of the fungi were placed in the centre of Petri dishes (90 mm in diameter) with 20 ml of PDA containing 5 g l−1 of oligochitosan, and then incubated at 25 ℃. Mycelial growth was determined by measuring colony diameter at 3 d after inoculation. The inhibitory rate on mycelial growth was calculated according to the following formula: Inhibitory rate (%) = 100 × (colony diameter of the control-colony diameter of the treatment) / (colony diameter of the control−7 mm).
Effect of OT on control of brown rot and blue mould rot in peach fruit
In order to investigate the action of oligochitosan on control of brown rot and blue mould rot caused by M. fructicola and P. expansum, respectively, we applied three treatment methods.
Method I: OT of M. fructicola or P. expansum followed by fruit inoculation. Two wounds (4 mm deep and 3 mm wide) were made with a sterile nail on the equator of each peach fruit. Ten microlitre oligochitosan-treated (5 min) spore suspension (1 × 104 spores ml−1 containing 5 g l−1 of oligochitosan) was administered into each wound. Water-treated spore suspension was used as a control.
Method II: OT was applied to fruit followed by inoculation of wounds with M. fructicola or P. expansum spore suspension. Fruits were immersed in oligochitosan solutions (5 g l−1) for 5 min and air-dried for 2 h. Fruits immersed in water for 5 min served as a control. Two wounds (4 mm deep and 3 mm wide) were made on the equator of each fruit with a sterile nail and each wound was inoculated with 10 µl of M. fructicola or P. expansum spore suspension (1 × 104 spores ml−1).
Method III: Inoculation of fruit with M. fructicola or P. expansum spore suspension followed by the application of an OT to the inoculated fruit. Two wounds (4 mm deep and 3 mm wide) were made on the equator of each fruit with a sterile nail and each wound was inoculated with 10 µl of M. fructicola or P. expansum spore suspension (1 × 104 spores ml−1) and air-dried for 2 h; they were subjected to oligochitosan solutions (5 g l−1) or water (control) immersion for 5 min.
After each treatment, fruits were placed in a covered plastic food tray, and each tray was enclosed with a polyethylene bag and stored at 25 ℃. Disease incidence and lesion diameter of peach fruit were determined after 3 d. Each treatment contained three replicates with 10 fruit per replicate, and the experiment was repeated three times. Incidence represented the percentage of fruit displaying rot, while lesion diameter was measured only on those wounds that were infected (Liu et al., 2012).
Sample collecting
The fruits treated with OT without wounding or dipping in M. fructicola or P. expansum spore suspension were used for evaluation of fruit response. Three and five grams of peach tissues were excised from each fruit every day and packed with aluminium foil individually. The excised tissues were rapidly frozen in liquid nitrogen and kept at −80 ℃ until biochemical analysis. Three replicates per treatment were made, each replicate containing 16 fruits and the entire experiment was performed twice.
Measurement of malondialdehyde (MDA) content
Three grams of frozen tissue was homogenised with a mortar and pestle in 5 ml of 10% trichloroacetic acid (w/v) and centrifuged at 12,000×g for 20 min at 4 ℃. The supernatant was used for MDA determination according to Ren et al. (2012). MDA formation was determined by measuring the absorbance (UV-2450, SHIMADZU, Japan) of the reaction product of MDA with 2 ml of 0.67% 2-thiobarbituric acid (TBA) in 1 ml of the supernatant at 532 nm, corrected for non-specific turbidity by subtracting the absorbance at 600 nm and interference generated by TBA–sucrose complexes at 450 nm. The MDA concentration was expressed as nmol g−1 FW (fresh weight).
Measurement of total phenolics, flavonoids and lignin contents
Total phenolics and flavonoids were detected according to the methods by Yin et al. (2012) with some modifications. Three grams of frozen tissue was homogenised with 5 ml of 1% HCl–methanol ice-cold solution, extracted for 2 h and then centrifuged at 4 ℃ for 30 min at 12,000×g. The supernatant was collected and absorbance measured at 280 and 325 nm for total phenolics and flavonoids, respectively. The contents of total phenolics and flavonoids were determined from calibration curve prepared with standard gallic acid and rutin solution, respectively, and expressed as mg g−1 FW.
Lignin was assayed according to the method proposed by Yin et al. (2012). Three grams of frozen tissue was homogenised with 5 ml of 95% ice cold ethanol solution and then centrifuged at 4 ℃ for 20 min at 14,000×g. The collected pellet was washed with 95% ethanol thrice, and then washed thrice with ethanol/hexane = 1:2 (v/v). Pellets were collected for drying (24 h at 60 ℃) and transferred to tubes, and then 1 ml of 25% bromised acetyl-acetic acid was added. The tubes were incubated in a water bath at 70 ℃ for 30 min, and then 1 ml of 2 mol l−1 NaOH was added to stop the reaction. Two millilitres of ice acetic acid and 0.1 ml of 7.5 mol l−1 hydroxylamine hydrochloric acid were added to each tube and centrifuged. Supernatant solution (0.5 ml) was removed. The absorbance of the supernatant was measured at 280 nm with a spectrophotometer. Lignin compounds were expressed as OD280 g−1 FW.
