
Abstract
Background:
Melatonin (N-acetyl-5-methoxy-tryptamine) is a promising antitumor agent and antioxidant. The pharmacological dose of melatonin has an oncostatic function by various means, including angiogenesis. Evidences indicate that melatonin, using multiple interrelated mechanisms, exhibits a variety of oncostatic properties in a variety of tumors during different stages of progression.
Methodology:
The present study was undertaken to investigate the epigenetic modulations in the vascular endothelial growth factor VEGF, TIMP3, and CDH1 genes in Dalton lymphoma (DL) cells; furthermore, melatonin may reverse these epigenetic modulations. Endothelial cells (ECs) are the precursors for the development of new blood vessels; therefore, an EC proliferation assay was performed with and without DL ascites (DLA). EC migration leads to capillary formation, in which matrix metalloproteinases (MMPs) play a very important role in providing space for EC proliferation and migration. Furthermore, ECs were checked for MMP-9 secretion through enzyme-linked immunosorbent assay. The MMP activity was also confirmed by gelatin zymography. Reactive oxygen species (ROS) level was also measured on ECs in the presence and absence of DLA. These activities of ECs or the tumor microenvironment were regulated by growth factors, such as VEGF secretion by cancer cells and the inhibition or silencing of endogenous antiangiogenic genes. In this, chromatin modification plays a very important role; also, in this, DNA methylation and histone deacetylases (HDACs) play a significant role. So, in the DL cells, the authors try to find the HDACs and DNMTs levels with respect to the control thymus cells by Reverse Transcription-polymerase chain reaction (RT-PCR). Further DNA methylation was checked by the MS-PCR technique. Proteins and melatonin were studied using VINA 2.0 software.
Results:
DLA promote a significant increase in EC proliferation and a high amount of MMP-9 in the presence of DLA. The gelatin zymography also showed the high activity of MMP-2 and MMP-9 with respect to normal ECs. The ROS levels are also found to be high in DLA-induced ECs. The melatonin treatment successfully decreases EC proliferation, MMP-9 levels, and the MMP-2 and MMP-9 activities. The ROS levels were also reduced. The RT-PCR analysis showed that the melatonin modulates the alterations in HDAC1, 2, 6, and 9 and DNMT1, DNMT3A and DNMT3B with respect to the control. The MS-PCR results signify one of the possible ways of action of melatonin on DL cells.
Introduction
Melatonin (N-acetyl-5-methoxy-tryptamine) is a major pineal lipophilic hormone that regulates various biological activities involving hypothalamic–pituitary, neuroendocrine regulation in animals, and various physiological processes.1,2 It is mainly synthesized and secreted by the pineal gland and also produced by other cells and tissues, including the bone marrow, retina, gastrointestinal tract, platelets, and lymphocytes. 3 It is highly lipid soluble in nature, and so, it can easily cross the blood–brain barrier. The evidences confirm that cerebrospinal fluid, as well as adipose tissue, has high concentrations of melatonin with respect to blood. These evidences of elevated levels of melatonin in certain tissues explain the requirement of high concentrations of melatonin to obtain the antitumor or oncostatic effect of melatonin. 4 Apart from animals, plants such as rice, corn, ginger, and barley grains have the highest concentrations of melatonin, at 149.8, 187.8, 142.3, and 87.3 ng/100 g, respectively. Pomegranate and strawberry have low levels of indolamines (8–12 μg/g serotonin, 4–9 μg/g tryptamine, and 13–29 ng/100 g melatonin).5,6 Also, both vegetative and reproductive growth development processes are affected by the exogenous application of melatonin. 7
A plethora of research showed that melatonin is associated with various pathological changes, including sleep deprivation and circadian rhythm disruption, and its suppression is associated with increased risk of cancer development. 8 Years ago, it was hypothesized that melatonin deprivation or high exposure to artificial light results in an increase in cancer incidence. It is widely studied for its protective nature in various pathophysiological conditions, including several types of cancer, such as breast, hepatic, and gastric carcinoma.9,10 Several reports suggest that pinealectomy stimulates breast cancer development. Studies confirm that melatonin has the potential to inhibit cancer cell growth and proliferation and induce apoptosis. Melatonin can successfully inhibit estradiol-induced MCF-7 breast cancer cell and BF-1 ovarian cancer cell proliferation by several mechanisms, including cell cycle arrest, enhancing DNA stability, inhibition of angiogenesis, increasing p53, antioxidant activity, inhibiting aromatase activity, and improving immune responses.11,12 Melatonin acts in both a receptor-dependent and a receptor-independent manner. It also scavenges reactive oxygen species (ROS) and reactive nitrogen species (RNS) independent of receptor-mediated stimulus by direct neutralization or activates antioxidant defense mechanism by increasing detoxifying enzyme activity.12,13 Studies show that the pharmacological dose of melatonin successfully inhibits the growth of tumors associated with mammary glands and liver cancer Melatonin acts as an oncostatic by regulating the expression and activity of various signaling molecules secreted by cancer and proximal cells, especially cytokines, which regulate aromatase enzyme activity in the surrounding fibroblast and endothelial cells (ECs). Recent studies reported that melatonin has paracrine interaction with vascular endothelial growth factor (VEGF) signaling and matrix metalloproteinase (MMP-2 and MMP-9) activity to inhibit EC proliferation and migration. Melatonin shows antiangiogenic effect in several types of cancer cells, such as human colon cancer cells (HCT116), human pancreatic carcinoma cells (PANC-1), human alveolar adenocarcinoma cells (A549), adenocarcinomic human alveolar basal epithelial cells, and cervical cancer cells (HeLa). It regulates the tumor microenvironment by regulating the pro- and antiangiogenic growth factors in both the tumor cells and in proximal ECs. Melatonin also shows proapoptotic properties via upregulating cell death-promoting processes, such as increasing the expression of CHOP and caspase-3, and downregulating proliferation-promoting processes, such as COX-2 and Bcl-2.
