
Abstract
Introduction:
Turnout, or external rotation of the lower limbs, is a fundamental skill in classical ballet. While the hip is turnout’s main contributor, the coordinated roles of the knee and ankle remain unclear under dynamic and aerial tasks, such as jumps. Previous studies have focused on static or bipodal tasks, overlooking unipodal landings in jumps. The aim was to analyze the kinematic contributions of the hip, knee, and ankle to turnout during three ballet jumps from fifth position: Assemblé Dessus (bipodal, with lateral aerial body displacement), Sissone Ouvert En Avant (unipodal, with anterior displacement), and Sissone Ouvert Devant (unipodal, with lateral displacement).
Methods:
In this observational cross-sectional study, we assessed 30 female pre-professional dancers (20.1 ± 2.6 years; 11.7 ± 4.1 years ballet experience) using a cluster-based wedge-shaped marker protocol to improve accuracy of hip and knee axial rotation measurements. We used a six-degree-of-freedom model to calculate peak rotations and their timing, which were compared across joints and jump phases (preparation, flight, and landing) using repeated measures ANOVA (P < .05).
Results:
The hip consistently exhibited the greatest external rotation across all jumps and phases (P < .001). In both Sissones, hip rotation remained stable from flight to landing, whereas in Assemblé, hip and knee rotations decreased upon landing (P < .001). Ankle rotation increased during landing (P < .001) and was the lowest during flight. Timing of rotation peaks varied across joints: the hip peaked earlier, while the ankle reached its maximum in late flight and late landing.
Conclusion:
The hip is the primary contributor to turnout in all jumps, whereas the knee and ankle act complementarily, with the knee reducing its contribution and the ankle increasing it from flight to landing. Jump type and landing support influence rotational contributions, reinforcing the need for training that optimizes coordination and joint alignment.
Key Points
The hip is the main contributor to turnout across all ballet jumps, while the knee and ankle act as complementary joints adjusting rotation during preparation and landing.
Turnout control is dynamic and phase-dependent, requiring coordinated timing of hip, knee, and ankle rotations to maintain alignment and prevent compensatory stress.
Training should emphasize functional turnout centered at the hip, promoting joint alignment and reducing harmful compensations at the knee and ankle during jump preparation and landing.
Introduction
The biomechanics of classical ballet is a key area of study for understanding the mechanisms that support dancers’ technique and performance. Among the ballet elements analyzed in the literature there is the turnout, which is the maximum external rotation of the lower limbs, essential for maintaining both the aesthetic and technical aspects of ballet particularly during jumps.1-6
While the hip joint has been identified as the main driver of turnout during bipodal jumps (Sauté), the knee and ankle also contribute significantly, albeit secondarily, through compensatory rotations and transverse plane adjustments. During the unipodal Sissone jump, the hip accounts for 60% to 70% of the total external rotation, followed by the knee, with 20% to 25%, and the ankle, contributing only around 5% to 10%, primarily involved in fine-tuning foot positioning. When considering turnout during a task performed in first-feet position, 7 reported that the largest share of external rotation occurred at the hip. However, the authors also highlighted the substantial role of the knee, particularly when fully extended, suggesting a potential stress on the knee if the turnout position is forced. These investigations, however, were predominantly conducted under static conditions or during bipodal support tasks, without considering the specific demands of unipodal or bipodal landings during jumps, in which the challenges of postural control and aerial displacement likely amplify the biomechanical requirements.
Although several authors6-8 have contributed to the understanding of ballet turnout, an important gap remains regarding turnout alignment during jumps that combine aerial displacement, transverse-plane control, and rapid postural transitions, characteristics inherent to classical ballet technique. Investigating more complex movements in which turnout is performed, such as jumps with unipodal landings that involve aerial body displacement and challenging foot positions, can contribute to the understanding of the different biomechanical strategies employed to preserve extreme lower-limb rotations under varying motor demands.8,9 Quanbeck et al, 10 in a recent cross-sectional study, partially addressed this gap by investigating the kinematic contributions of the hip, knee, and ankle joints to turnout during bipodal Assemblé Dessus and unipodal Sissone Ouvert jumps performed in fifth position, using the Plug-in Gait skin-marker protocol. However, this protocol lacks accuracy and reliability in measuring axial rotations,11-14 and the results obtained were subject to soft tissue artifacts influence.
