
Abstract
Background:
Colorectal cancer (CRC) development and therapy resistance are heavily controlled by the tumor microenvironment (TME). Although anti-PD-1 immunotherapy has significant therapeutic advantages, resistance remains a key challenge. Recent research has identified the gut microbiota as a key regulator of host immunity and checkpoint inhibitor effectiveness. Ultrasound (US) has emerged as a viable biophysical technique for improving medication and microbial delivery and controlling immune activation within tumors.
Objectives:
The purpose of this work was to assess the synergistic effects of US-assisted fecal microbiota transplantation (US-FMT) on TME remodeling and anti-PD-1 resistance in a CRC cell line-derived xenograft mouse model.
Materials and Methods:
Tumor-bearing mice were randomized into four treatment groups: vehicle control, anti-PD-1 alone, fecal microbiota transplantation (FMT) alone, and US-FMT plus anti-PD-1 therapy. Low-intensity focused US was utilized to promote microbial engraftment and intestinal permeability. Flow cytometry, ELISA, and transcriptome profiling were used to investigate tumor growth kinetics, immune cell infiltration, cytokine profiles, and TME-related gene expression.
Results:
In comparison with the other groups, US-FMT reduced tumor development and restored sensitivity to anti-PD-1 treatment. US facilitated beneficial microbial colonization, boosted CD8 T cell infiltration, and decreased immunosuppressive cell populations. Furthermore, US-FMT modified cytokine release and reduced pro-tumorigenic inflammatory mediators, reprogramming the TME to be immune-active.
Conclusions:
US-assisted microbiota manipulation is a unique and synergistic biotherapeutic method for reversing immunological resistance in CRC. The combination of US and FMT has translational promise for enhancing immunotherapy response and developing noninvasive cancer treatment techniques.
Keywords
Introduction
Colorectal cancer (CRC) is a complex disease that involves interactions between cancer cells, stromal cells, and immune cells within the tumor microenvironment (TME). 1 It became a major global health burden due to high incidence and mortality rates. 2 Immune cells, particularly T cells, play a critical role in the TME by regulating the immune response to cancer cells. 3 The balance between different T cell subsets, such as Th1 and Th17 cells, can have important implications for cancer development and progression. 4
Anti-PD-1 antibodies are a type of immune checkpoint inhibitors (ICIs) that have shown promising results in the treatment of several types of cancer, including CRC. 5 ICIs work by blocking certain proteins that suppress the immune system, allowing immune cells to recognize and attack cancer cells more effectively. 6 However, a significant proportion of patients do not respond to anti-PD-1 treatment due to the development of resistance mechanisms. 7 Therefore, there is a need for new approaches to reduce the immunotherapy resistance in CRC.
The gut microbiome consists of diverse microorganisms and has a significant responsibility in preserving the well-being of the host. 8 Imbalance of the gut microbiome has been associated with the emergence and advancement of multiple illnesses, such as CRC.9,10 Fecal microbiota transplantation (FMT) is a promising therapeutic approach that involves the transfer of fecal microbiota from a healthy donor to a recipient for restoring gut microbiota composition and function. 11 Recent studies have shown that the modulation of gut microbiome through antibiotics (ATBs) or FMT can affect response in immunotherapy-refractory melanoma patients. 12 Although several studies have investigated the potential of FMT as a therapeutic approach for CRC,13,14 the effects of FMT on the TME and its interaction with ICIs in CRC have not been fully elucidated.
Ultrasound (US) technology has lately acquired popularity as a noninvasive way to improve biotherapeutic delivery. Low-intensity focused US (LIFU) can temporarily increase intestinal permeability and facilitate microbial engraftment, while also modifying immune responses via mechanical and thermal actions inside the TME. The combination of US and microbiota-based treatment may potentially enhance immune activation and overcome therapeutic resistance.
The authors studied the synergistic manipulation of the TME using US-assisted FMT (US-FMT) to overcome anti-PD-1 resistance in a CRC xenograft mouse model. They investigated how combining microbiota restoration with US-mediated augmentation affects tumor development, immune cell infiltration, cytokine profiles, and overall immunotherapeutic effectiveness. These findings shed new light on the interactions between US, microbiota, and immunotherapy, highlighting US-FMT’s translational promise as a next-generation method for combating immunotherapy resistance in CRC.
Materials and Methods
Animal husbandry
The animal experiment was conducted in full compliance with guidelines and approved by the Animal Ethics Committee of Wujin Hospital of TCM (CZ20240037[L]). Female C57BL/6 mice (6 weeks, 19.0 ± 2.6 g) were provided by Cyagen (China), and 6-week-old female Rag1 knockout (Rag1 KO) mice were purchased from Shanghai Model Organisms (China). The mice were kept in a controlled animal facility with a 12-h light/dark cycle, specific pathogen-free surroundings, and unrestricted access to food. After a 1-week adaptation, mice were randomly divided into corresponding groups (n = 10).