Oxidative stress plays a major role in carcinogenesis; reported evidence recommends preventive measures in oxidative stress management.14,15 Prolonged oxidative stress may cause damage to large biomolecules such as DNA, lipids, and proteins, resulting in a high risk of cancer development. Scientific evidence showed that ROS cause oxidation of DNA, consequently in adenine to guanine transformation leading to G to T transversion. Cancer cells show 35%–50% oxidized adenine and guanine, which causes mutagenesis. ROS also influence epigenetic modulators, such as DMNTs, leading to alterations in gene expression. Epigenetic mechanisms are essential for the normal development and maintenance of tissue-specific gene expression patterns in mammals. Epigenetics is the causal interaction between genes and their products, which brings the phenotype into being. There are piling reports that have proved the association of epigenetic aberrations with the development of chronic disorders, especially in the early stages of life. 16 Epigenetic changes get perturbed easily in response to a change in the microenvironment of its cell. Epigenetic aberrations pave the path for cancer development or the development of chronic diseases later in life. 17
Global changes in the epigenetic landscape are a hallmark of cancer. These genetic and epigenetic alterations interact at all stages of cancer development, working together to promote cancer progression. Due to the reversible nature of epigenetic modifications and early incidence during the process of cancer development, epigenetic modifications have taken center stage as promising drug targets for cancer prevention. 18 Therefore, antioxidants, both exogenous and endogenous, can be effective in protection against oxidative stress, resulting in either inhibition or delay of cellular damage or reversal of the epigenetic landscape.19–21 In this study, the authors first evaluate the reversal of promoter methylation status with melatonin in Dalton’s lymphoma (DL) cells. DL, a kind of spontaneous T cell lymphoma, is well characterized, easily transplantable, and reproducible and has been considered to be a good and interesting model for screening of various drugs in cancer research because of its practicality in a preclinical system for evaluation of potential drugs for cancer treatment in a short period. In addition, many specific assays can be designed and performed to measure the efficacy of drugs.
Materials and Methods
Chemicals
Melatonin, Dulbecco's modified eagle medium (DMEM), fetal bovine serum (FBS), and antibiotics were purchased from Sigma-Aldrich. Melatonin, primers for VEGF, VEGFR, FGF, CDH1, TIMP3, β-ACTIN, and antibiotics were purchased from Sigma-Aldrich. Methanol, crystal violet, acridine orange, propidium iodide, and dihydroethidium (DHE) were purchased from HiMedia. The DL cell line was taken from Prof. Ajit Sodhi, Banaras Hindu University, Varanasi, India.
Animals
The animal experiments were carried out with BALB/c mice aged 3 to 4 months old of either sex, housed in a polypropylene cage, 3 to 4 animals per cage, obtained from the Animal House Facility of the Department of Zoology, University of Delhi, India. The animals were maintained under standard laboratory conditions, provided with a standard pellet diet, water ad libitum, temperature, and humidity, with an alternating 12-h light/dark cycle in accordance with the guidelines laid by the Animal Ethics Committee, University of Delhi, and protocols approved by the Committee for the Purpose of Control and Supervision of Experiments on Animals, India. 1
Maintenance of DL and collection of ascitic fluid
DL cells were sustained in the peritoneum of BALB/c mice by continuously passaging tumor cells through intraperitoneal injection, as described previously. 1 Briefly, 1 mL of 1 × 106 cells/mL of DL cell suspension (in Phosphate Buffered Saline [PBS]) was injected into the peritoneal cavity of mice and left to grow for 18–21 d. Increased body weight and abdominal swelling signified the development of DL, which was clearly visible from 10 to 11 d post-transplantation, and these mice survived for 26 ± 3 d.
To get DL ascites (DLA), the peritoneal lavage was drawn from mice having a fully grown tumor (18 ± 2 d) and was centrifuged at 3000 g for 10 min at 40°C. Supernatant was collected and filtered through a 0.45 µm membrane filter. Collected DLA was aliquoted and stored at −200°C until used in experiments. 1
EC proliferation assay
Most of the angiogenic studies were performed on human umbilical vein endothelial cells, which are already dividing cells. The ECs from adults are the true target with respect to tumor-induced angiogenesis. For this, the ECs from mice are considered best to understand the angiogenic impact of DL cells, as the in vitro conditions mimic the real tumor conditions. The ECs were isolated from the main thoracic aorta of mice following the protocol as described in an earlier report, with 0.23% collagenase treatment in DMEM at 37°C. 1 Finally, 1 × 105 cells/well were plated to form a monolayer of ECs in a 24-well plate in DMEM containing 20% FBS, gentamycin (20 mg/mL), streptomycin (100 mg/mL), and penicillin (100 IU), and were incubated in a CO2 incubator at 37°C and 70%–80% humidity. The media were replaced every 2 days. The cells were immunostained with the EC marker CD31. 1 Furthermore, ECs (1 × 104 cells/well) were treated with melatonin in the presence or absence of 25% DLA-containing medium for 72 h, and the proliferation of cells was analyzed using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay with slight modification. Four hours before termination, 20 µL of MTT solution (5 mg/mL in PBS) was added into each well. For termination, cells were collected in microfuge tubes, centrifuged at 3000 g for 5 min, and the medium was removed, washed with PBS, and 100 µL of dimethyl sulfoxide was added to the pellet to dissolve the formazan crystal formed in the cells. Finally, optical density was measured at 570 nm by using an enzyme-linked immunosorbent assay (ELISA) plate reader. 1
ROS in ECs
DHE, a superoxide indicator dye, was used to determine ROS levels in the DLA-induced ECs. The ECs were treated with DLA and melatonin (1 mM) and were incubated for 72 h in a CO2 incubator at 37°C and 70%–80% humidity. The cells were fixed with chilled methanol and washed with PBS; thereafter, the cells were stained with DHE (1 µg/mL) for 15 min and observed under a fluorescent microscope (Nikon). 22
Quantification of MMP-9 by ELISA
ECs were harvested from the mouse aorta and treated with DLA and melatonin (1 mM) for 72 h. The cells were lysed with radioimmunoprecipitation assay (RIPA) buffer, and protein concentration was estimated with Bradford assay reagents. ELISA was performed as directed by the Ray Biotech kit manual with an equal amount of protein. A standard graph was drawn with the MMP-9 protein as provided in the kit, where the x-axis is the concentration of MMP-9 in pg/mL and the y-axis is the net optical density. Finally, MMP-9 concentration was quantified with reference to the standard curve.
