
Abstract
The characterization of autochthonous cultures based on their contribution to cheese flavor is an additional selection criterion for their use in cheese making. The objective of the present work was to assess the ability of three strains of mesophilic lactobacilli: Lactobacillus casei 72 (Lc72), L. paracasei 90 (Lp90), and L. plantarum 91 (Lp91), one strain of thermophilic lactobacillus: L. helveticus 209 (Lh209), and the thermophilic–mesophilic combinations, to grow and produce aroma compounds in a hard cheese model. Microbiological counts, pH, and the profiles of carbohydrates, organic acids, and volatile compounds were analyzed during incubation for 14 days at 37 ℃. The population of mesophilic lactobacilli reached levels around 8.0 log CFU ml−1 at three days, but then decreased until ∼7.0 log CFU ml−1 toward 14 days. Thermophilic lactobacillus population reached and maintained levels around 7.7 log CFU ml−1 during incubation. Carbohydrates were absent in the hard cheese model, and so no change in the pH values and in the levels of lactic acid was detected. Mesophilic lactobacilli, inoculated individually or in association with Lh209, metabolized the citric acid and produced ethanoic acid. The profiles of volatile compounds of mesophilic lactobacilli (characterized mainly by butan-2-one, 3-hydroxybutan-2-one, 3-methylbutan-1-ol, hexan-1-ol, 2-phenylethanol, and ethanoic acid) were different from the profile of thermophilic lactobacillus Lh209 (characterized mainly by heptan-2-one, ethyl acetate, isoamyl hexanoate, pentan-1-ol, decanoic acid, and 2- and 3-methylbutanal). Cooperative effects in the production of compounds related to cheese flavor, such as 3-hydroxybutan-2-one, ethyl butanoate, ethanol, pentan-2-ol, hexan-1-ol, benzeneacetaldehyde, 2-phenylethanol, and heptanoic acid, were largely evidenced between Lh209 and Lp91; in a lesser extent, cooperative effects were also found for Lh209+Lp90 for the following compounds: 3-hydroxybutan-2-one, isoamyl acetate, and ethanoic acid. Of the mesophilic lactobacilli strains evaluated, Lp91 and Lp90 would be interesting candidates for its use as adjunct cultures in hard cheeses to improve and diversify the flavor.
Keywords
Introduction
During cheese ripening, several biochemical events occur, leading to the formation of a great number of flavor compounds which are associated with the enzymatic activities of the microbiota present in the cheese matrix (Steele et al., 2013). Particularly, in the long-ripening hard cheeses, thermophilic lactobacilli used as starter and adventitious mesophilic lactobacilli of non-starter lactic acid bacteria (NSLAB) origin coexist and develop dynamically (Gatti et al., 2014; Gobbetti et al., 2018; Montel et al., 2014). The main role of the starter is to act during cheese production, converting the lactose to lactic acid in few hours, with the consequent pH reduction; in addition, it could also contribute to the production of volatile compounds impacting in sensory characteristics of the cheese (Helinck et al., 2004; Klein et al., 2001). On the other hand, the influence of NSLAB in the cheese ripening is strain-dependent. In previous studies, several strains of NSLAB demonstrated good properties for their use as adjunct cultures in cheeses, because they were able to control the growth of potentially harmful microorganisms and contribute to the development of a mature and improved flavor, especially in cheeses made with pasteurized milk (Gobbetti et al., 2015; Hynes et al., 2003; Irigoyen et al., 2007; Rynne et al., 2007). In particular, the use of selected NSLAB strains with key enzymatic activities involved in the production of volatile compounds is an interesting strategy to improve, diversify, and modulate the cheese flavor (Settanni and Moschetti, 2010). However, in order to assure the usefulness of this strategy, the technological performance of the adjunct culture, the interaction with the starter, and the specific impact on the maturation profiles of cheeses must be previously studied (Sgarbi et al., 2013; Smit et al., 2009; Stefanovic et al., 2017). Some studies of the associative effect between lactococci (used as starter) and mesophilic lactobacilli (used as adjunct culture) in cheese flavor development have been reported (Amárita et al., 2001; Ayad et al., 2000; Kieronczyk et al., 2004), while there are no studies about the cooperation between mesophilic lactobacilli and thermophilic lactobacilli, according to our knowledge.
Many works about the evaluation of the role of the cultures in the cheese flavor development and their impact on the quality have been carried out in cheese made in pilot scale. Although these studies are the most representative, they demand high economic resources and take a considerable time. Alternatively, simpler model systems that mimic the ripening conditions of the cheese have been proposed to evaluate the effect of different cultures in a rapid, economical, and reproducible way (Budinich et al., 2011; Milesi et al., 2011; Sgarbi et al., 2013; Stefanovic et al., 2017).
The objective of the present work was to assess the ability of three strains of mesophilic lactobacilli: Lactobacillus casei 72 (Lc72), L. paracasei 90 (Lp90), and L. plantarum 91 (Lp91), one strain of thermophilic lactobacillus: L. helveticus 209 (Lh209), and the thermophilic–mesophilic combinations, to grow and produce aroma compounds in a hard cheese model. This approach is proposed as a selection step before the application of autochthonous cultures in cheese making.
Materials and methods
Strains
One strain of thermophilic lactobacillus: L. helveticus 209 (Lh209), and three strains of mesophilic lactobacilli: L. casei 72 (Lc72), L. paracasei 90 (Lp90), and L. plantarum 91 (Lp91), belonging to the INLAIN collection, were used. Lh209 was isolated from whey starters provided by Reggianito cheese-producing industries (Reinheimer et al., 1995, 1996) and the technological and biochemical properties have been studied in cheeses and model systems (Candioti et al., 2002; Cuffia et al., 2018; Hynes et al., 2003; Milesi et al., 2011; Perotti et al., 2005). Mesophilic lactobacilli were isolated from good quality Tybo cheeses and they have been characterized regarding their genetic and probiotic properties (Bude Ugarte et al., 2006; Burns et al., 2012). In addition, the technological performance in in vitro assays (salt tolerance, phage resistance, acidifying and proteolytic activities) and in soft and semi-hard cheese models (Briggiler-Marcó et al., 2007; Milesi et al., 2010; Peralta et al., 2014) has also been studied. The aminotransferase (AT) and glutamate dehydrogenase activities of the tested strains were previously determined (Peralta et al., 2016a).
