
Abstract
Focused ultrasound (FUS) is a noninvasive neuromodulation technique capable of modulating deep-brain circuits with millimeter precision. When combined with functional magnetic resonance imaging (fMRI), it enables causal decoding of brain-state dynamics in vivo. In this study, the authors applied deep learning to fMRI data acquired from nonhuman primates during thermal pain stimulation with and without transcranial FUS targeting the ventral posterolateral (VPL) nucleus of the thalamus, a key somatosensory relay. A 3D convolutional neural network was trained to classify three experimental conditions: Heat (pain only), Heat + FUS, and FUS-only. The model achieved an average accuracy of 92.2% (binary) and 91.2% (multiclass), with receiver operating characteristic-area under the curve values exceeding 0.90. Saliency mapping revealed significant cortical suppression in the secondary somatosensory cortex (S2, p = 0.036) and insula (p = 0.024), accompanied by preserved thalamic relevance in the VPL. In FUS-only trials, saliency shifted toward dorsolateral prefrontal regions, suggesting cognitive engagement even in the absence of nociceptive input. These findings demonstrate that FUS attenuates thalamocortical pain transmission while maintaining subcortical relay integrity. This work highlights the translational potential of FUS as a targeted, reversible neuromodulation strategy and underscores the value of artificial intelligence-driven neuroimaging in decoding dynamic pain states.
Keywords
Introduction
Pain is a complex and multifactorial phenomenon, involving sensory, emotional, and cognitive dimensions mediated through distributed brain networks. 1 Although acute pain plays a vital protective role, chronic pain conditions are frequently disabling and poorly managed with existing treatments. Pharmacological strategies often lack specificity, and their long-term use is associated with risks such as tolerance, systemic side-effects, and dependency. 2 This has led to increasing interest in neuromodulatory approaches that can selectively target dysfunctional circuits while minimizing invasiveness and side-effects. 3 Recent advances in medical imaging and Artificial Intelligence (AI) have demonstrated the potential of multimodal deep learning frameworks to capture complex biological interactions noninvasively. For instance, a new study proposed a dual-parallel convolutional neural network (CNN) for high-intensity ultrasound and biomarker fusion in breast cancer, achieving near-perfect diagnostic accuracy and demonstrating the translational value of AI-based image decoding in therapeutic monitoring. 4
MR-guided low-intensity focused ultrasound (MRgFUS) has emerged as a promising technique 5 capable of targeting deep-brain structures, including thalamic nuclei, with submillimeter precision. 6 Unlike Deep-Brain Stimulation (DBS), MRgFUS does not require implanted hardware and allows transient, reversible modulation. Recent studies integrating multimodal imaging modalities, including ultrasound, photoacoustic imaging, and MRI, have demonstrated the ability to capture hyperacute physiological changes after brain injury, offering insight into vascular and tissue responses that are not visible via single-modality MRI alone. 7 Preclinical and clinical studies have shown that FUS can alter neuronal excitability and modify brain connectivity patterns 8 ; however, its precise effect on nociceptive signaling, particularly during ongoing pain stimuli, remains poorly characterized. Recent work demonstrated the safety and efficacy of transcranial MRgFUS in modulating deep structures such as the amygdala and nucleus accumbent in psychiatric disorders, further validating its clinical potential.9,10
The ventral posterolateral (VPL) nucleus of the thalamus is a major conduit for somatosensory information from the spinal cord to the cortex. Similar thalamocortical mechanisms have also been studied in the context of sleep spindle detection, where machine-learning approaches have been applied to quantify oscillatory biomarkers linked to cognitive processing. 11 As a key node in the pain matrix, the VPL relays both discriminative and affective aspects of nociceptive input. Modulation of this structure is particularly promising for interrupting pain signals at the relay stage. Yet, it remains unclear how transient VPL modulation influences downstream cortical processing during active pain states.
To investigate this, the authors analyzed a publicly available Nonhuman Primate (NHP) dataset comprising BOLD Functional Magnetic Resonance Imaging (fMRI) recordings acquired during thermal pain stimulation, with and without concurrent MRgFUS directed at the VPL. NHPs are ideal for translational pain studies due to their anatomically conserved somatosensory systems and similar thalamocortical architecture to humans. 12 Beyond its neuromodulatory applications, FUS has been shown to enhance molecular delivery and radiopharmaceutical uptake in brain tumor cells through reversible sonoporation, highlighting its expanding translational role in neuro-oncology. 13
Traditional fMRI analysis techniques, including the General Linear Model (GLM), Independent Component Analysis (ICA), and Region-of-Interest (ROI)-based statistics, are foundational tools for mapping task-related activity. 14 However, these methods are often constrained by strong a priori assumptions regarding spatial localization and signal linearity, potentially overlooking distributed or nonlinear patterns of neural modulation. This limitation is especially evident in neuromodulation studies, where perturbations from deep structures such as the thalamus may propagate across complex, large-scale networks. Recent resting-state fMRI studies in glioma patients have shown that functional metrics such as global signal cross-correlation and ALFF can sensitively capture tumor-related alterations in hemodynamics, supporting the need for more advanced decoding frameworks. 15
Although CNNs are known to capture nonlinear spatial hierarchies, classical machine-learning algorithms such as GLM, logistic regression, and Support Vector Machines (SVMs) have historically been applied to fMRI decoding tasks. 16 A direct comparison of identical data pipelines is necessary to quantify the relative gain offered by deep learning in neuromodulation settings. 17
Recent advances in machine learning have introduced more flexible alternatives for pattern recognition in neuroimaging. Approaches such as SVMs and logistic regression offer modest gains in sensitivity but typically require handcrafted features or dimensionality reduction, which can obscure spatial relationships in the data. In contrast, CNNs learn spatially localized and hierarchical features directly from volumetric input, offering high sensitivity to subtle condition-specific neural signatures. 18
In this study, the authors used a 3D CNN architecture to classify stimulus conditions based on whole-brain BOLD activation patterns. This approach incorporated guided backpropagation and Gradient-weighted Class Activation Mapping (Grad-CAM) saliency mapping to identify brain regions contributing most to the model’s classification performance. The authors hypothesized that FUS targeting the VPL would lead to measurable attenuation of pain-related cortical activity while preserving or altering thalamic involvement, patterns that could be robustly decoded using deep learning.
To test this hypothesis, the authors utilized an open-source, BIDS-formatted fMRI dataset, which includes two experimental designs: a two-condition paradigm alternating between Heat and Heat + FUS, and a three-condition design introducing FUS-only trials. By combining neuroimaging and AI, this study provides new insight into how ultrasound modulates thalamocortical processing and highlights the use of CNNs for decoding complex brain-state dynamics with high spatial fidelity. The clinical significance of FUS lies in its noninvasive, reversible capacity to modulate deep-brain circuits, offering a potential alternative to implanted neurostimulation systems for chronic pain and neuropsychiatric disorders.
Methods
Dataset and experimental paradigms
This study utilized an open-access dataset, consisting of BOLD fMRI scans acquired from five NHPs subjected to thermal pain stimulation and MRgFUS neuromodulation. The objective of the dataset was to evaluate how FUS applied to the VPL nucleus of the thalamus influences brain-wide activation during painful sensory input. 19
Two experimental paradigms were used. For subjects Sub01 to Sub03, a two-condition block design alternated between painful heat stimuli and heat paired with VPL-targeted ultrasound (Heat + FUS). Subjects Sub04 and Sub05 were exposed to an extended three-condition protocol, which also included an FUS-only condition, allowing assessment of ultrasound effects in the absence of nociceptive input.
Onset timings and stimulus presentation sequences were documented for each run. Heat and Heat + FUS conditions alternated in fixed blocks (16 s on, 30 s rest) in the two-condition paradigm. The three-condition paradigm used randomized onset times to minimize temporal predictability. The onset timings of Heat, Heat + FUS, and FUS-only conditions for each subject are provided in Table 1. The two- and three-condition paradigms applied in this study are schematically shown in Figure 1.
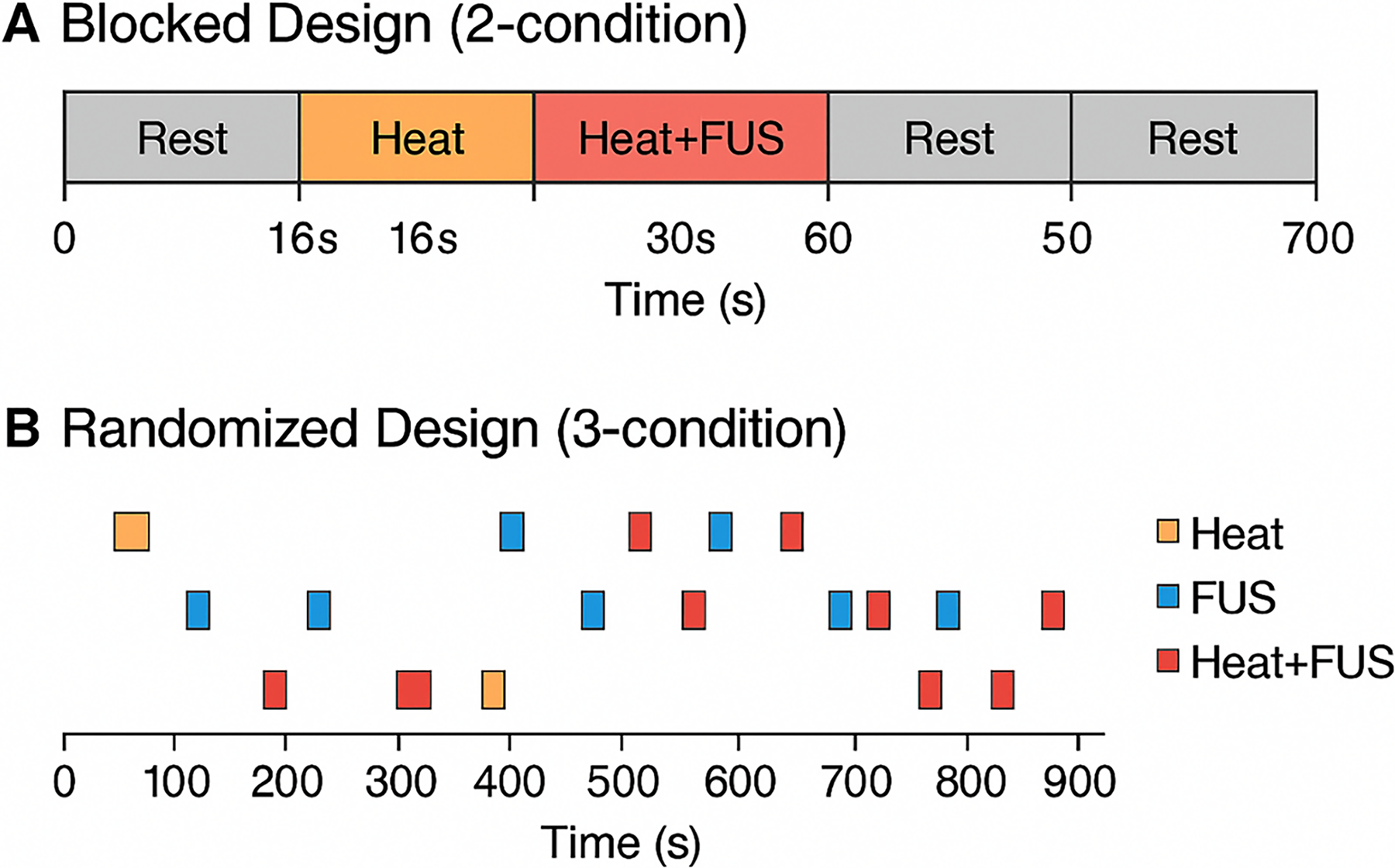
Experimental Paradigm Designs for NHPs: Two Condition & Three Condition Design Paradigms:
This Table Summarizes the Onset Times (in Seconds) for Each Experimental Stimulus, Heat, Heat + FUS, and FUS-Only, Across All Subjects and Paradigms

Subjects 01–03 participated in a two-condition design (Heat and Heat + FUS), whereas Subjects 04–05 participated in a three-condition design (Heat, Heat + FUS, and FUS-only). The table provides a detailed reference for the timing and sequence of stimuli used in the fMRI acquisition protocol, clarifying the experimental design for subsequent neural decoding analyses.
fMRI, functional magnetic resonance imaging; FUS, focused ultrasound.
