
Abstract
Recent research has significantly altered the understanding of the immunogenic profile of certain processes of cancer cell death, leading to the recognition of a new subclass of apoptosis called “immunogenic apoptosis.” This form of cell death, induced by specific chemotherapeutic agents, has been shown to elicit a “chemotherapy vaccine effect” in vivo, effectively stimulating an antitumor immune response. At the molecular level, “a collection of molecules” known as “damage-associated molecular patterns (DAMPs)” have been identified as key contributors to the immunogenicity of various cell death pathways. Intracellular molecules, such as heat-shock proteins, high-mobility group box 1 protein, and calreticulin, act as DAMPs when exposed or secreted in response to specific certain stressors, stimuli, and modes of cell death. These discoveries have fueled ongoing research focused on the identification of novel DAMPs, uncovering new mechanisms of their exposure or secretion and developing therapeutic agents capable of inducing immunogenic cell death (ICD). In addition, there is growing interest in addressing the current challenges and limitations within this emerging paradigm. The authors believe that this integrated strategy—combining DAMPs, ICD, and anticancer therapies—may hold the key to significantly reducing cancer-related mortality in the near future.
Introduction
Endogenous substances known as damage-associated molecular patterns (DAMPs) are released from host cells during cell death or injury. The loss of tissue homeostasis is closely associated with the release of DAMPs in tissues. 1 Innate immune receptors that recognize DAMPs trigger inflammatory responses, which can contribute to chronic inflammatory disorders but may also aid in restoring tissue homeostasis. 2 In recent years, the detection of intracellular DAMPs has garnered significant attention in the context of sterile inflammation. 3 Notably, DAMPs can originate not only from adjacent cells but also from distant organs or tissues, mediating systemic sterile inflammatory responses. This multilevel DAMP sensing facilitates trans-tissue, trans-organ, and intercellular communication. 4 DAMP-sensing receptors detect DAMPs from intracellular, intercellular, or distal tissue and organ sources and drive downstream immune response. Consequently, targeting DAMPs or their receptors presents a promising strategy for managing inflammatory disorders. 5 DAMPs are immune-stimulatory molecules released during cellular damage. They attract and activate immune cells and promote the release of inflammatory mediators. Depending on the nature of cellular injury or death such as pyroptosis, ferroptosis, necrosis (often induced by physicochemical insults, infections, or metabolic stress), or apoptosis (a tightly regulated form of programmed cell death essential for tissue homeostasis), the molecular pathways and biological outcomes may differ substantially. DAMPs may be released into the extracellular environment such as high-mobility group box 1 (HMGB1) and interleukin-1 α (IL-1α), or exposed on the plasma membrane, such as calreticulin (CRT). During the terminal stages of cell death, additional DAMPs are generated, including deoxyribonucleic acid (DNA) and adenosine triphosphate (ATP).6,7
Despite producing similar immune effects, DAMPs and pathogen-associated molecular patterns (PAMPs) originate from fundamentally different sources. DAMPs are endogenous signals that mediate “sterile inflammation,” whereas PAMPs are microbial components such as lipopolysaccharides (LPS). Moreover, during infection, PAMPs can induce the release of DAMPs, thereby intensifying the inflammatory response. 8 For instance, studies have shown that extracellular heat shock protein 72 (HSP72) is released from the airway epithelium in response to respiratory syncytial virus infection. Such findings are guiding the development of novel immunotherapy and chemotherapy approaches for patients with lung adenocarcinoma conditions.
Severe traumatic injury induces an immediate and prolonged state of systemic immunosuppression, increasing the risk of hospital-acquired infection. Following trauma, a broad array of DAMPs, including proteins, lipids, and DNA, are known to contribute to a systemic inflammatory response syndrome (SIRS). These DAMPs can appear within minutes and persist for days to weeks postinjury. Recent studies suggest that DAMPs not only initiate inflammation but also mediate post-traumatic immune suppression, suggesting additional immunomodulatory roles in regulating immune response. 9
To provide a comprehensive overview of the novel immune-tolerizing effects, this review summarizes findings from in vitro, in vivo, and ex vivo studies focusing on DAMPs originating from cytosolic, nuclear, and mitochondrial sources. 10 Among the immunosuppressive effects attributed to DAMPs, one hypothesis is that they may directly suppress neutrophil antimicrobial activities, potentially influencing tissue-resident immune cell behavior. While it is plausible that high concentrations of DAMPs at tissue damage sites could inhibit localized immune responses, given that these concentrations are higher than those found in circulation, there is currently limited evidence on their impact on tissue-resident immune cells.
Nevertheless, a few studies have demonstrated that DAMPs may impair the ex vivo function of T cells and macrophages and promote the expansion of tissue-resident suppressor cells derived from myeloid-derived suppressor cells (MDSCs). These findings suggest that the release of injured tissues may contribute to immune dysfunction across affected tissue compartments following severe trauma. 11
DAMPs in tissues have been linked to immunosuppressive characteristics. 12 However, a limited number of studies have demonstrated a role for DAMPs in the impaired ex vivo function of T cells and macrophages, as well as in promoting the proliferation of tissue-resident suppressor cells derived from MDSCs.
DAMPs Generated from Cytosol
Heme
Heme, an iron-containing porphyrin, is synthesized in the cytoplasm and mitochondria of erythrocytes. 13 It is a vital component of many biological functions, including oxygen transport and storage, electron transfer, and cellular metabolism. Heme serves as a prosthetic group for hemoproteins such as hemoglobin, myoglobin, and cytochrome P450.
However, when unbound heme is released from hemoproteins, it can catalyze the formation of highly reactive oxygen radicals, stimulate the complement cascade, and enhance toll-like receptor (TLR)-mediated immune responses. These processes contribute to oxidative stress, inflammation, and tissue damage. 14
In critical illness, elevated levels of circulating cell-free heme have been associated with the SIRS and organ injury response and organ destruction. The primary regulatory of circulating cell-free heme, which reduces its cytotoxic effects, is the endogenous heme-binding plasma protein hemoperin. 15 In addition, three heme/hemoglobin-binding plasma proteins—albumin, α−2-macroglobulin, and haptoglobin—aid in neutralizing, recycling, and clearing free heme and iron. 16
Heat shock proteins
HSPs are a broad family of molecular chaperons involved in protein folding, stabilization, and transport. Constitutively expressed HSPs can be passively released from necrotic tissue or actively secreted through nonclassical protein release pathways, either as free proteins or packaged within extracellular vesicles. These stressors include heat shock, toxic gases such as cigarette smoke, and ultraviolet.
HSPs are highly immunogenic and have been associated with the pathophysiology of several chronic inflammatory diseases. Patients with polytrauma and traumatic brain injury (TBI) have been found to have significantly higher levels of HSP60, HSP70, HSP72, and/or HSP90-α in their blood compared with healthy controls. 17 Following trauma, intracellular expression of HSPs also increases. Neutrophils and monocytes from patients with severe burns and trauma have been shown to express HSP27, HSP32, HSP60, HSP70, and HSP90 in response to injury.