Gelatin zymography
ECs were harvested from the mouse aorta and treated with DLA and melatonin (1 mM) for 72 h. Furthermore, cells were lysed with RIPA buffer. The protein concentration was evaluated with Bradford assay reagents for the melatonin-treated EC samples. MMP-9 protein was taken as a positive control and marked as M in the zymogram. MMP activity was measured by gelatin zymography as described by Kupai. Briefly, an equal amount of protein was separated in 10% polyacrylamide gels with 0.1% gelatin. After electrophoresis, the gels were first soaked in renaturing buffer (2.5% Triton X-100) and then in zymogram developing buffer (50 mM Tris base, pH 7.5, containing 200 mM NaCl, 5 mM CaCl2 · 2H2O). They were then incubated for 20 h at 37°C and were finally stained with 0.025% Coomassie Brilliant blue. Zymogram developing buffer contains divalent metal cations, which are required for the enzymatic activation of both the pro- and active enzymes. The areas of proteinase activity were visualized as clear bands by washing with the distaining solution. MMP-9 protein from Ray Biotech was taken as a marker.23,24
qPCR analysis of histone deacetylases (HDAC1, HDAC2, HDAC6, and HDAC9) and DNMT (DNMT1, DNMT3A and DNMT3B) mRNA expressions in DL cells
The effect of melatonin on the expression of histone deacetylases (HDACs) and DNA methyltransferase (DNMT) genes in the DL cells was measured by RNA isolation. DL cells treated with melatonin or control groups were obtained by the abovementioned experiment, followed by first-strand cDNA synthesis and then gene-specific PCR done by using the forward and reverse primers enlisted in Table 1 to obtain PCR amplicons and fold change of each transcript, calculated by the comparative ΔΔCT method and normalized with β-actin.
Free Binding Energy and Drug Interaction with Targeted Protein Residues with Melatonin
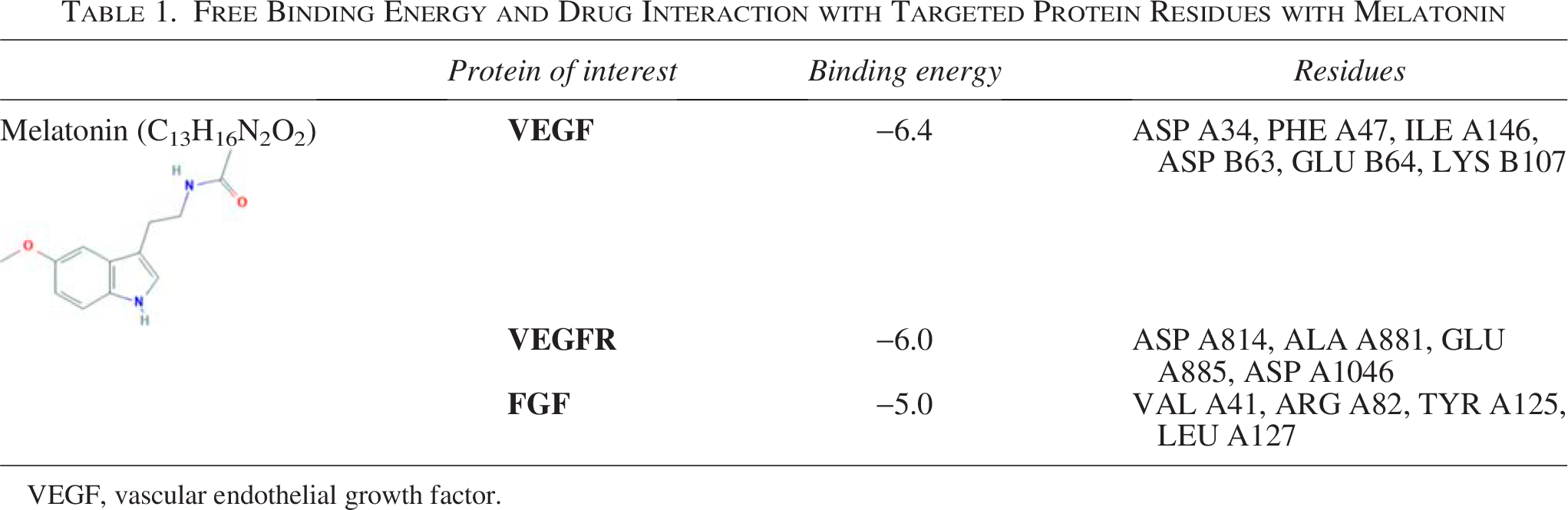
VEGF, vascular endothelial growth factor.
Molecular docking study
Database and software: PubMed Database (https://pubmed.ncbi.nlm.nih.gov/), PubChem (https://pubchem.ncbi.nlm.nih.gov/), RCSB-Protein Data Bank (PDB) (www.rcsb.org/), AutoDock Vina v1.2.× (2021–present): https://github.com/ccsb-scripps/AutoDock-Vina, interaction visualized with Discovery studio visualizer, and PyMOL used for molecular docking.