Preparation of hard cheese model
Preparation of the hard cheese model was performed according to Cuffia et al. (2018). Briefly, Reggianito cheeses, made in the pilot plant of our institute, using Lh209 as starter, were processed after three months of ripening with distilled water (1:1 w/v) using a food processor. Then, the suspension was centrifuged at 8000 g/15 min/5 ℃.The aqueous phase was extracted, filtered, and the NaCl content and pH were adjusted at 4% w/v and 5.25, respectively. Finally, it was sterilized by heat (70 ℃ for 30 min). These cheese extracts were used as hard cheese model.
Experimental design
Experimental design
pH and microbiological analysis
The pH was measured at 3, 7, and 14 days of incubation with a pH meter (Orion 3-star benchtop, Thermo Fisher Scientific Inc., USA) by direct immersion of the electrode.
Counts of mesophilic and thermophilic lactobacilli at 3, 7, and 14 days of incubation were made by spread-plate method (MRS-agar) using 0.1 ml of the decimal dilutions obtained in sterile peptone water. The plates were incubated for 48 h at 37 and 43 ℃, respectively. Thermophilic lactobacilli only grew at 43 ℃, while at 37 ℃ both mesophilic and thermophilic lactobacilli were able to grow, but in this case mesophilic lactobacilli were differentiated from Lh209 according to the different morphology of the colonies: pin-point opaque colonies with small center and transparent halo-shaped border for Lh209 and small, smooth, and creamy colonies for the three strains of mesophilic lactobacilli (Bude Ugarte et al., 2006; Reinheimer et al., 1996).
Carbohydrates and organic acids
The content of organic acids (lactic, citric, ethanoic, and butanoic) and carbohydrates (lactose, glucose, and galactose) in the extracts at 14 days of incubation was analyzed by high-performance liquid chromatography (with the UV and IR detectors in series) according to Peralta et al. (2016b). Samples were centrifuged (5000 g/10 min/10 ℃), and the supernatants were diluted in the mobile phase (1:4 v/v), filtered through 0.45 µm membranes (Millex, Millipore, São Paulo, Brazil) and a volume of 60 µl was injected into the chromatograph.
Volatile compounds
The profiles of volatile compounds were determined at the end of incubation (14 days). Solid-phase microextraction (SPME) was applied as sampling technique, using a divinylbenzene/carboxen/polydimethylsiloxane fiber (DVB/Car/PDMS 50/30 µm, Supelco, USA). Prior to analysis, frozen samples were thawed at 4 ℃ overnight. Aliquots (10 ml) were transferred to a screw-top glass vial (40 ml) containing a micro stirring bar and capped with a cap with a silicon rubber/teflon membrane and a hole enabling SPME sampling. Vial was introduced in an aluminum block placed on a heater/stirrer (IKA, USA) and preheated at 40 ℃ for 10 min with an agitation speed of 250 r min−1. Then, the fiber was exposed to the headspace for 30 min at 40 ℃. Separation, identification, and semi-quantitative analysis of volatile compounds by GC-FID/MS were performed as described by Wolf et al., 2016). Three replicates were analyzed for each sample and the results were expressed as peak areas (arbitrary units).
Statistical analysis
One-way analysis of variance with fixed factors was conducted on the results of pH, carbohydrates, organic acids, and volatile compounds using Minitab 18 software (Minitab Inc., State College, PA, USA). When differences were significant (p < 0.05), the means were compared using Tukey's test. In the case of microbiological analyses, Student's t-test was applied. In addition, volatile compounds were subjected to a multivariate analysis, specifically, principal component analysis (PCA) using XLSTAT 2014.5 software (Addinsoft SARL).
Results and discussion
pH and microbiological analysis
The population of mesophilic lactobacilli either inoculated individually or together with Lh209 increased until 7.7–8.4 log CFU ml−1 on the third day, remained at similar levels at seven days of incubation (except for Lc72 individually inoculated, which grew 0.5 log CFU ml−1), and finally decreased until ∼7.0 log CFU ml−1 toward 14 days (Figure 1). Thermophilic lactobacilli population reached levels of 7.3–8.0 log CFU ml−1 (data not shown), which were maintained during incubation period. No microbial growth was observed in the control extracts. The good survival of the lactobacilli reflects their ability to use alternative energy sources (free fatty acids, short peptides, amino acids (AAs), citrate) instead of simple sugars which were absent in the extracts (Skeie et al., 2008; Stefanovic et al., 2017).
