
Abstract
Mounting preclinical evidence in rodents and nonhuman primates has demonstrated that prolonged exposure of developing animals to general anesthetics can induce widespread neuronal cell death followed by long-term memory and learning disabilities. In vitro experimental evidence from cultured neonatal animal neurons confirmed the in vivo findings. However, there is no direct clinical evidence of the detrimental effects of anesthetics in human fetuses, infants, or children. Development of an in vitro neurogenesis system using human stem cells has opened up avenues of research for advancing our understanding of human brain development and the issues relevant to anesthetic-induced developmental toxicity in human neuronal lineages. Recent studies from our group, as well as other groups, showed that isoflurane influences human neural stem cell proliferation and neurogenesis, whereas ketamine induces neuroapoptosis. Application of this high throughput in vitro stem cell neurogenesis approach is a major stride toward ensuring the safety of anesthetic agents in young children. This in vitro human model allows us to (1) screen the toxic effects of various anesthetics under controlled conditions during intense neuronal growth, (2) find the trigger for the anesthetic-induced catastrophic chain of toxic events, and (3) develop prevention strategies to avoid this toxic effect. In this article, we reviewed the current findings in anesthetic-induced neurotoxicity studies, specifically focusing on the in vitro human stem cell model.
Introduction
A large body of experimental work has demonstrated that exposure of developing animals to most general anesthetics used clinically today, either volatile or intravenous, can induce widespread neuronal cell death followed by long-term memory and learning abnormalities.1 -9 Anesthetic-induced neurotoxicity may depend on the following variables: (1) anesthetic dose, exposure duration, and number of exposures;10,11 (2) the receptor type being activated or inactivated;1,12 (3) single anesthetic or combination of different anesthetic agents; 1 and (4) the stage of brain development.12 -14 The window of the greatest vulnerability of the developing brain to anesthetics is restricted to the period of rapid synaptogenesis or the so-called brain growth spurt. This vulnerable period for anesthesia-induced neuroapoptosis appears to be very brief in animals, occurring in rodents primarily during the first 2 weeks after birth. For rhesus monkeys, this period ranges from approximately 115-day gestation up to postnatal day 60. In humans, it starts from the third trimester of pregnancy and continues 2 to 3 years following birth.12,15 -17 The underlying mechanisms of increased neurotoxicity following anesthetic exposure are not well understood. In addition, it is entirely unclear if anesthesia-induced cognitive impairment occurs in humans. Recent studies from our group and others showed that an in vitro human stem cell model can be used to study the effects of anesthetic agents on developing human neurons to determine whether or not anesthetic agents can induce toxicity in humans. In this article, we reviewed the current findings obtained from intact animal and in vitro primary neuron culture models and the underlying mechanisms. We specially focused on the discussion of studies utilizing the human stem cell model.
In Vivo Animal Model
Anesthetic-induced neurotoxicity appears to only affect young animals. For example, isoflurane-induced neurodegeneration was only observed in young rats but not in adult rats.13,14 Similar observations were also reported in mice and monkeys. Ketamine administered subcutaneously to 7-day-old mice for 5 hours resulted in a significant increase in neuronal cell death. 18 Intravenous administration of ketamine for 24 hours caused an increase in cell death in the cortex of rhesus monkeys at 122 days of gestation and postnatal day 5. 19 In a recent study, both fetal and neonatal monkeys were exposed to ketamine for 5 hours. Ketamine caused a less widespread pattern and less dense concentration of neuroapoptosis in neonatal brains than in fetal brains. 12 Additionally, sevoflurane anesthesia in pregnant mice (gestational day 14) also induced neurotoxicity in fetal and neonatal mice. 4 Table 1 is a summary of some of the key animal studies to date examining anesthetic-induced neurotoxicity.
Representative In Vivo Animal Studies Regarding the Effect of General Anesthetic–Induced Developmental Neurotoxicity.

Abbreviations: GABAAR, gamma-aminobutyric acid type A receptor; MAC, minimum alveolar concentration; PD, postnatal day; NMDAR, N-methyl-D-aspartate receptor.
In Vitro Primary Neuron Culture Model
In vitro experimental evidence from cultured neonatal animal neurons confirmed the in vivo findings.20 -22 Vutskits et al 20 showed that 1 hour of exposure to ketamine at a concentration of 40 µM or greater induced toxicity in a primary culture of GABAergic neurons from postnatal day 0 rats. Treatment of neurons derived from fetal rats (gestational days 18-19) with 100 µM of ketamine for 48 hours resulted in the loss of 45% of neurons by apoptosis. 21 Similar toxic effects were also found in rhesus monkeys at postnatal day 3 after 24 hours of 10 µM ketamine exposure. 22 The frontal cortex was the brain region that was found to be the most vulnerable to ketamine-induced neurotoxicity during development. 23 The in vitro findings have been summarized in Table 2.
Representative In Vitro Studies Regarding the Effect of General Anesthetic–Induced Developmental Neurotoxicity.