Therapeutic studies involving HSP70 have shown that it can enhance the immunosuppressive function of regulatory T cells (Tregs), induce a tolerogenic phenotype in monocyte-derived dendritic cells (DCs), inhibit the maturation of DC, and suppress the development of T helper 1 cells, ultimately leading to reduced T cell responses. 18
Given the substantial evidence supporting the immunosuppressive properties of HSPs, and the hypothesis that their previously reported pro-inflammatory effects may be attributed to contamination with LPS, it is increasingly believed that HSPs act more as “DAMPERs” (immune suppressors) than classical DAMPs (immune activators). 19
Proteins called S100
The cytoplasm of leukocytes, microglia, and astrocytes contains a family of calcium-binding proteins known as S100 proteins, which regulate cellular division, metabolism, and proliferation. 20 Patients with burns, polytrauma, and TBI have been reported to exhibit elevated levels of S100A8, S100A9, and S100B in their bloodstream upon hospitalization. These proteins are recognized by the receptor for advanced glycation end-product (RAGE), the pattern recognition receptors (PRRs), TLR2 and TLR4, and their elevated levels have been observed to persist for up to 7 days. Prospective studies have shown that patients with poor clinical outcomes—such as mortality or long-term disability—exhibit significantly elevated levels of S100 proteins as early as the first day post-injury. 21 These protein levels show positive correlations with the Glasgow Coma Scale score, percentage of total body surface area burned, and injury severity score. The pro-inflammatory and antiapoptotic properties of S100 proteins, such as neutrophil activation, leukocyte transepithelial migration, and monocyte-driven amplification of tumor necrosis factor (TNF) production, enhance the SIRS responses and may explain why injury-induced elevations in S100 proteins are associated with unfavorable clinical outcomes. 22
It has been demonstrated that S100A8, S100A9, and/or calprotectin inhibit neutrophil chemotaxis and the production of neutrophil reactive oxygen species (ROS) induced by LPS and phorbol 12-myristate 13-acetate (PMA). This inhibition is attributed to adenosine metabolite generation mediated by S100A8 and S100A9. They also suppress T cell priming by reducing DC antigen presentation. Elevated S100 protein levels at sites of tissue damage may create a localized immunosuppressive environment in addition to their direct effects on immune cells. Both calprotectin and S100A8 homodimers have been shown to act as chemoattractants for MDSCs. Upon activation, MDSCs synthesize and secrete S100A8/A9 heterodimers. 23 Thus, S100 proteins from damaged tissue may attract MDSCs, whose activation and subsequent calprotectin production exacerbate immune dysfunction and promote further immunosuppression, creating a vicious postinjury cycle of immunosuppression. 24 Another nuclear-derived DAMP, CIRP, functions as an RNA chaperone that, under normal conditions, resides in the nucleus and regulates mRNA translation and cell proliferation. However, extracellular CIRP (eCIRP) has been shown to enhance the cytotoxic profile of cluster of differentiation 8 (CD8)+ T cells, stimulate CD4+ T cell differentiation into Th1 cells, and promote neutrophil transcription of intercellular adhesion molecule as well as extracellular trap formation. eCIRP also induces TNF production, ROS generation, and extracellular trap formation by macrophages and monocytes through TLR4, triggering receptor expressed on myeloid cells-1 (TREM-1), and NF-κβ signaling pathways. 25 Therefore, it is not surprising that eCIRP enhances systemic inflammatory responses in in vivo models of sepsis and hemorrhage, which lead to organ dysfunction, damage, and higher mortality. 26 CIRP has been detected in the bloodstream of critically ill patients in intensive care units, as well as in the serum of rats subjected to hemorrhagic shock and cecal ligation and puncture. As observed with other DAMPs, innate immune cells preexposed to CIRP exhibit diminished responses upon secondary stimulation. When eCIRP binds to the IL-6 receptor (IL-6R), it activates the signal transducer activator of transcription3 (STAT3), explaining the observed endotoxin tolerance. This eCIRP-induced macrophage tolerance was reversed by using anti-IL-6R antibodies or pharmacological inhibitors of STAT3. Moreover, arginase-1 (ARG-1) and CD206 expression levels were elevated in the (RAW264.7) macrophage cell line treated with eCIRP, indicating its role in promoting M2 anti-inflammatory macrophage polarization. 27 Interestingly, these cells also secreted HMGB1, another DAMP capable of inducing endotoxin tolerance, alongside inflammatory cytokines. 28
Interleukin-33
IL-33, a member of the IL-1 cytokine family, is constitutively produced by epithelial, endothelial, and stromal cells, and its expression increases in macrophages and DCs during inflammatory and infectious events. IL-33 is a dual-function protein that resides in the nucleus and acts as both a cytokine and transcriptional repressor. 29 When released into the extracellular environment, either through active secretion or cellular damage, IL-33 binds to its receptor, suppression of tumorigenicity (ST2). 30 Activation of ST2 signaling pathway leads to T cell polarization, degranulation, and cytokine release. ST2 is expressed by various immune cells, including mast cells, eosinophils, Th2 cells, group 2 innate lymphoid cells, and natural killer (NK) cells. In contrast to a mixed control population of polytrauma patients and healthy individuals, plasma IL-33 concentrations were found to be significant 6 to 72 h postinjury in an independent study of 14 polytrauma patients. 31 Although it remains uncertain whether injury severity directly influences IL-33 levels, variations in IL-33 concentrations have been observed among patients with differing clinical outcomes. CD16+ granulocytes isolated from patients with blunt or penetrative trauma have been shown to exhibit increased (ARG-1) expression and enzymatic activity for up to 2 weeks following injury, linking these findings to trauma-induced immune changes. 32 By promoting the development of M2 macrophages, which release the anti-inflammatory cytokine IL-10, and facilitating the accumulation of Tregs, IL-33 has been proposed as a potential mediator of long-term immunosuppression in murine models of sepsis. 33
The cellular process of inflammation is triggered by physicochemical injury, such as infection or metabolic stress. This cycle includes cell death for homeostasis, programmed necrosis, apoptosis, secondary necrosis, immune cell activation, DAMP release, and inflammation. The body responds to injury and stress at the cellular level through both cell death pathways and immune activation. The release of inflammatory mediators, including IL-1α, IL-33, IL-18, and HMGB1, sustains this cycle, illustrating the body’s cellular response to injury and stress, as shown in Figure 1.
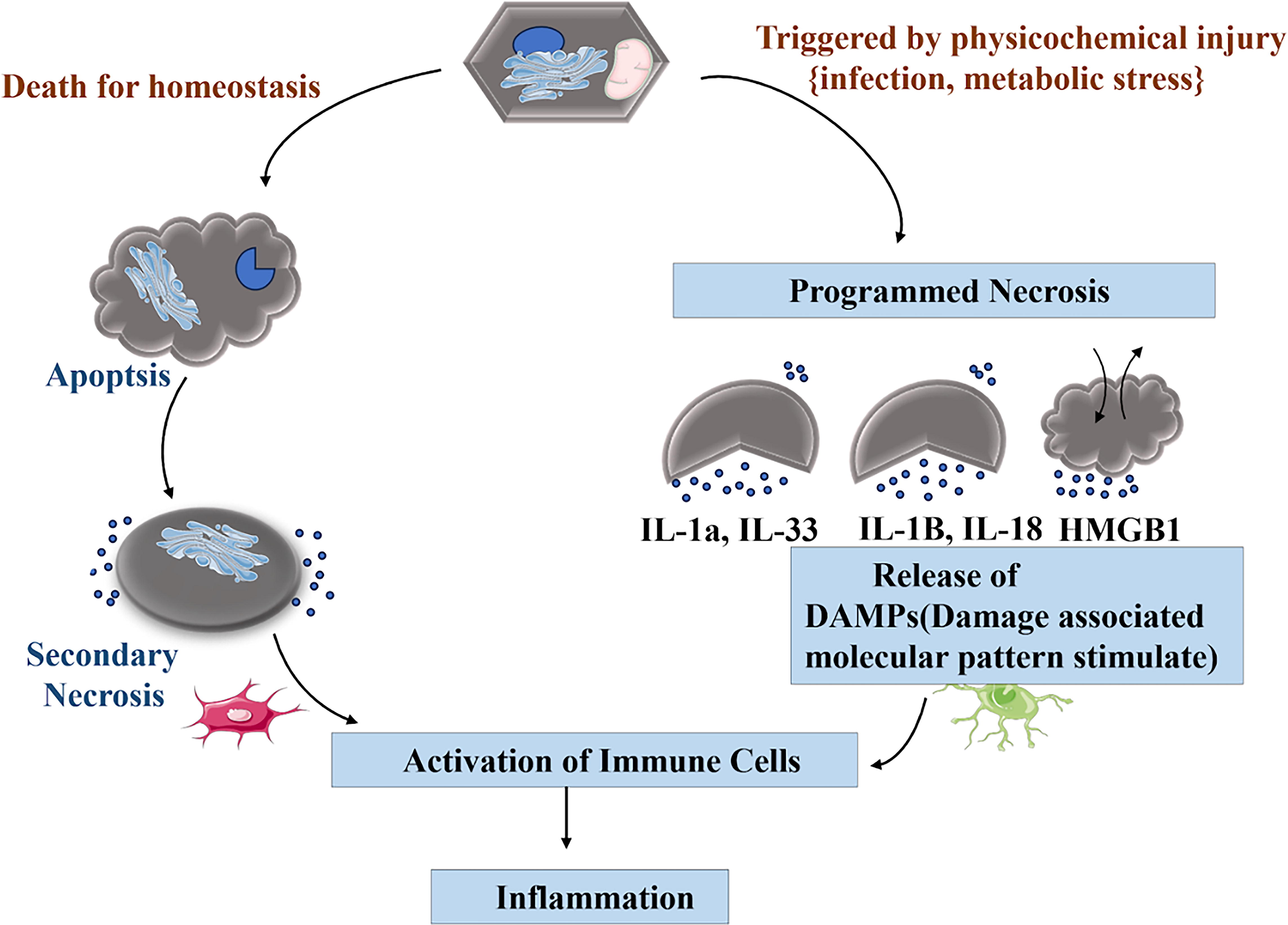
The image describes the process of ferroptosis, a form of regulated cell death distinct from apoptosis or necrosis. It is characterized by iron-dependent lipid peroxidation.
