
Abstract
Industrialising edible insects goes along with quality control and hazard analysis and critical control points. One of those steps is assessing heat treatment. For the present contribution, the potential of enzymatic heat assessment tests used in the dairy industry (alkaline phosphatase and lactoperoxidase) to detect heat treatment in several insect species (Acheta domesticus, Gryllus assimilis, Gryllus bimaculatus, Locusta migratoria, Schistocerca gregaria, Chilecomadia moorei, Galleria mellonella, Bombyx mori, Pachnoda marginata, Tenebrio molitor, Zophobas atratus, Apis mellifera, and Hermetia illucens) was evaluated. Insect material was homogenised, diluted, and the enzymatic tests (Lactognost®, Peroxtesmo®) were carried with these liquids as if they were milk. All species but C. moorei, B. mori, P. marginata, and A. mellifera showed alkaline phosphatase activity in raw samples and none in heated (10 min at 100 ℃) ones, while only G. mellonella, T. molitor, and Z. atratus reacted accordingly with lactoperoxidase. In trial 2 focusing only on alkaline phosphatase activity, inactivation of the enzyme after 5, 10, and 15 min of heating occurred species specific within a range of 60–86 ℃, i.e. within ordinary pasteurisation schemes. Thus and for the time being, heat treatment in many edible insect species can be assessed using alkaline phosphatase activity test kits. In contrast to milk samples, positive results may display bluish or greenish colours, and the time until a reliable reading is possible is extended to 1–1.5 h (24 h in the case of Gryllidae).
Introduction
Despite some cases of raw consumption, edible insects are typically eaten after heating. In traditional entomophagy, insects are cooked, roasted, or deep fried. Like in other foodstuffs, heating is necessary to eliminate pathogenic and spoilage microorganisms (Jay et al., 2005). Apart from that, some toxic species are made edible if their venom may be inactivated by heat (Nishimune et al., 2000).
Food microbiology is a crucial factor in the food safety of edible insects. It was seen that there is a common set of bacteria and fungi all insects seem to share, while other microorganism genera and species may occur specifically in subsequent taxa, from order down to species. Wild-caught insects (including edible species) may harbour also pathogenic microorganisms, e.g. salmonellae, Escherichia coli, staphylococci, and Aspergillus spp. (Grabowski and Klein, 2017a; Grabowski et al., 2017). In this way, edible insects are comparable to more common foodstuffs, e.g. meat or milk.
Now, entomophagy starts in areas in which it had been practically absent for centuries, e.g. Europe. Understanding edible insects as a foodstuff just like meat or milk, it is necessary to develop methods by which quality and safety may be verified. One of them is validating heat treatment.
Heat treatment in meat is usually assessed microbiologically and in case of need, i.e. foodstuff is processed according to hazard analysis and critical control points (HACCP) principles (including heating), and samples of each batch are retained to ensure quality and safety. Surpassing specific microbial growth thresholds is interpreted as lack of hygiene and/or insufficient heating (Klein and Schütze, 2011). Microbiological techniques are, however, time consuming and costly.
Milk differs from other foodstuffs by being even more susceptible for bacterial growth. This is what edible insects and milk have in common. The need to heat a foodstuff before putting it on the market in order to prevent pathogenic and spoilage microorganisms was recognised at latest when drinking milk of tuberculosis-positive cows was shown to lead to human tuberculosis. Boiling milk is an old practise, but milk pasteurisation was established less than 150 years ago (Von Soxhlet, 1886). Nowadays, there are several official pasteurisation and other heating techniques to treat liquid milk. Traditional pasteurisation refers to heating milk to 72–75 ℃ for 15–30 s, ultrahigh-temperature (UHT) treatment to 135–150 ℃ for 2–3 s. Between these two, several other treatments leading to so-called extended shelf life milk are allowed by the EU food regulation.