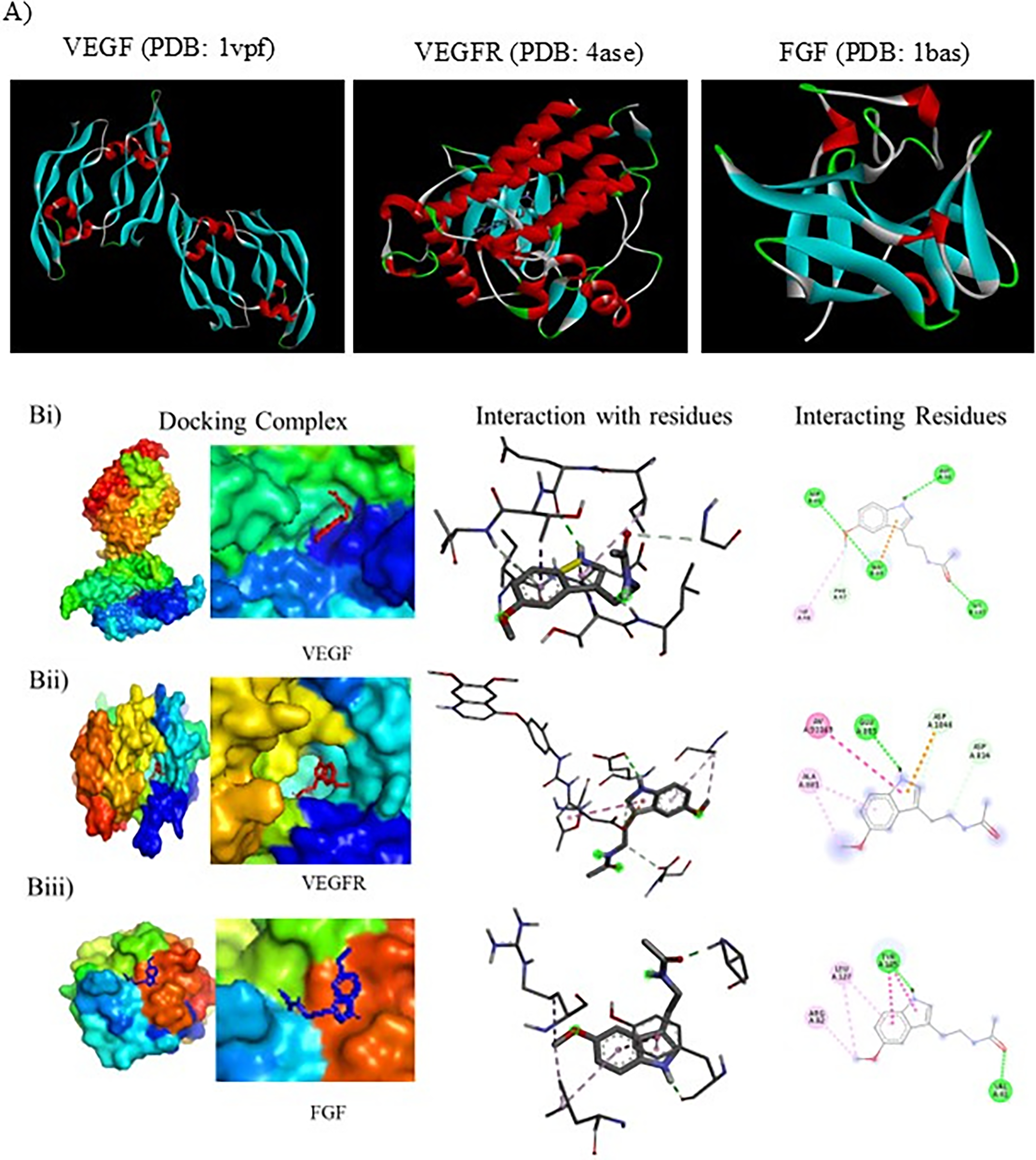
Preparation of protein and melatonin: The crystal structures of VEGF (PDB: 1vpf), VEGFR (PDB: 4ase), and FGF (PDB: 1bas) were retrieved from the RCSB PDB site (www.rcsb.org). The VEGF structure contains a total of 98 amino acids with 2.50 Å resolution. The VEGF structure contains a total of 98 amino acids with 2.50 Å resolution. VEGFR structure contains 353 amino acids with 1.83 Å resolution, and FGF structure contains 154 amino acids at 1.90 Å resolution. The structure of melatonin was obtained from PubChem. Docking coordinates were obtained by removing water molecules, bound receptors, inhibitors, and undesired heteroatoms using Discovery Studio 2017R2. The polar hydrogens were added, and the Kollman charges were assigned to stabilize the charges. The PDB file of structures was converted into a PDBQT file format using Pyro.
Docking protocol using AutoDock Vina: The previously reported binding pockets in the proteins were used to generate a grid box to encapsulate the active site. The auto grid program was used to precalculate the grid maps of interaction energies between the protein and various atom types present in the inhibitors. Docking calculations were performed with default parameters. The binding energy was evaluated using the following scoring function. Representative docked conformation of various constituents of VEGF, VEGFR, and FGF with melatonin. The interaction of three-dimensionally oriented molecules with the active site of melatonin is shown (left); the interacting residues in the active site and the localization of the inhibitors are shown (middle); and the two-dimensional interactions with various residues in the active site and the nature of interactions are depicted (right).
Statistical analysis
Data shown are representative images, and the difference among groups of cells was analyzed using Student’s t-test and analysis of variance. A p-value of 0.05 was considered statistically significant. The p-value ≤ 0.05 is marked as *, p-value ≤ 0.01 is marked as **, and p-value ≤ 0.001 is marked as ***. All experiments were performed in triplicate and repeated three times unless otherwise stated.
Results
Growth inhibitory effect of melatonin on ECs
First, the authors checked the effect of melatonin on DLA-induced EC proliferation. The DLA-treated ECs showed immense proliferation compared with the control group. The pharmacological dose of 1 mM showed significant inhibition of DLA-induced EC growth with respect to the DLA control group. Whereas there was no significant change in the rate of proliferation in control ECs with melatonin treatment (Fig. 1).

Growth inhibitory effect of melatonin on endothelial cells (ECs), where
Melatonin reduces DLA-induced ROS in ECs
To determine ROS production in the DLA-induced ECs, the superoxide indicator DHE dye was used. The ECs were treated with DLA and melatonin (1 mM). First fixed with chilled methanol, after washing with PBS, the cells were stained with DHE (1 µg/mL) for 15 min and observed under the fluorescent microscope (Nikon). The ROS levels in the DLA-treated ECs were high with respect to the control ECs. On the contrary, melatonin treatment of DLA-induced ECs shows a reduction in ROS levels with respect to the DLA-induced ECs (Fig. 2).