High-mobility group box 1
The nuclear-resident, DNA-binding chaperone protein HMGB1 controls the transcription of genes. In the minutes, hours, and days following severe trauma, HMGB1 concentrations are noticeably elevated.34,35 It can reach the bloodstream through a secretory vesicular route, necrosis, pyroptosis, or active secretion. Poor clinical outcomes, including multiple organ dysfunction syndrome (MODS), organ damage, and mortality, have been linked to elevated levels of HMGB1. These elevated levels exhibit a favorable correlation with pro-inflammatory mediators (e.g., MPO, IL-6, IL-8), base deficit, and the severity of the damage. TLR2, TLR4, TREM-1, and RAGE are among the PRRs that recognize HMGB1. HMGB1 has the ability to both stimulate and suppress immune cell activity, depending on its redox state. 36 The immune-suppressive actions of HMGB1 are caused by the terminal oxidation of three cysteine residues located in the protein’s acidic tail and DNA-binding domains. Similar to many other DAMPs, HMGB1 suppresses the pro-inflammatory responses of monocytes. Reduced phosphorylation of the nicotinamide adenine dinucleotide phosphate oxidase component p40phox is associated with this impairment. It has been shown that cultivating neutrophils from healthy controls with sera from critically ill septic shock patients significantly reduces ROS production by these cells, which has practical implications. HMGB1 has been demonstrated in numerous studies to support MDSC activation, migration, survival, and differentiation. 37 A greater number of MDSCs have been observed in the bone marrow, blood, and spleen of mice that have experienced peripheral tissue trauma. After the injury, treatment with an inhibitory anti-HMGB1 antibody can prevent this increase. A significant rise in ARG-1 expression in CD11b+ monocytes has also been associated with an increase in spleen-localized MDSCs in a rat model of neurological damage. The reduction in MDSC frequency in RAGE−/− mice and the absence of the trauma-induced ARG-1 upregulation in monocytes from mice pretreated with an anti-HMGB1 antibody indicate a role of this DAMP in mediating systemic immunosuppression. 38
Although not all studies have found this, existing research suggests that ATP is a DAMP with potent pro-inflammatory effects and initiates the SIRS response. For example, signaling through ATP and the P2 family of purinergic receptors induces the production of TNF, IL-6, and IL-1α by macrophages, activates T cells, enhances IL-2 secretion, increases superoxide production by neutrophils in response to N-formylmethionine-leucyl-phenylalanine, and promotes inflammasome-mediated release of IL-1β by macrophages primed with LPS. In addition, in a mouse model of LPS-induced shock, systemic administration of the ATP-hydrolyzing enzyme apyrase reduced the cytokine storm, immune cell infiltration into peripheral organs, and apoptotic cell death observed in littermates treated with a vehicle control. 39 According to this research, poststress immunosuppression may be mediated by interactions between ATP, mitochondrial DNA (mtDNA), HSPs, and heme metabolism. In a small cohort study of 11 trauma patients, the frequency of CD8+ CD39+ T cells was positively correlated with plasma mtDNA levels. This suggests that mtDNA sensing may enhance T cell-mediated conversion of ATP/ADP into its immune-suppressive metabolite, adenosine. In a series of in vitro experiments, it was demonstrated that adenosine and HSP32 form a positive feedback loop that prevents macrophages from producing TNF in response to LPS. 40
The pathogenesis of chronic inflammatory processes in chronic obstructive pulmonary disease (COPD) involves a sequence of cellular and molecular events that lead to airway inflammation. Damaged airway epithelial cells release DAMPs, which activate PRRs, leading to triggering immunogenic cell death (ICD) and the release of pro-inflammatory cytokines. These cytokines activate various immune cells, thereby sustaining chronic inflammation in COPD. The diagram provides a simplified overview of the complex mechanisms underlying COPD-related inflammation, highlighting the role of environmental factors, cellular damage, and the immune system in disease progression, as shown in Figure 2.
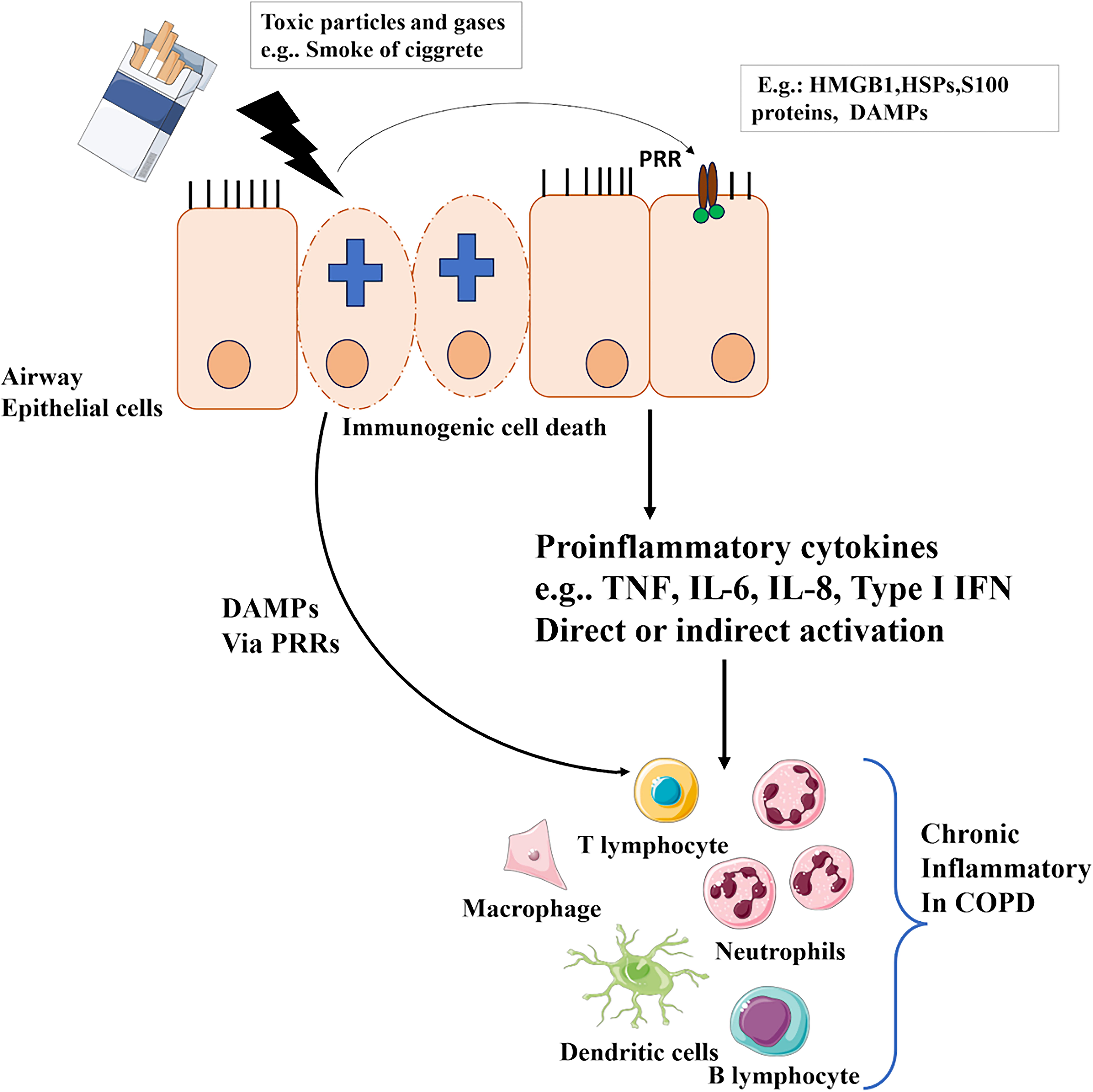
The image depicts the process of inflammatory cell death, specifically in the context of chronic obstructive pulmonary disease (COPD).