Evolution of the population of mesophilic lactobacilli during the incubation: extracts inoculated only with the mesophilic lactobacilli (black bars) and extracts with mesophilic–thermophilic combination (grey bars). (A) Lactobacillus casei 72 (Experiment 1), (B) Lactobacillus paracasei 90 (Experiment 2), (C) Lactobacillus plantarum 91 (Experiment 3). Note: Different letters at each sampling time indicate significant differences (p < 0.05).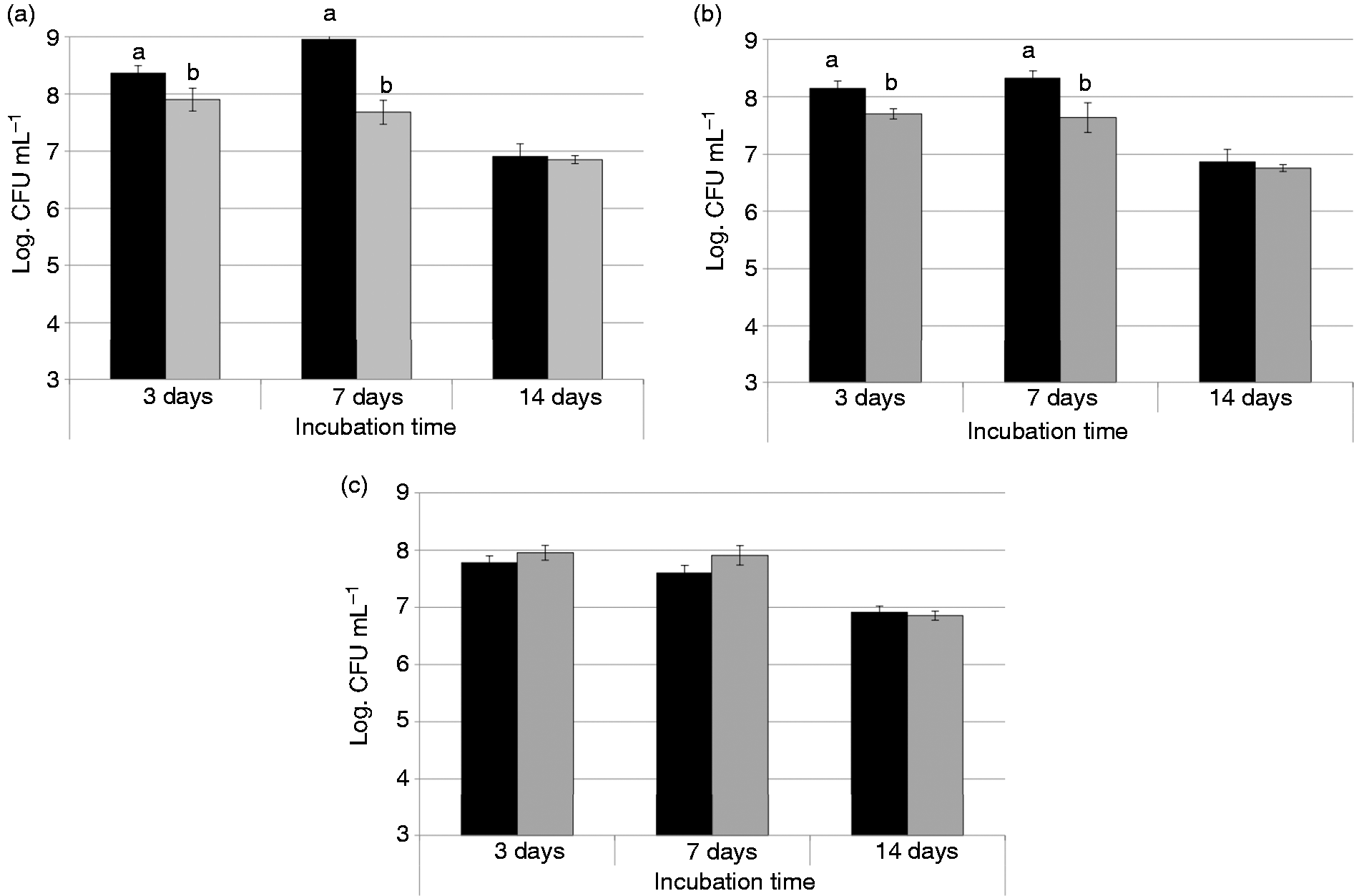
At three and seven days, counts of Lc72 and Lp90 were significantly higher (p < 0.05) in the extracts inoculated individually with these strains than in the extracts inoculated together with Lh209 (Figure 1(a) and (b)). This could be due to a competitive effect between mesophilic and thermophilic lactobacilli in the first days of incubation, which could indicate that Lh209 would have better capacity to metabolize the nutrients of the medium in comparison to mesophilic lactobacilli (Gatti et al., 2014). However, the final levels of mesophilic lactobacilli were high.
In relation to pH, the values for the inoculated extracts were similar to those of the controls in the three experiments and remained constant during incubation; the values were in the range of 5.02–5.10.