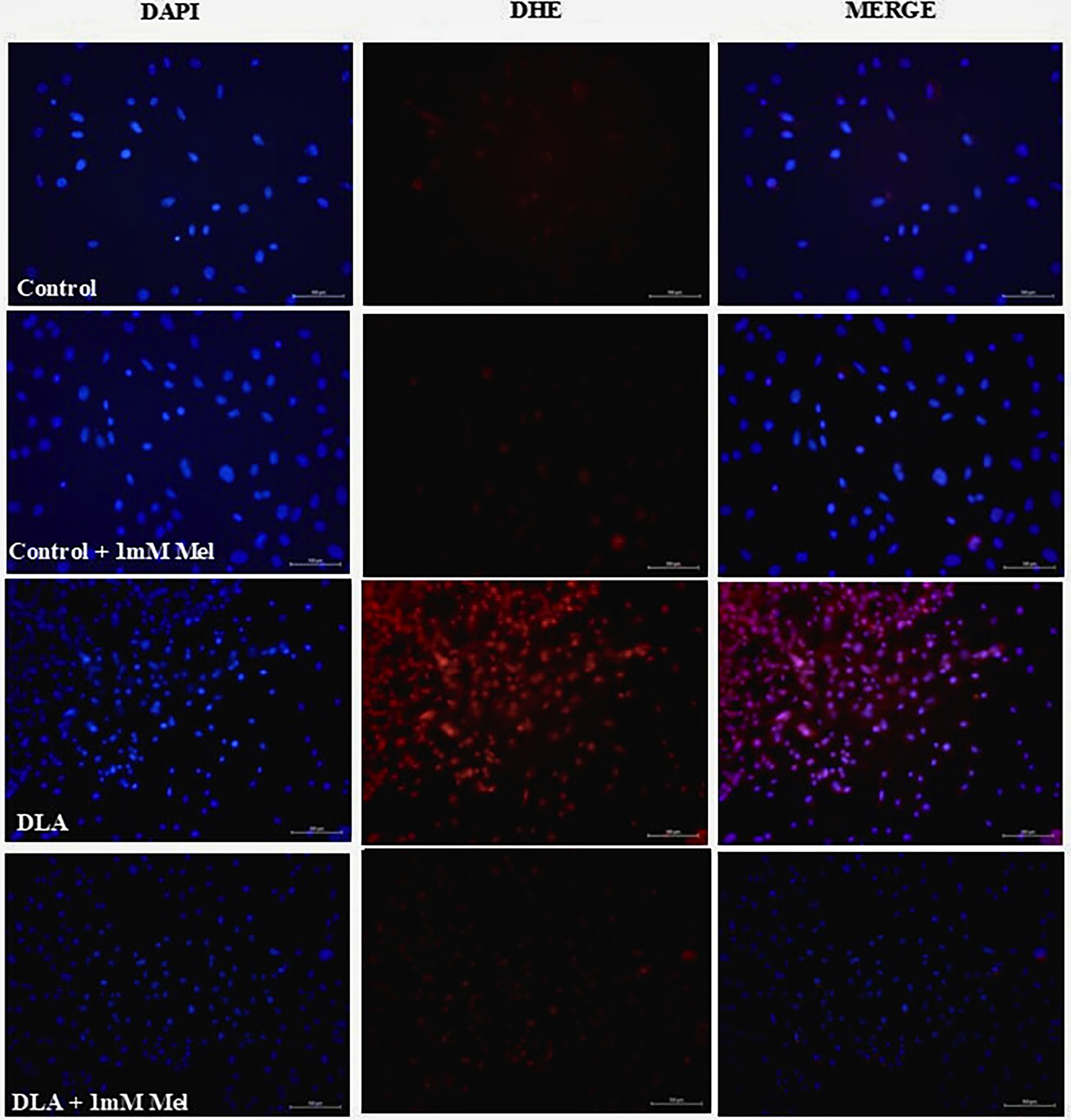
Reduction of Dalton lymphoma (DL)-induced reactive oxygen species (ROS) formation in endothelial cells. Representative images from control, DL ascites (DLA), and melatonin treated (1 mM) at 20× magnifications.
Melatonin reduces MMP-9 in DLA-ECs
ECs were harvested from the mice aorta and treated with DLA and melatonin (1 mM) for 72 h. The cells were lysed with RIPA buffer, and an ELISA was performed as directed by the kit manual. A standard graph was made with the MMP-9 protein as provided in the kit, where the x-axis is the concentration of MMP-9 in pg/mL and the y-axis is the net optical density. Finally, MMP-9 concentration was quantified with reference to the standard. Melatonin treatment showed a significant reduction in MMP-9 concentration (Fig. 3A).

Melatonin reduces matrix metalloproteinases (MMPs) in endothelial cells (ECs).
Melatonin treatment decreases MMP activity in DLA-activated ECs
ECs were harvested from mice aorta and treated with DLA and melatonin (1 mM) for 72 h. Furthermore, cells were lysed with RIPA buffer, and equal protein was loaded on an acrylamide gel as discussed earlier. Melatonin treatment significantly reduces the MMP activity. The MMP-9 activity was increased by 1.47-fold in the DLA-treated group, and a significant decrease of 1.87-fold was observed in the melatonin-treated ECs. Whereas 1.67-fold increased MMP-2 activity was reduced by 1.88 times upon melatonin treatment (Fig. 3B).
Effect of melatonin on HDAC (HDAC1, HDAC2, HDAC6, and HDAC9) and DNMT (DNMT1, DNMT3A, and DNMT3B) mRNA expressions in DL cells
The core histones undergo a wide range of posttranslational modifications, including acetylation, controlled by histone acetyltransferases and associated with gene expression, whereas deacetylation is controlled by HDACs and associated with gene inactivation. Similarly, DNMTs balance the promoter and global methylation of DNA, causing gene expression or suppression. In this experiment, HDAC1 was 2.1-fold downregulated in DL cells, which were upregulated with melatonin treatment up to 1.5-fold (Fig. 4Ai). Whereas HDAC2 was upregulated in DL cells with respect to the normal thymus. Melatonin treatment downregulates HDAC2 expression by fourfold (Fig. 4Aii). HDAC6 was downregulated 3.18-fold in DL cells compared with the normal thymus. Melatonin treatment upregulates 0.589-fold (Fig. 4Aiii). HDAC9 was downregulated 2.785-fold, which was upregulated to 2.6-fold with melatonin treatment (Fig. 4Aiv). The DNMT1 was 5.8-fold increased in DL cells, which were reduced to 1.2-fold (Fig. 4Bi). DNMT3a is 3.2-fold higher in DL cells, which were reduced 2.6-fold with melatonin treatment (Fig. 4Bii). DNMT3b is also elevated by 3.8-fold, which was reduced to 1.46-fold (Fig. 4Biii).