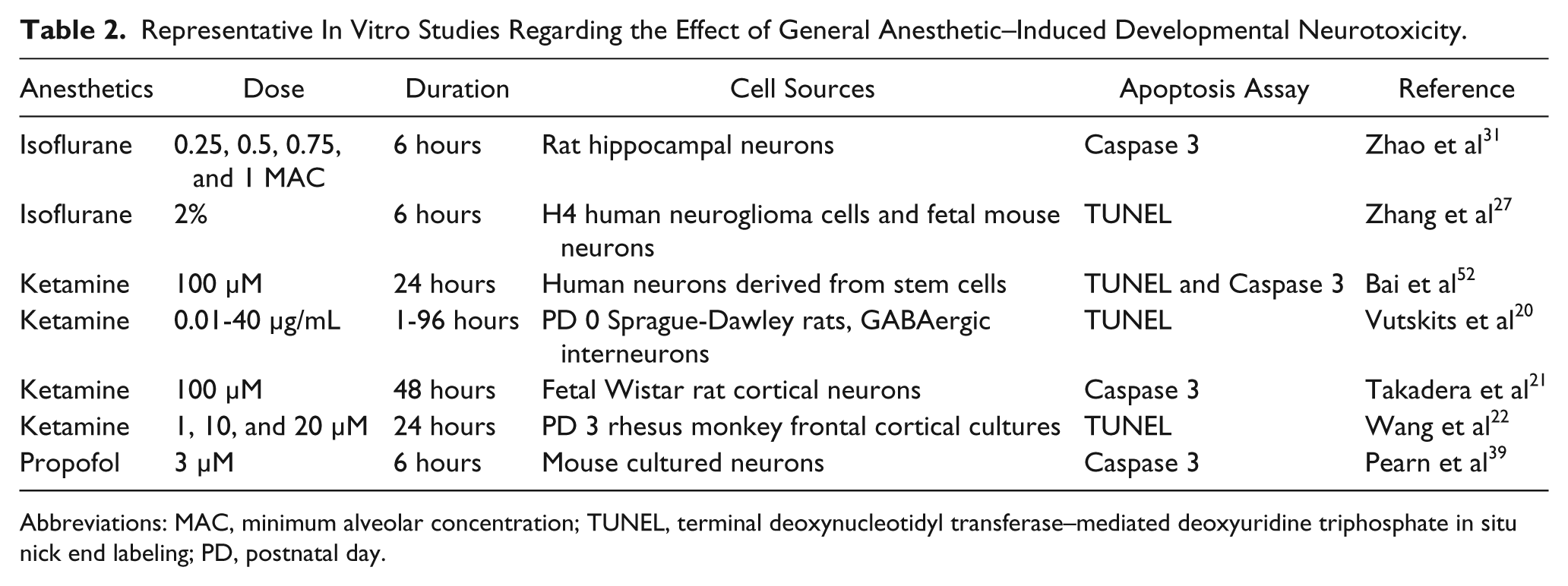
Abbreviations: MAC, minimum alveolar concentration; TUNEL, terminal deoxynucleotidyl transferase–mediated deoxyuridine triphosphate in situ nick end labeling; PD, postnatal day.
Mechanisms of Anesthetic-Induced Neurotoxicity
The underlying mechanisms of increased neurotoxicity following anesthetic exposure are complex and are not well understood. Although many different mechanisms and pathways have been implicated to play a role in anesthetic-induced neurotoxicity, the mitochondria appear to play key roles in this process through their crucial involvement in cellular processes and apoptosis. 24 However, many additional mechanisms have been proposed.
Trigger of Neurotoxicity
Apoptosis was shown to be involved in anesthetic-induced neuronal cell death.25 -27 The mechanistic details by which anesthetics induce neurotoxicity have yet to be established. The vast majority of general anesthetics are N-methyl-D-aspartate receptor (NMDAR) antagonists and/or gamma-aminobutyric acid type A receptor (GABAAR) agonists. The NMDAR is involved in a variety of physiological processes, including memory, learning, and neuronal development. In the immature brain, GABA is the first neurotransmitter to become functional in developing networks and does not function as an inhibitory neurotransmitter as in the adult.28 -30 GABAAR activation results in chloride efflux and depolarization of neurons and induces a cytosolic calcium increase. 31 Upregulation of NMDAR and/or overactivation of GABAAR may lead to excessive calcium entry, exceeding the buffering capacity of mitochondria. This can lead to a cascade of events, including membrane potential depolarization, reactive oxygen species (ROS) production, cytochrome c release into the cytosol, and caspase activation, ultimately resulting in neuroapoptosis.14,18,32,33 Ketamine is a noncompetitive NMDAR antagonist. Withdrawal of ketamine induces compensatory upregulation of NMDAR expression followed by a toxic influx of calcium into neurons, leading to elevated reactive ROS generation and neuronal cell death. Administration of the antisense of NR1 can attenuate the ketamine-induced neuronal death.19,22,23,34 -36 Isoflurane, a GABAAR agonist, was found to induce neurotoxicity in hippocampal neurons in culture via a GABAAR-mediated increase in intracellular calcium concentration.31,37 Some anesthetics might be less toxic, such as dexmedetomidine, because they do not affect the cellular receptors that are most commonly influenced by the majority of general anesthetics. 38
Neuroinflammation
In addition to a direct influence on NMDAR and GABAAR, it was reported that anesthetic-induced neuroinflammation also plays an important role in the developing mouse brain and cognitive impairment. Anesthesia with 3% sevoflurane for 2 hours daily for 3 days induced cognitive impairment and neuroinflammation (eg, increased interleukin-6 levels) in the developing mouse brain but not in adult mice. Anti-inflammatory treatment (ketorolac) attenuated the sevoflurane-induced cognitive impairment, implicating neuroinflammation as a key mediator of anesthetic-induced neurotoxicity. 11
Neurotrophins
Alterations in the levels of a variety of neurotrophins have been implicated to play a role in anesthetic-induced neurotoxicity in the developing rodent brain. Brain-derived neurotrophic factor (BDNF) is important to prosurvival signaling pathways. BDNF is stored as proneurotrophin (proBDNF) within synaptic vesicles and is proteolytically cleaved to mature BDNF (mBDNF) in the synaptic cleft by plasmin, a protease activated by tissue plasminogen activator. mBDNF triggers prosurvival signaling while proBDNF binds to the p75 neurotrophin receptor (p75NTR) and activates RhoA that regulates actin cytoskeleton polymerization resulting in apoptosis. It has been shown that exposure of immature mouse neurons to propofol can induce neuroapoptosis through a proBDNF/p75NTR pathway. Knockout of p75NTR or administration of TAT-Pep5 (a specific p75NTR inhibitor) attenuated propofol-induced neuroapoptosis. 39 Additionally, it was observed that exposure of rat pups to propofol induced a significant decrease in the thalamus at the level of nerve growth factor, a protein that is critical in the survival and growth of neurons. Propofol exposure was also shown to alter the expression levels of a variety of key neurotrophic factor receptors and downstream targets such as AKT and Erk. 40 Taken together, these data suggest that altered neurotrophic factor expression following anesthetic exposure may also play a role in anesthetic-induced neurotoxicity.