Measurement of enzyme activities in peach fruit
Enzyme extraction: all enzyme extracts were conducted by homogenising 5.0 g of frozen tissue in homogeniser (T-25, IKA, Germany) on ice, using the following extraction media: 20 ml of 0.2 mol l−1 phosphate buffer (pH 6.4) containing 0.1 g polyvinylpolypyrrolidone (PVPP) for POD, polyphenoloxidase (PPO), CAT and SOD. Twenty millilitres of 0.1 mol l−1 phosphate buffer (pH 7.5) containing 1 mmol l−1 ethylene diamine tetraacetic acid (EDTA) and 3 mmol l−1 ascorbic acid for ascorbate peroxidase (APX). Twenty millilitres of 0.1 mol l−1 sodium borate buffer (pH 8.7) containing 0.037% (EDTA) (w/v), 0.137% β-mercaptoethanol (v/v) and 3% PVPP (w/v) for PAL. The extracts were then centrifuged at 12,000×g for 30 min at 4 ℃.
SOD activity was determined by the method of Ren et al. (2012). Reaction mixture contained 1.5 ml of 50 mmol l−1 phosphate buffer (pH 7.8), 0.3 ml of 130 mmol l−1 methionine, 100 µmol l−1 EDTA–Na2, 750 µmol−1 nitro blue tetrazolium (NBT), 0.4 ml of crude enzyme extract and 0.3 ml of 20 µmol l−1 riboflavin. The formation of blue formazan was monitored by taking the absorbance at 560 nm. One unit of SOD activity was defined as the amount of enzyme that caused a 50% inhibition of NBT and the SOD activity was expressed as U g−1 FW.
CAT activity was assayed according to Ren et al. (2012) by monitoring the disappearance of H2O2 by recording the decrease in absorbance at 240 nm of a reaction mixture containing 50 mmol l−1 phosphate buffer (pH 7.5), 20 mmol l−1 H2O2 and 100 µl of crude enzyme extract. The CAT activity was expressed as ΔOD240 min−1 g−1 FW.
PPO activity was examined with the method reported by Jiang et al. (2002). The assay mixture contained 0.1 ml of enzyme extract, 2 ml of sodium phosphate buffer (100 mmol l−1, pH 6.4) and 1 ml of catechol (65 mmol l−1). Absorbance was measured at 420 nm. The activity was expressed as ΔOD420 min−1 g−1 FW.
APX activity was determined as described by the method of Ren et al. (2012). The assay mixture consisted of 2 ml of 100 mmol l−1 phosphate buffer (pH 7.5), 800 µl of 3 mmol l−1 ascorbic acid, 500 µl of 30% H2O2 (v/v) and 200 µl of crude enzyme extract. The APX activity was expressed as ΔOD290 min−1 g−1 FW.
PAL activity was assayed by using a method modified from Yin et al. (2012). One millilitre of enzyme extract was incubated with 2 ml of borate buffer (50 mmol l−1, pH 8.8) and 1 ml of
The assay of POD was conducted following the method of Meng et al. (2008). 0.5 ml of enzyme in 2 ml buffer substrate (100 mmol l−1 sodium phosphate, pH 6.4 and 8 mmol l−1 guaiacol) was incubated for 5 min at 30 ℃ and the increase in absorbance at 460 nm for 2 min after adding 1 ml of H2O2 (24 mmol l−1) was measured. Specific enzyme activity was defined as ΔOD460 min−1 g−1 FW.
Gene expression analysis by real-time quantitative PCR
Total RNA from OT and control peach samples was isolated after 12 h of oligochitosan or water treatment following a cetyltrimethylammonium bromide-based protocol (Yu et al., 2014). First-strand cDNA was synthesised from 500 ng of total DNA-free RNA with a PrimeScript® RT reagent Kit (TaKaRa, Japan) according to the manufacturer’s instructions. The cDNA was diluted 20-fold and 2 µl of the diluted cDNA was used as the template for real-time quantitative PCR analysis.