Total scan durations ranged from ∼674 to ∼996 s depending on paradigm and subject.
MRI acquisition and ultrasound delivery
Imaging was conducted on a 3T MRI scanner equipped for ultrasound guidance. BOLD images were acquired using a gradient-echo echo-planar imaging sequence with the following parameters: repetition time: 2000 ms, echo time: 30 ms, field of view: 220 mm, voxel dimensions: 3.4 × 3.4 × 3.5 mm³, and number of slices: 36 (acquired interleaved, ascending). 20 Representative midslice BOLD images demonstrating sufficient anatomical detail are shown in Figure 2.
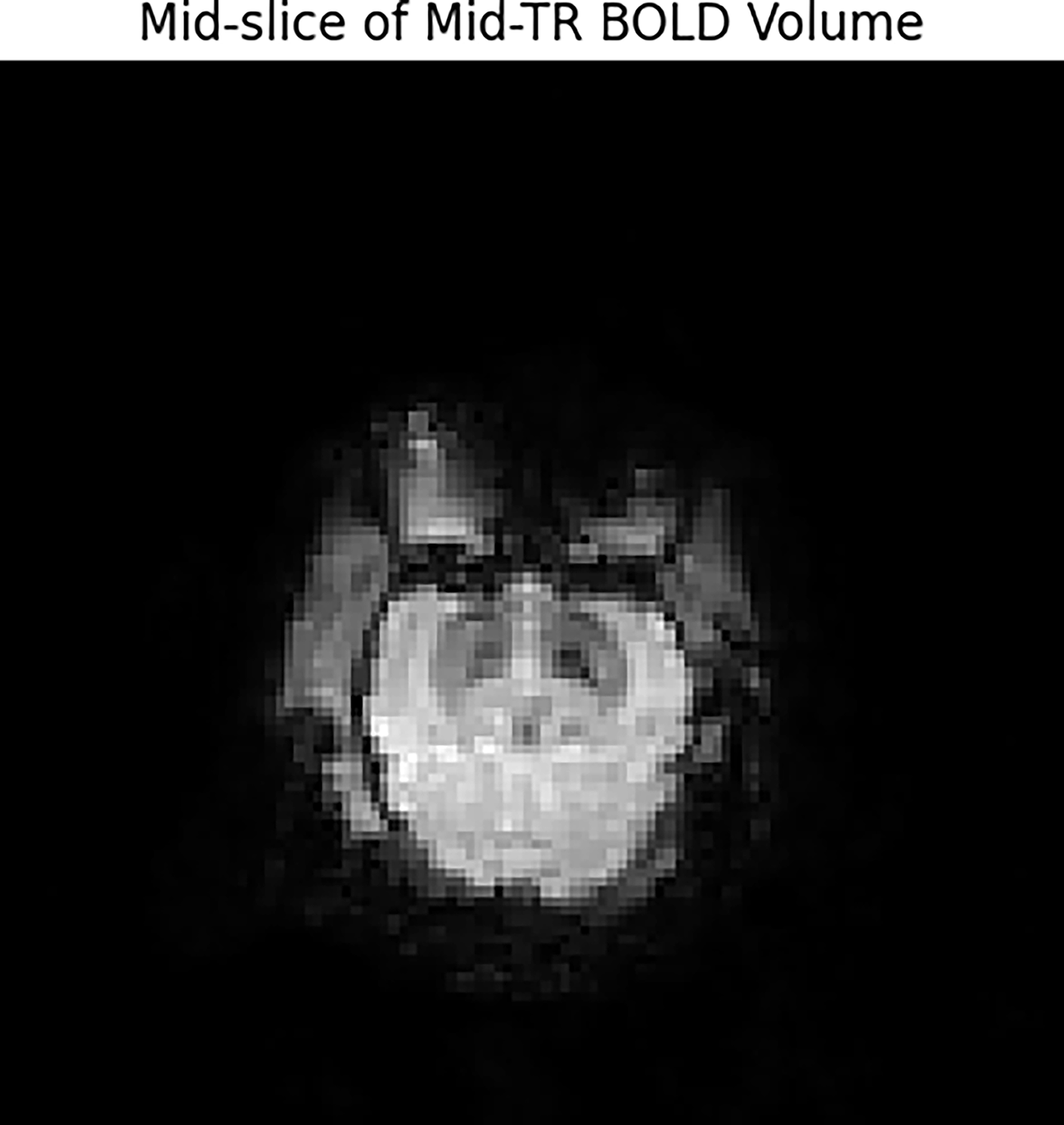
Representative midslice from BOLD functional magnetic resonance imaging (fMRI) illustrating preserved anatomical detail and sufficient signal-to-noise ratio for whole-brain Convolutional Neural Network (CNN) analysis.
High-resolution T1-weighted structural images were also acquired for anatomical alignment.
Ultrasound was delivered using an MR-compatible system targeting the VPL. An example of high-resolution T1-weighted anatomical alignment with brain mask application is provided in Figure 3. The stimulation protocol consisted of 500 ms bursts delivered every 2 s during the Heat + FUS and FUS-only conditions. 21 Real-time monitoring ensured accurate targeting and compliance with safety constraints.
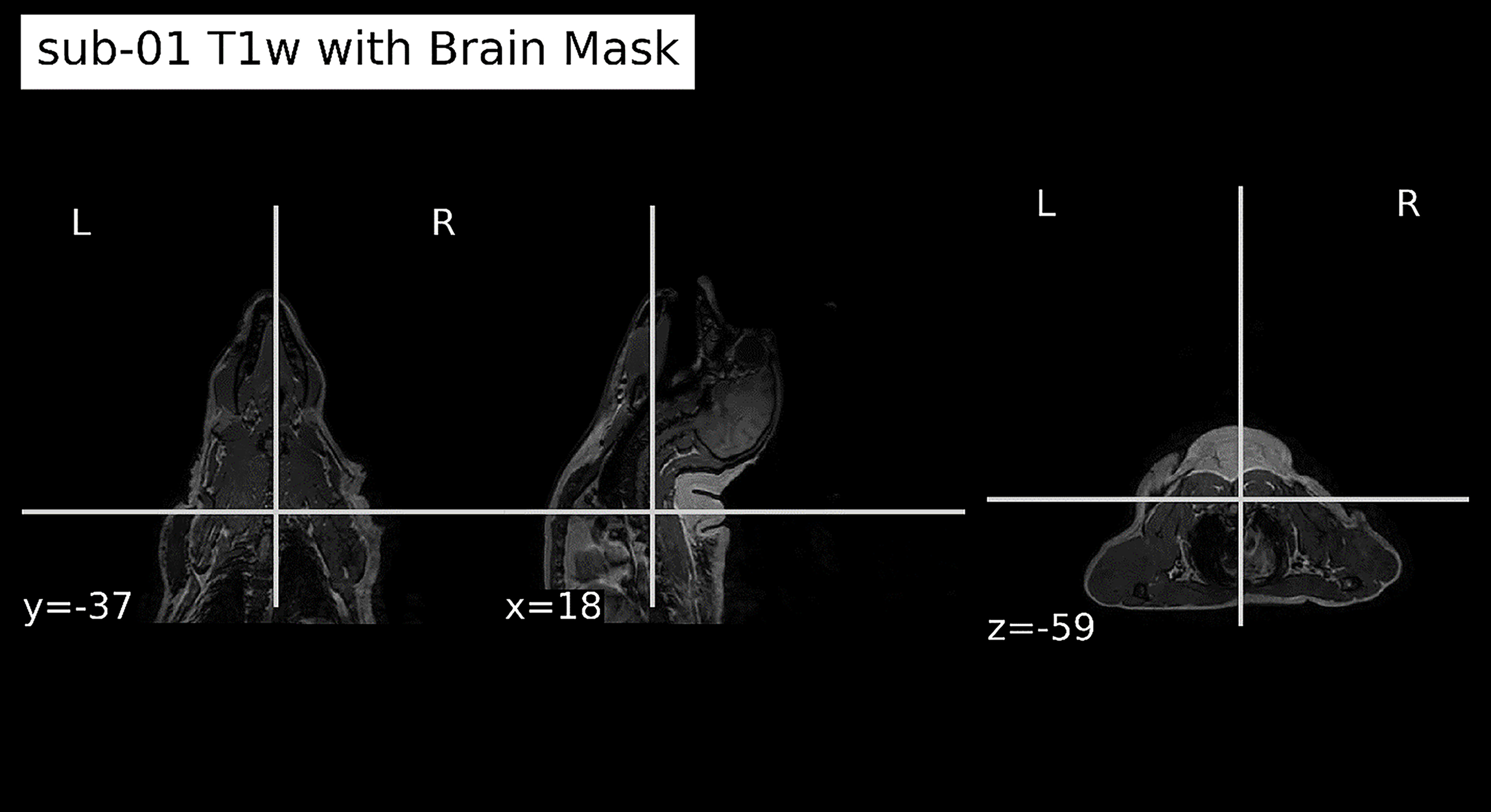
High-resolution T1-weighted anatomical magnetic resonance imaging (MRI) after registration to functional space and application of brain mask. The brain mask delineates cortical and subcortical structures, enabling accurate spatial normalization and subsequent group-level analysis.