Calcium Signaling
Intracellular calcium levels are tightly regulated under normal conditions as a result of key roles that calcium balance plays in various physiological processes. Persistent increases in intracellular calcium, beyond normal levels, can induce apoptosis. 41 It has recently been shown that intracellular calcium balance is disrupted in neurons following anesthetic exposure, implicating an important role for calcium disruption in anesthetic-induced neurotoxicity. 33 Ketamine has been shown to suppress intracellular calcium oscillations and decrease the levels of Ca2+/calmodulin-dependent protein kinase II, a key protein in calcium regulation in a hippocampal cell culture model. 42 It is well established that NMDA receptors play important roles in the regulation of intracellular calcium levels. Therefore, blockade of these receptors by anesthetics such as ketamine can lead to disrupted calcium levels, indicating that calcium dysregulation may be a probable mechanism of anesthetic-induced neurotoxicity.
Mitochondrial Pathway
Mitochondria are highly dynamic organelles that undergo continuous cycles of fusion and fission to maintain cellular homeostasis. The dynamics of mitochondrial fusion and fission result in changes in mitochondrial shape: either elongated, tubular, and interconnected mitochondrial networks or fragmented and discontinuous mitochondria. Unbalanced fission-fusion can affect a variety of biological processes such as apoptosis and mitosis, 43 leading to various pathological processes, including neurodegenerative disease.44 -47 Dynamin-related protein 1 (Drp1) is a key regulator of mitochondrial fission. Drp1 is primarily distributed in the cytoplasm of a healthy cell but shuttles between the cytoplasm and the mitochondrial surface. Phosphorylation of Drp1 at serine 616 stimulates mitochondrial fission, whereas phosphorylation at serine 637 inhibits mitochondrial fission. Drp1 activates, translocates, and oligomerizes from the cytoplasm to the mitochondrial surface at fission sites to induce mitochondrial fission. 48 Previous studies have shown that exposure of neonatal rat pups to general anesthetics induces significant increases in mitochondrial fission and leads to decreases in the level of Drp1 in the cytosol while increasing mitochondrial Drp1 levels. 49 Inhibition of mitochondrial fission was shown to prevent mitochondrial cytochrome c release and apoptotic cell death. 50 Loss of ΔΨm and release of cytochrome c from mitochondria are key events in initiating mitochondria-related apoptosis. 51 Ketamine-induced apoptosis in stem cell–derived human neurons was accompanied by a significant decrease in ΔΨm and an increase in cytochrome c release from mitochondria into the cytosol. In addition, most control neurons showed elongated and interconnected tubular mitochondria, whereas much shorter and smaller mitochondria were prevalent in the ketamine-treated culture (Figures 1A and 1B), 52 indicating the increased mitochondrial fission as possibly playing an important role in the toxic effects.
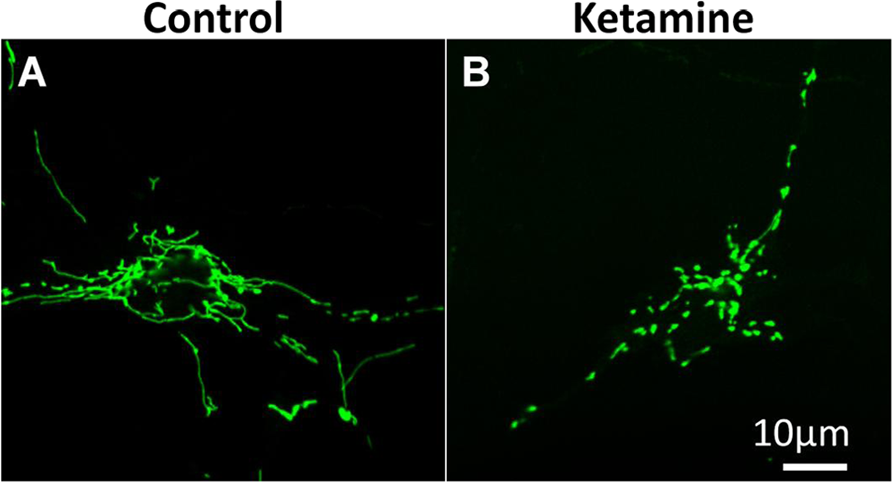
Ketamine increases mitochondrial fission: mitochondrial shape in the cells treated with or without 100 µM ketamine for 24 hours. Differentiated neurons were labeled with CellLight mitochondria–green fluorescent protein (GFP) reagent and expressed GFP in the mitochondria. Mitochondria were elongated and interconnected in the control cells (A), whereas mitochondria were short and disconnected in the ketamine-treated culture (B). 52
ROS Production
In response to ketamine, there was a significant increase in ROS production in the cytosol and superoxide generation within mitochondria in the ketamine-treated human embryonic stem cell (hESC)-derived neurons, indicating a mitochondrial origin of ROS. Trolox, a ROS scavenger, prevented ketamine-induced ROS production and apoptosis in the differentiated human neurons. 52 ROS are involved in the regulation of many physiological processes. However, overproduction of ROS under various cellular stresses can cause cell death. Mitochondria are a main source of ROS, and the primary ROS is superoxide, which is converted to H2O2 either by spontaneous dismutation or by the enzyme superoxide dismutase. H2O2 can be further transformed to OH·. 53 In agreement with our data, several recent animal studies from other groups suggested that accumulation of ROS was associated with anesthetic-induced mitochondrial damage.27,32 Application of the antioxidant 7-nitroindazole (nitric oxide synthase inhibitor) provided protection against ketamine-induced neuronal cell death. 54 These data suggest that the toxic effect can be prevented.