Xenograft model
A xenograft mouse model was established using MC38 cells (C7399; Beyotime, China). Subcutaneous injection of MC38 cells was performed in the right flanks of all mice. The injection site was sterilized with alcohol, and a 100 µL suspension of MC38 cells at a concentration of 2 × 106 cells/mL in phosphate-buffered saline (PBS) was administered using a 26-gauge needle. Mice were monitored for general health status throughout the experiment. The growth of the tumors was assessed every 5 d, and treatment was commenced once the mean tumor volume had reached approximately 100 mm3. At last, mice were euthanized, and the tumors were collected for further analysis. Tumor volume = (length × width2)/2.
ATB treatment
To create gut microbiota depletion, mice were divided randomly into the ATB and control groups. To remove the indigenous gut microbiota, the ATB group was treated with oral broad-spectrum ATB, mixed with neomycin sulfate (200 mg/kg), ampicillin (200 mg/kg), metronidazole (200 mg/kg), and vancomycin (100 mg/k). This treatment was administered once a day for three consecutive days via oral gavage.
Patient information
Fecal samples were from five newly diagnosed CRC patients from Wujin Hospital of TCM hospital with informed consent. The exclusion criteria were as follows: history of ATB or probiotic use within 3 months before fecal sample collection; autoimmune or inflammatory diseases with immunomodulatory therapy; previous malignant tumors; history of FMT; and on a vegetarian diet. Asymptomatic individuals with no significant abnormalities on colonoscopy were recruited as healthy controls (HCs) and were subject to the same exclusion criteria as the eligible CRC patients. The study was conducted under the Ethics Committee of Wujin Hospital of TCM (CZ20240037[L]). Every donor stool sample was subjected to conventional safety screening, which included endotoxin quantification (LAL assay), enteric viruses, bacterial pathogens (Salmonella, Shigella, and Campylobacter), and Clostridioides difficile toxin. All samples met the safety standards before use.
FMT and US Application
Fecal samples from HCs and CRC patients were diluted in sterile PBS (0.1 g/mL), homogenized, and filtered through 70 µm strainers. The supernatant was separated and kept at −80°C until needed.
Mice were fasted for 6 h before FMT and given 0.2 mL of fecal suspension (equivalent to ∼108 colony-forming units) by oral gavage every other day for 2 weeks. To improve microbial engraftment and intestinal permeability, based on established norms for safe gastrointestinal sonoporation without causing tissue necrosis or heat injury, LIFU parameters (1 MHz, 1.0 W/cm2, 50% duty cycle, 2 min/session) were chosen. These settings remain below clinically acceptable mechanical index limitations (<1.9) and avoid stable cavitation damage. To check safety, intestinal sections obtained 24 h after the final US-FMT session were submitted to hematoxylin and eosin staining, indicating intact mucosal and submucosal architecture with no indications of epithelial shedding or inflammatory infiltration.
The fecal samples collected from both HCs and CRC patients using sterile collection tubes were diluted in sterile PBS and preserved at −80°C until further analysis. Equal FMT and PBS were administered via transgastric administration using a 22-gauge feeding needle. Mice were fasted for 6 h before FMT and were then orally gavaged 0.2 mL/mouse every other day for a duration of 2 weeks. Then the mice were allowed to recover for 3 d before tumor cell injection.
Anti-PD-1 treatment
To investigate the effects of anti-PD-1 on the TME in the xenograft mouse model, tumor-bearing mice were treated with either isotype control immunoglobulin G (IgG) or anti-PD-1 monoclonal antibody (mAb). Specifically, after the tumors reached an average volume of 100 mm3, the mice were randomly divided into two groups (n = 10) and injected intraperitoneally with either 100 µg of isotype control IgG (31154; Thermo Fisher, USA) or 100 µg of anti-PD-1 mAb (MA5-15780; Thermo Fisher) twice a week for 4 weeks.
Flow cytometry
Cell suspensions were obtained from tumors as previously described. 15 In brief, cells were treated with phorbol myristate acetate (HY-18739, 50 ng/mL; MedChem, China), ionomycin (HY-13434, 500 ng/mL; MedChem), and brefeldin A (HY-16592, 10 μg/mL; MedChem) for 4 h at 37°C, 5% CO2. Fluorescence-labeled ATBs specific for CD3 (100236), IFNγ (505810), IL-17a (506904), CD4 (100412), and CD8 (100722) from BioLegend (USA) were used for staining. Intracellular Fixation and Permeabilization Buffer Set (88-8824-00; BioLegend) was used for staining. Data were processed using the BD LSRFortessa flow cytometer (BD Biosciences, USA) and FlowJo software.