Mitochondrial DNA
Exposure to this DAMP induces the synthesis of IL-8 and activates mitogen-activated protein kinase. MtDNA levels increase abruptly and permanently after trauma. Further research has validated these findings and demonstrated that mtDNA also triggers neutrophil degranulation and neuroendocrine tumor (NET) formation in a TLR9-dependent manner. While plasma mtDNA levels positively correlate with severity in models of trauma and shock, injecting mtDNA into mice induces hepatic inflammation.41,42 Furthermore, the mtDNA burden is significantly higher in nonsurvivors and in patients who develop complications such as acute respiratory distress syndrome (ARDS) and MODS compared with survivors. Interestingly, there are two types of circulating cell-free mtDNA (cf-mtDNA): a larger form that constitutes approximately 95% of total cf-mtDNA and a low-molecular-weight form. 43 Low-molecular-weight cf-mtDNA levels were positively associated with poor clinical outcomes, including longer stays in the intensive care unit, increased use of a ventilator, and extended duration of MOF, according to comparison of these two fractions in plasma samples from 25 patients who had experienced significant trauma. 44 Patients with sterile brain trauma who developed health care-associated infections exhibited markedly higher circulating levels of mtDNA. By promoting cortactin (CTTNs) degradation via histone deacetylases through the activation of G protein-coupled receptor kinase 2 (GRK2), mtDNA treatment inhibits CTTN activation. Neutrophils isolated from trauma patients have shown reduced CTTN acetylation and increased GRK2 activation, both of which correlate with elevated plasma mtDNA levels. In traumatized neutrophils, GRK2 activity was elevated and CTTN acetylation was reduced during days 1 to 6 postinjury, suggesting the potential clinical relevance of these findings. 45 In addition, mtDNA exerts a detrimental effect on DC function, which may compromise the effectiveness of adaptive immune responses. 46
N-formylated peptides produced from mitochondria
Mitochondria synthesize 13 formylated peptides that are molecularly identical to the formyl peptides produced by bacteria. All but one of these peptides, specifically Cox1, have been detected in the bloodstream of trauma patients. Among them, the concentration of ND6, a subunit of mitochondrial NADH dehydrogenase, increases significantly within an hour of injury and continues to rise for up to 72 h. Neutrophils are attracted to mitochondrial N-formyl peptides (mtFPs), via surface-expressed FP receptors, FPR1 and FPR2. The molecular basis for ND6-mediated downregulation of G-protein-coupled receptors, which leads to translational receptor desensitization and a reduction in antimicrobial activity, may involve GRK2 activation. In animal models, peripheral fracture damage has been shown to reduce lung neutrophil recruitment following pulmonary contusion, which is associated with impaired bacterial clearance. 47 In vitro replication of these impairments revealed that phorbol myristate acetate (PMA)-induced NET formation was inhibited when control neutrophils were treated with whole mtDAMP preparations, but not with purified mtDNA. This deficiency is associated with reduced aerobic glycolysis and mtFP-dependent activation of AMP-activated protein kinase, a negative regulator of NET formation. 48
Research has examined mtFPs, a type of DAMP that suppresses several neutrophil antimicrobial functions, as a potential therapeutic target to reverse immunosuppression following trauma. Several studies have shown that pretreating neutrophils with FPR1 inhibitors not only prevents mtFP-induced deficiencies in calcium mobilization and chemotactic responses but also reduces ND6-mediated downregulation of GPCRs. 48 Another strategy under investigation is the intratracheal instillation of neutrophils, which leverages mtFPs generated at sites of tissue injury to redirect neutrophils from regions of pathogenic challenge. In preclinical models of pseudo-fracture and bacterial inoculation, intratracheal administration of exogenous bone marrow-derived murine neutrophils or neutrophils isolated from human peripheral blood has been shown to prevent and/or treat pneumonia by reversing the mtDAMP-induced impairment in bacterial clearance from the lungs. 49
ICD refers to the death of cancer cells triggered by the initiation of an immunomodulatory response. The enhanced immunogenicity of cancer cells undergoing ICD is what characterizes this form of regulated cell death. In cancer treatment, ICD plays a critical role in enhancing immune system activation. 50 Therefore, ICD may represent a novel strategy to boost immune responses against cancer by generating DAMPs. Several conventional and chemotherapeutic agents, along with preclinically studied compounds derived from natural sources, have demonstrated immunostimulatory properties through the induction of ICD. 51 Due to their low toxicity, affordability, and ability to inhibit cancer cells by targeting multiple pathways essential to cancer progression, natural compounds have attracted considerable interest in cancer therapy. Identifying natural substances (NSs) capable of inducing ICD therefore holds a promising potential for cancer immunotherapy. Because naturally occurring compounds trigger DAMPs and cancer-related stress responses, they are believed to function as immunoadjuvants. Acute exposure to DAMP monomers can activate DCs and other antigen-presenting cells (APCs). This in turn stimulates NK cells and cytotoxic T lymphocytes (CTLs) to take further immune action. Although using natural compounds as inducers of ICD may be a promising approach, it is critical to understand the factors that determine a compound’s suitability for this purpose. In this context, the authors discuss how various ICD inducers affect DAMP release and the activation of signaling cascades that initiate immune responses. 52 They primarily focused on natural agents, including plant-derived substances, marine molecules, and bacterial products. In addition, the potential of synthetic drugs to induce ICD is also discussed. Cancer remains one of the major global challenges to increasing life expectancy. Recently, numerous anticancer therapies have been developed to activate ICD, which alerts the immune system to the presence of dying cancer cells. ICD, a unique form of cell death, triggers T cell-mediated immunity in response to antigens released by dead cells. To trigger an immune response, ICD is initiated through the translocation and release of various DAMPs, which can be promoted by treating cells with specific agents. 53
The Main Characteristics of ICDs
As previously mentioned, DAMPs are intracellular molecules released during the ICD process. When DAMPs are exposed to or released by dying cells, they exhibit immunostimulatory effects. DAMPs are also released during necrosis in inflammatory or pathological conditions. Multiple lines of evidence demonstrate that cancer cells exposed to radiation or chemotherapy can generate DAMPs and attract and activate immune cells. In addition, DAMPs act as signals that stimulate CTLs to eliminate cancer cells by promoting phagocytosis. 54 ROS and endoplasmic reticulum (ER) stress contribute to the generation of DAMPs during ICD. There are three major subclasses of DAMPs based on their location or site of release: DAMPs that occur on the cell surface (such as HSP 90, HSP 70, and Rtg), extracellular DAMPs (such as pro-inflammatory cytokines such as TNF-α, IL-10, IL-8, HMGB1, and uric acid), and higher levels of HMGB1 (Table 1) in patients with squamous cell carcinoma of the esophageal were associated with improved survival in those receiving chemoradiotherapy.
Modulating Damage-Associated Molecular Patterns for Cancer Therapy: Mechanism and Effects on Antitumor Immunity
DAMPs, damage-associated molecular patterns; TLR4, toll-like receptor 4; RAGE, receptor for advanced glycation end-product.