Controversy as to the Applicability of Animal Data to Clinical Anesthesia Practice
Although anesthesia neurotoxicity has been reported in several animal models,23,55 -58 considerable controversy remains as to whether these findings are relevant to humans because of the following issues. First, the experimental conditions under which the data are collected from animals are different from those experienced by human neonates undergoing surgery and general anesthesia. It is clearly very hard to separate the effects of anesthetics alone from other quite relevant variables, including the impact of surgery and other factors related to acute/chronic diseases.5,59 -61 Second, there may be interspecies differences in development and brain plasticity across mammalian species that allow humans to accommodate injury without clinically significant effects on neurodevelopment. For instance, the central components involved in apoptosis are a group of proteolytic enzymes called caspases, which can be activated by various types of stimulation. 62 There are species-specific (human vs mouse) differences in the way caspases process their targets. 63 Third, the neurodevelopmental impairments observed in animals, such as disruption of spontaneous behavior and poor performance in the Morris water maze, are of uncertain relevance to humans.13,58 Finally, experimental evidence provided thus far from in vitro cell culture and in vivo animal studies demonstrate that very high doses and prolonged exposure times are required to produce neurotoxic effects.22,31,36 A study from Wang’s group demonstrated that 6 injections of 20 mg/kg ketamine produced the most severe neuronal damage in neocortical areas, especially in layers II and III of the frontal cortex. However, no significant effects were observed in the animals injected either 1 or 3 times with 20 mg/kg ketamine. 22
Limitations of Clinical Evidence
It is well documented that exposure to general anesthetics during very active brain growth and formation of synapses in animals results in considerable death of brain neurons and subsequent learning disabilities.1,64 So far, there is no direct clinical evidence showing any such effect in fetuses, infants, or children at any dose. If and how general anesthetics induce human neural cell toxicity is unknown. Similar studies in humans are not feasible. There are many obvious reasons why studying the neurotoxic effects of various anesthetic agents in pediatric patients is not possible. It is impossible to thoroughly investigate the neurotoxic effects of anesthetics in fetuses, infants, and children. In addition, it is clearly very hard to separate the effects of anesthetics alone from other quite relevant variables, including the impact of surgery and the factors associated with diseases. 5 Moreover, it is not practical to either delay surgery in sick children or expose healthy children to anesthetic drugs in order to study these effects. It is also impossible to determine anesthetic-induced neurotoxicity using primary cultures of neonatal human neurons because of limited access to human tissue.
Several epidemiological studies in humans have implicated that children exposed to anesthesia in early life have a higher incidence of learning disabilities later in life.59,60,65,66 These and subsequent studies evaluating multiple anesthetic exposures (eg, 3 times) have shown detrimental effects on cognitive development in children (eg, before age 4) and significant delays in measured intelligence, adaptive functioning, and academic performance.61,67 -69 However, others found no significant differences. For instance, monozygotic twins discordant for having received anesthesia showed no significant difference in learning outcomes. 70 There were no differences in educational outcomes at 15 to 16 years of age between 2500 children with or without inguinal hernia repair. 71 The discrepancy in the results of the epidemiological studies highlights the need for a better model by which to study this phenomenon. There is a considerable ongoing effort to more fully understand the clinical significance of anesthetic neurotoxicity. For instance, the US Food and Drug Administration and the International Anesthesia Research Society have formed a unique public-private partnership called SmartTots (smarttots.org), and several retrospective human studies are under way.
Each year, up to 2% of pregnant women in North America undergo anesthesia during their pregnancy for surgery unrelated to delivery of the fetus. In addition, millions of human fetuses, infants, and children are exposed to anesthetic drugs every year in the United States and throughout the world. Thus, it is imperative to find a reliable mechanistic model to study whether or not clinically relevant doses of anesthetics induce developmental toxicity in human neurons. With the development of an in vitro neurogenesis system using hESCs, induced pluripotent stem cells (iPSCs), and neural stem cells (NSCs), investigators can (1) study mechanisms of brain development, (2) screen the toxic effects of various anesthetics under controlled conditions (dose, number of exposures, or developmental stage) on human neuronal lineages such as NSCs and neurons, (3) dissect the underlying molecular mechanisms of anesthetic-induced neurotoxicity, and (4) develop potential strategies for avoiding the toxic effect of anesthetics.