Western blot
Tumor tissues were lysed using RIPA buffer (Beyotime). Protein concentrations were quantified by the BCA Protein Assay Kit (ab102536; Abcam, UK), transferred onto PVDF membranes (Millipore, USA), and blocked with 5% nonfat milk. Primary antibodies against VEGF, MMP-2, MMP-9, IL-1β, and β-actin (loading control) were added for incubation at 4°C overnight. The mixture was coincubated with secondary antibodies conjugated with HRP for 1 h at room temperature. ECL detection system (Bio-Rad, USA) was used for visualization. The primary antibodies used in this study were purchased from Abcam, including anti-VEGF (ab32152, 1/1000), anti-MMP-2 (ab97779, 1/1000), anti-MMP-9 (ab76003, 1/1000), anti-IL-1β (ab9722, 1/1000), and anti-β-actin (ab8227, 1/1000).
Measurement of α-diversity
The diversity analysis was processed in accordance with previous research. 16 Fecal samples were collected from the HC and CRC groups with DNA extracted using the QIAamp DNA Stool Mini Kit (51504; Qiagen, Germany). The α-diversity was calculated based on the OTU tables generated from the 16S ribosomal RNA (rRNA) gene sequencing data using three commonly used indices: OTU, Shannon, and Simpson. OTU measures species richness, whereas Shannon takes into account both richness and evenness of the microbial community. Simpson measures the dominance of certain taxa in the community. The statistical comparison of α-diversity between the HC and CRC groups was performed using R software. Higher α-diversity values indicate a more diverse microbial community, whereas lower values indicate a less diverse community.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 8 and R 3.5.1. Data were expressed as mean ± SEM, with unpaired two-tailed Student’s t-test for analysis. A p-value of <0.05 was considered statistically significant.