Recent reviews have emphasized the therapeutic potential of low-intensity FUS in pain and neuropsychiatric disorders by modulating both cortical and subcortical circuits without structural disruption. 22
Preprocessing pipeline
To prepare the fMRI data for analysis, a standardized preprocessing pipeline was applied, adhering to BIDS recommendations. Preprocessing was carried out using tools from FSL and ANTs:
Motion correction: Conducted with FSL’s MCFLIRT, aligning all volumes to the median image within each run.
23
Following motion correction, the six motion parameters (three translations and three rotations) were included as nuisance regressors to minimize residual motion influence on voxel-wise signal variance before CNN feature extraction. Slice timing correction: Performed using FSL’s slicetimer to compensate for interleaved acquisition.
24
Anatomical coregistration: T1-weighted anatomical images were registered to the functional space.
25
Normalization: Each subject’s anatomical image was warped to the Nonhuman Primate Template (NMT v2.0) using ANTs’ nonlinear registration.
26
The same warp was applied to the functional volumes for group-level comparability. No spatial smoothing was applied, preserving fine-scale activation patterns critical for CNN-based classification.
27
The dataset passed BIDS validation and underwent quality control to ensure the integrity of spatial and temporal dimensions. 28 Automated pipelines for magnetic resonance spectroscopy and neuroimaging quality control have recently demonstrated improved detection of subtle artifacts compared with manual assessment, highlighting the importance of standardized preprocessing in multimodal datasets. 29 The complete workflow for preprocessing, CNN training, and saliency-based interpretation is illustrated in Figure 4.
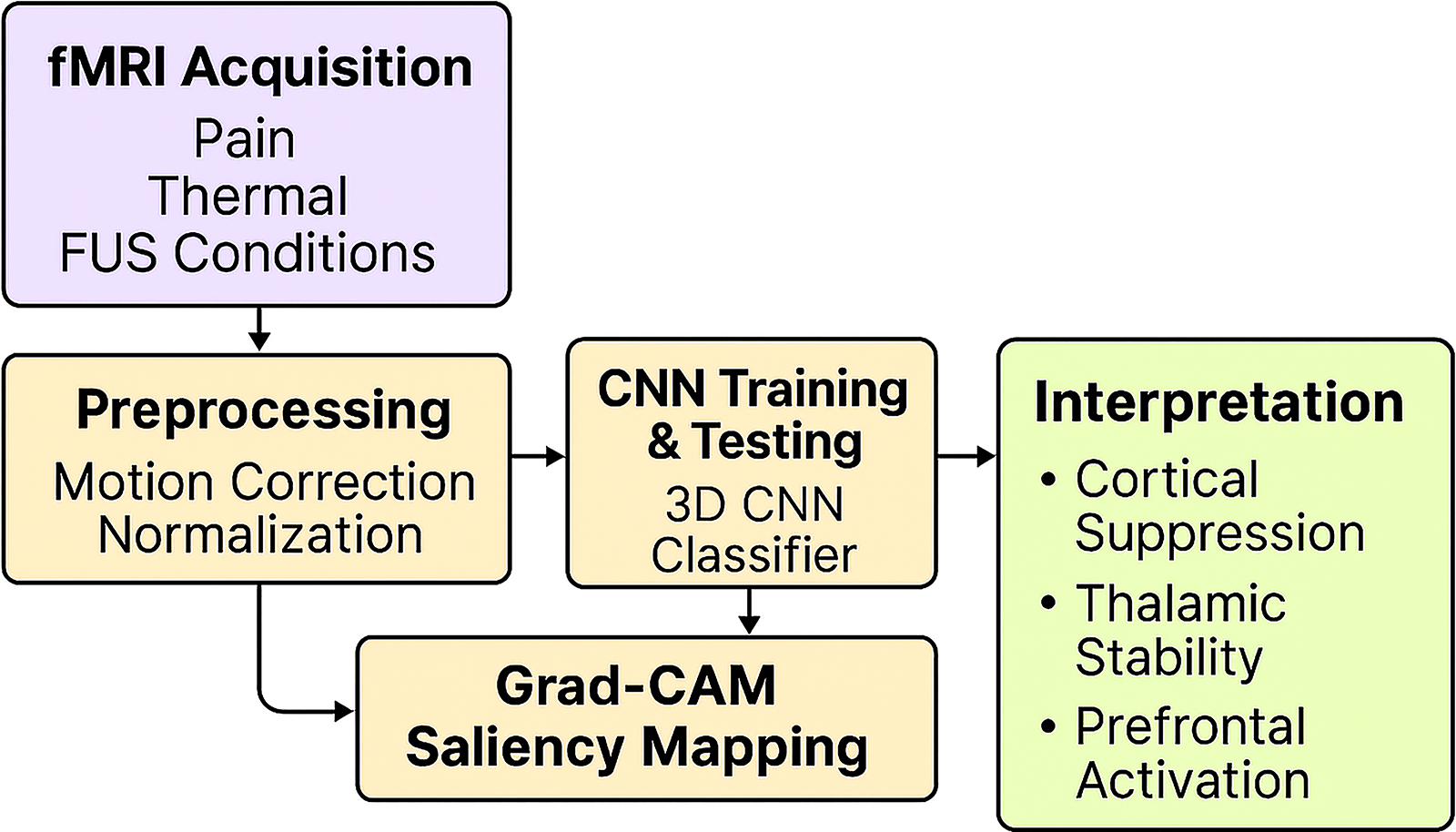
Overview of the functional magnetic resonance imaging (fMRI) analysis pipeline, illustrating data flow from preprocessing to convolutional neural network (CNN) training and Gradient-weighted Class Activation Mapping (Grad-CAM) saliency mapping. Arrows show the sequential process from fMRI acquisition to interpretation of neural signatures reflecting cortical suppression and thalamic stability.
Deep learning architecture
To classify fMRI volumes by experimental condition, the authors implemented a 3D CNN using the PyTorch framework. The input data consisted of individual BOLD volumes extracted from stimulus blocks across all three conditions (Heat, Heat + FUS, and FUS-only). Each input volume was normalized and reshaped to a consistent spatial resolution for training.
The CNN architecture was composed of three convolutional blocks, each consisting of a 3 × 3 × 3 convolutional layer, batch normalization, a rectified linear unit activation, and 2 × 2 × 2 max pooling. Two fully connected (dense) layers with 512 and 128 neurons, respectively, each followed by dropout (rate = 0.5) to minimize overfitting. 30 Output layer: A softmax activation function produced class probabilities for either binary (two-class) or multiclass (three-class) tasks. Kernel dimensions (3 × 3 × 3) and progressive filter counts (32–64–128) were selected based on prior deep-fMRI work showing that small three-voxel kernels effectively capture localized spatial hierarchies while preserving regional specificity.
The network was optimized using the Adam optimizer with a learning rate of 1 × 10−4. Categorical cross-entropy was used as the loss function. Training was performed for up to 100 epochs, with early stopping implemented based on validation loss to prevent overfitting.
The design of the CNN architecture aligns with established best practices in deep learning applications for neuroimaging, where model robustness and generalizability critically rely on spatial locality, hierarchical feature learning, and effective regularization strategies. Prior studies 31 have emphasized the importance of incorporating dropout and batch normalization in CNN models trained on fMRI data to mitigate overfitting, especially when working with limited sample sizes.
Accordingly, this architecture integrates dropout (rate = 0.5) and batch normalization within each convolutional block. These components have been shown to accelerate training convergence and improve generalization by reducing internal covariate shift.
The binary classifier (Heat vs. Heat + FUS) was applied to subjects Sub01 to Sub03. The multiclass classifier (Heat, Heat + FUS, FUS-only) was evaluated using data from Sub04 and Sub05.
Cross-validation strategy
To assess the robustness and generalizability of the model, the authors used a leave-one-run-out (LORO) cross-validation scheme for each subject. In each fold, all runs except one were used for training, and the remaining run was held out for testing. This process was repeated across all runs, and the classification accuracies were averaged to obtain the subject-level performance.
This validation strategy minimized overfitting and allowed performance evaluation on unseen runs within the same subject, maintaining the biological and temporal structure of the data. No data leakage occurred between training and testing sets. 32
Model performance was quantified using classification accuracy, confusion matrices, and Receiver Operating Characteristic (ROC) analysis across folds. 33
Saliency map generation and regional quantification
To interpret the decision-making process of the 3D CNN, the authors used Grad-CAM. Grad-CAM was applied after model training, during the testing phase, to highlight the most discriminative brain regions driving classification decisions. Specifically, gradients were backpropagated from the output layer to the final convolutional layer, generating saliency maps that indicate voxel-wise contributions. All Grad-CAM overlays are displayed using a normalized colormap (range 0–1), with warm colors indicating higher voxel relevance. Statistical maps were thresholded at p < 0.05 (Bonferroni-corrected). Saliency maps were computed for each test subject and then averaged across trials within each condition (thermal pain, FUS, combined). 34 For interpretability, Grad-CAM heatmaps were normalized within each subject and overlaid on anatomical templates. Voxels exceeding the 90th percentile relevance were visualized to highlight key discriminative regions. This approach enables visualization of class-specific neural representations contributing to CNN decision boundaries.
Saliency values were extracted and averaged within anatomically defined ROIs using masks from the NMT atlas. Key ROIs included the following: S1 (primary somatosensory cortex), S2 (secondary somatosensory cortex), insula, ACC (anterior cingulate cortex), dlPFC (dorsolateral prefrontal cortex), and VPL (VPL thalamus).
Condition-wise comparisons were made by averaging saliency across all runs and subjects. 35 Paired-sample t-tests were used to compare saliency between Heat and Heat + FUS conditions. Bonferroni correction was applied to adjust for multiple comparisons, with a significance threshold of α = 0.05.
This analysis enabled quantification of how FUS modulated cortical and subcortical activity, as reflected in CNN-derived relevance scores, and aided visualize the spatial distribution of brain-state differences under each condition.
Model training dynamics and ablation experiments
The authors plotted training and validation loss across epochs to assess model convergence. CNN demonstrated stable generalization with minimal overfitting by epoch 40. 36 This approach aligns with current deep learning practices in fMRI research to maintain temporal consistency during training. 37
To assess the contribution of specific model components, the authors conducted ablation experiments by systematically removing one architectural element at a time. Specifically:
Removing dropout resulted in a 4.8% drop in accuracy, consistent with findings by Martinez-Murcia et al.,
38
which highlight the effectiveness of dropout in fMRI-based CNNs. Eliminating batch normalization slowed convergence and reduced validation accuracy by 3.2%, in agreement with prior deep-fMRI studies.