Profiles of carbohydrates and organic acids
Concentration of organic acids in the extracts at the end of incubation (expressed as mg 100 ml−1 of extract)
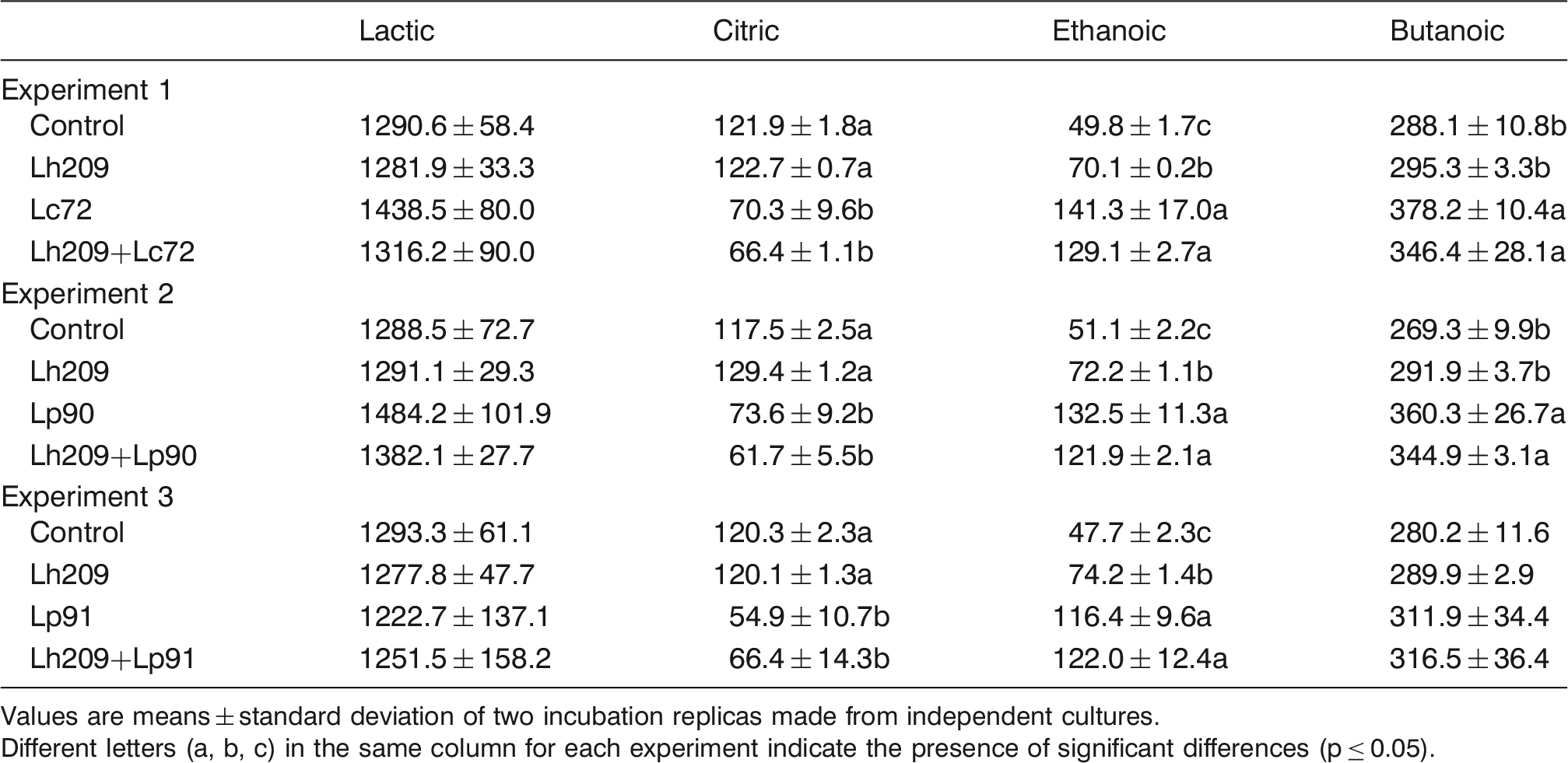
Values are means ± standard deviation of two incubation replicas made from independent cultures.
Different letters (a, b, c) in the same column for each experiment indicate the presence of significant differences (p ≤ 0.05).
Lh209 increased the levels of ethanoic acid. This strain did not metabolize citric acid, which was expected because thermophilic lactobacilli are considered citrate-negative (McSweeney, 2011). In contrast, in the extracts inoculated with the mesophilic lactobacilli strains, individually or in association with Lh209, a consumption of this compound was verified, which was correlated with an increase in the content of ethanoic acid. The degradation of citrate with the concomitant production of ethanoic acid by mesophilic lactobacilli strains was reported previously (Sgarbi et al., 2013). In addition, ethanoic acid can also be synthesized via AA catabolism such as serine and aspartic acid (Liu, 2003), which is related with the enzymatic activities found for these strains (Peralta et al., 2016a).