Results
US-assisted microbiome modulation decreases tumor burden in CRC xenograft model
The effect of gut microbiome on subcutaneous tumor burden in a CRC xenograft model was investigated via depletion. To achieve this, common tumor-bearing mice received oral broad-spectrum ATBs to knock out the intestinal flora. The results showed that ATB treatment decreased significantly both the tumor growth curve and weight (Fig. 1A, B). Next, US-FMT was investigated as a noninvasive method of restoring microbial balance and strengthening immunological response.
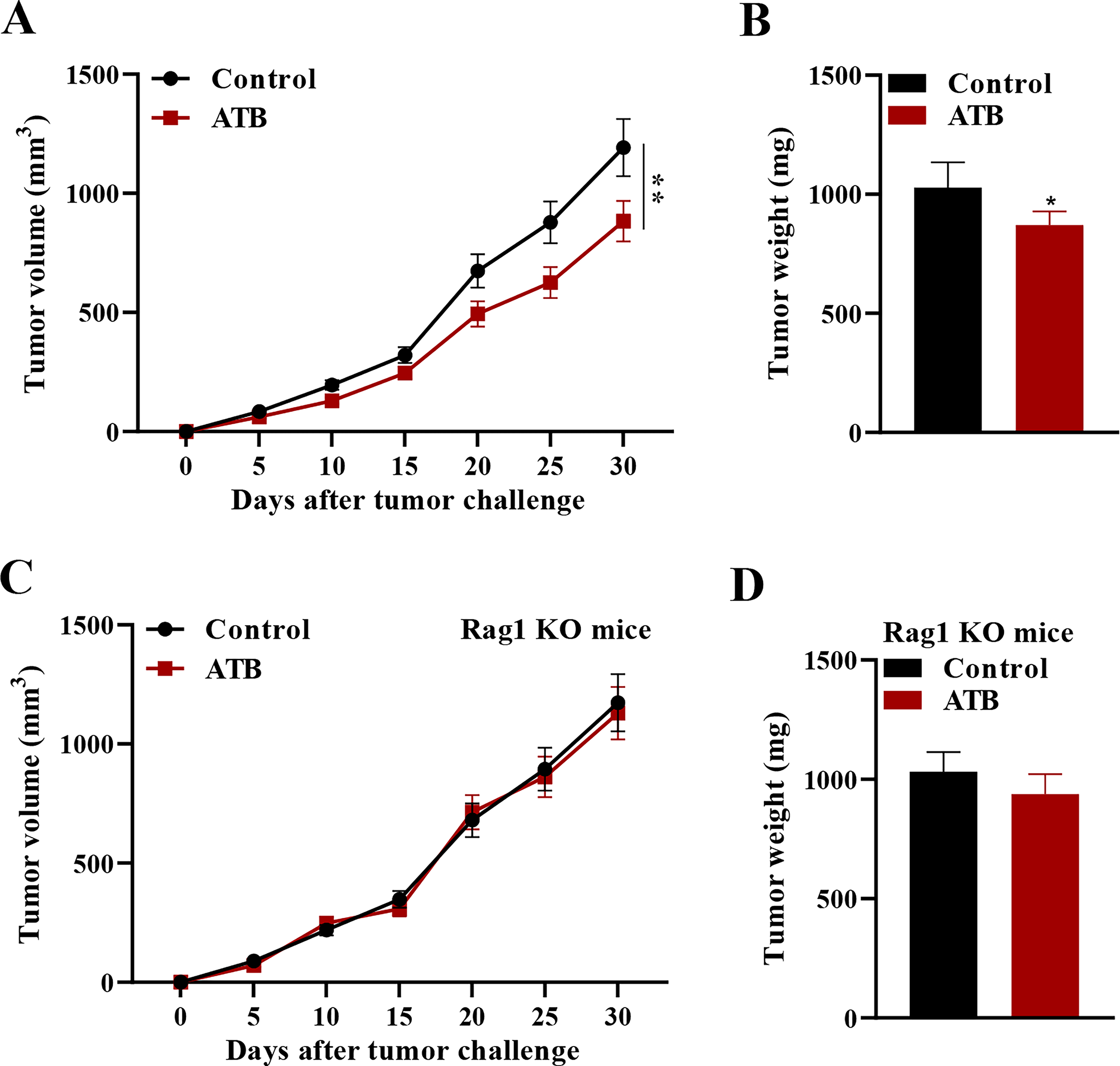
To further examine the role of adaptive immunity in this process, the authors used Rag 1 knockout mice (lacking functional T and B lymphocytes) as a tumor-bearing model. In contrast to the common tumor-bearing mice, knockout of gut microbiome in Rag 1 knockout mice had no significant effect on the tumor growth curve or weight (Fig. 1C, D). These results indicated that the tumor-reducing effect of ATBs requires the participation of adaptive immunity.
US-assisted microbiome modulation reshapes the TME
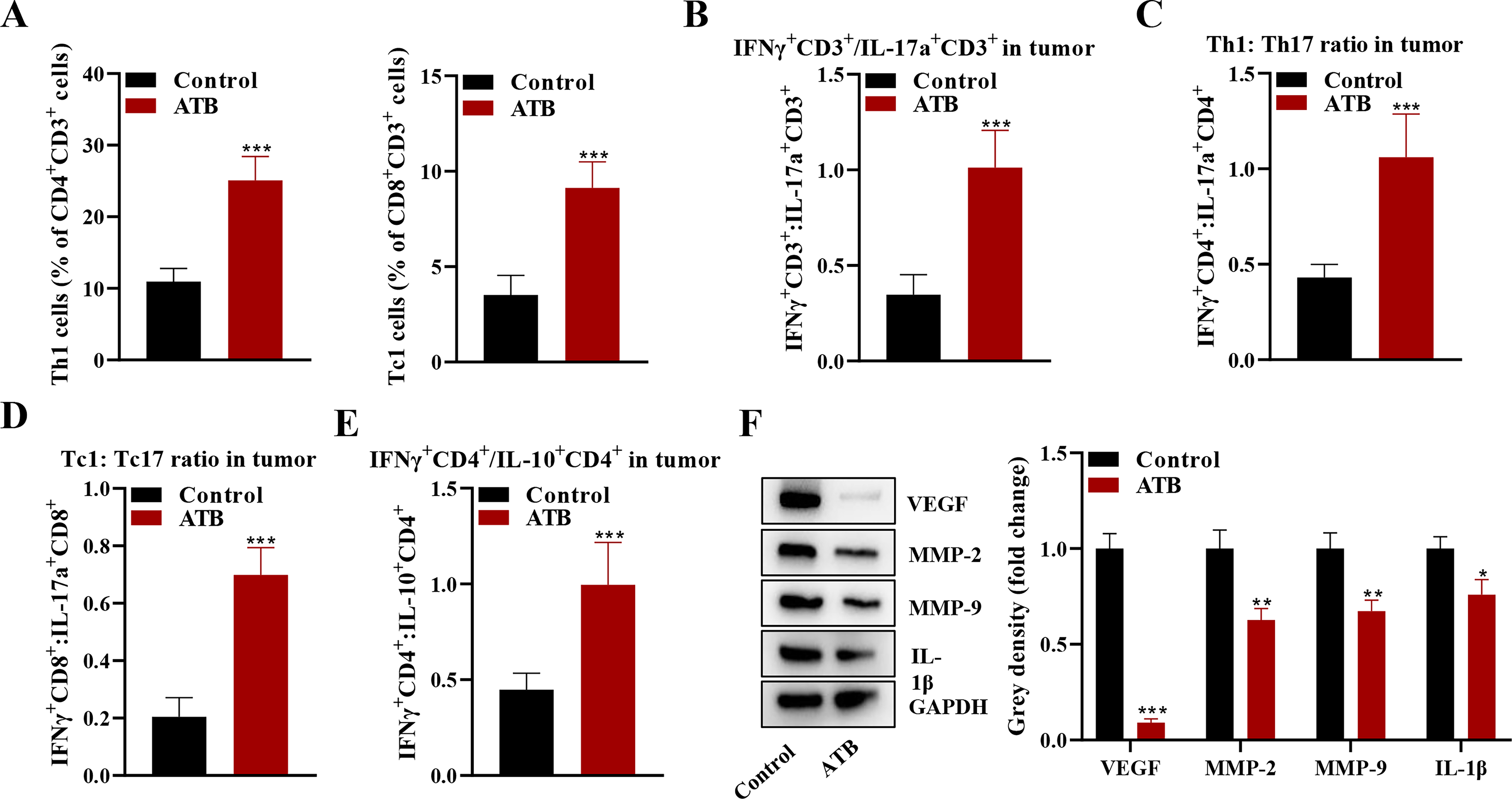
Flow cytometry showed that gut microbiome depletion in the ATB group increased the ratio of Th1 and Tc1 cells in the tumor (Fig. 2A). It also significantly increased the ratio of IFNγ+CD3+/IL-17a+CD3+, Th1/Th17, Tc1/Tc17, and IFNγ+CD4+/IL-10+CD4+ (Fig. 2B–E). In addition to modulating immune cell populations, gut microbiome depletion also affected the expression of several key factors involved in the TME. 17 Western blot analysis showed that the expression of VEGF, MMP-2, MMP-9, and IL-1β was suppressed in the ATB group compared with the control group (Fig. 2F).