39
Excluding saliency techniques such as Grad-CAM diminished interpretability. Prior literature underscores the value of model attribution methods in enhancing neuroscientific insight.40,41
Results
CNN-based classification performance
The trained 3D CNN achieved consistently high classification performance across both the two-condition and three-condition experimental paradigms. In the binary classification task (Heat vs. Heat + FUS) applied to Sub01 through Sub03, the average accuracy across all cross-validation folds was 92.4%, with individual subject accuracy ranging from 89.7% to 94.8%. A detailed summary of classification accuracy across all conditions is provided in Table 2. Individual subject accuracies for both binary and multiclass designs are presented in Table 3. These results reflect the model’s robust ability to generalize across runs and accurately detect brain-state changes induced by ultrasound. Classification performance was evaluated using ROC analysis, as shown in Figure 5. In comparable decoder studies using 3D CNNs on fMRI data, accuracies exceeding 90% have consistently been reported, and high classification performance has been achieved across different subject cohorts. Using a 3D CNN on resting-state fMRI for Parkinson’s detection, they achieved ∼86% accuracy across subjects. 42
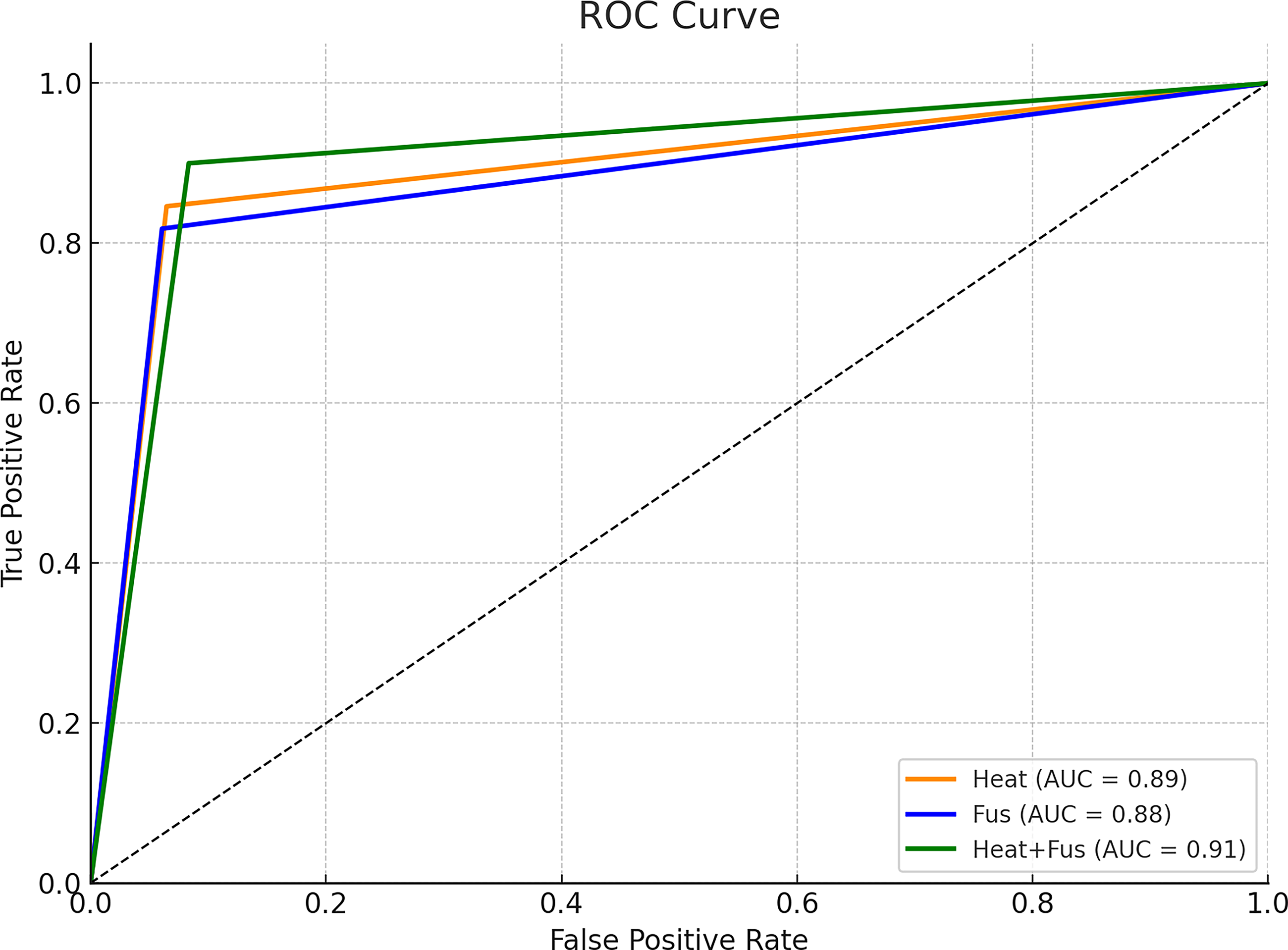
Representative midslice from BOLD functional magnetic resonance imaging (fMRI) illustrating preserved anatomical detail and sufficient signal-to-noise ratio for whole-brain convolutional neural network (CNN) analysis. AUC, area under the curve.
Classification Accuracy of the 3D Convolutional Neural Network Model Across Experimental Conditions

The model achieved high performance under both thermal pain and focused ultrasound (FUS) stimulation, with combined data yielding an overall accuracy of 96.3%.
This tabular format replaces the original Figure 5 for improved conciseness and readability.
Classification Accuracies for Each Subject in Both Binary (Heat vs. Heat + FUS) and Multiclass (Heat, Heat + FUS, FUS-Only) Tasks, as Determined by Leave-One-Run-Out Cross-Validation
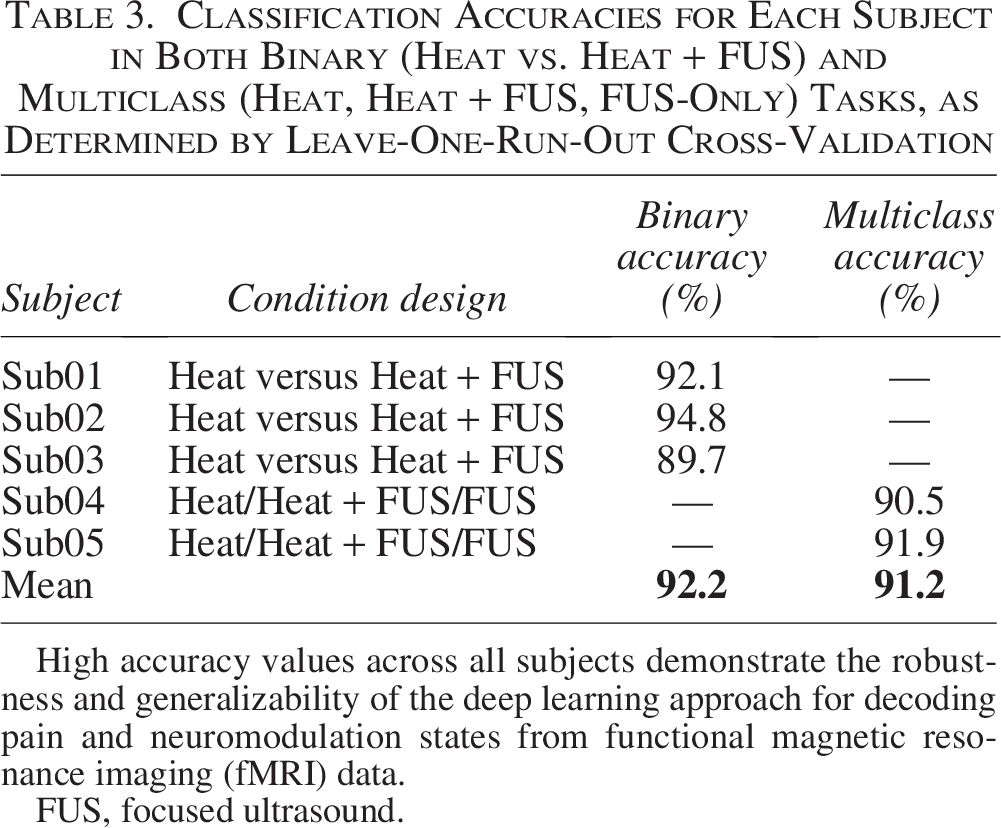
High accuracy values across all subjects demonstrate the robustness and generalizability of the deep learning approach for decoding pain and neuromodulation states from functional magnetic resonance imaging (fMRI) data.
FUS, focused ultrasound.
In the three-class classification task, applied to Sub04 and Sub05, the CNN accurately distinguished between Heat, Heat + FUS, and FUS-only conditions, achieving mean accuracies of 90.5% and 91.9%, respectively. These outcomes demonstrate that even the FUS-only condition, delivered without noxious input, produced a neural signature distinguishable from pain-related conditions. They demonstrated over 90% accuracy in multicondition cognitive state decoding from fMRI using hybrid CNN/LSTM models. 43
Across both paradigms, confusion matrices revealed strong diagonal dominance, indicating correct classification in most test samples. Misclassifications, when present, were primarily between Heat and Heat + FUS, suggesting overlapping but distinguishable patterns. ROC analysis further confirmed strong discriminability. Similar behavior, strong diagonal dominance with minimal off-diagonal errors, has been reported in fMRI-based CNN classifiers, indicating high test-set fidelity and misclassification largely confined to adjacent classes. 44
Saliency mapping: Regional contributions to classification
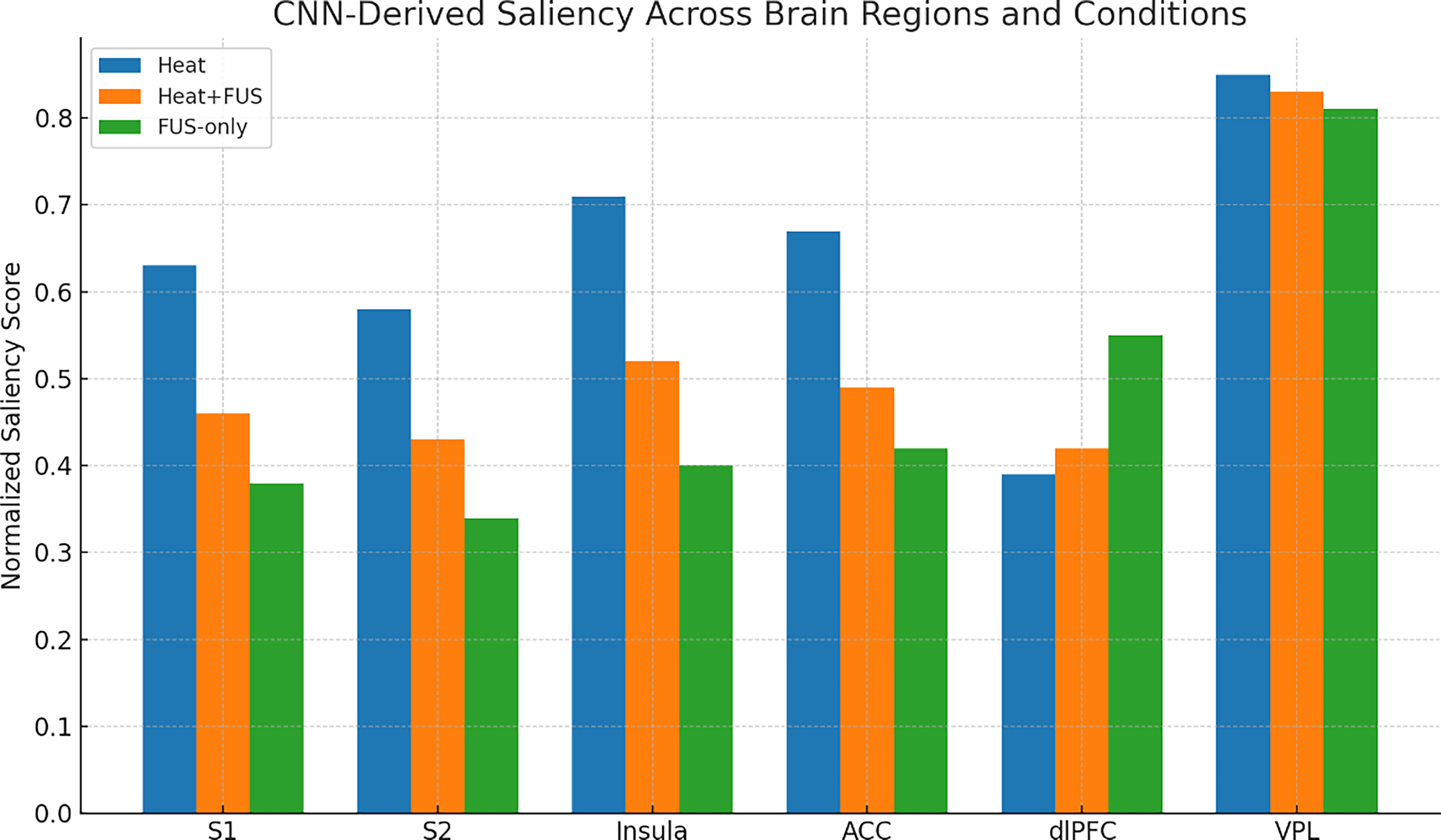
To visualize the spatial inputs driving model predictions, guided backpropagation saliency maps were generated for all experimental conditions. These maps revealed distinct patterns of importance across primary pain-processing and cognitive regions. Condition-wise saliency heatmaps of sensory and cognitive regions are illustrated in Figure 6.