Heat treatment assessment in milk is done chemically. Pasteurisation and UHT treatment are confirmed by using enzymes which are thermally inactivated at temperature–time regimes similar to these heat treatments. There are millions of enzymes in living beings, but some of them share common functions, although structural and compositional differences exist (isoenzymes). The alkaline phosphatase (ALP) present in milk hydrolyses organophosphates and is located in fat globule membrane, playing an important role in the basic metabolism of animals. Lactoperoxidase (LP), part of the whey proteins, removes oxygen from peroxides, thus detoxifying them. They are inactivated at 70–75 ℃ and ≥85 ℃, and/or Although there are more elaborate reference methods, commercially available rapid test kits are used in the dairy industry. The principle is the same in both cases: the sample containing the enzyme is presented with the substrate for the enzyme under favourable conditions, and the work of the enzyme is made visible by colour. If the sample was raw and/or heated below the inactivation regime for the enzyme, the latter could work properly, leading to the desired colour change in the sample. If the sample was heated, the enzyme becomes denaturised, and either no colour change or a different colour occurs. Regarding ALP, a raw milk sample becomes blue and a heated one acquires a dusky greyish brown colour (Sinkiewicz and Kirst, 2006). Previous kits for LP detection used 2-ethoxyethanol (Traventol®) and used guaiacol as substrate for LP. Raw milk turned reddish brown when producing p-benzoquinones (Olszewski and Reuter, 1992). Due to the toxicity of Traventol®, safer systems were developed, e.g. Peroxtesmo® with a test mechanism not revealed by the manufacturer. Raw milk on the test paper turns blue, suggesting that the test is based on H2O2 and 3,3′,5,5′-tetramethylbenzidine, from which benzidine blue is synthesised (Kratz, 2014).
In insects, ALP has been detected in many species and instars. It was found in the digestive, excretory, and reproductive system, in haemolymph, the fat body, the cuticle, and the appendages. Where existent, ALP is involved in many metabolic pathways, from development and stimulus conduction to hormones synthesis, general metabolism, and even caste disposition in social species. It was studied moreover in pest insects since ALP seems to increase the animals’ resistance towards insecticides. In addition, the pathogenic potential of some entomopathogens may consist in impairing ALP activity (Yán et al., 2009). LP appears to be restricted to vertebrates where it also plays an important role in the defence of the digestive tract; this goal is achieved in the insect gut by a series of specialised cells (Boucias and Pendland, 1998). However, there are many different peroxidase isoenzymes which also occur in insects (Anderson, 2012).
The present paper intended to assert and, if possible, adapt the well-established heat assessment tests (ALP, LP) for edible insects in two steps. In the first step, the ability of the test to tell cooked insects from raw ones was assessed, leading to a ranking list (trial 1). Then, a precise time–temperature regime for the inactivation of the most promising enzyme for several species of edible insects was determined (trial 2).
Material and methods
Material
For trial 1, several edible species (A. domesticus (house cricket), G. assimilis (Jamaican field cricket), G. bimaculatus (Mediterranean cricket), L. migratoria (migratory locust), S. gregaria (desert locust), C. moorei (Chilean moth, butterworm), G. mellonella (greater wax moth), B. mori (silkworm), P. marginata (sun beetle), T. molitor (mealworm), Z. atratus (superworm), A. mellifera (European honeybee), and H. illucens (black soldier fly) were purchased alive and killed by freezing. Pooled samples per species of 10 g were homogenised (at 24,000 r/min for two times 30 s in order to avoid overheating of the sample) and diluted by 1:5 in distilled water.
Subsamples were separated into portions, and while one portion was kept raw, the other one was boiled for 10 min (Figure 1). Then, each subsample was submitted to enzyme testing using ALP (Lactognost®, Heyl Chemisch-Pharmazeutische Fabrik, Berlin, Germany) and LP (Peroxtesmo®, Macherey-Nagel, Düren, Germany) test kits. Samples were treated according to manufacturers’ recommendations, as if they were milk samples. For ALP, the Lactognost® kit contains one tablet with the substrate for ALP (disodium phenyl phosphate), one with a pH buffer (pH approx. 9.5) which are mixed in 10 ml of distilled water at 37 ℃ and 1 ml of sample, incubated at 37 ℃, and dibromoquinochlorimide is added to make the production of indophenols visible. Bluish or greenish colours were rated as ‘positive’, no colour changes or dusky, olive colours as ‘negative’. For LP, a drop of the sample was placed on the test paper, and while a blueish colour was regarded as ‘positive’, no colour change was indicative of a negative result. In this way, colour rating was similar to that in milk. Each test was repeated twice.