On the other hand, the extracts with Lc72 and Lh209 + Lc72, and Lp90 and Lh209 + Lp90 had higher contents of butanoic acid in relation to control extracts and those inoculated only with Lh209 (Table 2). Butanoic acid is an important flavor compound in cheeses, formed mainly from metabolism of milk fat (Collins et al., 2003), or from AA deamination (Buffa et al., 2004).
Profiles of volatile compounds
Volatile compounds in the extracts of experiment 1 at the end of incubation (expressed as areas in arbitrary units)
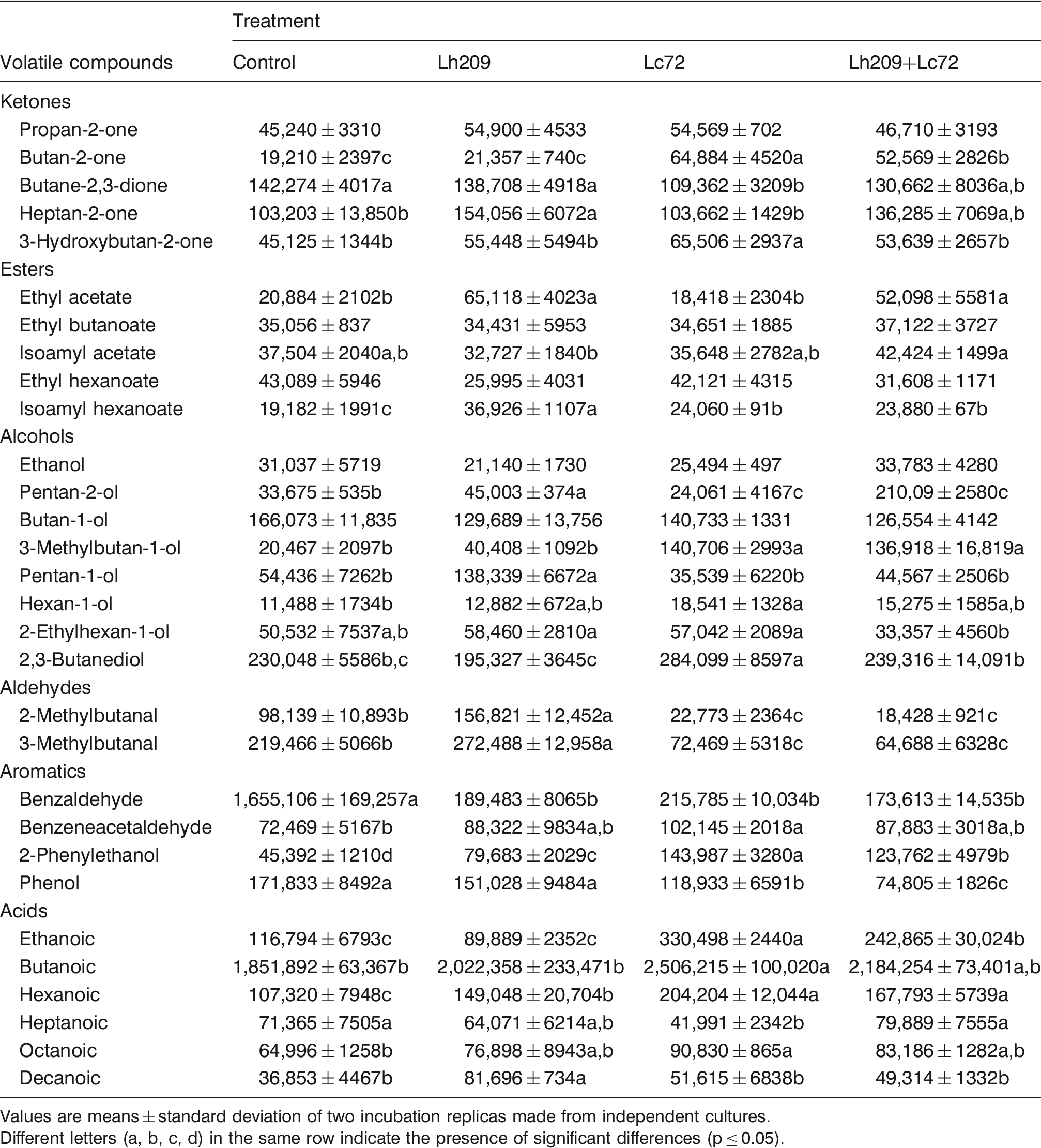
Values are means ± standard deviation of two incubation replicas made from independent cultures.
Different letters (a, b, c, d) in the same row indicate the presence of significant differences (p ≤ 0.05).
Volatile compounds in the extracts of experiment 2 at the end of incubation (expressed as areas in arbitrary units)
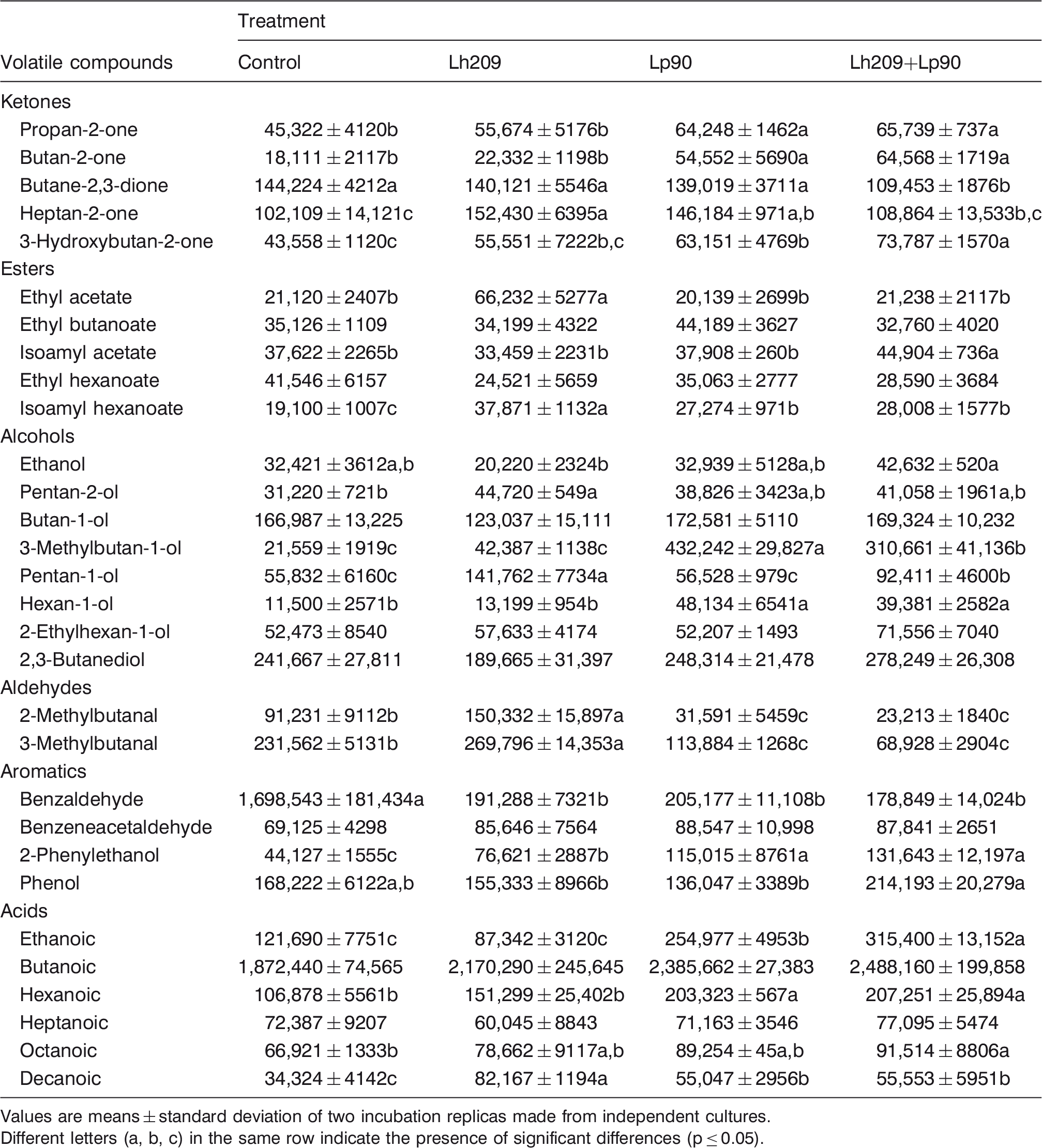
Values are means ± standard deviation of two incubation replicas made from independent cultures.