The specific genes were amplified using gene-specific primers as previously described (Liu et al., 2012; Wang et al., 2013).The sequences of primers used were as follows: PpNPR1 (DQ149935), forward 5′-GACCCAAACATGCCAGCAGTG-3′ and reverse 5′-ATCCTTCGGCCTTGTCAACCT-3′; PpPR1 (AF362989), forward 5′-ATCAACTGGGACTTGCGTACT-3′ and reverse 5′-TAGTCGCCACAGTCAACAAA G-3′; PpPAL (AF206634), forward 5′-TTGACCGCGAGTACGTTTT-3′ and reverse 5′-CTGTTTGGGGTTGCTGATT-3′; Ppα-tubulin (DY650410), forward 5′-CAGATGCCCAGTGATGCCTCAG-3′ and reverse 5′-ACCAGTACCACCACCAACAGC-3′. PCR reactions were carried out in a 48-well system (20 µl per well). The reaction mixture contained 10.0 µl of SYBR® Premix Ex Taq™ (TaKaRa, Japan), 0.4 µl of each primer (10 µmol l−1), 0.4 µl ROX Reference Dye (50 ℃), 2 µl of cDNA and 6.8 µl of RNase-free water. The reactions were performed on a Step One Real-Time PCR System (Applied Biosystems) at 95 ℃ for 30 s, followed by 40 cycles at 95 ℃ for 5 s and at 60 ℃ for 30 s. After each run, a dissociation curve was created to confirm the specificity of the product and to avoid the production of primer dimers. α-tubulin was used as a housekeeping gene. Calculation of the relative quantification was performed by the comparative 2−ΔΔCT method (Livak and Schmittgen, 2001). All reactions were performed in triplicate and each sample was further amplified without reverse transcription to avoid DNA contamination of the sample.
Statistical analysis
All the data were presented as the mean ± standard errors. Statistical analyses were performed by using SPSS software (Version 14.0; SPSS, Chicago, IL).
Results
Effect of oligochitosan on spore germination and mycelial growth of M. fructicola and P. expansum
After 12 h of incubation on PDA at 25 ℃, the oligochitosan-treated (5 g l−1) M. fructicola and P. expansum spores exhibited a significantly (P < 0.05) lower level of germination as compared to the water-treated control. The germination rates in water-treated control reached 95%, while that in oligochitosan-treated M. fructicola and P. expansum spores were only 47.6 and 54.6%, respectively (data were not shown). Likewise, a similar pattern was observed for mycelial growth, with the inhibitory rate of 67.5 and 57.6% for M. fructicola and P. expansum, respectively (data were not shown). The results indicated that oligochitosan could directly inhibit the growth of the two fungi in vitro and M. fructicola was more sensitive to oligochitosan as compared to P. expansum.
Efficacy of OT on control of brown rot and blue mould rot in peach fruit
Three different methods of OT were employed: (I) spores were oligochitosan treated and inoculated in wounds of untreated fruits, (II) fruits were oligochitosan treated and untreated spores were applied to wounds and (III) untreated spores were inoculated in fruit wounds and then the fruits were treated by oligochitosan. Among the three treatments, method III gave the best level of control of brown rot and blue mould rot. Disease incidence reached 100% in the control fruit after 3 d storage at 25 ℃. At that time, incidence for the three treatments (I, II and III) was 50, 65 and 42%, respectively, for infecting by M. fructicola and 58, 67 and 46%, respectively, for infecting by P. expansum (Figure 1). The lesion diameter was also significantly reduced by the three treatments (P < 0.05), the largest reduction was obtained by OT in Method III for both M. fructicola and P. expansum (Figure 1).