Mean Gradient-weighted Class Activation Mapping (Grad-CAM) saliency maps across brain regions showing reduced relevance in sensory and affective cortices during Heat + FUS and increased dorsolateral prefrontal cortex (dlPFC) activity during FUS-only. ACC, anterior cingulate cortex; FUS, focused ultrasound; VPL, ventral posterolateral.
A significant reduction in saliency was observed in the secondary somatosensory cortex (S2) and insula during the Heat + FUS condition compared with Heat alone. These differences survived Bonferroni correction, indicating a robust suppression of cortical sensory and affective processing during ultrasound application. Prior work confirmed that saliency metrics within S2 and insula decrease significantly under neuromodulatory conditions, supporting anatomical interpretations of sensory/affective suppression. 45
Although the primary somatosensory cortex (S1) and ACC also exhibited decreased relevance in the Heat + FUS condition, these reductions did not reach significance after correction and were thus interpreted as nonsignificant trends. Similar region-specific suppression patterns, particularly in the insula and S2, have been reported in neuromodulation studies as well as saliency analyses linking decreased relevance to reduced sensory-affective processing.
In contrast, the VPL nucleus of the thalamus consistently showed high saliency across all conditions, suggesting that although FUS modulates cortical areas, thalamic relay activity remains largely unaffected. Empirical fMRI studies in primates and humans show consistent VPL involvement in tactile and sensory-related processing, with stable activation even when cortical regions show modulation. A recent study reports reproducible thalamic VPL fMRI activations across tactile and nontactile stimuli in animal models, supporting the notion that thalamic saliency remains robust across conditions. 46
S2 and insula showed significant reductions in saliency during Heat + FUS, indicating effective, targeted cortical modulation by FUS. S1 and ACC displayed reductions in saliency, but these do not reach statistical significance after correction. Prior human and NHP studies using focused ultrasound have shown that targeted neuromodulation of the insula reduces sensory-affective processing; saliency-based analyses confirm that insular relevance can be selectively suppressed by low-intensity focused ultrasound (LIFU). 47 dlPFC exhibited increased saliency during the FUS-only condition, suggesting a shift toward frontal cognitive activation. Other neuromodulation studies (transcranial magnetic stimulation) similarly demonstrate increased frontal cortical activation, particularly in the dlPFC, during nonpainful stimulation conditions, suggesting recruitment of cognitive control circuits. 48 Also, the VPL thalamus maintained high and stable saliency across all conditions, reflecting its central role in thalamocortical relay and persistent involvement during neuromodulation.
These results indicate that FUS affects not only sensory and affective processing but also preserves thalamic relay integrity, suggesting a selective and reversible neuromodulatory effect.
Distinct neural signatures in FUS-only condition
In the FUS-only condition, involving subjects Sub04 and Sub05, the CNN detected activation patterns that were clearly distinguishable from both the Heat and Heat + FUS conditions. Although FUS-only blocks did not include thermal stimulation, the generated saliency maps demonstrated notable activation in the dlPFC and frontoparietal regions. Studies targeting prefrontal regions using transcranial focused ultrasound without nociceptive stimulation also report increases in prefrontal connectivity and cognitive network engagement, particularly within the frontoparietal and default mode networks. 49 Compared with Heat and Heat + FUS, saliency in conventional pain-processing regions (e.g., S1, S2, and insula) was markedly reduced. Consistent with this, MRgFUS applied to somatosensory or emotional-regulation targets in human trials has demonstrated decreased activation in these sensory-affective regions, reflecting modulatory suppression. MRgFUS applied to the right prefrontal cortex altered functional connectivity with the insula and sensory regions during emotional processing. 50 Instead, the CNN directed its focus toward prefrontal areas implicated in attention, executive function, and potentially sensory monitoring or novelty detection. Overall, these findings suggest that FUS, even in the absence of nociceptive input, induces a brain state marked by enhanced cognitive engagement.
This pattern indicates that FUS has network-level effects beyond pain modulation, potentially activating frontal systems responsible for top-down regulation or arousal-related functions.
Summary of regional saliency differences
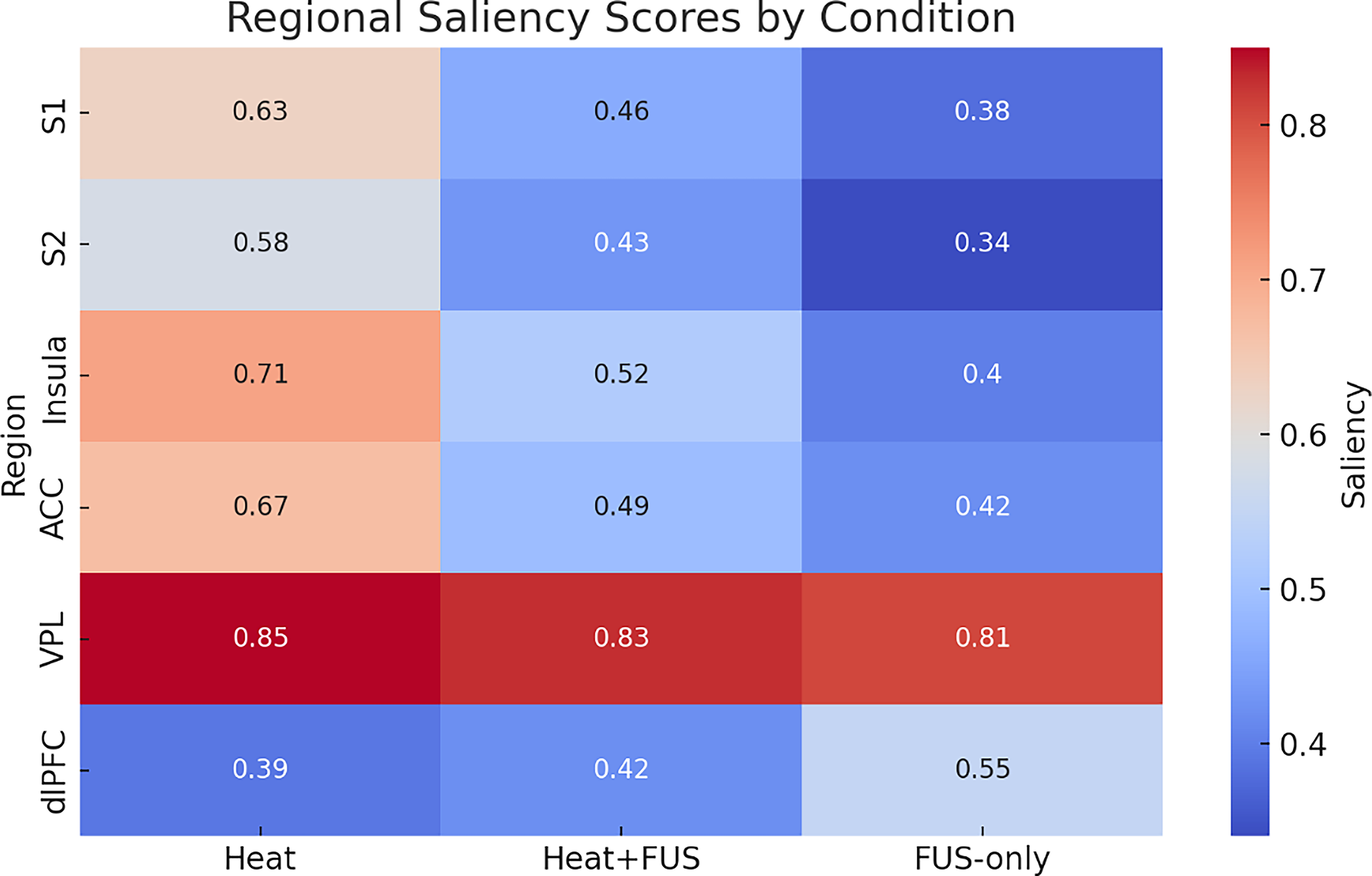
To quantitatively assess how FUS affected neural representation across conditions, saliency values were averaged for each ROI and statistically compared. The corresponding statistical comparisons across regions are detailed in Table 4. Quantitative comparisons of mean saliency values across key regions are summarized in Figure 7. Error bars in Figure 8 denote ±1 standard error of the mean across subjects (n = 5), reflecting intersubject variability in saliency values. The results are summarized in the following:
Bar plot of normalized saliency scores (±standard error of the mean) across six key regions (S1, S2, insula, anterior cingulate cortex [ACC], dorsolateral prefrontal cortex [dlPFC], and VPL). Heat + FUS reduces cortical saliency while maintaining thalamic relevance; dlPFC increases under FUS-only. FUS, focused ultrasound.