Pluripotent Stem Cells
Pluripotent Capacity
hESCs are pluripotent stem cells derived from the inner cell mass of human blastocysts. These cells are able to replicate indefinitely in an undifferentiated state and differentiate into virtually every cell type found in the adult body. 72 The differentiation ability of hESCs into committed stem cells or specific lineages is potentially valuable for: (1) tissue regeneration, (2) studying cellular and molecular events involved in early human development, (3) disease modeling, and (4) drug development and toxicity screening.34,73 -76
iPSCs are reprogrammed from somatic cells such as skin fibroblasts and blood cells by transferring pluripotency factors (eg, Oct-4, Klf4, Sox2, and cMyc). They are similar to hESCs in cell morphology, pluripotent marker expression (eg, Oct-4 and SSEA4), proliferation, and differentiation potential. Despite the significant advances in hESC biology, issues such as ethical controversies and rejection with hESCs limit their utility. Development of iPSC technology provides an alternative pluripotent cell source. Specifically, iPSCs can be generated from any patient, including those with heritable diseases and, therefore, carry the genotype of the patient they were derived from. Thus, development of iPSC technology offers the unique possibility of investigating the cellular consequences influenced by genetic versus environmental factors in a human model and the underlying mechanisms. For instance, iPSCs obtained from patients with long-QT syndrome recapitulate the long action potential phenotype features of inherited arrhythmias in the context of the patient’s genetic background in cellular culture.77,78
Neuronal Lineage Differentiation
Both hESCs and iPSCs could differentiate into any type of human cells, including NSCs by culturing them in a chemically defined medium. During the differentiation process hESCs were induced into NSCs and neurons following embryonic neuronal developmental principles.52,79,80 hESCs, NSCs, and neurons were characterized by cell morphology and cell-specific marker expression as follows. hESCs were grown as uniform flat colonies on mouse embryonic fibroblast feeder cells (Figure 2A). hESCs expressed pluripotent stem cell markers Oct-4 (Figure 2B) and SSEA-4. NSCs showed triangle-like morphology (Figure 2C) and expressed the NSC marker nestin (Figure 2D). They had strong proliferation potential and were passaged every 5 to 6 days. 52 In addition, they were able to differentiate into neuronal lineages: neurons, astrocytes, and oligodendrocytes. NSCs can also be directly isolated from fetal or adult nervous tissue. 81
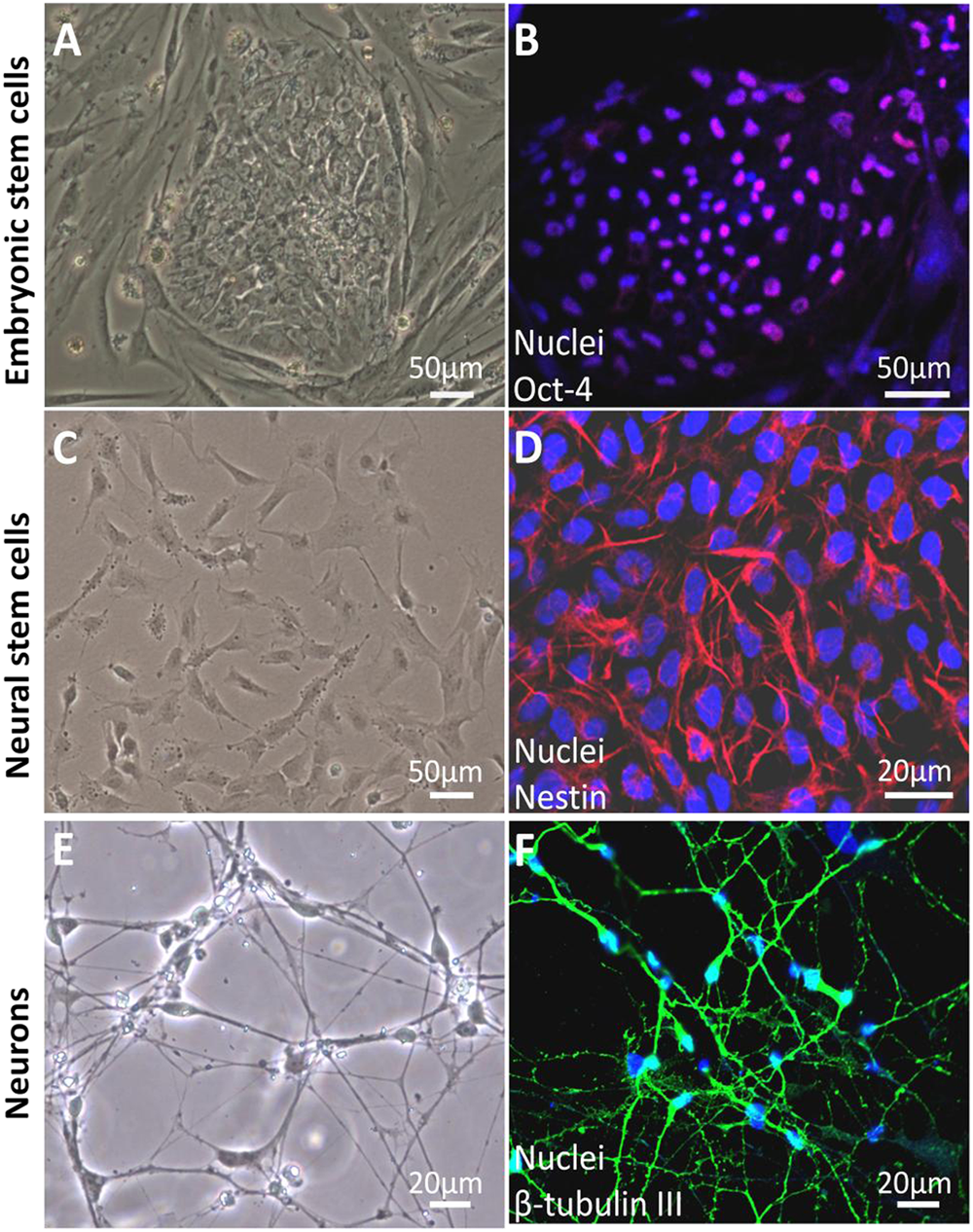
Differentiation of human embryonic stem cells (hESCs) into neural stem cells (NSCs) and neurons. hESCs grew as uniform flat colonies on mouse embryonic fibroblasts (A). hESCs expressed pluripotent stem cell markers Oct-4 (b, pink). Blue are cell nuclei (B). NSCs grew as a monolayer in the dish coated with Matrigel (C). NSCs were positive for the NSC-specific marker, nestin (g, red; D). Two weeks after culturing NSCs in neuronal differentiation medium, cells demonstrated neuron-like morphology with small round cell bodies and extending long projections (E). Differentiated cells were positive for the neuron-specific marker, β-tubulin III (b, green) (F). 52
Neuronal differentiation was observed by morphological assessment in the culture after 6 days of culturing the NSCs in neuronal differentiation medium. Differentiated neurons exhibited a round cell shape with small projections. Cell projections extended, and the extensive neuron networks were further formed in the culture over time (Figure 2E). Two-week-old neurons expressed neuron-specific markers β-tubulin III (Figure 2F) and microtubule-associated protein 2. In addition, these differentiated neurons expressed the synaptic marker synapsin-1, which is exclusively localized in the regions occupied by synaptic vesicles. They also expressed postsynaptic protein Homer 1, a family of postsynaptic scaffolding proteins.52,82,83 In addition, differentiated neurons exhibited a functional synapse. 84 The differentiation efficiency of NSCs into neurons was more than 90%. 52 Recapitulation of neurogenesis from hESCs in vitro provides a valuable and promising tool for the investigation of varying drug-induced developmental neurotoxicity, which is difficult to study in human patients.
Advantages of the In Vitro Stem Cell Neurogenesis Model