CRT is one of the most well-known DAMP located in the lumen of the ER. It plays a crucial role in immunity by contributing to chaperone activity and Ca2+ homeostasis. CRT also regulates various cellular processes, including nuclear transport, invasion, adhesion, protein synthesis, and cell proliferation. On the surface of apoptotic cells, CRT acts as a general recognition ligand. Increasing evidence suggests that CRT strongly influences the immunogenicity of dying cells and enhances antitumor immune responses. This was accomplished through lowering CRT levels and cell surface exposure. CRT exposure improves the ability of CTLs and DCs to recognize and eliminate tumor cells. 55
Heat Shock Proteins
Another key feature of ICD is the involvement of HSP, which facilitates protein folding and refolding under stress conditions. When cells are exposed to stress (oxidative stress, radiation, or chemotherapy medications), HSPs are overexpressed within the intracellular environment and translocated to the plasma membrane. Among the various HSPs, HSP70 and HSP90 are the most commonly associated with ICD. These proteins perform different functions in cancer, depending on their subcellular localization. 56 Intracellular HSPs are primarily protective, while membrane-bound or extracellular HSPs play immunological roles. The mobilization of intracellular HSPs to the plasma membrane leads to strong immunostimulatory activity. HSP70 and HSP90 exhibit immunostimulatory effects when they interact with the extracellular surface of dying cells. Ecto-HSP70 and HSP90 regulate the immunogenicity of dying cells by engaging with APC surface receptors such as lectin-like receptors, oxidized low-density lipoprotein receptor-1 (LOX1), CD40, and CD91. 57
HMGB1 Is Released
There is growing evidence that extracellular HMGB1 can activate anticancer immune system responses during ICD. DNA molecules can be bound and modified by three members of the HMGB family—HMGB1, HMGB2, and HMGB3. The first known member of the family, HMGB1, is a nonhistone chromatin-binding protein. Its subcellular location determines its function. 58 While extracellular HMGB1 is involved in angiogenesis and chemotaxis, nucleus HMGB1 mediates several processes, such as transcription, recombination, nucleosome maintenance, and DNA repair. Another DAMP released during ICD is ATP. The release of ATP from dying cells is an essential component for the successful initiation of ICD. Lysosomal-associated membrane protein 1 (LAMP1) is required for the mechanism that facilitates ATP release following its translocation from lysosomes to autolysosomes in response to ICD inducers. LAMP 1 then translocates to the plasma membrane in a caspase-dependent manner. ATP secretion is also mediated by caspases and pannexin 1 (PANX1). PANX1 is required for the translocation of LAMP1 to the cell surface and for lysosomal exocytosis during ICD. 59 For APCs and their progenitors, extracellular ATP acts as a strong chemotactic agent by binding to purinergic receptors P2RY2 and P2RX7.
The figure illustrates the process of DAMP release and the subsequent immune response, particularly in cancer treatment. It shows how dying tumor cells release DAMPs, which activate immune cells and initiate an antitumor immune response. This involves tumor cell death and DAMP release, which activates DCs. These activated DCs present tumor antigens to T cells and NK cells, activating cytotoxic immune cells that target and destroy the remaining tumor cells. The process also includes feedback and regulation through signaling molecules and receptors that modulate the immune response. This complex biological process highlights the key events in DAMP-mediated antitumor immunity, as shown in Figure 3.
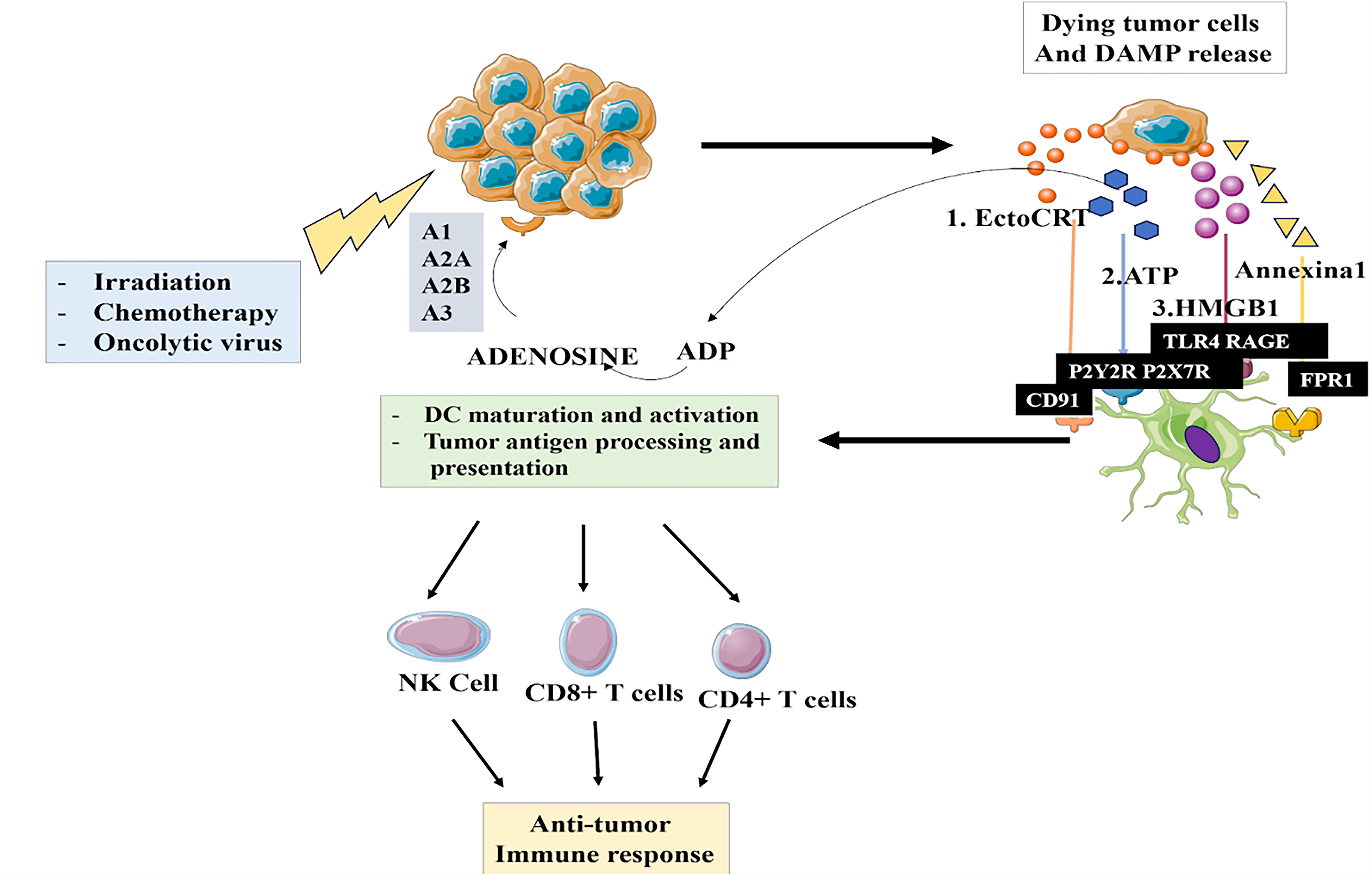
The image depicts the process of how dying tumor cells release damage-associated molecular patterns (DAMPs), which initiate an antitumor immune response.