Different letters (a, b, c) in the same row indicate the presence of significant differences (p ≤ 0.05).
Volatile compounds in the extracts of experiment 3 at the end of incubation (expressed as areas in arbitrary units)
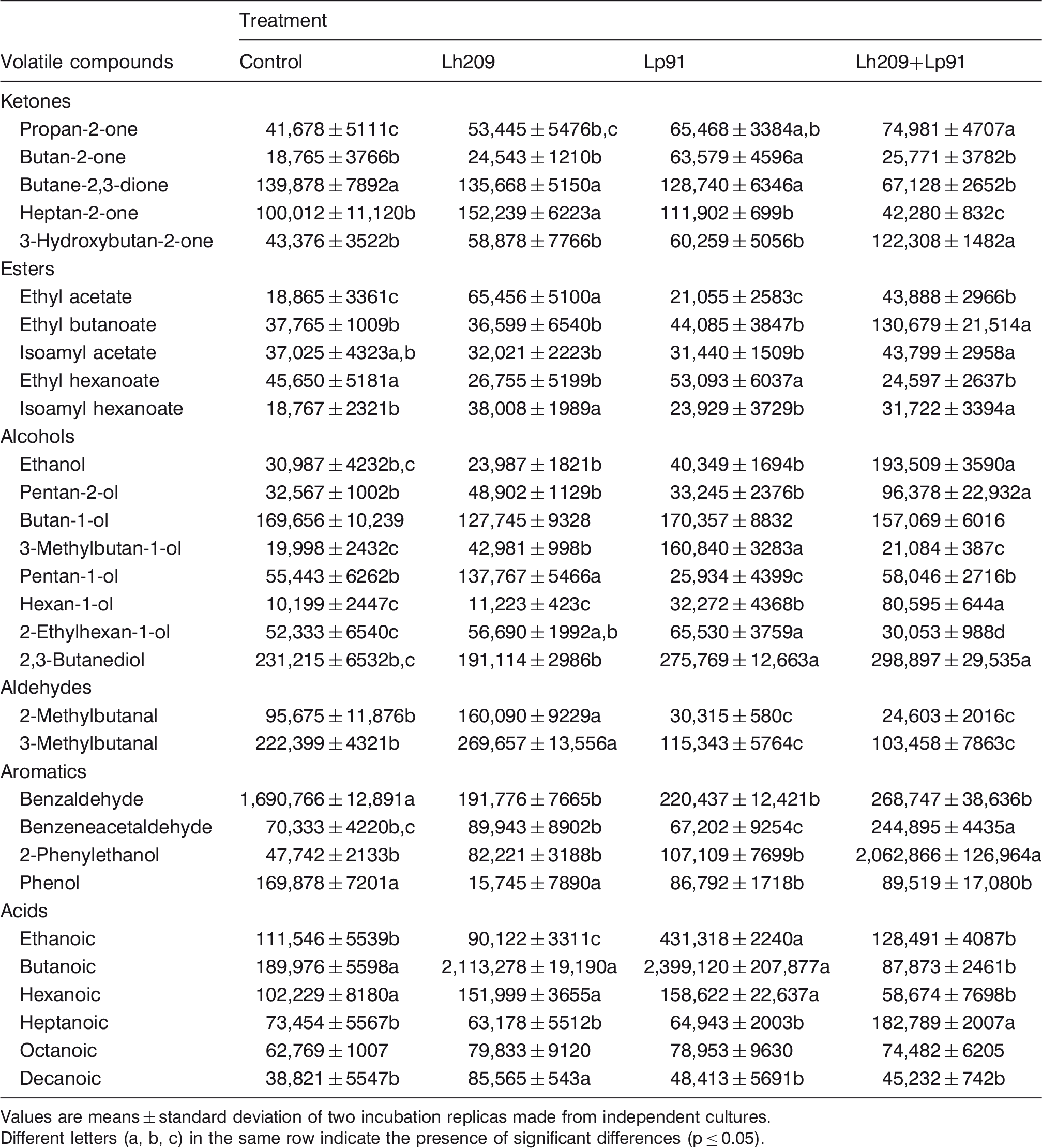
Values are means ± standard deviation of two incubation replicas made from independent cultures.
Different letters (a, b, c) in the same row indicate the presence of significant differences (p ≤ 0.05).