Effect of oligochitosan on disease incidence and lesion diameter of peach fruit caused by M. fructicola and P. expansum after 3 d storage at 25℃.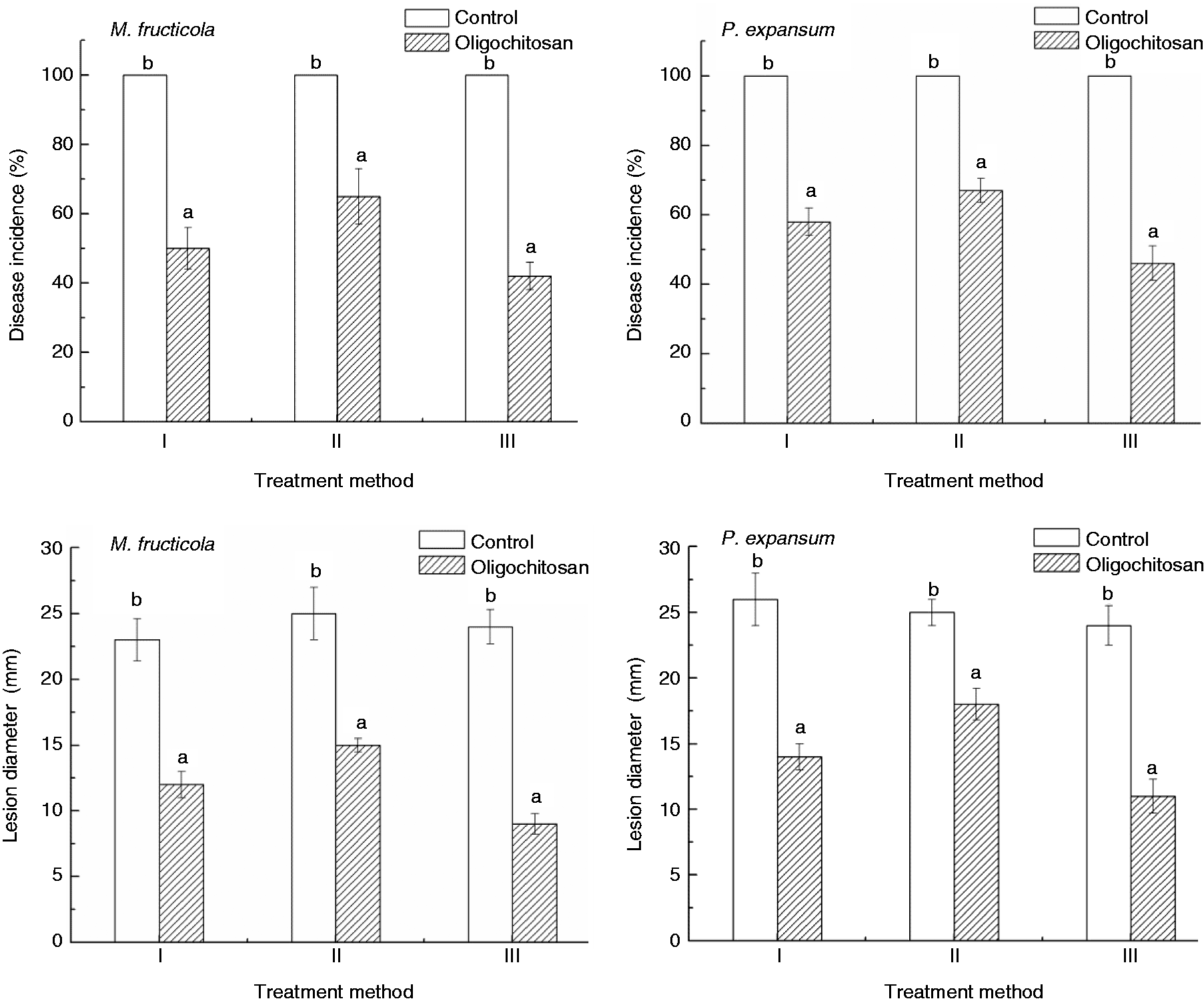
Effect of OT on MDA content
MDA content of peach fruit increased quickly during storage at 25 ℃, but OT (5 g l−1 for 5 min) significantly (P < 0.05) inhibited the increase (Figure 2). The maximum difference appeared at 3 d, showing a 63.9% lower MDA content in OT peach fruit than that in the control.
Effect of oligochitosan treatment (5 g l−1 for 5 min) on MDA content in peach fruit during storage at 25℃.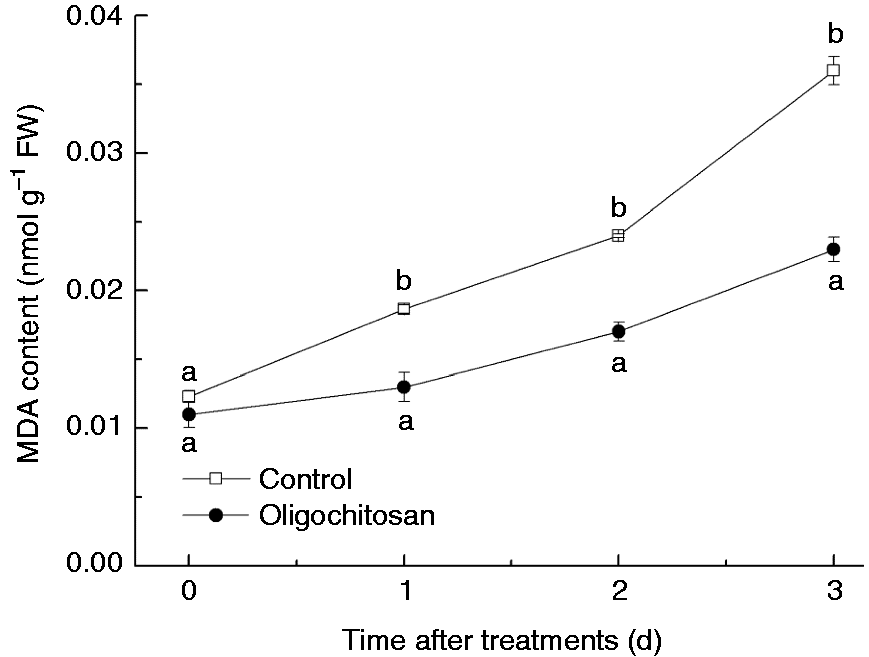
Effect of OT on total phenolics, flavonoids and lignin contents
The contents of total phenolics (Figure 3(a)) and flavonoids (Figure 3(b)) in the OT (5 g l−1 for 5 min) peach fruit showed rapid increases during the first day storage and decrease afterwards. Both total phenolics and flavonoids contents in OT peach fruit were significantly higher at days 1 and 2 (P < 0.05), which was approximately 1.3- to 1.5-fold than that in the water control. The content of lignin in OT peach fruit increased at the beginning of storage time and reached maximum activity at second day and then declined (Figure 3(c)). In comparison to the control, the lignin content in the OT peach fruit was higher at days 2 and 3, approximately 1.4- and 1.5-fold than that in the water control, respectively.