Sample preparation and processing in trial 1.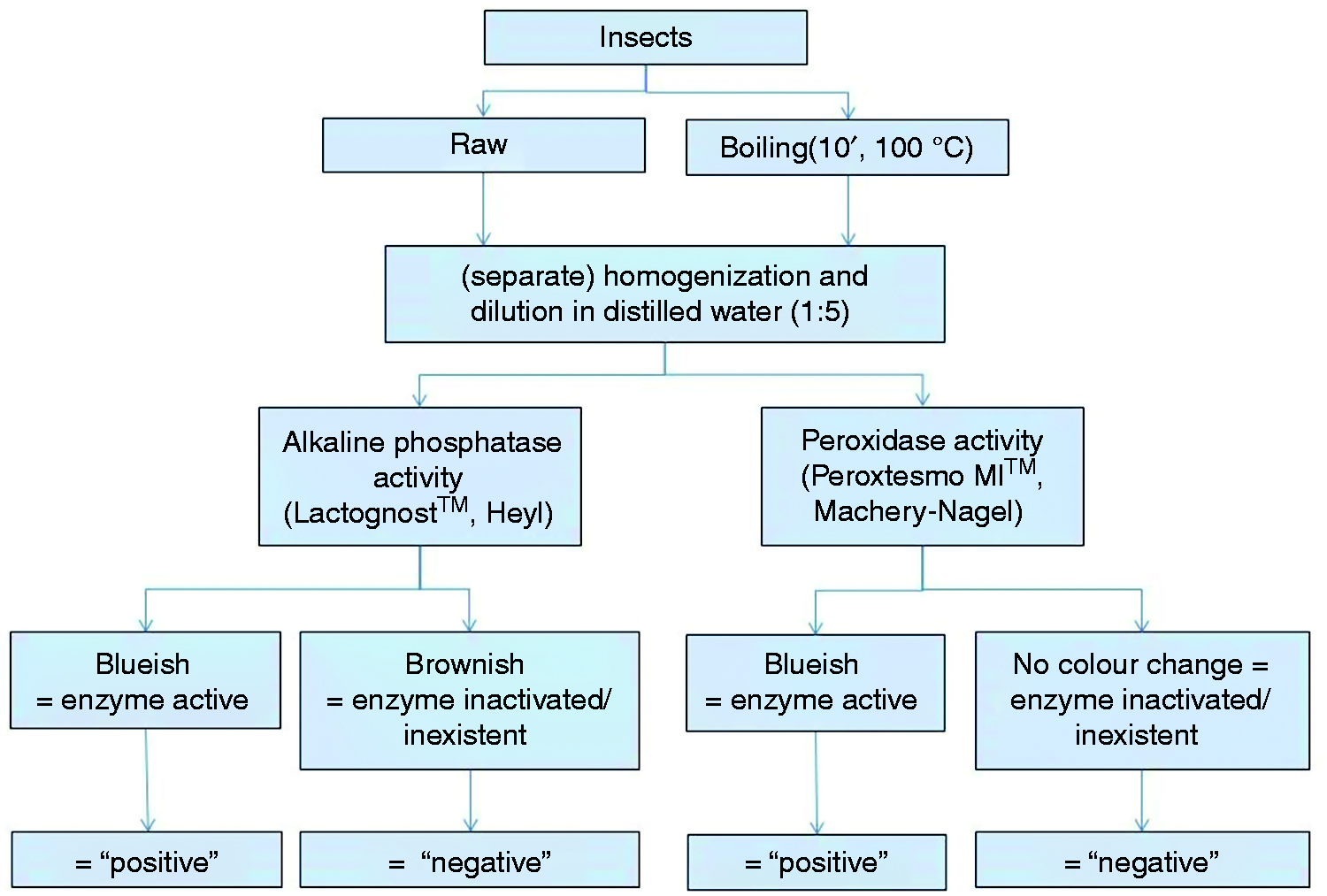
The trial 2 focused on ALP activity. Those species that did not react in trial 1were excluded, and fresh specimens of the reacting ones were purchased and processed in a similar way. However, samples were diluted by 1:10 and subsamples were submitted to different temperature–time regimes. Initially, heating starting at 20 ℃ for 10 min and in steps of 5 ℃, the samples were tested for ALP in same way as for trial 1, until the test became negative, plus another two heating steps. Then the interval was narrowed between the last positive and the first negative result, going in steps of 1 ℃. When it became clear that ALP became inactivated within a range similar to that of milk, temperatures between 60 and 90 ℃ were chosen. The procedure was repeated with heating times of 5 and 15 min. Colour evaluation was done like in trial 1.