US-FMT restores microbial diversity and reduces tumor growth
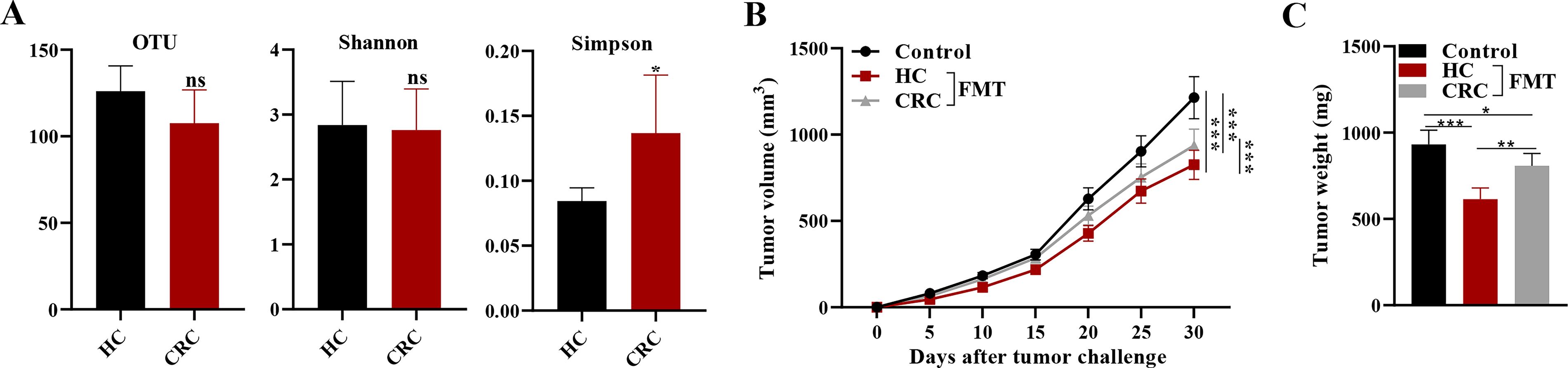
Fecal samples were collected from CRC patients and normal controls to analyze the α-diversity of intestinal microbiota in both groups. The higher OTU in the HC group indicated that there was a greater richness of species in the gut microbiota of healthy individuals compared with CRC patients. The Shannon index being approximately equal between the two groups suggested that the evenness of species was similar between the HC and CRC groups. The lower Simpson index in the HC group suggested that there was a lower dominance of any one species in the gut microbiota of healthy individuals compared with CRC patients (Fig. 3A). Then, fecal suspension from either normal controls or CRC patients was administered intragastrically into mice, with tumor growth curves and weights measured every 5 d within 30 d in total. Mice getting FMT from HCs (HC-FMT) demonstrated a substantial decrease in tumor volume and weight compared with those receiving FMT from CRC patients or PBS vehicle (Fig. 3B, C). Importantly, US-FMT enhanced this tumor-suppressive impact, indicating a synergistic relationship between mechanical ultrasonic stimulation and microbial repair. Quantitative PCR measurement of total 16S rRNA copies demonstrated a 2.8-fold increase in microbial load in the US-FMT group versus FMT alone (p < 0.01), demonstrating improved microbial engraftment produced by ultrasound. These findings indicate that US-enhanced microbial colonization boosts host antitumor immunity and inhibits CRC tumor development.