Effect of oligochitosan treatment (5 g l−1 for 5 min) on total phenolics (a), flavonoids (b) and lignin (c) contents in peach fruit during storage at 25℃.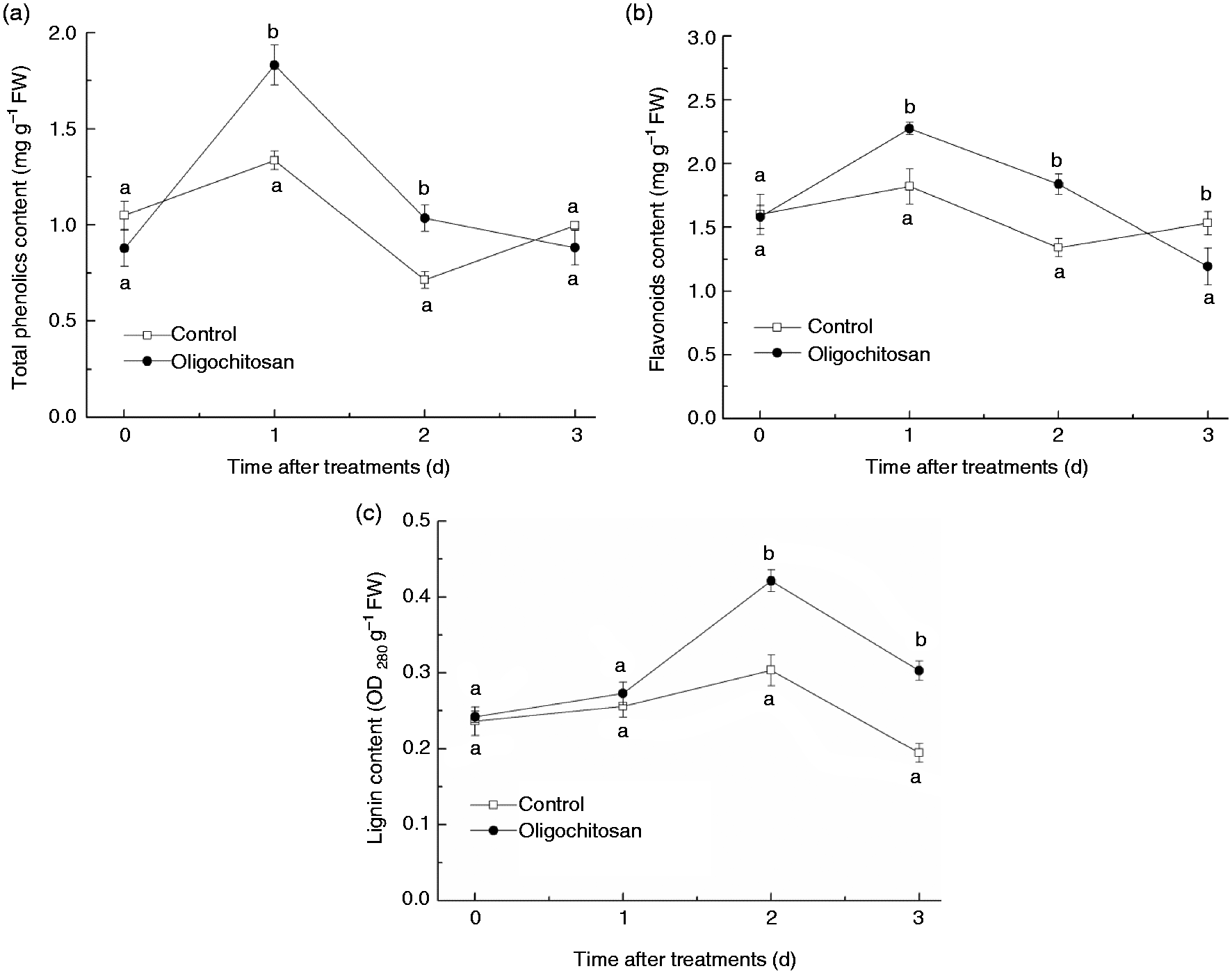
Effect of OT on defence-related enzyme activities
The changes in CAT, SOD, PPO, APX, PAL and POD activities in response to OT (5 g l−1 for 5 min) were shown in Figure 4. During the storage at 1–3 d, CAT and SOD activity (Figure 4(a) and (b), respectively) in peach fruit under treatment with oligochitosan remained a stable higher level than the water control, especially on the third day, approximately 4.5- and 1.8-fold than that of the control for CAT and SOD activity, respectively. The PPO activity (Figure 4(c)) in OT peach fruit was 2.3- and 6.9-fold than that of the control at 0 and 1 d, respectively. Then the activity of PPO in OT peach fruit began to decrease and no significant difference was found between the treatment and the control after 2 d storage (P > 0.05). In comparison to the control, the APX activity (Figure 4(d)) in OT peach fruit was significantly higher at days 2 and 3 (P < 0.05), which was approximately 2.9- and 1.3-fold than that of the water control, respectively. The PAL activity (Figure 