Results
Trial 1
Reactivity of alkaline phosphatase (ALP) and lactoperoxidase (LP) in heated (100 ℃ for 10 min) edible insect species
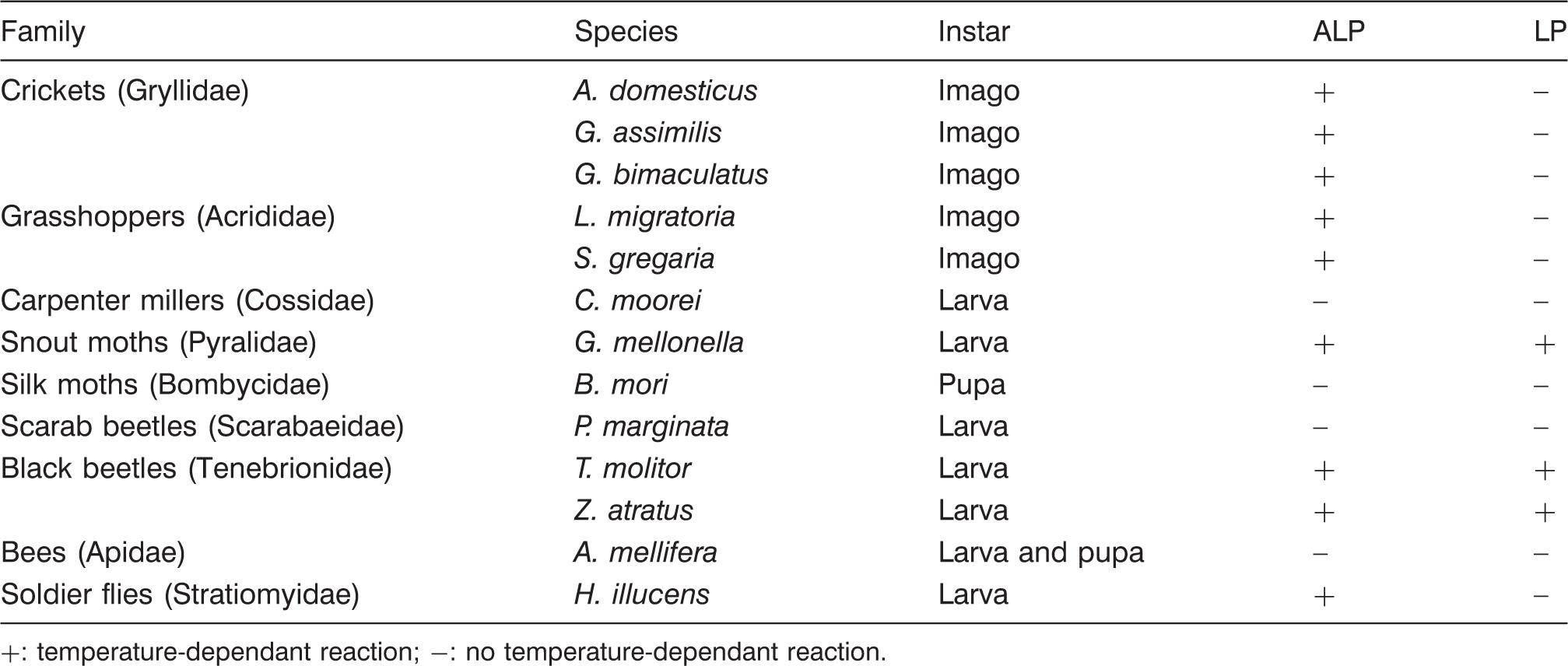
+: temperature-dependant reaction; −: no temperature-dependant reaction.