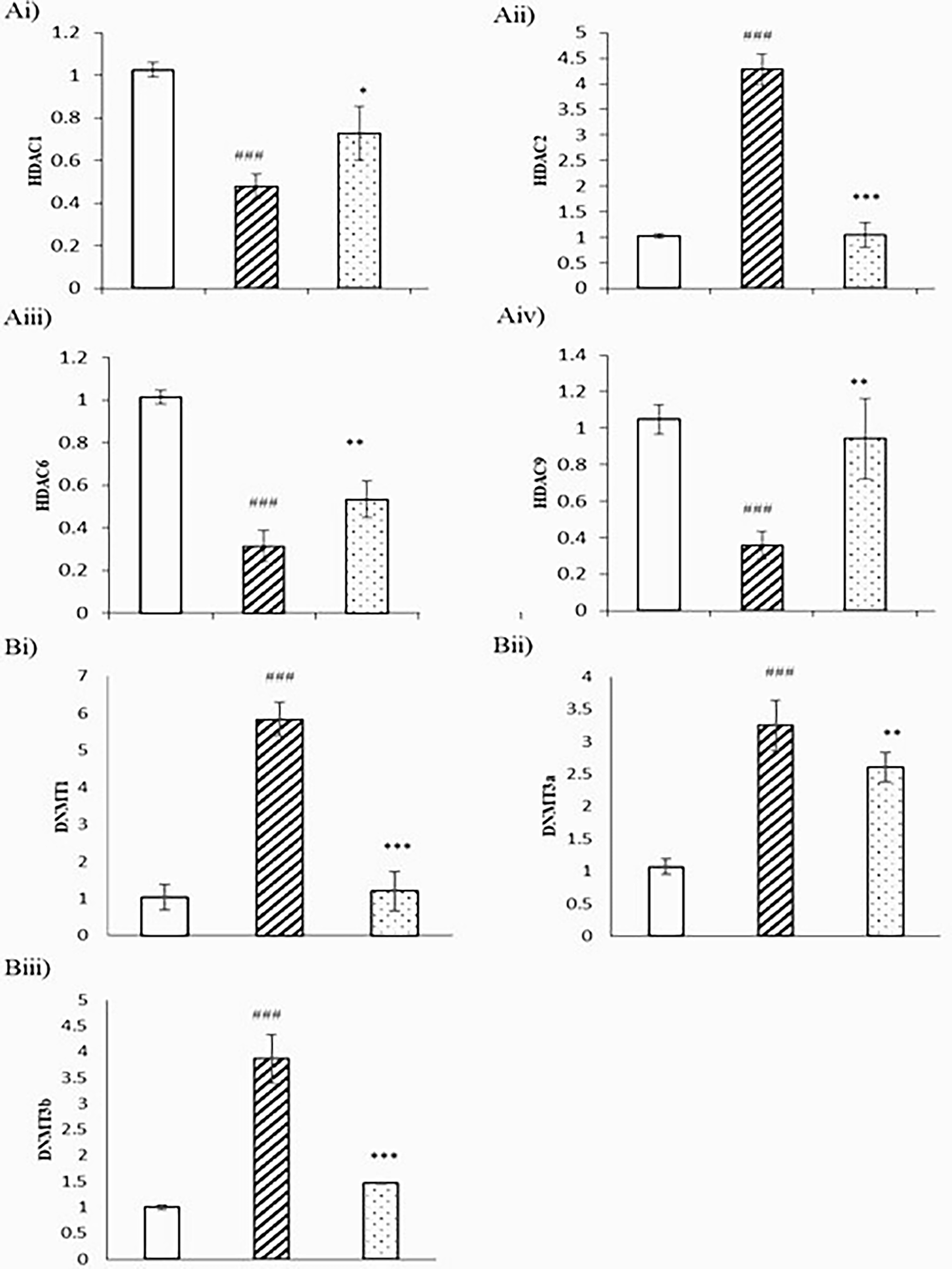
Effect of melatonin on the expression of HDACs and DNMTs in Dalton lymphoma (DL) cells. qPCR analysis of DL cells (2 × 106 cells/mL) grown with or without melatonin (24 h) for mRNA expression of
Effect of melatonin on promoter methylation of VEGF, CDH1, and TIMP3
The DL cells showed high promoter methylation with respect to the normal thymus. Melatonin treatment promotes an increase in unmethylated PCR amplification. This indicates a reversal of the trend with respect to the DL cells. The TIMP3 was highly suppressed in DL cells, as observed in the previous results. One of the main reasons is promoter methylation, which was very much clarified in the MS-PCR with respect to the normal thymus DNA. With melatonin treatment, there is no improvement in amplification of the unmethylated PCR product. This directly correlates with the ineffectiveness of melatonin on promoter unmethylation. The DL cells show an increase in the unmethylation PCR product with respect to the control group. This was reduced with melatonin treatment (Fig. 5).
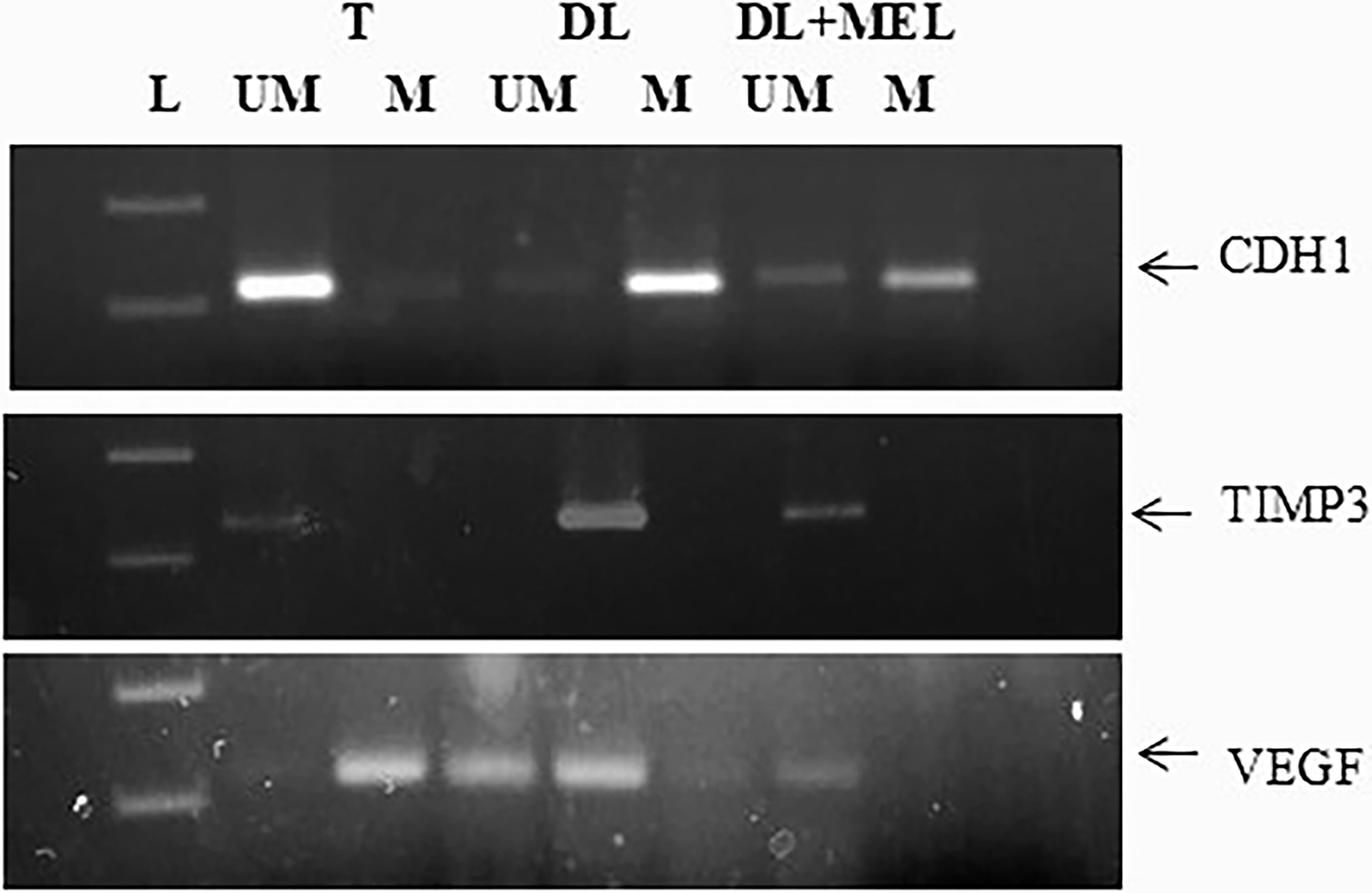
Gel electrophoresis, 2%, showing expression of promoter methylation status of CDH1, TIMP3, and VEGF genes in DL cells treated with 1 mM melatonin for 6 d. Where T: thymus, DL: Dalton lymphoma cells, DL + Mel: the DL cells treated with 1 mM melatonin for continuous 6 d, um: unmethylated PCR product, m: methylated PCR product.