4(e)) increased in OT peach fruit in the first 2 d of storage and was 3.3-fold than that of the water control at 2 d. Then the activity of PAL in OT peach fruit began to decrease along with the extension of storage time, which dropped significantly below the control at 3 d (P < 0.05). However, the effect of OT on the POD activity (Figure 4(f)) in peach fruit followed a different pattern from the other five enzymes. There was no significant effect on POD activity between the control and OT peach fruit during storage (P > 0.05).
Effect of oligochitosan treatment (5 g l−1 for 5 min) on CAT (a), SOD (b), PPO (c), APX (d), PAL (e) and POD (f) activity in peach fruit during storage at 25℃.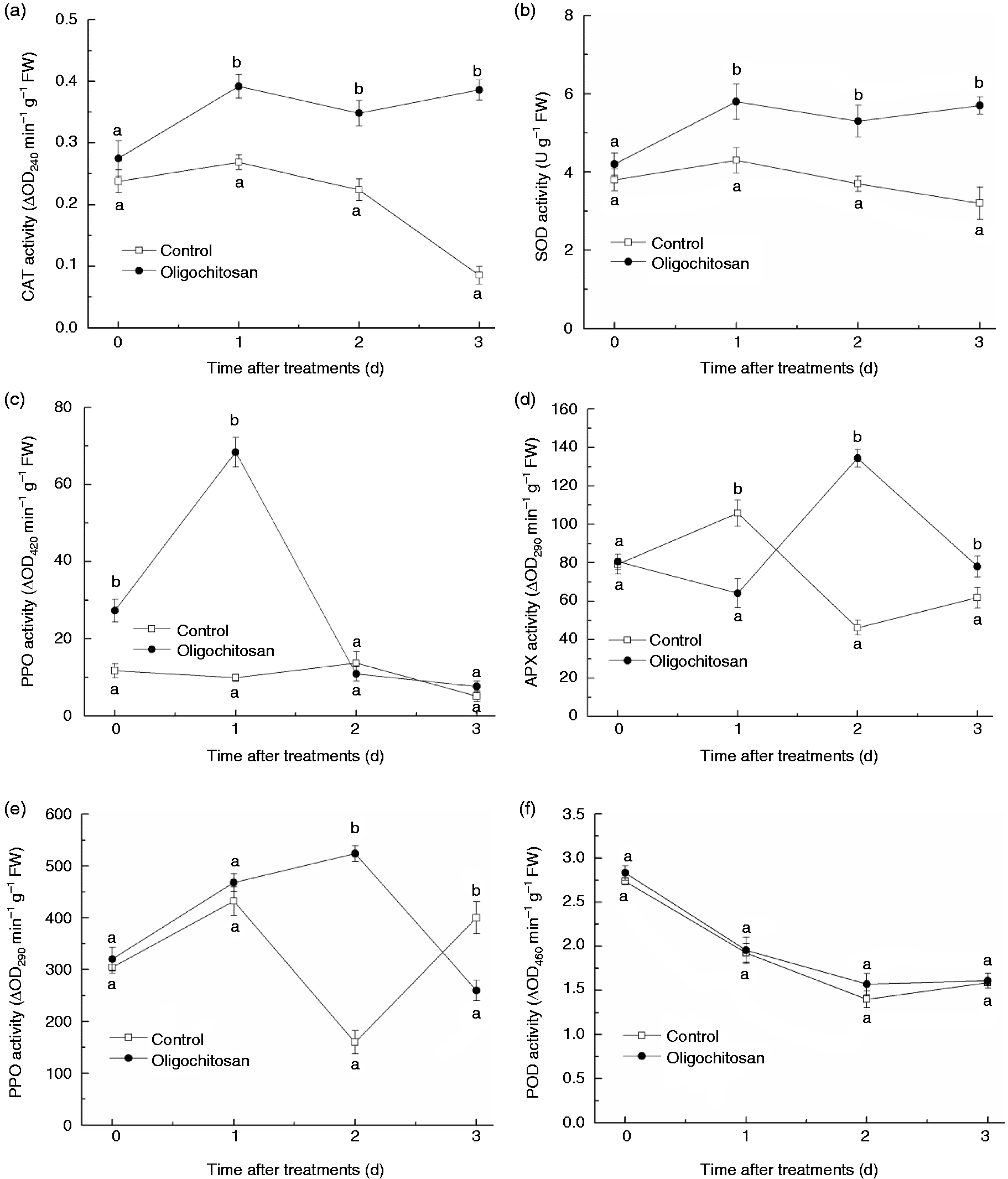
Effect of OT on gene expression of SA signalling pathway-related genes
The expression of SA signalling pathway-related genes (NPR1, PR1 and PAL) was analysed by qRT-PCR in OT (5 g l−1 for 5 min) and control (water for 5 min) peach fruit. As shown in Figure 5, 12 h after OT, NPR1, PR1 and PAL increased 5.8, 3.9 and 3.6 times as compared to the untreated samples, respectively.
Relative expression levels of SA signalling pathway-related genes (NPR1, PR1 and PAL) in oligochitosan treated and untreated peach tissues.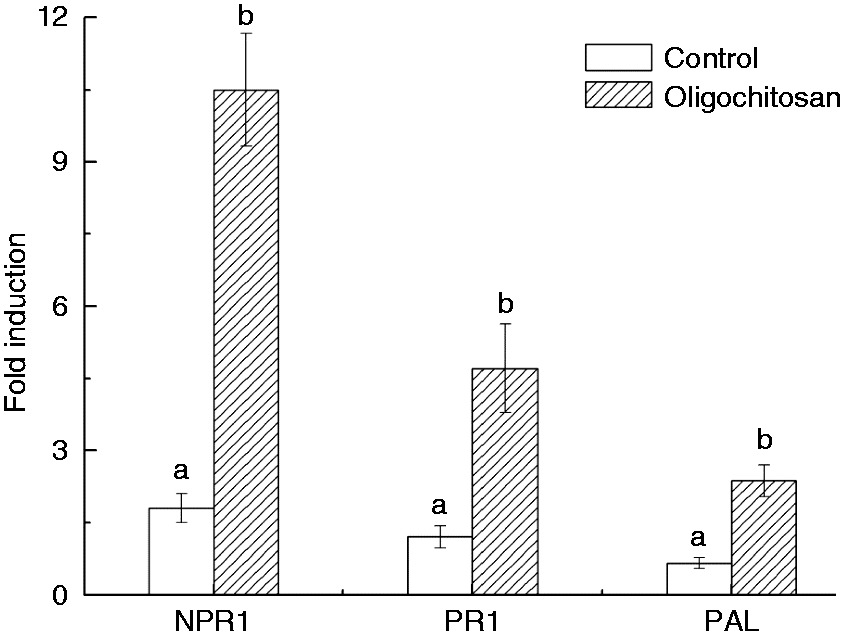
Discussion
OT of fruits has been reported to be an effective method of managing postharvest diseases by both its inhibitory or germicidal effect on decay pathogens and eliciting defence mechanisms in plant host (Deng et al., 2015; Ma et al., 2013; Meng et al., 2010; Yan et al., 2011). In the present study, it was found that OT directly inhibited both spore germination and mycelial growth of M. fructicola and P. expansum in vitro. The results confirmed previous findings about the effect of OT on M. fructicola reported by Yang et al. (2010, 2012). In order to investigate whether the mode of action of OT on control of brown rot and blue mould rot involved an effect on both the pathogen and the peach, we used three different methods of OT. Data indicated that oligochitosan treating of the fruit after it was inoculated with the pathogen provided the best control. This suggests that the mode of action of oligochitosan on control of fungal diseases involved an effect on both pathogen and the host. Oligochitosan directly inhibited the growth of M. fructicola and P. expansum and was also able to induce host defence mechanisms in the fruit itself.