Trial 2
Inactivation temperature (℃) of alkaline phosphatase (ALP) in heated edible insect species
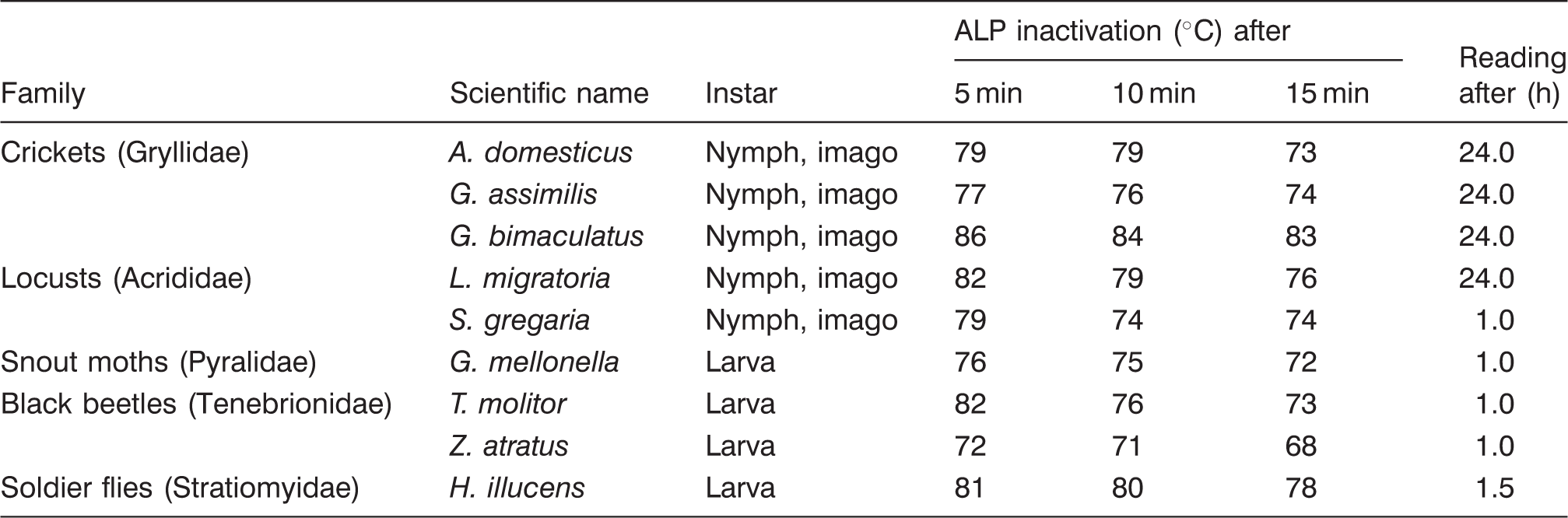
The inactivation temperature of ALP varied according to the species and the duration of the heat treatment, with increasing treatment duration resulting in lower inactivation temperatures. Each species displayed an individual inactivation curve. Taxon-related issues could not be determined since marked differences occurred within families (e.g. Tenebrionidae) and even genera (Gryllus spp.). However, a taxon-related influence was noted with regard to the times elapsing until a trustworthy reading was possible and to the extent of the colour change. Most orthopterans (crickets and locusts) required one day of reacting time, while a positive result within 60–90 min was typical for holometabolous insects. While locusts, the house cricket (A. domesticus), the tenebrionid beetles, and the black soldier fly (H. illucens) displayed clear bluish or greenish patterns in contrast to a light olive of negative samples, this distinction was more difficult in both Gryllus crickets and the wax moth (G. mellonella), even in the raw samples. The results as presented in Table 2 reflect the first temperature at which the light olive-brownish colour of inactivated insect ALP corresponded to that of cooked (100 ℃ for 10 min; see trial 1) insects. In milk terms, this would mean that a distinction was made between ‘slightly positive’ and ‘negative’.
During the trials, the Gryllus crickets were a particular challenge, and since a reaction was observed in trial 1, a second round was conducted, using the original 1:5 dilution. However, the colour intensity did not increase. Thus, the degree of dilution did not play a major role.
Finally, colour became more intensive in most species when the samples were stored for more time. Samples from tenebrionid beetles and wax moths, however, did not change.
Discussion
To our knowledge, using ALP and LP to determine the heating of edible insects is a novel approach. In their review, Pritchard et al. (2012) mention ALP in the context of stabilising silk which is the primary product from the silkworm (B. mori), but do not refer to ALP in the producing animal. Both enzymes are used in allergy assessment, also in insects (e.g. Phiriyangkul et al., 2015), but are part of the reactives and taken from other sources, e.g. horseradish.
However, the role of peroxidase to mitigate allergic responses is known, and accordingly, Sun-Waterhouse et al. (2016) propose peroxidase treatments on insect-based products.