Type I Interferon
Type I interferon (IFN) plays a critical role in ICD. It significantly influences both innate and adaptive immunity. Type I IFN promotes antigen presentation and enhances T cell-mediated antitumor activity. ICD-inducing factors cause tumor cells to upregulate genes involved in the type I IFN response pathway. These factors include radiation, oncolytic viruses, and anthracyclines. An IFN-I-induced gene, chemokine ligand 10, acts as a chemotactic factor, recruiting immune cells to target and kill cancer cells. Moreover, a type I IFN-related gene expression profile can predict patient responses to anthracycline-based chemotherapy. Annexin A1 binds phospholipids in the presence of calcium.60,61
ICD Inducers: An Emphasis on Organic Substances
Agostinis et al. proposed the first classification scheme for ICD inducers in 2013. ICD inducers are divided into categories, based on whether they target the ER directly (type II) or indirectly (type I). directly. Type II ICD inducers directly target the ER and more directly enhance immunogenicity. However, available type II inducers are extremely limited. For example, hypericin localizes primarily in the ER and, upon light activation, induces ROS-driven ER stress, which leads to ICD. 62
Natural Substances
NSs are organic compounds derived from living organisms, such as plants, fungi, and marine organisms. They exhibit remarkable structural and functional diversity and have been a rich source of bioactive molecules for drug discovery and development. 63 Plants, in particular, are a diverse source of NSs and have been used in traditional medicine systems such as Ayurveda, Chinese medicine, and Unani medicine. They produce a variety of secondary metabolites, including alkaloids, flavonoids, terpenoids, phenolics, and glycosides, which have a range of biological activities and therapeutic potential. 64 NSs and their derivatives are gaining attention as potential sources of novel anticancer drugs due to their low toxicity, affordability, and availability. Natural anticancer substances can remodel the tumor microenvironment and overcome drug resistance in cancer immunotherapy by targeting and regulating immune cells, such as T cells, macrophages, mast cells, and inflammatory cytokines within the tumor microenvironment. 65 Personalized therapy is one of the most promising applications of precision medicine in immune-related disorders. NSs can be used as personalized immunotherapy by targeting specific genes, proteins, and pathways involved in immune regulation, thereby triggering specific immune responses. Synergistic effects observed when NSs are combined with conventional anticancer treatments may improve outcomes and reduce side-effects. 66
Dual-action compounds are NSs that exhibit both stimulatory and suppressive effects on immunity. The compounds can modulate the immune system in a balanced, context-dependent manner, depending on factors such as type, dose, and exposure duration, as well as the phenotype and functional state of immune cells. 67 Dual-action NSs are useful in treating chronic inflammatory diseases such as atherosclerosis, obesity, and neurodegeneration, and they can enhance vaccine efficacy. 68 Other NSs derived from fungi, insects, and marine organisms exhibit diverse therapeutic potentials, including antimicrobial, anticancer, antiviral, and immunosuppressive properties. Further research is needed to determine optimal combinations, dosages, and treatment schedules to maximize synergistic effects and advance personalized cancer therapy. 69
Plant-Based Substances
Natural ICD inducers include capsaicin, alternol, curcumin, silibinin, wogonin, ginsenoside, alantolactone, gallotannin-rich fractions, shikonin, lentinan, chalcones, coumarin, OT52, plumbagin, cardenolides, bufadienolides, cardiac glycosides (CGs), and colchicine. 70 Capsaicin, a pungent compound found in chili peppers, has pharmacological effects, including lowered blood pressure, enhanced immunity, and reduced pain and inflammation. Alternol, derived from microbial fermentation, induces ICD in prostate cancer cell lines by increasing pro-inflammatory cytokines, DAMPs, and tumor-targeted immune responses. Curcumin, found in turmeric, has been used for generations to treat inflammatory diseases. 71 Silibinin, isolated from milk thistle, inhibits p-STAT3 in cancer cells and promotes apoptosis. Wogonin, a plant-derived flavonoid, has immunomodulatory, anticancer, antioxidant, and anti-inflammatory properties. Ginsenosides, derived from ginseng, destroy tumor cells and modulate the immune system. 72 Alantolactone, from Chinese medicinal plants, includes ICD in microsatellite-stable colorectal cancer. Gallotannin-rich fractions from Caesalpinia spinosa stimulate apoptosis by externalizing annexin V and activating caspases 3 and 9. Shikonin promotes apoptotic pathways and inhibits melanoma cell proliferation.
Celastrol, known for its anti-inflammatory, anticancer, and antiobesity effects, can induce ICD at extremely low, nontoxic doses. Bullatacin, from Annona atemoya, is a potent anticancer compound. OT52, a coumarin compound, regulates inflammation and exhibits anti-inflammatory, anticancer, and antioxidant properties. It inhibits non-small-cell lung cancer cell growth by suppressing STAT3 activation and alleviating ER and Golgi stress. 73
Plumbagin, a naphthoquinone from Salvia miltiorrhiza, enhances ICD by generating ROS. Nanodelivery of these compounds significantly improves their half-life and tumor-targeting efficiency in hepatocellular carcinoma mouse models. Cardenolides and bufadienolides are the two subtypes of CGs (type I ICD inducers). Digoxin, digitoxin, ouabain, and lanatoside are the most effective ICD inducers. Vesiculated A-tocopheryl succinate, a nontoxic form of vitamin E, can enhance DC-mediated cancer immunotherapy by directly killing tumor cells and promoting DC maturation via HSPs as danger signals. Colchicine, from Colchicum autumnale, plays a crucial role in cytokine signaling and ICD induction. 74
Marine-derived substances, including microalgae and fish oil, have been linked to ICD induction in cancer cells. These substances may be useful in cancer prevention and treatment, alone or in combination with other immunotherapy approaches. Microalgae-derived compounds have shown immunomodulatory and anticancer properties, although their role in ICD remains unclear. 75 Sulfavant A, derived from marine dinoflagellates, has shown efficacy in a melanoma vaccination mouse model. Alexandrium minutum produces a glycopeptide that includes mitophagy in cancer cells without harming healthy cells. Fish oil, which contains docosahexaenoic acid (DHA), enhances the cytotoxicity of several anticancer drugs by generating ROS and increasing cancer cell sensitivity. DHA has cardio-protective effects, which can be highly advantageous when combined with DOX. 76
Three diatoms—Skeletonema costatum, Thalassiosira rotula, and Pseudonitzschia delicatissima—produce polyunsaturated aldehydes that can cause necroptosis in colon and lung cell lines. The marine alkaloid lepadin A can induce ICD, leading to the overexpression of major histocompatibility complex class and its costimulatory factors, which are necessary for DC development and the activation of naive T cells, thereby boosting the effective immune response. MHO7, a substance originating from marine sources, acts as a potent inducer of ICD via the ER stress-C/EBP-homologous protein cascade, offering potential treatment for triple-negative breast cancer. Microalgal compounds from Spirulina maxima and Schizochytrium sp. influence the microbiota, and recent studies have shown that using microalgal species as dietary supplements may exert immunomodulatory effects. 77 Bacterial compounds can also induce ICD in cancer cells. For example, lactation-induced cell death is associated with in vitro ICD markers. Septacidin, an antibiotic produced by Streptomyces fimbriatus, induces ICD by promoting HMGB1 expression, ATP release, and CRT exposure in both mouse fibrosarcoma cells and modified human osteosarcoma cells. Patupilone, a therapeutic compound isolated from the Sorangium cellulosum bacterium, can cause ICD and promote CRT translocation in a mouse model, highlighting its potential as a vaccination agent in immunocompetent mice.68,78
DAMPs or Endogenous Ligands
The activation of the immune system in response to tissue damage and pathological cell death is triggered by endogenous ligands, also known as DAMPs. These molecules include nuclear and cytosolic proteins such as HSPs and HMGB1, components of the extracellular matrix such as fibrinogen and hyaluronan, mitochondrial DNA from damaged or fragmented organelles, and constituents of the plasma membrane. Galectins, particularly galectin-3, have been recognized as important mediators of inflammation due to their ability to bind to TLRs, especially TLR-4, thereby influencing immune cell activation and tissue damage responses. PRRs, such as TLRs, serve as the primary sensors that detect DAMPs. The recognition of DAMPs by TLRs triggers signaling cascades that lead to the production of type I IFNs, pro-inflammatory cytokines, and other immune mediators, facilitating an effective immune response to tissue injury. To maintain homeostasis and defend against a variety of disease states, the immune system uses a complex mechanism to distinguish self from non-self and to differentiate between distinct forms of cell death.