Antioxidant enzymes, such as SOD, CAT, POD and APX, are associated with improving disease resistance in fruits by regulating metabolism of ROS. MDA, a product of lipid peroxidation, can reflect the extent of lipid peroxidation induced by oxidative stress (Zhang et al., 2017). A higher level of SOD, CAT, POD and APX activities and a lower level of MDA content are equal to a more effective antioxidant defence system and closely associated with delayed senescence in harvested fruit (Cai et al., 2015). Preharvest oligochitosan sprays enhanced the activities of SOD and APX in jujube (Zizyphus jujuba Mill. cv. Dongzao) fruit against M. fructicola and A. alternata, which were related to enhanced disease resistance (Yan et al., 2012). Deng et al. (2015) also found that the enzymatic activities of SOD, CAT, POD, APX increased in citrus fruit peels by OT, all of which were correlated with the onset of induced disease resistance. In the present study, we found that OT effectively (P < 0.05) enhanced activities of CAT, APX, SOD and inhibited the increase of MDA in peach fruit during storage at 25 ℃. Thus, it was speculated that postharvest application of oligochitosan induced the defence mechanisms which could be associated with increasing the antioxidant metabolism of peach fruit.
Meanwhile, phenylpropanoid metabolism has been also associated with disease resistance as reported by Cai et al. (2015) and Yuan et al. (2013). PAL is the key enzymes in the activation of systems, which catalyses the conversion of phenylalanine to trans-cinnamic acid, a key intermediate in the synthesis of SA. Moreover, PAL plays important roles in biosynthesis of phenolics, phytoalexins and lignin, the key factors responsible for disease resistance (Joe et al., 2012). Yan et al. (2011) reported that the PAL activity of Chinese jujube was stimulated by OT. PPO and POD are the terminal enzymes in the phenylpropanoid metabolism. PPO is involved in the lignification of plant cells and the oxidation of polyphenols into quinines which can restrict the growth of the plant pathogen (Mohammadi and Kazemi, 2002). POD participates in the cell wall-building processes, such as oxidation of phenols, suberisation and lignification in the cross-linking of cell wall protein during the defence reaction against pathogenic agents (Brisson et al., 1994). Although OT had no effect on POD activity, it could enhance the PPO and PAL activities in peach fruit, which could result in disease resistance.
In addition, accumulation of antifungal compounds is thought to be another important mechanism of resistance to various diseases. Numerous researches have shown that a high level of phenolic compounds can restrict or slow the growth of the pathogen (Mayr et al., 1997), flavonoid compounds can directly kill the fungal pathogen (Shadle et al., 2003) and the deposition of lignin can increase the resistance of plant cell walls against pathogens and limit pathogenic bacteria access to water and nutrients from the host (Deepak et al., 2007; Tronchet et al., 2010; Yuan et al., 2013). In this work, levels of total phenolics, flavonoids and lignin contents in peach fruit were also significantly enhanced by OT. The results suggest these changes might have a role in enhancing disease resistance in OT peach fruit.
Plant defence responses are regulated by a complex network of signalling pathways, which are dependent on the plant hormones (Yang et al., 2017). Multitude of studies has reported that SA is essential for the activation of systemic acquired resistance (Kumar, 2014) and involved in oligochitosan activated signal pathway (Obara et al., 2002). PAL is the rate-limiting enzyme involved in SA biosynthesis (Métraux, 2002). NPR1 is an important regulatory that acts downstream of SA and induces the expression of PR genes (An and Mou, 2011). Particularly, PR1 proteins are often used as markers of the SA-dependent resistance. Sun et al. (2013) reported that burdock fructooligosaccharide treatment induced the upregulation of PAL, NPR1 and PR1 genes in postharvest Kyoho grapes, which indicated that the SA-dependent signalling pathway was induced. In our experiment, the treatment of peach fruit with oligochitosan increased the transcript levels of several SA signalling pathway-related genes (PAL, NPR1 and PR1 were upregulated 3.6-, 5.8- and 3.9-fold, respectively), suggesting OT-induced disease resistance partly depends on the SA signalling pathway. However, hormone-mediated signalling pathways are known to crosstalk resulting in an antagonistic or synergistic interaction. Therefore, further research should be done in order to explore how SA interacts with other hormone.
Conclusions
The present study showed that OT could have a significant impact on postharvest diseases control in Dongxue peach fruit. The control effect by OT was associated with the direct antimicrobial effects on pathogen propagules and induction of defence mechanisms in peach fruit. The defence mechanism might be closely related to its ability to increase antioxidant and phenylpropanoid metabolism and accumulate antifungal compounds by activating the SA-dependent pathway. However, the detailed modes of its molecular mechanisms need to be elucidated further.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Key R & D Program of China (No. 2016YFD0400903), the National Natural Science Foundation of China (No. 31601517, 31471923) and The Fundamental Research Funds for the Central Universities (DC201502020405).