In experiment 1 (Table 3), Lh209 was characterized by the production of compounds such as heptan-2-one, ethyl acetate, isoamyl hexanoate, pentan-1-ol, pentan-2-ol, 2- and 3-methylbutanal, and decanoic acids. The production of volatile compounds by L. helveticus strains has been little studied in comparison with other LAB. However, their ability to produce aroma compounds through different metabolic pathways has been demonstrated in cheeses and in cheese models (Collins et al., 2003; Helinck et al., 2004; Klein et al., 2001). In fact, the formation of 2- and 3-methylbutanal could be correlated with the AT activity toward branched AAs (leucine and isoleucine) (Peralta et al., 2016a). Esterases of LAB can catalyze ester-forming reactions such as esterification and alcoholysis (reactions between medium or short chain fatty acids and partial glycerides with alcohols, respectively) (Holland et al., 2005). The information related to the esters production by L. helveticus strains is scarce. Methylketones are also important flavor compounds, produced by different LAB, including L. helveticus (Klein et al., 2001). Primary alcohols, such as pentan-1-ol, are formed by the reduction of aldehydes derived from AAs and fatty acids, while secondary alcohols, such as pentan-2-ol, are formed from methyl ketones (Collins et al., 2003). Lc72 showed the ability to produce butan-2-one, 3-hydroxybutan-2-one, 3-methylbutan-1-ol, hexan-1-ol, 2,3-butanediol, 2-phenylethanol, benzeneacetaldehyde, and ethanoic, butanoic, hexanoic, and octanoic acids. Contrary, this strain consumed 2- and 3-methylbutanal, both when was inoculated alone or combined with Lh209. This fact showed the ability of Lc72 to metabolize these aldehydes to their respective alcohols (e.g. 3-methylbutan-1-ol) or other volatile compounds. In addition, benzaldehyde was also consumed in the Lc72, Lh209, and Lc72+Lh209 extracts, and phenol in the Lc72 and Lh209+Lc72 extracts. When Lh209 and Lc72 were grown together increased levels of butan-2-one, isoamyl hexanoate, ethyl acetate, 3-methylbutan-1-ol, 2-phenylethanol, and ethanoic and hexanoic acids were found. The levels of these compounds were similar or intermediate to those in extracts inoculated with individual strains. Thus, no cooperative effect was observed in the production of volatile compounds for the combination of Lh209 and Lc72. A cooperative or associative effect takes place when the production of compounds in the extract inoculated with both strains exceeds to that found in the extracts inoculated with the individual strains.
In the experiments 2 and 3, the behavior of Lh209 (Tables 4 and 5) was similar to that described in the E1. Additionally, a diminution of benzaldehyde, and 2- and 3-methylbutanal in extracts with mesophilic lactobacilli (Lp90 and Lp91), inoculated alone and with Lh209, was also observed in these experiments.
In E2, Lp90 had the ability to synthesize propan-2-one, butan-2-one, heptan-2-one, 3-hydroxybutan-2-one, isoamyl hexanoate, 3-methylbutan-1-ol, hexan-1-ol, 2-phenylethanol, and ethanoic, hexanoic, and decanoic acids. These compounds were also increased in the Lh209 + Lp90 extracts, suggesting that Lp90 led the production of volatile compounds when both strains were together. In addition, cooperative effects between Lh209 and Lp90 were found for 3-hydroxybutan-2-one, isoamyl acetate, and ethanoic acid.
In E3 (Table 5), Lp91 produced propan-2-one, butan-2-one, hexan-1-ol, 3-methylbutan-1-ol, ethylhexan-1-ol, 2,3-butanediol, and ethanoic acid. Similar or intermediate levels for propan-2-one, ethyl acetate, and 2,3-butanediol were detected in the Lh209+Lp91 extracts in comparison to extracts inoculated with the individual strains. Cooperative effects were detected for 3-hydroxybutan-2-one, ethyl butanoate, ethanol, pentan-2-ol, hexan-1-ol, benzeneacetaldehyde, 2-phenylethanol, and heptanoic acid.
Among the volatile compounds detected in the extracts, 3-hydroxybutan-2-one has an important role in the flavor development of fermented milk products and it is produced by the reduction of butane-2,3-dione that can be generated via citrate metabolism (Le Bars and Yvon, 2008). In our work, a diminution of the citrate levels and an increase of 3-hydroxybutan-2-one were detected in the extracts inoculated with Lc72 and Lp90, and with the combination of Lh209 + Lp90 and Lh209 + Lp91.
Many authors have reported the production of volatile compounds by strains of L. casei, L. paracasei, and L. plantarum through different mechanisms, mainly associated with AA and citrate metabolism (Gobbetti et al., 2015; Le Bars and Yvon, 2008; Milesi et al., 2010; Pogačic et al., 2015), although the specific metabolic pathways of NSLAB in aroma compounds formation are still poorly understood.
In addition, volatile compounds profiles produced by the individual lactobacilli strains and by the three-pairing thermophilic–mesophilic lactobacilli were analyzed jointly by a multivariate approach.
For that, PCA was applied in order to identify groupings of the samples and visualize associations or differences between strains or combinations of strains, using the correlation matrix (centered and scaled values). A two-PC model representing 78.1% of the cumulative total variance of the system (eigenvalues >1) was selected; seven compounds or variables were eliminated since their communalities were low (<0.75). PC1 represented 51.5%, while PC2 contributed 26.6% of the total variance. As visualized in the scores plot (Figure 2(a)), the samples were grouped in several groups in the plane defined by the two first PC. It was appreciated that the inoculated extracts with individual strains and with the combinations of thermophilic–mesophilic lactobacilli were distinguished perfectly from the controls. Lc72, Lp90, and Lp91 extracts were quite similar and completely different from Lh209 extracts. PC1 separated the samples clearly into two groups: the Lh209+Lp91 extracts were located at the positive end and were characterized by the compounds that contributed positively on this PC (pentan-2-ol, heptanoic acid, ethyl butanoate, benzeneacetaldehyde, 2-phenylethanol, ethanol, 3-hydroxybutan-2-one, and hexan-1-ol); the rest of the samples was grouped at the negative end and were associated with butanoic, hexanoic, octanoic, and ethanoic acids; butane-2-one; butane-2,3-dione; heptan-2-one; and 3-methylbutah-1-ol. Four groups were distinguished from the positive end toward the negative end on the PC2 axis: 1—Lh209; 2—control; 3—Lh209+Lc72 and Lh209+Lp91; and 4—Lc72, Lp90, Lp91, and Lh209+Lp90, respectively. The two branched chain aldehydes (2- and 3-methylbutanal) were positively related to PC2, whereas ethanoic acid, 3-methylbutan-1-ol, and butan-2-one contributed negatively to this PC (Figure 2(b)). Results obtained show that the production of volatile compounds of the three strains of mesophilic lactobacilli inoculated individually or combined with Lh209 was clearly distinguished from the thermophilic lactobacilli; associative effects were mainly observed for Lh209+Lp91.