The tumor-specific immune response involves DCs and T cells and is essential for recognizing and eliminating tumor cells. This response begins when dying tumor cells release tumor-associated antigens and DAMPs, which initiate immune activation. DCs capture tumor antigens and present them to naive T cells, leading to the activation of both CD8+ and CD4+ T cells. Upon activation, CD8+ T cells differentiate into cytotoxic T lymphocytes (CTLs) that directly eliminate target cells, whereas CD4+ T cells support the immune response primarily through cytokine production. CLTs then recognize and eliminate tumor cells expressing specific tumor-associated antigens. Tumor cells may undergo ICD, which further amplifies the immune response. This cyclical process illustrates the intrinsic interplay among immune cells involved in antitumor activity, as shown in Figure 4.
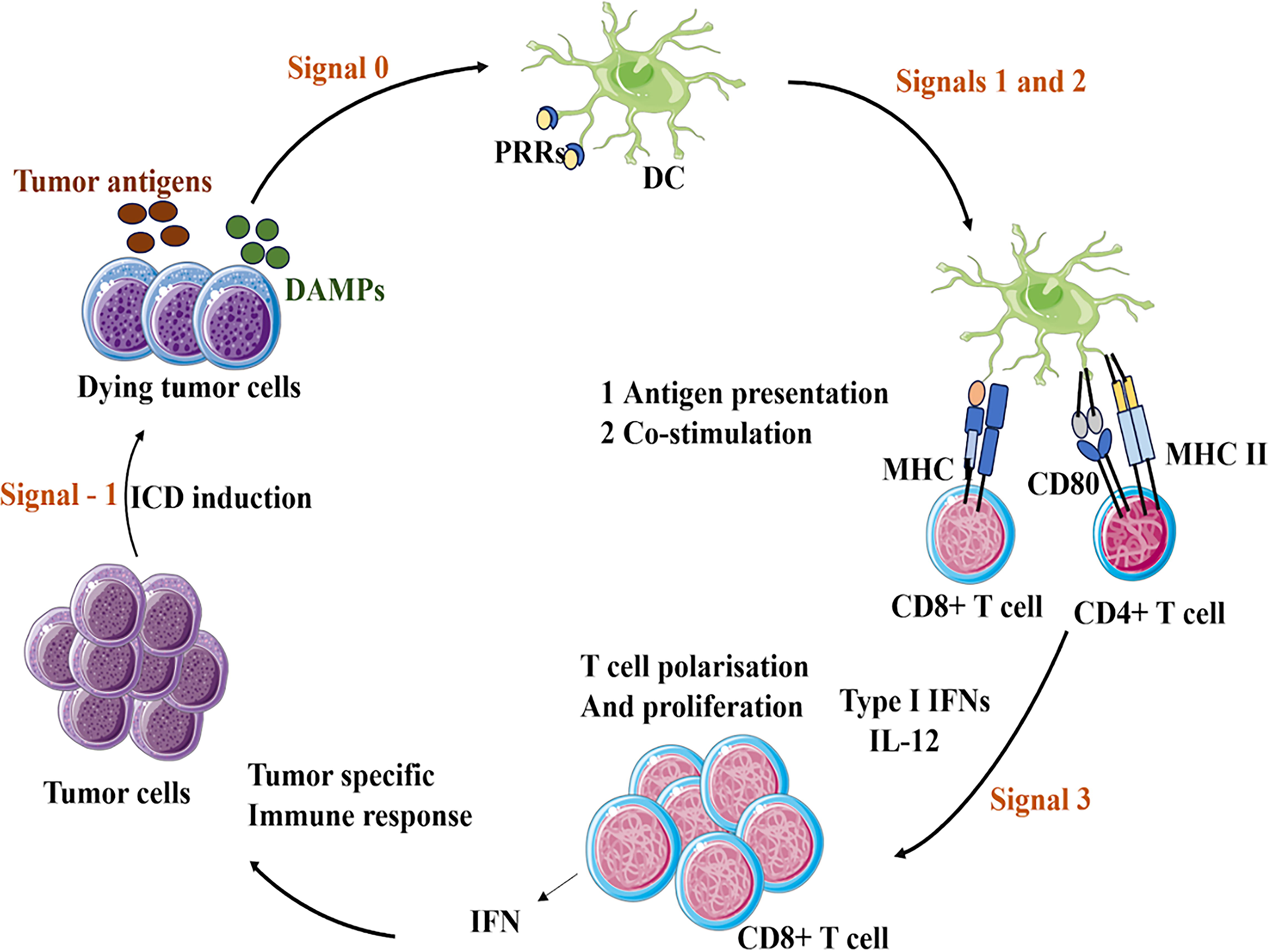
The image illustrates the mechanism of action for tumor vaccines, which are a type of immunotherapy that stimulates the body’s immune system to attack cancer cells.
TLRs are widely distributed across various cell types and tissues and play a crucial role in immune surveillance. Upon ligand binding, TLRs initiate a signaling cascade that leads to the production of type I IFNs and pro-inflammatory cytokines, both of which are essential for orchestrating the immune response. Immune homeostasis depends on the proper regulation of TLR signaling. Dysregulation is associated with various illnesses, including cancer, chronic inflammation, and autoimmune diseases, making TLRs promising targets for therapeutic intervention. The recognition of DNA fragments by TLR9 highlights the intricate cross-regulation within type I IFN signaling pathways, emphasizing the critical role of TLRs in the pathophysiology of lifestyle-related disorders. Studies demonstrating the connection between endosomal transmembrane (TM) TLRs (including TLR3) and various diseases that may activate the IFN signaling pathway support this finding. In conclusion, the critical role of TLRs in immune surveillance and their involvement in multiple immune and inflammatory pathways underscore the importance of tightly regulated TLR signaling. Their potential as therapeutic targets underscores the need for further research in this area and offers new avenues for the treatment of autoimmune, inflammatory, and chronic disorders. 79 TLRs, a class of proteins, constitute the first line of defense in the immune system. The leucine-rich repeat (LRR) structures of each TLR confer ligand specificity, enabling the recognition of specific PAMPs or DAMPs. DAMPs are molecules released by damaged or stressed cells that serve as danger signals to the immune system. DAMPs bind to PRRs on immune cells, triggering the production of pro-inflammatory cytokines and potentially contributing to a vicious cycle of inflammation, which can lead to tissue damage and the development of inflammatory diseases. This specificity allows the immune system to distinguish between various infections and injured host cells, thereby triggering appropriate immune responses. Members of the functionally diverse TLR family are located either on the cell membrane or within intracellular compartments such as lysosomes, endosomes, and the ER.80,81 Microbial membrane components such as lipoproteins, flagellin, and LPSs are the primary targets of cell membrane TLRs, including TLR2 (in heterodimeric interaction with TLR1 or TLR6), TLR4, TLR5, and TLR10. The system’s ability to adapt to various molecular signals indicative of infection or cellular damage is highlighted by the specialization of intracellular TLRs, such as TLR3, TLR7, TLR8, and TLR9, which are specialized in sensing nucleic acids. The 10 functional TLRs (TLR1–TLR10) in humans are each uniquely adapted to recognize distinct molecular patterns associated with pathogens and cellular damage, reflecting their role in immune surveillance and response. These receptors possess a single TM helix, a C-terminal cytoplasmic TIR domain, and an N-terminal domain with LRRs. 66
The Ligands for TLRs
According to both clinical and experimental research, TLR signaling enhances the immune response against allograft and xenograft antigens and influences susceptibility to autoimmunity. TLRs are expressed by nearly all immune cells, including NK cells, T cells, B cells, DCs, and macrophages. These receptors are essential for recognizing conserved molecular patterns associated with a wide range of pathogens, including viruses, fungi, and bacteria. TLRs are now recognized for their roles in activation and signaling. The host immune response against infections is triggered by intracellular downstream signaling pathways initiated when TLRs are engaged by various agonists. Immune responses to specific pathogens are generated when PAMPs interact with PRRs, leading to the production of pro-inflammatory cytokines and type-1 IFN. 82
TLRs’ Function in Cell-Mediated Immunity
As a vital bridge between innate and adaptive immunity, TLRs play a key role in mediating the activation and maturation of DCs and macrophages. Naive T cells are activated and differentiated into Th1, Th2, and cytotoxic T cell subtypes by professional APCs such as DCs. TLRs have a significant impact on DC function, which in turn influences the generation of effector T cell responses. TLR ligands such as LPS, CpG DNA, and lipoproteins can stimulate the production of costimulatory molecules. 83 TLRs function in regulating T cell activity.