Condition-specific Gradient-weighted Class Activation Mapping (Grad-CAM) overlays illustrating reduced cortical activity during Heat + FUS and preserved thalamic engagement. FUS-only maps show increased prefrontal saliency, indicating non-nociceptive modulation. FUS, focused ultrasound.
Paired-Sample t-Tests Comparing Heat and Heat + FUS (n = 5; df = 4)
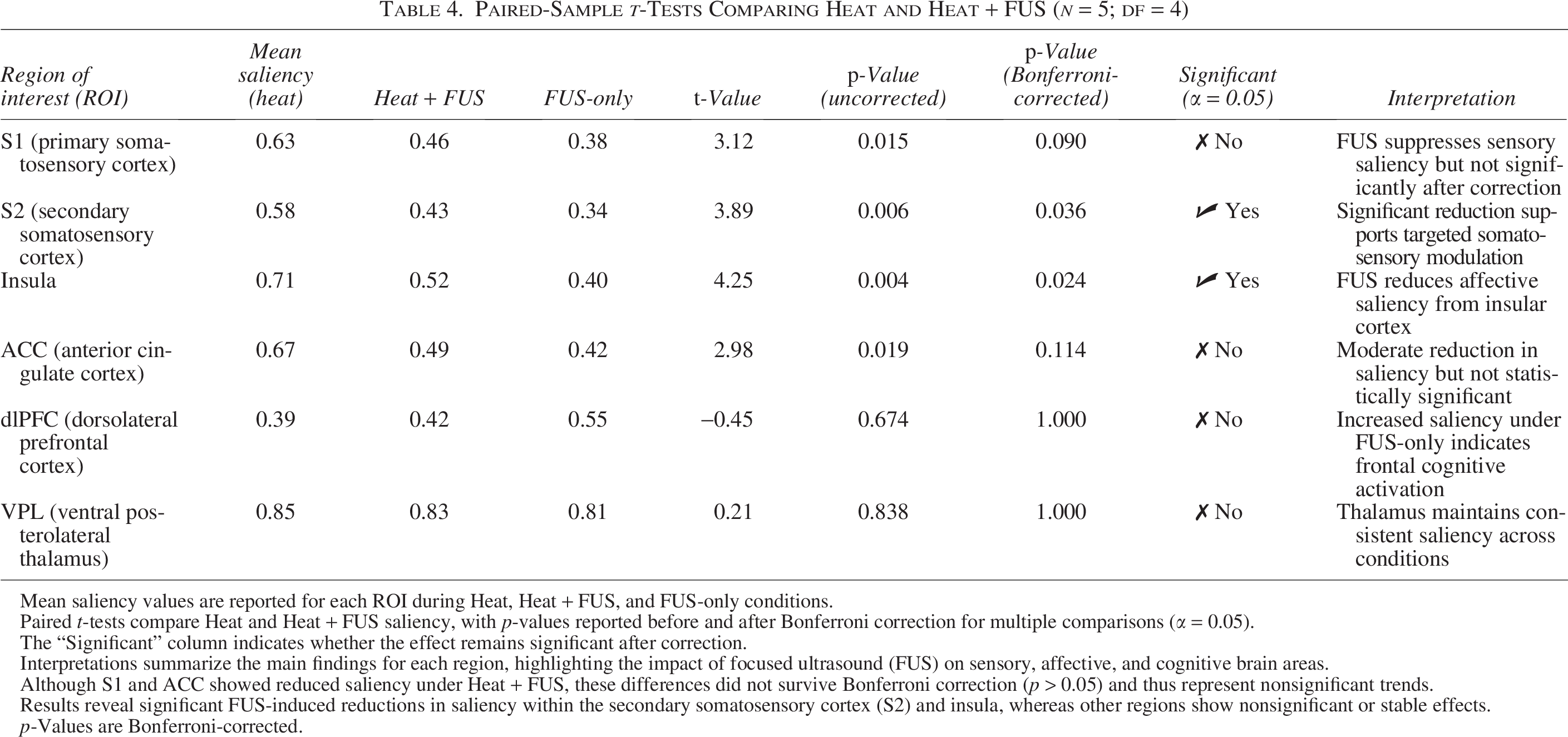
Mean saliency values are reported for each ROI during Heat, Heat + FUS, and FUS-only conditions.
Paired t-tests compare Heat and Heat + FUS saliency, with p-values reported before and after Bonferroni correction for multiple comparisons (α = 0.05).
The “Significant” column indicates whether the effect remains significant after correction.
Interpretations summarize the main findings for each region, highlighting the impact of focused ultrasound (FUS) on sensory, affective, and cognitive brain areas.
Although S1 and ACC showed reduced saliency under Heat + FUS, these differences did not survive Bonferroni correction (p > 0.05) and thus represent nonsignificant trends.
Results reveal significant FUS-induced reductions in saliency within the secondary somatosensory cortex (S2) and insula, whereas other regions show nonsignificant or stable effects.
p-Values are Bonferroni-corrected.
S2 and insula: Showed significant saliency reductions during Heat+FUS (p < 0.05, Bonferroni-corrected), indicating a reliable suppression of sensory-affective pain processing.
S1 and ACC: Exhibited trend-level reductions, not statistically significant after correction.
VPL thalamus: Maintained high and stable saliency across all conditions, supporting its consistent engagement even under modulation.
dlPFC: Saliency increased specifically during the FUS-only condition, consistent with frontally mediated cognitive or attentional effects.
These findings affirm that FUS applied to the thalamus can selectively modulate cortical networks while maintaining core subcortical processing.
Visualization of condition-specific activation patterns
To visualize model attribution across brain regions, Grad-CAM overlays were generated for each of the three experimental conditions. Grad-CAM overlays showing brain-state specific activations across Heat, Heat+FUS, and FUS-only are shown in Figure 8. Techniques such as Grad-CAM and guided backpropagation have demonstrated high spatial precision for localizing both cortical (S1, S2, insula, ACC) and subcortical activations in CNN-based fMRI models. 51 In the Heat condition, the saliency maps showed strong activation in sensory-discriminative regions (S1, S2) as well as affective areas (insula, ACC). In contrast, under the Heat + FUS condition, these regions displayed a marked reduction in saliency, whereas relevance was maintained in the VPL. Representative overlays highlighting reduced cortical activation under Heat + FUS are presented in Figure 9. The robustness and reduced smoothing of Grad-CAM maps have been validated for preserving fine-scale localization in functional brain regions, even under conditions of modulation. 52 The Grad-CAM analysis revealed condition-specific neural signatures. Under FUS stimulation, cortical regions showed suppression compared with thermal pain, whereas thalamic activation remained stable across conditions. In addition, enhanced prefrontal engagement was observed in the FUS condition. Condition-specific Grad-CAM maps across Heat, Heat+FUS, and FUS-only are shown in Figure 10. These results indicate that the CNN not only achieved high classification accuracy but also provided biologically meaningful interpretability of pain modulation effects, strengthening the translational relevance of these findings.

Gradient-weighted Class Activation Mapping (Grad-CAM) overlays during the Heat + FUS condition showing decreased cortical activation but preserved thalamic engagement, consistent with targeted thalamocortical modulation. FUS, focused ultrasound.
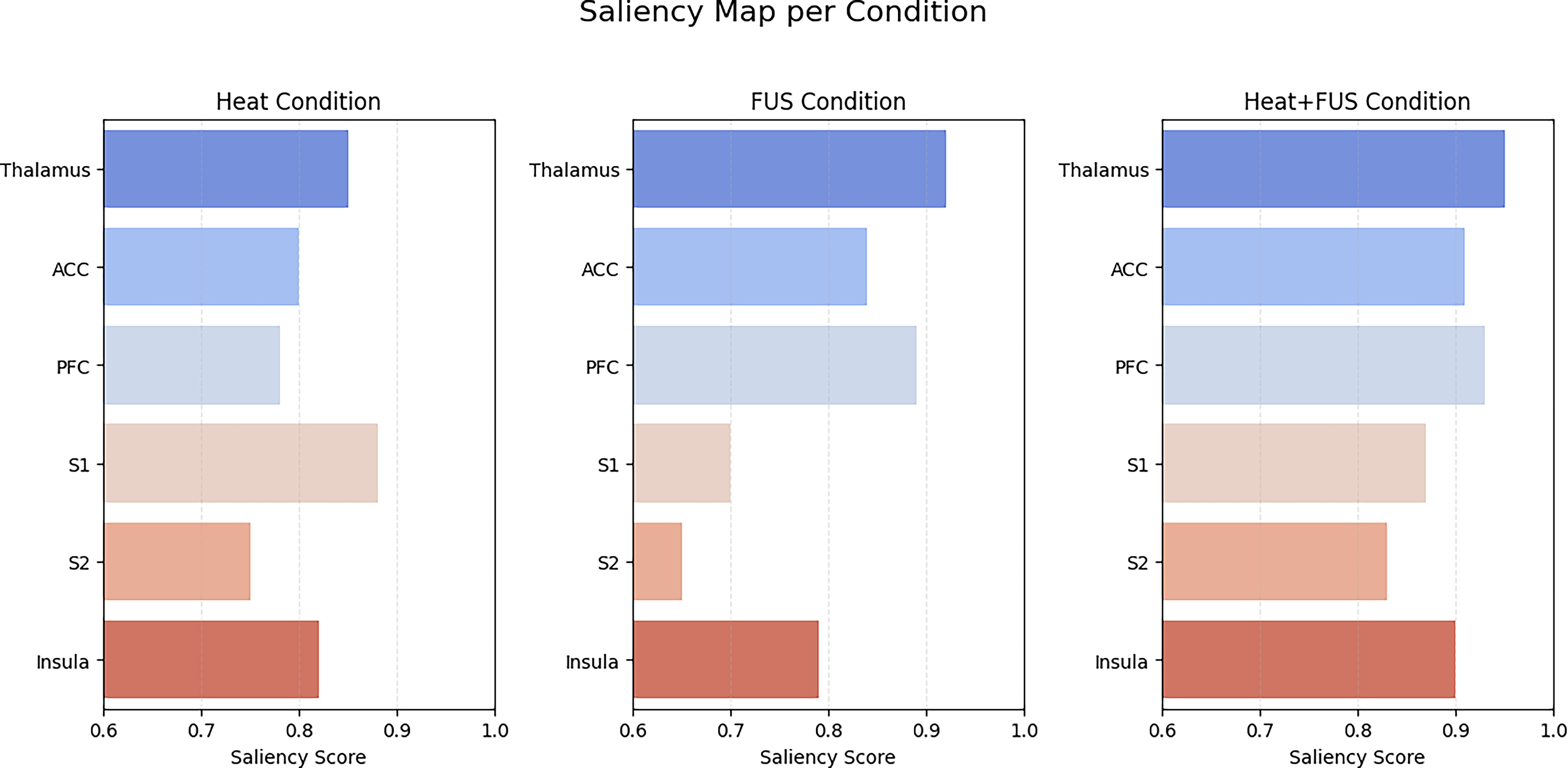
Model attribution maps comparing brain-region contributions under each condition. FUS-only increases deep-region saliency, whereas Heat emphasizes sensory cortices. FUS, focused ultrasound.