Score plot (a) and loading plot (b) (PC1 vs. PC2) of the PCA applied on the volatile compounds of the hard cheese extracts.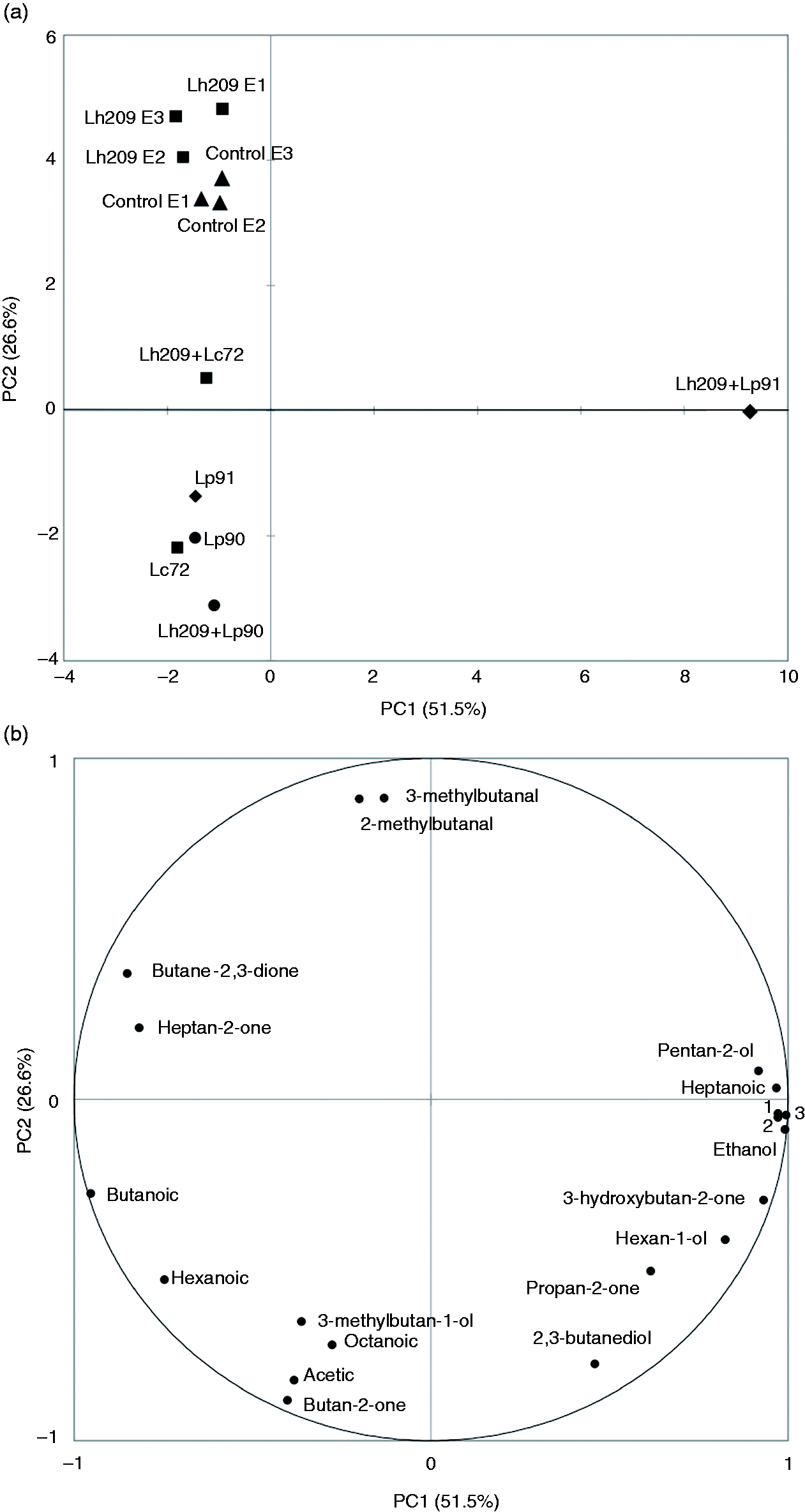
Conclusion
The abilities of four autochthonous lactobacilli strains, three mesophilic: Lc72, Lp90, and Lp91, and one thermophilic: Lh209, and the thermophilic–mesophilic combinations, to grow and produce volatile compounds in a hard cheese model, were studied. The profiles of volatile compounds produced by mesophilic lactobacilli and thermophilic–mesophilic combinations were differenced from that of thermophilic lactobacilli. Cooperative effects in the production of some volatile compounds related to cheese flavor were detected, mainly for the combination of Lh209 and Lp91, but also for Lh209 and Lp90. This fact suggests that Lp91 and Lp90 would be interesting candidates as adjunct cultures in hard cheeses to improve and diversify the flavor. The selection of individual strains and the detection of associations between them, based on the production of volatile compounds, represent a potential tool for designing cultures in order to obtain cheeses with distinctive flavor attributes.
Supplemental Material
FST881512 Supplemental Material - Supplemental material for Evaluation of autochthonous cultures to improve the cheese flavor: A case study in hard cheese model
Supplemental material, FST881512 Supplemental Material for Evaluation of autochthonous cultures to improve the cheese flavor: A case study in hard cheese model by Facundo Cuffia, Carina V Bergamini, Érica R Hynes, Irma V Wolf and María C Perotti in Food Science and Technology International
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors acknowledge Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, Argentina) for the doctoral fellowship of Facundo Cuffia. The present work was financially supported by grants from Universidad Nacional del Litoral (CAI+D No. 501-201-10100082-LI).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.