The greatest vulnerability of the developing brain to anesthetics occurs at the time of the growth spurt period in the brain. Many developmental events, including proliferation, migration, differentiation, formation of axons and dendrites, and synaptogenesis occur within this period. Thus, neuronal cell death may not be the only consequence of general anesthetics. Anesthetics may perturb neural development by influencing NSC proliferation, neurogenesis, synaptogenesis, and neuronal survival.85,86 Thus, our in vitro human stem cell model is promising for high-throughput examination of anesthetic-induced developmental neurotoxicity.
There are 5 main advantages of this experimental model in addressing the critical issues relevant to anesthetic neurotoxicity: (1) providing unlimited human NSCs and neurons for neurotoxicity study; (2) screening the neurotoxic effect of various anesthetics under controlled conditions (eg, dose, duration, and frequency of drug exposure); (3) allowing the dissection of the toxic effects of varying anesthetics on various stages of neuronal cell development, the individual neurodevelopment process, and the underlying molecular mechanisms; (4) investigating the potential preventive strategies to avoid this toxic effect; and (5) eliminating the need for a large number of animals. This approach was recently also used to examine the effects of alcohol on human NSCs, showing a dramatic phenotype of proliferation in response to ethanol. 87
Current Findings From the Stem Cell Neurogenesis Model
Using this in vitro human stem cell neurogenesis approach, we have examined for the first time the effect of anesthetics on hESC-derived NSCs and neurons.52,79 We used the intravenous anesthetic agent ketamine, which is widely used in pediatric anesthesia to provide sedation/analgesia to children during surgery or imaging. Ketamine and isoflurane are 2 of the most widely studied anesthetics for addressing neurotoxicity issues in rodent and primate models and the cell culture model.5 -9,18 -22,88 Using this in vitro stem cell neurogenesis model, our most recent study and that of others showed that these 2 drugs could influence neuronal developmental progress, including NSC proliferation, neurogenesis, and neuronal viability as detailed below.52,89
Neural Stem Cell Proliferation
In our study, NSCs were treated with 100 µM ketamine for 6 hours. Ki67 immunofluorescence staining and bromodeoxyurindine (BrdU) assay were performed for evaluation of cell proliferation. Ki67 is a nuclear, nonhistone protein and is preferentially expressed during late G1, S, G2, and M phases of the cell cycle, whereas resting, noncycling cells (G0 phase) lack Ki67 expression. There were more Ki67-positive cells in the 6-hour ketamine-treated culture compared with the control group. BrdU is a thymidine analog and can be incorporated into newly synthesized DNA strands of actively proliferating cells. The extent of BrdU incorporation allows for the assessment of cell proliferation rate. Consistent with the Ki67 staining, ketamine significantly increased BrdU incorporation after 6 hours of exposure. 52 The ketamine-induced increase in NSC proliferation was also observed in rats. 90 However, a recent in vitro study showed that ketamine decreased rat cortical NSC proliferation. 91 The observed differences in the ketamine on NSC proliferation may be a result of the use of different models. A recent study also found the enhanced proliferation in human neural progenitor cell line, ReNcell CX (immortalized by retroviral transduction with the c-myc oncogene). 89 These cells were derived from the cortical region of the human fetal brain. Increased proliferation of these neural progenitor cells were shown at a low concentration (0.6%) of isoflurane, whereas no effect was seen using a clinically relevant concentration (1.2%). Decreased proliferation was observed at a high concentration of isoflurane (2.4%). 89 Abnormal proliferation of NSCs could influence neurodevelopment. Ethanol, an NMDA antagonist, has long been recognized to be neurotoxic to the developing brain. Exposure to ethanol during brain development might promote neurodevelopmental defects.92,93 In one recent study, Nash et al 87 used the similar in vitro hESC-based neurogenesis system to study ethanol-induced early developmental toxicity. They found that ethanol induced a complex mix of phenotypic changes, including an inappropriate increase in stem cell proliferation and loss of trophic astrocytes. Thus, ketamine-induced alterations in NSC proliferation might also contribute to abnormal brain development.