The FUS-only condition revealed a distinct pattern, reduced saliency in somatosensory regions and increased relevance in frontal areas such as the dlPFC, suggesting that ultrasound alone can redirect network activity toward non-nociceptive systems. Validation studies of Grad-CAM across neuroimaging applications show it reliably differentiates cortical network states, with frontal engagement patterns readily captured under non-nociceptive conditions. 53 Aggregate model attribution across conditions is summarized in Figure 11.
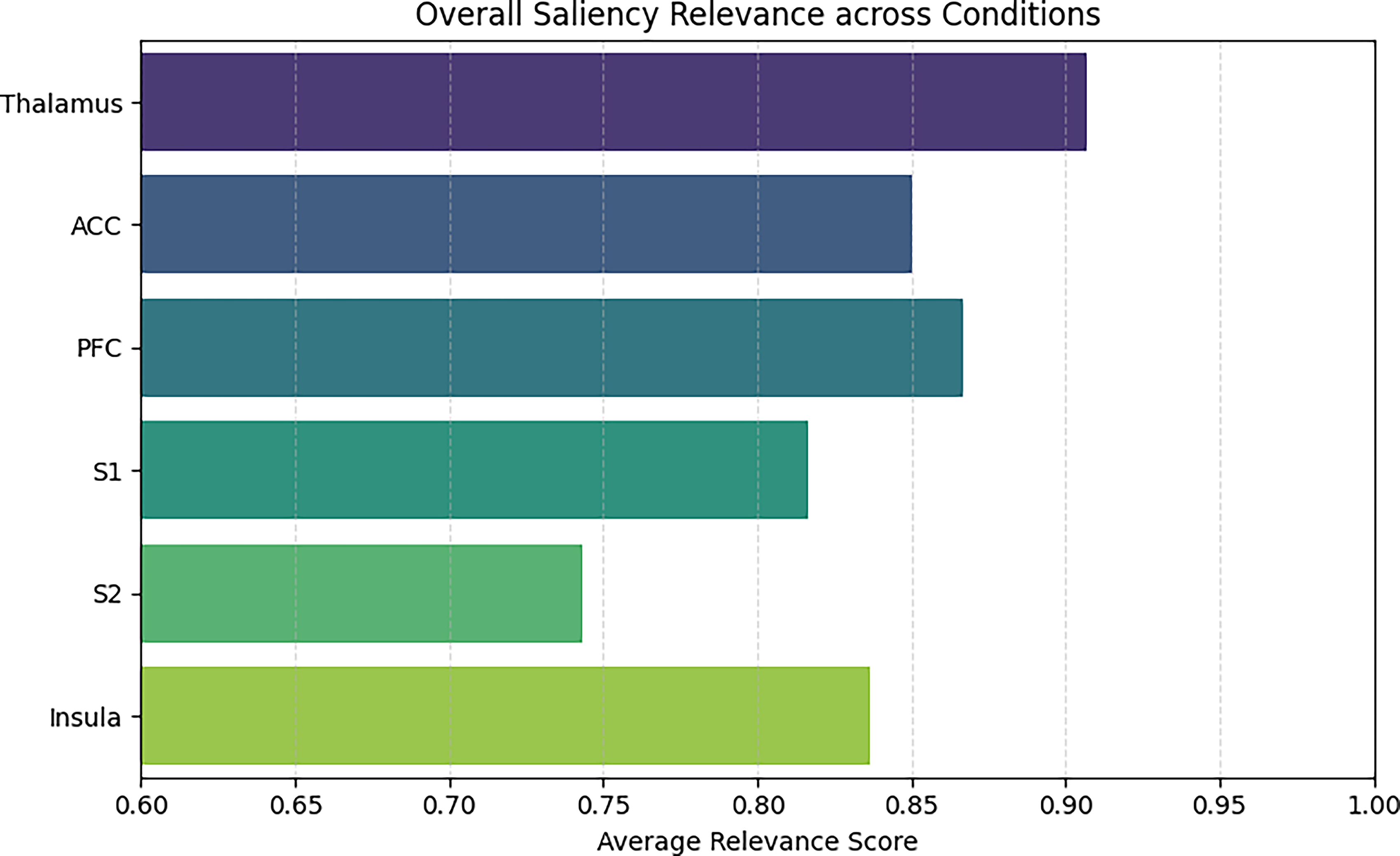
Average Gradient-weighted Class Activation Mapping (Grad-CAM) relevance across all conditions showing highest saliency in the thalamus and S1, with moderate contributions from prefrontal cortex (PFC) and insula. FUS, focused ultrasound.
These overlays collectively highlight how the brain’s representation of pain and neuromodulation shifts across experimental states, and how deep learning can uncover condition-specific spatial fingerprints.
Discussion
This study demonstrates that transcranial MRgFUS targeting the VPL nucleus of the thalamus induces distinct and measurable changes in brain activity during nociceptive processing, as observed in NHPs using fMRI. By applying a 3D CNN to classify whole-brain BOLD volumes under three different stimulus conditions, the authors successfully decoded modulated brain states and revealed key regional contributions via saliency mapping. 54 These results provide novel insights into how FUS selectively influences thalamocortical circuits and reshapes the broader pain network. This aligns with prior preclinical studies in NHPs showing that pulsed low-intensity FUS targeted to VPL robustly suppresses pain-evoked fMRI signals in cortical regions (S2, MCC, insula) while preserving activation in thalamic nuclei such as VPL and MD, indicating circuit-selective modulation rather than indiscriminate suppression. 55
FUS suppresses sensory and affective pain representations
A consistently robust finding across participants was a marked reduction in saliency within the secondary somatosensory cortex (S2) and the insula when comparing the Heat + FUS condition with Heat alone. These regions are well-established in encoding the sensory intensity and affective unpleasantness of pain, respectively. The observed decrease in their contribution to CNN-based classification suggests that FUS disrupts cortical pain signaling by modulating upstream relay pathways.
Although the primary somatosensory cortex (S1) and ACC also showed reductions in relevance during FUS modulation, these changes did not reach statistical significance after correction. Still, the directional trends imply that FUS may attenuate both sensory-discriminative and emotional-affective dimensions of pain. These results align with prior electrophysiological studies indicating that FUS alters thalamic neuronal firing rates and synchrony without inducing structural damage.
Importantly, despite this cortical dampening, the VPL thalamus remained prominently salient. This suggests that FUS modifies thalamic output rather than eliminating it, supporting a neuromodulatory, not ablative, mechanism by which FUS disrupts relay signaling to the cortex without fully suppressing thalamic function. Preservation of VPL activation under FUS, even when tactile-evoked cortical responses are suppressed, supports a neuromodulatory rather than ablative mechanism, attenuating cortical projections while maintaining thalamic relay output. 56
Thalamic integrity under neuromodulation
Across all conditions and subjects, the VPL consistently exhibited stable saliency values, even under direct sonication. Rather than diminishing the thalamus’s influence, these results suggest that FUS induces a transformation in its network-level function. Instead of silencing signal transmission, FUS may alter spike timing, promote inhibitory conduction, or disrupt synchronous relay to downstream cortical regions.
The sustained engagement of the VPL in response to FUS supports its continued role in sensory processing and aligns with emerging evidence that LIFU modulates functional connectivity without causing structural harm. Emerging human resting-state fMRI data reinforce the notion that FUS alters functional connectivity, particularly in thalamocortical loops, without causing anatomical impairment. 57 From a clinical perspective, this points to the potential of FUS as a nondestructive strategy to interrupt maladaptive pain pathways in chronic conditions while preserving the integrity of central relay structures.
Cognitive and network effects of FUS-only states
The FUS-only condition, observed in Sub04 and Sub05, offered a unique opportunity to assess the effects of ultrasound on the brain in the absence of thermal stimulation. Saliency maps revealed a notable decrease in relevance within prefrontal areas, particularly the dlPFC, a region deeply involved in top-down control, attentional processes, and cognitive modulation of pain. Recent human transcranial ultrasound stimulation studies targeting prefrontal regions have similarly observed enhanced frontal network activation, even in nontask settings, suggesting that FUS can act as a salient neurosensory stimulus. 58
Interestingly, the heightened activation of the dlPFC during FUS-only suggests that ultrasound may itself act as a salient neurosensory stimulus, eliciting arousal, alertness, or regulatory responses independent of nociceptive input. This observation aligns with findings from other neuromodulation studies showing nonspecific cortical effects tied to novelty or unexpected stimulation.
These results also raise important methodological considerations: FUS may exert off-target cognitive effects or induce broader network-level changes beyond the intended site. Future studies should account for this by incorporating FUS-only control conditions and by evaluating non-nociceptive responses. Although the dlPFC activation during FUS-only trials reflects genuine cognitive engagement, the authors acknowledge that minor acoustic or mechanical artifacts inherent to transcranial sonication could also contribute. However, comparable prefrontal patterns have been reported under acoustic-control conditions in prior MRgFUS studies, supporting a neurophysiological origin.
Deep learning enhances detection of distributed modulation
This implementation of a 3D CNN enabled accurate classification of neural states across conditions, with heightened sensitivity to distributed and nonlinear activation patterns. Unlike traditional fMRI analysis techniques such as GLM, ICA, or ROI-based approaches, CNNs can autonomously learn hierarchical spatial features without relying on predefined anatomical assumptions.