Docking simulations
Many reports in the literature suggest melatonin alone or in combination with chemotherapy as a possible candidate for the treatment of cancer. The multipronged therapeutic effect of melatonin has paved the way for it as a potent regulator. In the previous report, the authors found that the regulatory impact of melatonin on DL-induced angiogenesis, where melatonin acts at the transcription as well as the translation levels of pro- and antiangiogenic genes. Herein, the authors have chosen VEGF, a proangiogenic protein; VEGFR, the receptor for VEGF; and FGF, another proangiogenic gene, to interact with melatonin. With these targets, they have carried out the molecular docking studies with melatonin. The results (Table 1) indicate that melatonin exhibits strong interaction with ASP A34, PHE A47, ILE A46, ASP B63, GLU B64, and LYS B107. The kinase domain receptor (KDR) binding sites’ five side chains most important for binding were Ile-46 from strand b2, Ile-83 from strand b5, Glu-64 from loop b3-b4, Phe-17 from helix a1, and Gln-79 from strand b5. 25 VEGFR ASP A814, ALA A881, GLU A885, ASP A1046: similar protein binding interaction with axitinib (VEGFR type IVa TKI), GLU 885 and ASP1046, makes H-bonds to the key pivot point NH backbone of D1046 and a second direct H-bond to the carboxylate side chain of E885 26 (Fig. 6).
Discussion
Angiogenesis is the main reason behind the complexity in the treatment and survival of the patient. In 1971, Folkman proposed that tumor growth, survival, and metastasis are angiogenesis-dependent. Angiogenesis provides nutrition, oxygen, and the removal of waste. Later, blocking angiogenesis became a strategy to arrest tumor growth and metastasis. The strategy of using antiangiogenic compounds targeting nontransformed tumor microenvironment cells, such as ECs, is a modern approach. These cells are genetically stable and less prone to drug resistance. So, the nontoxic agents are very effective in targeting the pathological angiogenesis by regulating the tumor microenvironment. EC proliferation is a very essential step in new blood vessels. A tube-like assembly of ECs was stabilized with pericytes and smooth muscle cells; furthermore, fibroblasts strengthen the vessels. The authors were interested in checking the effect of melatonin on EC proliferation. They found that there is immense EC proliferation in the presence of tumor milieu DLA. This was decreased to a great extent with a pharmacological dose of melatonin treatment. On the contrary, melatonin does not have any toxic effect on normal cells (Fig. 1).
ROS/RNS are important intracellular signaling molecules that play key roles in various physiological processes, including apoptosis. Increased levels of reactive species are associated with oncogenic stimulation, and oxidative stress can be considered an important class of carcinogens. Cancer development is characterized by the cumulative action of multiple events in a single cell with initiation, promotion, and progression stages; ROS are involved in all stages.18,27 The initiation stage involves a nonlethal mutation in DNA. Both ROS and RNS are involved in DNA damage. The DNA mutations caused by reactive species include point mutations, deletions, insertions, chromosomal translocations, cross-links, and other modifications. An early study demonstrated that DNA alterations by oxidative stress through 8-hydroxyguanine (8-OH-G) mutations, which may arise from the formation of 8-OH-dG, involve the GC → TA transversion. This type of modified DNA is relatively easily formed, is mutagenic and carcinogenic, and can be used as a potential biomarker of carcinogenesis. Direct DNA damage or genomic instability, coupled with altered gene expression and changes in protein conformation, occurs simultaneously in cancer development.27,28 The promotion stage is characterized by clonal expansion of initiated cells by induction of cell proliferation and failure to induce cell death. Oxidative stress is present continuously at a relatively low level, and cell division and subsequent tumor growth are stimulated. Progression is an irreversible stage of the carcinogenic process. Further genetic damage and disruption of chromosome integrity occur at this stage, corresponding to a cell transition from benign to malignant. 29 In this experiment (Fig. 2), the authors also found increased ROS levels in DLA-induced ECs compared with the normal EC group. Melatonin treatment induces mild ROS production in normal cells with respect to the control group. Whereas melatonin treatment causes a reduction in ROS levels in DLA-induced ECs compared with the DLA-induced EC control group.