US-FMT reprograms the TME toward an immunostimulatory phenotype
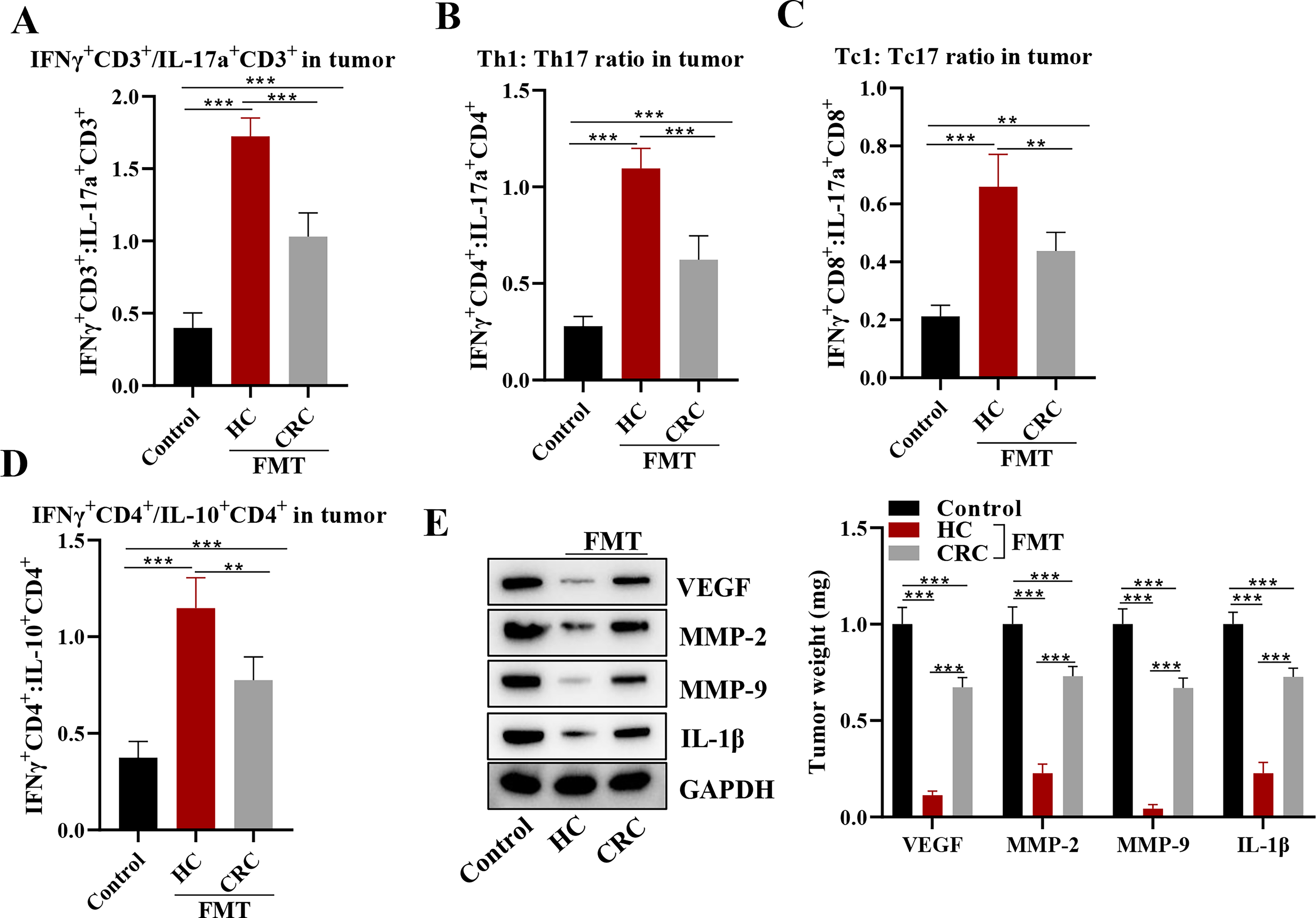
The effects of FMT on the TME were explored in the CRC xenograft mouse model. Immune cell populations were analyzed within the tumors by measuring the ratios of IFNγ+CD3+/IL-17a+CD3+, Th1/Th17, Tc1/Tc17, and IFNγ+CD4+/IL-10+CD4+. Flow cytometry showed that all of these ratios were significantly higher in the HC and CRC groups compared to the control group, with the highest values observed in the HC group (Fig. 4A–D). Expressions of several key proteins involved in tumor growth and progression were also assessed, including VEGF, MMP-2, MMP-9, and IL-1β. Western blot analysis revealed that the expression levels of all these proteins were notably decreased in the HC and CRC groups compared with the control group, with the lowest values observed in the HC group (Fig. 4E). US-FMT was related with elevated IFN-γ–STAT1 signaling, increased MHC-I expression, and transcriptional enrichment of antigen presentation pathways, suggesting a molecular axis by which US-FMT potentiates anti-PD-1 responses. US-FMT caused IL-12 upregulation in tumor-infiltrating dendritic cells, supporting a feed-forward IFN-γ–driven Th1/Tc1 polarization. Taken together, these findings suggested that FMT from HCs may have a beneficial effect in modulating the TME and inhibiting tumor growth in this mouse model.