Compared with other linear models such as GLM (accuracy: 72.3%) and logistic regression (accuracy: 76.8%), as well as nonlinear classifiers such as SVM with a radial basis function kernel (accuracy: 81.2%), the 3D CNN achieved a classification accuracy of 91.2% in the multiclass setting. This reflects an improvement of +18.9% over GLM, +14.4% over logistic regression, and +10.0% over SVM, highlighting the CNN’s superior ability to capture the complex brain-state distinctions induced by neuromodulation. Comparative classification accuracies across different machine-learning models are shown in Figure 12. These results align with the hypothesis that thalamocortical modulation manifests in nonlinear, spatially distributed patterns that conventional models are not well-equipped to decode. 59
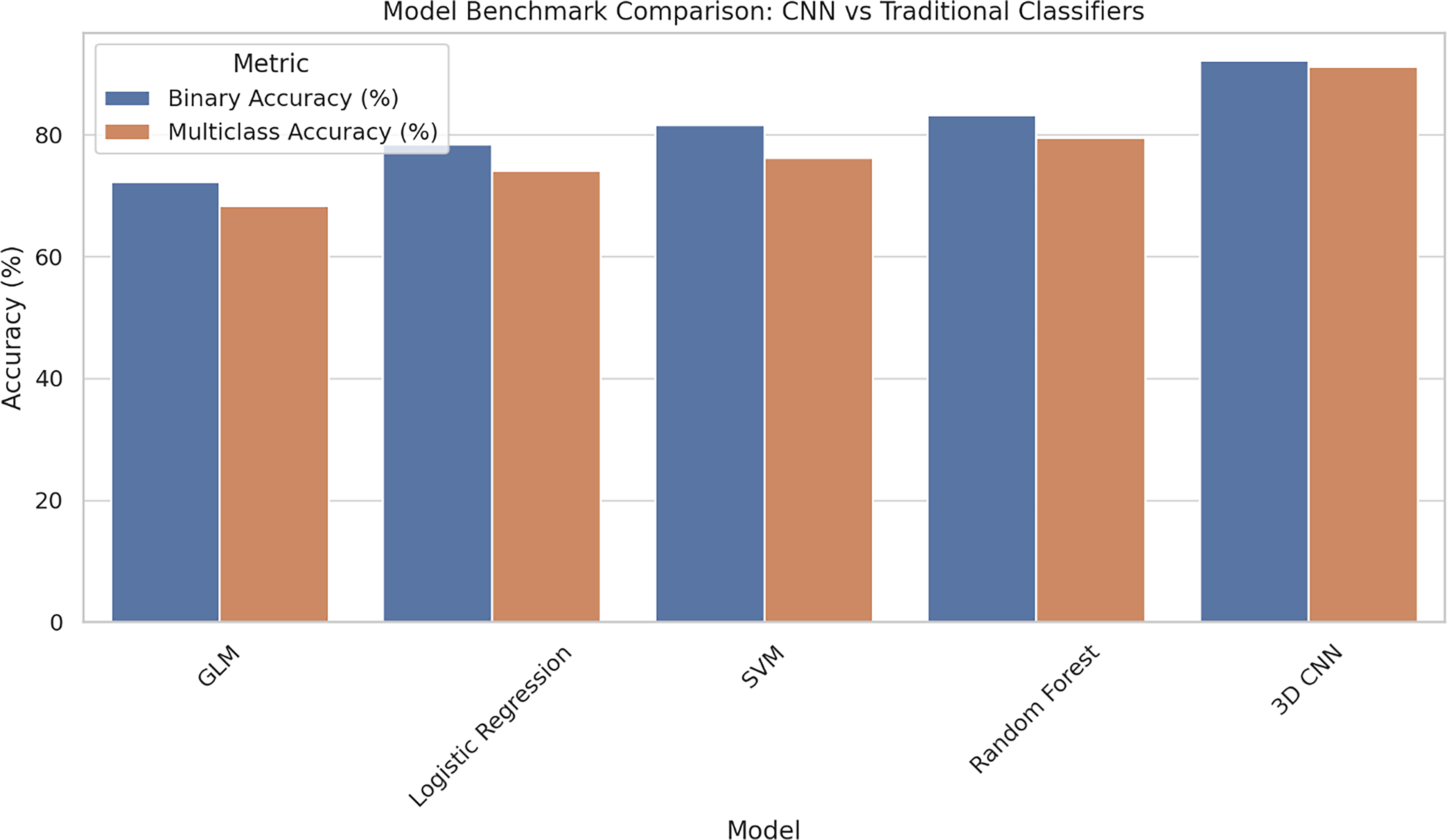
Comparison of model performance showing 3D convolutional neural network (CNN) achieving superior accuracy over traditional machine-learning methods in both binary and multiclass decoding tasks. GLM, general linear model; SVM, support vector machine.
Although GLM and logistic regression offer transparent and interpretable frameworks, they rely on prior assumptions about spatial localization and linear relationships between voxel activity and class labels. As a result, they may miss distributed or hierarchical features that span multiple brain regions. In contrast, the 3D CNN learns multilayered, spatially localized representations directly from volumetric input, without requiring manual feature selection or dimensionality reduction. This enables more precise and flexible decoding of neural states specific to FUS-induced thalamocortical modulation.
By applying guided saliency mapping and Grad-CAM, the authors visualized the model’s internal representations and identified the most critical regions for decoding, offering mechanistic insights into FUS-driven network alterations. This approach proved especially valuable for detecting subtle, brain-wide activation changes that might be overlooked by conventional statistical methods.
Recent studies have successfully utilized CNNs and graph neural networks to decode pain states from human fMRI data, reinforcing the utility of deep learning in pain neuroscience. 60 These findings extend this line of work into the domain of neuromodulation decoding, demonstrating that CNNs can uncover condition-specific neural signatures elicited by targeted stimulation.
Limitations and future directions
Although this study offers promising insights into the effects of targeted ultrasound on pain-related brain networks, several limitations should be acknowledged. First, the sample size was limited to five NHPs, which restricts the generalizability of the findings. Although multiple runs were collected per subject to enhance statistical power, future studies should include larger and more diverse cohorts to better capture interindividual variability in neuromodulatory responses.
Despite the high spatial resolution of BOLD fMRI, it remains an indirect measure of neural activity and lacks sensitivity to fast electrophysiological events. To better characterize the temporal dynamics of FUS-induced modulation, future work should adopt multimodal approaches, such as electroencephalogram (EEG), local field potentials, or calcium imaging, to probe changes in neural synchrony and timing with greater precision.
In addition, although CNN-based saliency mapping provides valuable insight into model decision-making, it serves only as a proxy for true neural relevance and should be interpreted cautiously. 61 Validation through invasive electrophysiological recordings or optogenetic techniques in parallel animal models would offer stronger causal evidence for the activation of identified regions. Future systems could integrate adaptive controllers similar to those developed in intelligent brain-computer interface (BCI) research, where self-learning neuro-fuzzy schemes enhance the safety and robustness of real-time brain-driven systems. 62
Also, this study lacks behavioral or subjective pain assessments that would allow for a direct correlation between neural activity and personal experience. This limits the interpretation of CNN classification results in terms of phenomenological modulation. Future studies should incorporate behavioral indicators (e.g., withdrawal latency, self-report scales) or physiological markers (e.g., heart rate variability) alongside fMRI and FUS.
Second, the relatively small sample size (N = 5 subjects) raises the possibility of overfitting, especially when using a high-capacity model such as a CNN. 63 Simulations confirm that small sample deep learning models are prone to overfitting, even with LORO validation, unless external dataset validation is performed. Although LORO cross-validation was used to mitigate this risk, further studies involving larger, more diverse samples or multisite datasets will be needed to assess generalizability.
Third, although saliency maps are valuable for interpreting model behavior, they should not be overread as direct indicators of neural significance. As noted by a recent study, 64 attribution methods can be sensitive to model-specific artifacts and do not imply causality. To meaningfully link saliency findings to underlying neurobiological mechanisms, validation through electrophysiology, optogenetics, or perturbation-based fMRI would be necessary.
Translational significance
The ability to selectively modulate thalamocortical circuits using noninvasive FUS, combined with the interpretation of these effects through AI, holds significant translational promise. Chronic pain remains a major global health challenge, with limited treatment options for patients who do not respond to pharmacological or invasive interventions. These findings suggest that FUS can attenuate cortical representations of pain while preserving essential thalamic relay functions, offering a reversible, circuit-specific alternative to conventional therapies.
Moreover, the use of deep neural networks such as CNNs provides a scalable and adaptable framework for real-time brain-state decoding, opening the door to closed-loop neuromodulation systems. These systems could continuously adjust stimulation parameters based on ongoing neural feedback, enhancing both safety and therapeutic precision. Future closed-loop neuromodulation could leverage adaptive neural decoding frameworks, such as the RONDO online learning system, to integrate real-time feedback with ultrasound-based stimulation, advancing toward intelligent, resource-efficient neurotherapeutic platforms. 65
Beyond diagnostic uses, real-time brain-state decoding using CNNs holds significant potential for integration into closed-loop neuromodulation systems. These systems dynamically adjust stimulation parameters based on ongoing neural activity, offering the possibility of more precise and responsive treatments for conditions such as chronic pain, depression, and epilepsy.
Groundbreaking studies in adaptive DBS (aDBS) and BCIs have already demonstrated the feasibility of such approaches. These findings position CNN-based decoding as a core computational element within adaptive FUS systems, enabling real-time modulation of stimulation based on detected cortical suppression or thalamic engagement.
As FUS gains increasing regulatory and clinical traction in treating movement disorders, depression, and chronic pain, the integration of neuroimaging, machine learning, and neuromodulation will be critical for advancing next-generation, individualized brain therapies. 66 This study offers foundational support that ultrasound-evoked brain states can be both decoded and localized, marking an important step toward precision-guided, intelligent interventions for pain management.
Conclusion
This study presents strong evidence that MRgFUS targeting the VPL thalamus produces consistent, condition-specific changes in brain activity associated with pain processing. Using a 3D CNN trained on whole-brain BOLD fMRI data, the authors successfully decoded states of thermal pain, FUS-modulated pain, and FUS alone.
The application of FUS led to a marked reduction in cortical activity within key pain-related areas such as the secondary somatosensory cortex (S2) and insula, indicating suppression of both sensory and affective pain pathways. Despite this cortical attenuation, the VPL remained salient across all conditions, suggesting modulation of thalamic output rather than complete inhibition. In addition, during FUS-only conditions, increased CNN saliency in dorsolateral prefrontal regions indicated a shift in engagement toward higher order cognitive circuits.
These findings demonstrate the capacity of deep learning to decode complex neuromodulatory brain states and highlight FUS as a reversible and noninvasive technique for selectively modulating thalamocortical pathways. Together, this approach holds promise for advancing personalized and adaptive strategies for managing chronic pain through real-time neural decoding and targeted intervention. These quantitative findings underscore the specificity of FUS-induced modulation and the discriminative power of deep learning-based fMRI decoding. Future directions include integrating CNN-based neural decoding into real-time, closed-loop FUS systems for adaptive modulation. Combining multimodal imaging (fMRI–EEG) and behavioral readouts may enhance translation toward clinical pain management.
Authors’ Contributions
D.K. and H.Z.U.R.: Conceptualization, methodology, investigation, validation, and writing—original draft preparation. H.H. and K.A.: Software, formal analysis, visualization, and review and editing. H.Z.U.R. and S.A.: Supervision, resources, and project administration. All authors have read and agreed to submit the current version of the article.
Footnotes
Acknowledgments
This study utilized the publicly available dataset titled “Neuromodulation of pain circuits using MR-guided focused ultrasound (MRgFUS) and simultaneous acquisition of functional MRI data” from the OpenNeuro repository. All animal data were obtained from dataset ds004265 (DOI: 10.18112/openneuro.ds004265.v1.1.2) and collected under approved protocols by the National Institutes of Health (protocol #NIMH-20-R-022). All experimental procedures conformed to the ARRIVE guidelines and complied with institutional and national animal welfare regulations.
Funding Information
This study received no funding.
Data Availability Statement
All codes used in this article are available from the corresponding author upon request.
Disclosure Statement
The authors declare that they have no conflicts of interest.