Neurogenesis
Shorter exposure (1 hour) to a high dose of isoflurane (2.4%) had no effect on the differentiation of neural progenitor cell line (ReNcell CX) into neurons and astrocytes. 89 However, prolonged exposure (24 hours) to the same concentration of isoflurane significantly suppressed neuronal differentiation and promoted glial differentiation. This toxic effect may be attributed to differential regulation of calcium release through activation of endoplasmic reticulum–localized inositol-1,4,5-trisphosphate and/or ryanodine receptors. Pretreatment of ReNcell cultures with inositol-1,4,5-trisphosphate or ryanodine receptor antagonists (xestospongin C and dantrolene, respectively) mostly prevented isoflurane-mediated effects on the neuronal differentiation. 89
Ultrastructure in Neurons
Our electron microscopic images demonstrate the toxic effects on the cellular ultrastructure in ketamine-treated human neurons derived from hESCs. The neurons without ketamine treatment had very elongated mitochondria, whereas the ketamine-treated neurons displayed fragmented mitochondria. In addition, in the 100 µM culture treated with ketamine for 24 hours, large oval-shaped autophagosomes having some membrane-like material within them were found in almost every cell occupying the majority of the cytosol volume. Another observation was the disappearance of Golgi structures in the ketamine-treated neurons (Figure 3). 52
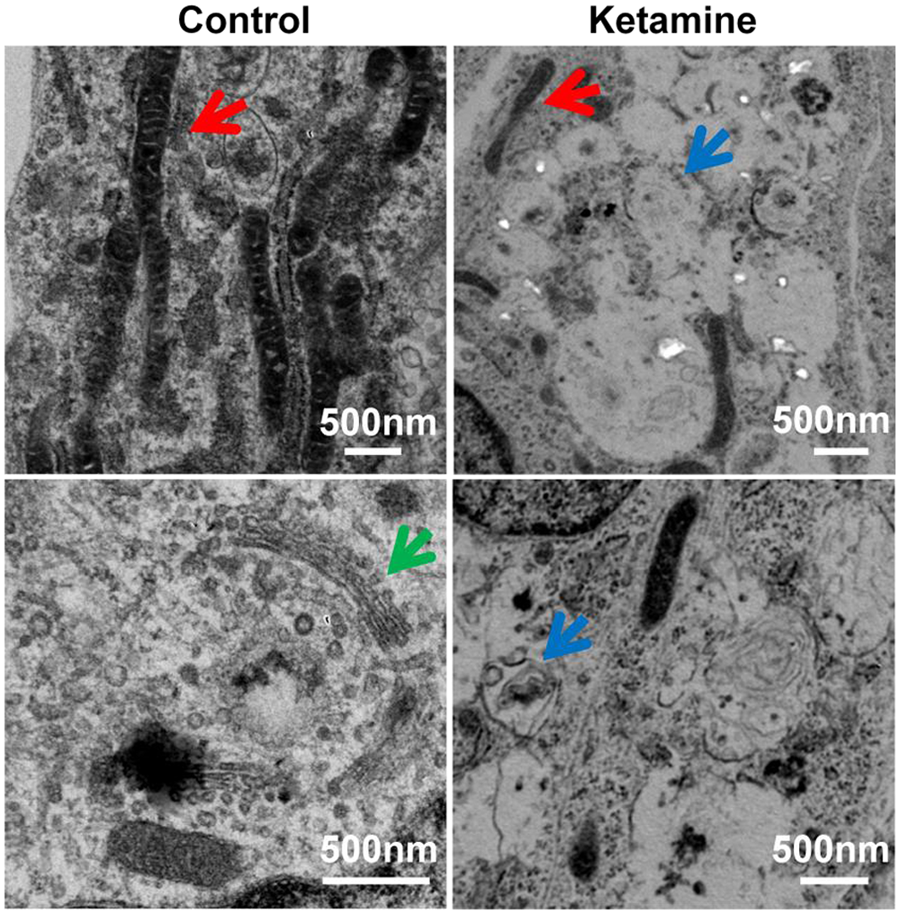
Ketamine causes abnormal cellular changes in ultrastructure: representative electron microscope images of differentiated neurons treated with the indicated concentrations of ketamine for 24 hours. Ketamine-treated neurons showed clear signs of the toxicity on the cellular ultrastructure. Abnormal ultrastructure of neurons included mitochondrial fragmentation and many autophagosomes with or without being packed with dense amorphous material. In addition, Golgi structures in the 100 µM ketamine-treated cells were not observed. Red, blue, and green arrows indicate mitochondria, autophagosome, and Golgi, respectively. 52
Neuroapoptosis
Neuronal apoptosis, or programmed cell death, is commonly recognized as one of the detrimental effects of anesthetics.2,21,94 However, how anesthetics induce neuronal apoptosis is not well understood. The central components of the programmed cell death are a group of proteolytic enzymes called caspases, which can be activated by various types of stimulation. The released cytochrome c activates caspase 9, which consequently induces caspase 3 activation, resulting in the cleavage of several cellular proteins, finally leading to the typical alterations related to cell apoptosis, such as DNA fragmentation in cell nuclei.27,94 DNA fragmentation can be used to detect apoptotic cells by terminal deoxynucleotidyl transferase–mediated deoxyuridine triphosphate in situ nick end labeling (TUNEL) staining. When compared with no treatment, prolonged exposure (24 hours) to 100 µM ketamine significantly increased caspase 3 activity. Consistent with caspase 3 activation, more TUNEL-positive cells with condensed and fragmented nuclei were observed in ketamine-treated cultures. 52
These findings from human stem cell–derived neurons were largely in line with several previous studies showing that only prolonged exposure and/or high doses of anesthetics could induce neuronal death. 57 For instance, 24 hours of 10 µM ketamine exposure induced a significant loss of postnatal-day-3 monkey frontal cortical neurons in culture. 22 In another study, treatment with 100 µM of ketamine for 48 hours resulted in the loss of 45% of neurons from fetal rats at 18 to 19 days of gestation. 21 Several recent reports showed that only a superclinical concentration of anesthetics could induce neurotoxicity. Mak et al 95 demonstrated that administration of 4000 µM ketamine for 48 hours caused significant death of both primary human neurons and differentiated neurons from the human neuroblastoma SH-SYY5 cell line. This finding was supported by Braun et al 94 , who showed that there was a significant increase in the number of apoptotic neurons differentiated from the human neuroblastoma SHEP cell line after 24 hours of 2000-µM ketamine treatment. A similar finding was also reported in primary cortical neurons from postnatal days 2 to 8 in mice. 96