ALP and LP in insects
As mentioned before, ALP is an enzyme which probably occurs in most insects species if not all. According to McComb et al. (2011), it was found in crickets, locusts, bees, some moths, and some beetles. Kim et al. (2011) detected ALP in H. illucens. No ALP records for C. moorei have been published, but McComb et al. (2011) mention its existence in the goat moth Cossus cossus, of the same family (Cossidae). It may be thus presumed that ALP may occur in all insects tested.
Although there are no records of LP in insects, peroxidases by themselves are central enzymes in insect physiology. Wu and Yi (2014) encountered a peroxidase in G. mellonella, Bruun Jensen et al. (2016) in honeybees, and since Anderson (2012) describes their role in the exoskeleton physiology, it may also be suggested that peroxidases do occur in all species.
In the present survey, ALP was detected in most species while the test for LP was only positive in pyralid moths and tenebrionid beetles. Since the employed test was designed for milk (LP), it is possible that insects ALP bear a closer resemblance to milk ALP than insect peroxidases do to LP. Thus, the milk test kit can only be recommended to test the heating of these three species. If higher temperature treatments are to be asserted in other species, alternatives must be developed.
Sources of variation of ALP activity
As seen in the ‘Results’ section, the degree of colour change, i.e. the activity of ALP, varied strongly between species. In their review, McComb et al. (2011) cite several reasons for a varying enzyme activity. One of them is the localisation within the insect’s body which may vary among species. Although for the survey, entire insects were used (for which the localisation per se is of no concern), it is possible that the presence in several organs, e.g. the digestive tract, the salivary glands, and the Malpighian tubes, may lead to more enzymes, resulting thus in a stronger response.
The sex of the individuals plays a role in some species (e.g. Musca domestica), but does not in others (e.g. cockroaches). This factor may also be excluded for the present study, as the insects were not grouped according to their sex; instead, both sexes were included in the homogenised samples.
However, there is an instar-related influence that is prone to having affected the results. Not all instars display the same amounts of ALP. In many species, recently laid eggs show no ALP activity, but do so shortly before hatching or after it. In some holometabolous species, ALP activity tends to increase towards pupation, in others like T. molitor, the peak activity was recorded during the early stages of the larva and imagines. Pupation is typically characterised by a very low or undetectable ALP activity. This is the reason why the test did not function with B. mori, the harvested instar being the pupa. These physiological fluctuations may become impaired in the case of disease. This at least was recorded in larvae of the cabbage moth Mamestra brassicae (Noctuidae) suffering from nosemiasis, where the physiological drop in ALP activity was not observed (McComb et al., 2011).
Finally, there is the possibility that the insect ALP isoenzymes in some species and/or instars are not compatible with the test system as such. Although present in mare milk and colostrum (Hank et al., 1993), ALP could not be detected via Lactognost® in a raw mare milk sample sent to the authors’ laboratory several years ago, probably to structural incongruences.
Using ALP activity to assert pasteurisation of edible insects
The observed inactivation temperature–time combinations ranged within the typical pasteurisation schemes (between 60 and 100 ℃). However, it will be necessary to premeditate on the goals when heat treating edible insects. If a minimal approach is intended, e.g. to preserve as much of the original flavour as possible, then the microbial efficacy of the chosen heat treatment must be evaluated first. It was seen that there are also species-specific reactions to heat treatment (Grabowski and Klein, 2017b), and since there are many spore-forming bacteria present in edible insects, these may not be inactivated accordingly. This however also occurs in liquid milk, leading to a short shelf life of this product.
Yet, if a longer shelf life is the goal, then a prolonged heating of the insects (i.e. 100 ℃ for 10 min) will be the safer option. However, it is likely that this measure will reduce, but not eliminate microbiological spoilage of the insects, just as is the case with other foodstuffs, e.g. milk.
From the technical point of view, dilution of the sample did not play a major role, but the time until a reliable reading was possible. This should be considered when planning a larger operational unit. Finally, the colours obtained also varied with the species, ranging from blue (as seen in milk) to bluish-greenish nuances.
Conclusions
Rather than LP, ALP resulted as a suitable tool to assert pasteurisation in most tested insect species. For the time being, the ALP activity test designed for milk as an economical and relatively timely option may be used. However, results regarding the temperature–time combination were species and instar specific, and reading results were obtained later than dairy samples.
Footnotes
Acknowledgement
The authors wish to thank VMTA Barbara Bouda for carrying out the analysis of trial 2.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.