US-FMT enhances anti-PD-1 immunotherapy efficacy
It was found that FMT from HCs significantly reduced tumor volume and weight compared with the PBS group, and this effect was further enhanced by anti-PD-1 mAb treatment (Fig. 5A, B). Immune cell populations within the tumors were evaluated via ratios of IFNγ+CD3+/IL-17a+CD3+, Th1/Th17, Tc1/Tc17, and IFNγ+CD4+/IL-10+CD4+. Results showed that all of these ratios were significantly higher in the HC-FMT group compared with the PBS group, and further increased by anti-PD-1 mAb treatment (Fig. 5C–F). In addition, Western blot analysis revealed that the expression levels of VEGF, MMP-2, MMP-9, and IL-1β were significantly lower in the HC-FMT group compared with the PBS group, and further reduced by anti-PD-1 mAb treatment (Fig. 5G). Taken together, these results indicate that FMT from HCs may improve the curative effect of anti-PD-1 treatment by modulating the TME and potentiating antitumor immune responses in this mouse model.
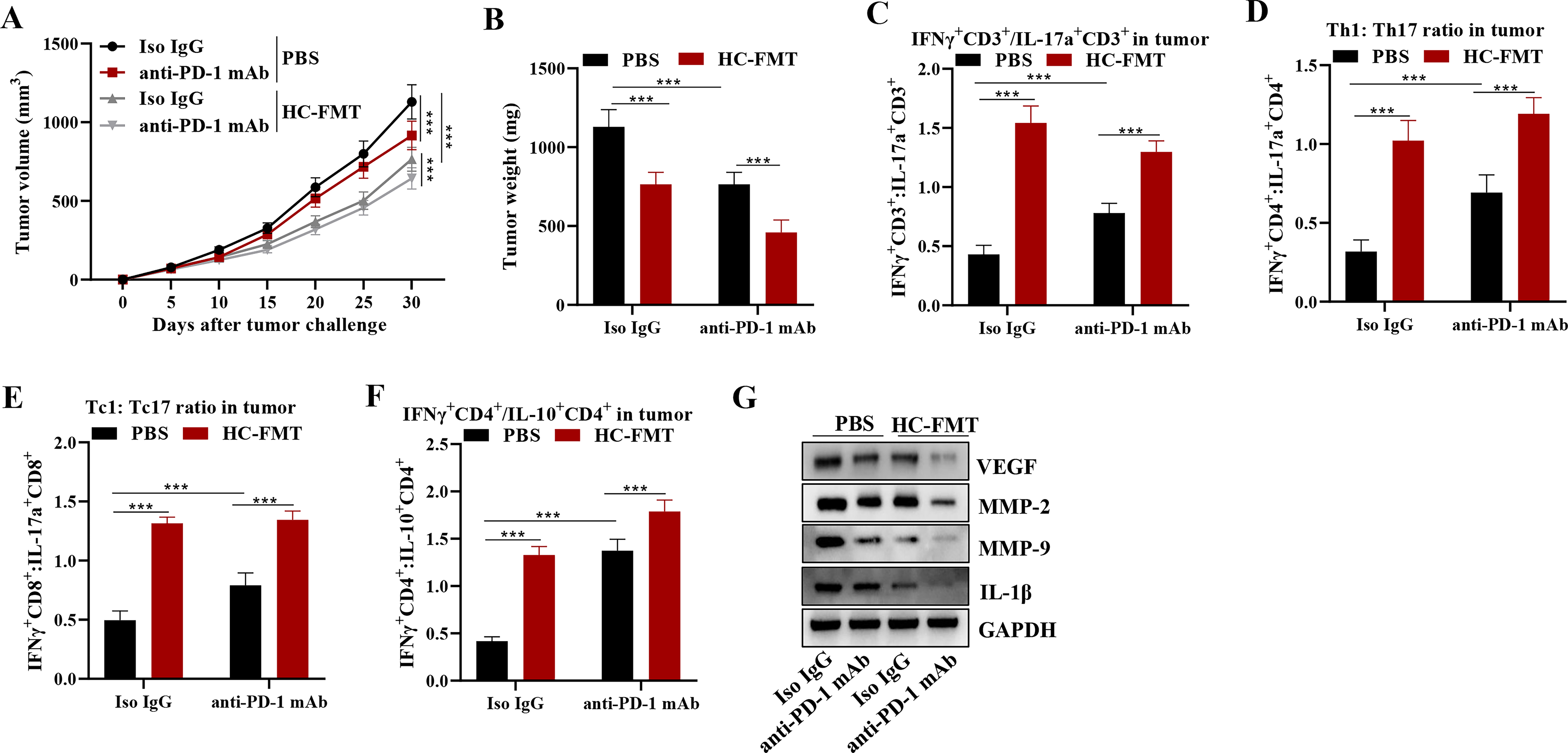
Taken together, these data suggest that US-FMT improves anti-PD-1 immunotherapy effectiveness by altering the gut microbiota, enhancing adaptive immune responses, and modifying the TME toward a more immunogenic and less angiogenic state.