Research suggests that TLRs play a crucial role in regulating T cell activity by acting as stimulatory receptors involved in memory preservation and antigen-specific T cell maturation. The functional state of various T cell subtypes is associated with TLR expression. TLR2 and TLR4 mRNA levels are typically low in naive CD4 T cells. However, TLR1, TLR2, and TLR4 are expressed on the surface of certain cells, including CD3-positive T cells. Notably, the surfaces of naive, effector, and memory T cells exhibit significant levels of TLR2 and TLR4. 84 TLRs as adjuvants in cell-mediated immunity are essential for activating the innate immune system, which subsequently initiates the adaptive immune response. To enhance immunity against infectious diseases, adjuvants play a crucial role as components of vaccines. Monophosphorylated lipid A (MPL), a modified form of LPS that activates TLR4, was the first TLR agonist to be used as a vaccine adjuvant. In 2009, the Cervarix vaccine was approved, incorporating AS04, an adjuvant system that combines aluminum salts with MPL, an additional TLR agonist. 75
DAMPs play a crucial role in inflammatory diseases, particularly in the context of stress and tissue injury. They are released from damaged or stressed cells and interact with PRRs on immune cells, initiating an inflammatory response. This response, which can be triggered by pro-inflammatory cytokines, may lead to the development or worsening of various inflammatory diseases, such as psoriasis, rheumatoid arthritis, and cardiovascular diseases. DAMPs can serve as biomarkers for disease progression and severity, and they represent potential therapeutic targets for managing inflammatory conditions by disrupting the inflammation cycle. The direction of DAMPs in blood samples from critically injured patients provides a mechanistic explanation for the simultaneous development of inflammation in such cases, as shown in Figure 5.
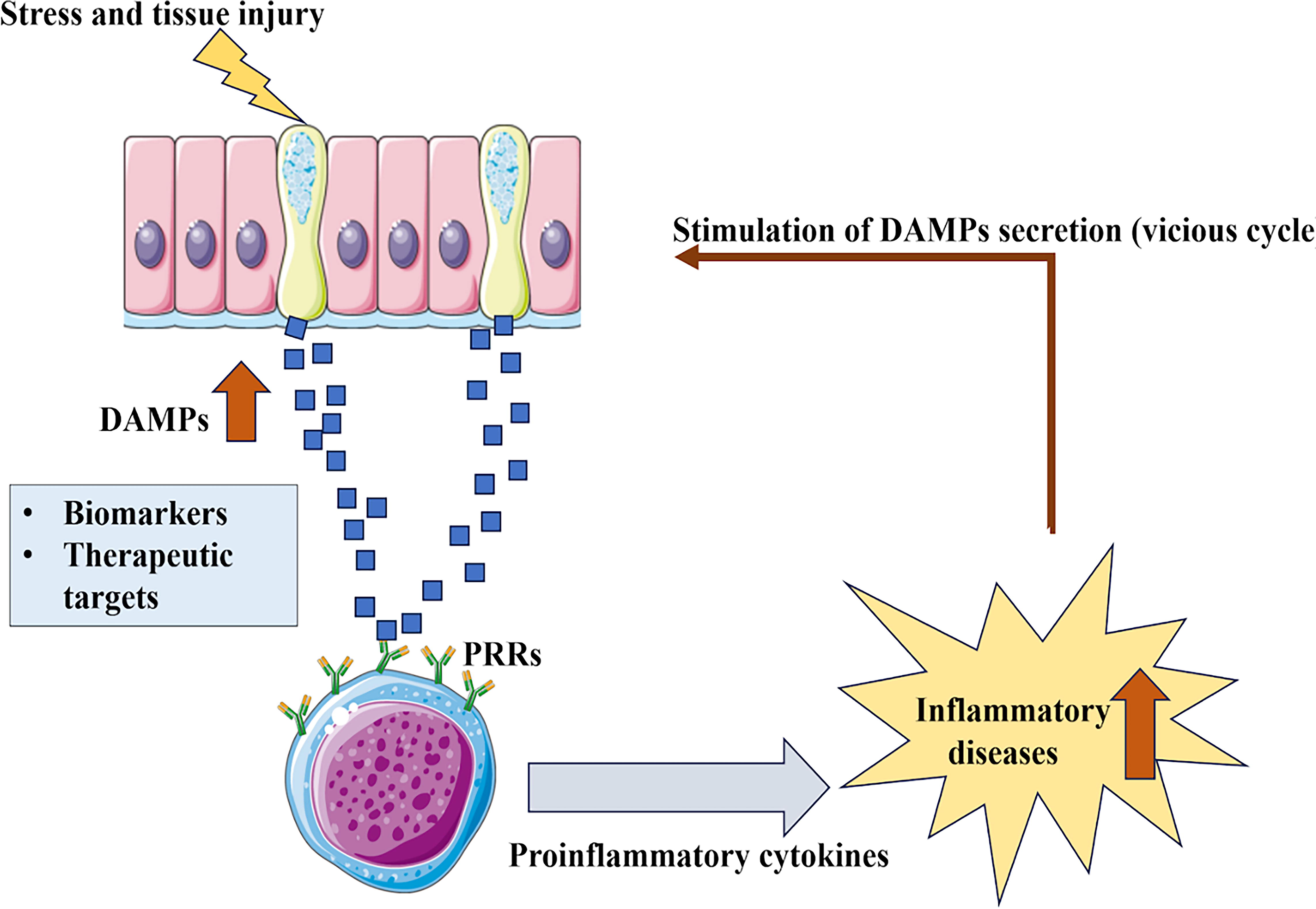
The image depicts the role of damage-associated molecular patterns (DAMPs) in inflammation. They act as “danger signals” that alert the immune system to tissue damage or stress.
Conclusions
The discovery of DAMPs in blood samples obtained from critically injured patients in a prehospital setting provides a mechanistic explanation for the simultaneous development of SIRS and the compensatory anti-inflammatory response syndrome within minutes after injury. If DAMPs are relevant in trauma patients, the question arises whether they and/or their associated signaling pathways represent promising therapeutic targets for preventing or treating post-traumatic infections. Microbe-associated molecular patterns, along with various subclasses of activating DAMPs and suppressing DAMP (SAMPs), work together to regulate controlled pro-inflammatory responses, inflammation resolution, and uncontrolled inflammation during bacterial, viral, and fungal infections. Hyperinflammatory, hyperresolving, and nonresolving/persistent immune responses are key patterns observed in several infections. Three clinical examples illustrate the inflammation-hyper-resolution dynamics driven by counterbalancing SAMPs and the hyperinflammatory responses caused by excessive DAMP release: ARDS caused by severe virus-induced respiratory infections, sepsis triggered by life-threatening bacterial infections, and malaria caused by a highly destructive protozoan. The illness is generally considered to result from a complex interplay between environmental risk factors and predisposing genetic variations.
The “outside-in” model proposes that the disease is initiated by a peripherally driven inflammatory or autoimmune response targeting degraded myelin, while the “inside-out” model suggests that the disease originates from a primary cyto degenerative process of cells within the central nervous system, which subsequently triggers secondary reactive inflammatory or autoimmune responses against myelin debris. These two complementary preclinical models are currently under discussion for investigating the etiopathogenesis of the disease.
Footnotes
Acknowledgments
The authors are thankful to the School of Pharmaceutical Education and Research, Jamia Hamdard, New Delhi, and St. Pauls College of Pharmacy, Telangana, for encouragement and support.
Authors’ Contributions
Writing—original draft, conception, and design: R.S., S.B., M.S., and S.F.S. Data collection and writing—review and editing: D.V. and A.M. Drafting the editorial article: H.K. Writing—conceptualization, design and drafting, methodology, and supervision: R.S. and K.M.
Disclosure Statement
The author declares no conflicts of interest.
Funding Information
No funding was received for this article.