The observed toxic threshold of ketamine varies in different studies because (1) the vulnerability of neurons from different species to ketamine may differ and/or (2) the neurons used in different studies might be at different developmental stages or the percentages of neurons could be different, influencing the vulnerability to ketamine. It is commonly considered that developing mammals are at greatest apoptotic risk during the most rapid period of growth of their central nervous system. Many developmental events, including neurogenesis, synaptogenesis, and neuron structure remodeling, occur within this period. Thus, neurons at different stages of brain growth burst might also exhibit different sensitivities to ketamine. It is also possible for ketamine at low concentrations to be able to alter other cell physiological activities, including neuronal receptor expression, the structure and branching of dendrites in neurons, and synaptogenesis, eventually resulting in impaired neuronal function. Additionally, the in vitro neuron culture system excludes the influence of other environmental factors (such as astrocytes and growth factors) in vivo that may increase the toxic threshold of ketamine. Short-term and low-dose anesthetics might be a safer strategy for a brief procedure or other sedation in children.
Conclusions and Future Perspectives
A large body of recent work suggests that neuroapoptosis is a major mechanism responsible for the progression of anesthetic-induced neurotoxicity. However, as we showed earlier, developing mammals are at greatest apoptotic risk during the most rapid period of brain growth. Many developmental events (eg, neurogenesis, synaptogenesis, and neuron structure remodeling) occur within this period. Thus, we have considered that neuronal cell death may not be the only consequence of general anesthetics. The in vitro hESC-related neurogenesis model can mimic the in vivo neuronal development process, providing a simple and promising in vitro human model and unlimited cell source for addressing such important anesthesia-related issues. It is important to note that this approach has a potential translational application because identification of the cellular mechanisms and signaling pathways that underlie anesthesia neurotoxicity will allow targeting of the molecules that can prevent this toxic effect. In addition, this established in vitro experimental model will provide numerous possibilities for future studies because one could use this high-throughput approach to test the effects of various conditions and anesthetics (eg, isoflurane and midazolam) on developing human neurons, and this could lead to major advances toward ensuring the safety of anesthesia for fetuses and infants.
As to the clinical practice, clearly more evidence is needed to guide clinical decision making on the safety of anesthesia during pregnancy as well as pediatric anesthesia. The National Center for Toxicological Research, an internationally recognized research center at the US Food and Drug Administration, and several universities are conducting research regarding the effects of anesthetics on the nervous systems of animals during developmental periods of rapid brain growth (smarttots.org). Currently, there is not sufficient evidence to determine whether these findings are translatable to the millions of young children who receive anesthesia each year. Despite the unequivocal neurotoxic effect of anesthetics in animal models, there is no direct clinical evidence showing any such effect in fetuses, infants, or children at any dose. The development of research in the area of stem cell biology and neuroscience will advance our understanding of anesthetic neurotoxicity. The ultimate goal would be not only to find the trigger for this catastrophic chain of events but also to prevent neuronal cell death itself.
One of the major caveats with this in vitro stem cell study lies in the relevance of the in vitro model to a true in vivo system. Specifically, there are many cell types present in the human brain, including neurons and glial cells, all of which interact extensively. Utilizing cultures of pure neurons may not allow for the accurate assessment of the effects of anesthetics on intact brains. Thus, it will be necessary to confirm the in vitro findings in animal models, including nonhuman primates. A second limitation of this model is the use of neurons derived from human stem cells. The ideal model for these types of studies would be neurons that have been harvested from a developing human brain. However, such a model is currently not feasible and would likely not yield enough cells for the studies. Although neurons derived from stem cells may not exactly mimic the human brain environment, they express several important neuronal markers and functional synapse, representing an acceptable and promising model.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by P01GM066730, R01HL034708 from the NIH, Bethesda, MD, and by FP00003109 from Advancing a Healthier Wisconsin Research and Education Initiative Fund (to Dr Bosnjak).