A family of 23 Zn2+ dependent nonproteolytic activities of MMPs plays a very essential function in tissue remodeling, including various components of the extracellular matrix. These are one of the major regulators of angiogenesis by EC proliferation; migration leads to cancer invasion and metastasis. Several studies suggest the role of MMP-2 and MMP-9 in facilitating cell movement by basement membrane degradation of collagen type IV. MMP-2 is secreted in the form of pro-MMP-2 and activated by an MT1MMP-dependent mechanism by TIMP2. 30 MMP-9 (92 and 240 kDa), or gelatinase B, are the basement membrane collagens IV-degrading MMPs. MMP-2 and MMP-9 are expressed in leukocytes, fibroblasts, cardiomyocytes, neutrophils, macrophages, vascular smooth muscle cells, and ECs and are involved in the degradation of structural proteins. The authors observed high amounts of MMP-9 in the DLA-induced ECs, which is very significantly decreased after melatonin treatment (Fig. 3A). Both MMP-9 and MMP-2 are gelatinases, so both have a common substrate, gelatin. With the gelatin zymography assay, the authors can check the activity of both the enzymes simultaneously. Zymography showed that MMP-9 and MMP-2 activity was high in DLA-induced ECs with respect to the normal ECs. The activity of both proteins was significantly decreased in the presence of melatonin treatment (Fig. 3B). The activity of MMPs is dependent on tissue inhibitor of matrix metalloproteinases (TIMPs). TIMP1, TIMP2, TIMP3, and TIMP4, each one, inhibit all the members of MMPs. Of the TIMPs, TIMP3 is a multicellular protein integrated into the extracellular matrix tissue by charge exchange with sulfated glycosaminoglycans. Knockout of TIMP3 results in lung emphysema-like alveolar impairment and apoptosis in mammary epithelial cells. 31 These evidences propose that TIMP3 is a key regulator of MMP activities. It also regulates innate immunity by controlling the mechanism of tumor necrosis factor-α by ADAM. TIMP3 was found to be downregulated in cancer, which is due to gene methylation. The authors checked the promoter methylation status of TIMP3 in DL cells by using the MS-PCR technique with genomic DNA. The results showed that the promoter region of TIMP3 is methylated, while the DNA isolated from normal thymic cells showed the amplification of unmethylated specific primers. This showed that normal thymus cells have no methylation at the promoter region. The DL cells showed that methylation at the promoter region leads to silencing of the gene (Fig. 5). The DNA having high CpG repeats is heavily methylated and responsible for the maintenance of chromosomal stability. CpG islands occupied ∼60% of human gene promoters. 32 Increased methylation at the promoter region causes gene silencing, and hypomethylation of global DNA causes genomic instability; both conditions cause a high risk of cancer. 33 Global hypomethylation causes instability of the genome and promotes the expression of quiescent proto-oncogenes and prometastatic genes, resulting in tumor progression. Not only is CpG island promoter methylation required for normal silencing of genes, but also the methylation of non-CpG promoters is important; for example, non-CpG island OCT-4 promoter methylation is essential for its expression. Differential methylation at promoters is involved in the regulation of gene expression, and altered promoter methylation levels are molecular signatures for conditions ranging from cancers to psychiatric disorders. 34 Next to TIMP3, the authors also check the methylation status of cadherin 1 (CDH1). CDH1 is an endogenous antiangiogenic gene; it also showed methylation at the promoter region compared with normal thymic tissue. Melatonin treatment leads to reversing the condition, causing a decrease in methylation at the promoter region (Fig. 5). Keqin Hua et al. also showed the upregulation of TIMP3, TSP1, and CDH1 during dormancy of breast cancer and downregulation in recurrence, where TIMP3 and CDH1 are epigenetically regulated. According to M.X. Da, the overexpression of VEGF-C leads to poor prognosis in gastric cancer, and the treatment of S-adenosylmethionine, a methyl group donor, causes global methylation, further leading to the downregulation of VEGF. The authors are interested in knowing the promoter methylation status in DL cells. Interestingly, DL cells showed the expression of unmethylated-specific amplification in comparison with methylation in DL cells. Normal thymic cells showed high methylation compared with nonmethylation (Figs. 4B and 5). Histone proteins, which comprise the nucleosome core, aid in the packaging of DNA and contain a globular C-terminal domain and an unstructured N-terminal tail. Histone modifications work by either changing the accessibility of chromatin or by recruiting and/or occluding nonhistone effector proteins, which decode the message encoded by the modification patterns. Histone deacetylation is catalyzed by HDACs to remove the acetyl groups. 35 Histone acetylation leads to an open chromatin structure and enables transcription factors to bind to DNA, whereas deacetylation leads to chromatin condensation and transcriptional repression. HDACs play a very important role in maintaining the homeostasis of histone acetylation in euchromatin and heterochromatin. 36 In DL cells, the authors observed the downregulation of HDAC1, HDAC6, and HDAC9, whereas HDAC2 is upregulated compared with the normal thymus (Fig. 4A). Melatonin maintains homeostasis by regulating the expression toward normal. HDAC1 regulates various DNA damage responses, such as by inducing P-53 acetylation and by inhibiting P21 and MDM2 gene expression. HDAC6 interferes with HSP90 by degrading HSP90-interacting proteins. 36 High expression of HDAC2 is correlated with colorectal carcinoma. HDAC9 showed variation in expression in different tumor types.
VEGF is a homodimeric member of the cystine knot family of growth factors. VEGF has a helical N-terminal, two identical symmetrical KDRs present symmetrically at each pole, containing two functional hot spots. The most important five side chains for KDR binding are Ile-46, Ile-83, Glu-64 from the loop, Phe-17 from the helix, and Gln-79, which account for 70% of free binding energy. Ile-43, Lys-84, and Pro85 are responsible for 20% of the binding energy. Apart from the KDR site, another hot spot region consisting of Ile-46 and Ile-83 is highly important, whereas Glu-64 is moderately important, and Ile-43, Lys-84, and Pro85 have less impact. The smaller hotspot includes Gln-79 and Phe-17 at the N-terminal helix. Melatonin showed −6.4 binding energy and interacts with ASP A34, PHE A47, ILE A146, ASP B63, GLU B64, and LYS B107. Sunitinib, an inhibitor of VEGFR, by making a direct H-bond of D1046 and E885 at the JM domain near the catalytic domain, results in the confirmation change. Melatonin showed −6.0 binding energy with ASP A814, ALA A881, GLU A885, and ASP A1046. FGF showed −5.0 binding energy at the D1 domain with VAL A41, ARG A82, TYR A125, and LEU A127 (Fig. 6).
Conclusion
The present study may conclude that melatonin is a potent antiangiogenic compound. It targets tumors by targeting angiogenesis by various means, including regulation of ROS production, by regulating MMP secretion and controlling their activity. Melatonin directly acts on DL cells and reverses epigenetic modifications in endogenous inhibitors TIMP3 and CDH1, resulting in an increase in inhibition of angiogenic growth.
Footnotes
Disclosure Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding Information
RC/2015/9677/D-1813, Dean(R)/R&D/2014 R&D, Delhi University.