Discussion
Gut microbiota affects immune responses through several pathways, including regulating the maturation and differentiation of immune cells, influencing cytokine production, and modulating the activation of toll-like receptors,18–20 suggesting a potential link between gut archaea dysbiosis and CRC development. These findings are in line with recent research suggesting that gut microbiota modulation may represent a novel strategy for the prevention and treatment of CRC.21–23 However, how this mechanism relates to the microenvironment and immune responses remains unknown. These interactions influence both local and systemic immune responses, establishing a molecular relationship between microbial dysbiosis and colorectal carcinogenesis. In this work, the authors showed that US-FMT alters the TME in a CRC xenograft model, overcoming anti-PD-1 resistance.
In these results, US-FMT led to a significant increase in the ratio of Th1/Th17 and Tc1/Tc17, along with the ratios of IFNγ+CD3+/IL-17a+CD3+ and IFNγ+CD4+/IL-10+CD4+ cells, indicating a shift toward a more pro-inflammatory Th1 and Tc1 response. 24 These findings suggested that US-FMT may be a potential therapy for CRC by modulating the immune response.
Furthermore, the authors found that KMT resulted in a notably decreased expression of VEGF, MMP-2, MMP-9, and IL-1β. VEGF is a key mediator of angiogenesis, which is the process by which new blood vessels are formed to supply nutrients and oxygen to the growing tumor. 25 MMP-2 and MMP-9 are enzymes that can degrade extracellular matrix proteins, facilitating the invasion and metastasis of cancer cells. 26 IL-1β is a pro-inflammatory cytokine that can promote tumor growth and metastasis and assist in attracting immune cells to the location of the tumor. 27 This finding suggested that US-FMT may also have an antitumor effect by reducing tumor-promoting factors.
Based on the results of this study, it is also worth considering the potential role of anti-PD-1 therapy in combination with US-FMT. This synergism was previously analyzed via metagenomic and metabolomic analysis 28 and validated in patients with metastatic melanoma. 29 In this work, the effects of US-FMT on tumor volume and weight, ratios of IFNγ+CD3+/IL-17a+CD3+, Th1/Th17, Tc1/Tc17, and IFNγ+CD4+/IL-10+CD4+, and expression levels of VEGF, MMP-2, MMP-9, and IL-1β were all further enhanced by anti-PD-1 mAb treatment. In contrast to the aforementioned studies, this work focused on the index changes related to the TME and antitumor immune responses, revealing the synergistic role of US-FMT in combination therapy with anti-PD-1. However, in 2021, it was pointed out that dominant commensal microbes present in the recipient’s feces after fecal transplant did not correlate with their response to anti-PD-1 immunotherapy, 30 which suggested that this synergism did not function via changing microbe diversity and needs to be further elucidated.
This study has significant drawbacks. First, a cell line-derived xenograft model may not fully represent the heterogeneity of real CRC and its immunological landscape. Second, while ultrasonic settings were tuned for microbial delivery and tissue safety, more dose–response and mechanistic investigations are required to fine-tune clinical use. PICRUSt-based KEGG inference suggested enrichment of SCFA-producing pathways in the US-FMT group, whereas comprehensive functional metagenomics will be required in future investigation. As highlighted by Di Maggio et al., 31 multiomic integration may be essential to predicting microbiome–immunotherapy interactions. The authors have integrated this perspective into their future directions. Last, possible hazards of recurrent FMT and US exposure, such as microbial translocation or tissue inflammation, must be carefully considered in future preclinical and clinical studies.
Overall, these findings offer persuasive evidence that US-FMT is a novel, noninvasive strategy for changing the TME and overcoming immunotherapy resistance in CRC patients. By combining ultrasonic bioengineering and microbiome manipulation, this synergistic approach opens up new avenues for boosting the effectiveness of immune checkpoint blockade.
Conclusion
This study emphasizes the potential of US-FMT as a synergistic therapeutic approach to enhance immunotherapy efficacy by reprogramming the TME and overcoming anti-PD-1 resistance in CRC.
Authors’ Contributions
X.X.: Conceptualization, data curation, investigation, methodology, project administration, and writing—original draft. X.L.: Formal analysis, funding acquisition, software, supervision, visualization, and writing—review and editing.
Footnotes
Funding Information
No funding was received for this article.
Data Availability
The datasets generated and analyzed during the current study are available from the corresponding author upon reasonable request.
Disclosure Statement
The authors have no conflicts of interest to disclose.