While previous studies have contributed to understanding the turnout movement, information is scarce on lower limb biomechanics during complex ballet jumps that involve unipodal landings and multidirectional aerial displacements. In particular, movements such as the Sissone Ouvert Devant, which is characterized by lateral displacement combined with sustained hip abduction and external rotation during flight, remain largely unexplored in biomechanical research. These tasks impose substantial demands on postural control, requiring precise lumbopelvic stability, control of pelvic orientation, and coordination of the lower limbs to maintain alignment over a reduced base of support. 15 Additionally, the interaction between lower-limb rotation and the dynamic role of the working limb as a counterbalance further increases neuromuscular and coordinative demands. 16 Understanding how turnout is maintained under these conditions is essential, as such dynamic tasks may reveal joint-specific strategies and compensatory mechanisms not observable in static and bipodal conditions.
We also addressed the methodological limitation of the previous study, 10 by using rigid clusters to improve the reliability of axial rotation tracking and minimize the systematic errors frequently observed in protocols based on isolated skin-mounted markers.13,17 In particular, we combined the use of a conventional rigid cluster on the leg with a wedge-shaped cluster on the thigh, which showed to be advantageous for assessing the external rotation required in ballet turnout.18,19
Therefore, we aimed at analyzing the kinematic contributions of the hip, knee, and ankle joints to lower-limb external rotation (turnout) during three classical ballet jumps: a bipodal laterally displaced jump (Assemblé Dessus), a unipodal anteriorly displaced jump (Sissone Ouvert En Avant), and a unipodal laterally displaced jump (Sissone Ouvert Devant). Additionally, we aimed at comparing lower-limb kinematics across the different jumps, identifying potential compensatory patterns or specific motor strategies depending on aerial displacement and postural configuration. The study was based on three hypotheses: (1) the hip would contribute most to turnout across all jump phases, primarily due to its anatomical capacity for axial rotation; 7 (2) the knee would contribute minimally to turnout because of its limited transverse-plane motion, while remaining aligned with the ankle during preparation and landing phases; 20 and (3) the ankle would exhibit greater external rotation during preparation and landing phases by taking advantage of foot-ground friction to increase its range of motion. 21 The outcome of the present investigation is expected to contribute both to technical improvement and injury prevention in classical dancers, providing biomechanical insights to support more precise pedagogical and therapeutic interventions.
Methods
Study Design and Participants
This observational cross-sectional study was approved by the Research Ethics Committee of the School of Medicine of the University of São Paulo (CAAE 70365223.5.0000.0068). We recruited 30 young pre-professional ballet dancers (20.1 ± 2.6 years old; body mass index of 20.7 ± 2.2 kg/m2; 11.7 ± 4.1 years of classical ballet practice; 8.2 ± 7.0 hours of weekly training) from ballet schools and amateur classical ballet companies in the Greater São Paulo area, Brazil. All participants (or their legal guardian, if younger than 18 years) signed an informed consent form. The inclusion criteria were female dancers with at least 5 years of ballet practice and a minimum of 4 hours of training per week. The sample size calculation (n = 30) was based on a pilot study and performed using G*Power, 22 considering a moderate effect size (f = 0.24) for peak hip external rotation, defined as the primary outcome, within a repeated-measures ANOVA model, with a significance level of 5% and statistical power of 80%. Only datasets meeting stringent technical criteria for marker tracking and processing (data acquisition and marker tracking quality), without significant gaps or loss of data continuity, were included in the analysis to ensure the reliability and accuracy of the kinematic measurements. By significant gap, we mean any loss of marker visibility or tracking exceeding 10 consecutive frames. Gaps within this limit were filled using fifth-order spline interpolation; beyond this threshold, interpolated data were considered likely to reduce kinematic accuracy in the transverse plane.
We assessed passive hip external rotation to establish a reference parameter for the hip maximum rotational amplitude at rest, without the influence of postural or dynamic compensations. For this purpose, maximum hip external rotation was measured with participants lying prone on an examination table, with the knee flexed at 90º, using a biaxial electrogoniometer (Biometrics, Newport, UK). The fixed arm of the electrogoniometer was attached to the examination table, and the moving arm was secured to the proximal portion of the tibia. The mean (±SD) of the participants’ maximum passive hip external rotation was 61.1° ± 8.5°.
Ballet Tasks
The tasks included three typical ballet jumps initiated from the fifth-feet position: Sissone Ouvert En Avant (Figure 1, upper panel), Sissone Ouvert Devant (Figure 1, middle panel), and Assemblé Dessus (Figure 1, lower panel). In the Sissone Ouvert En Avant (Figure 1, upper panel), the preparation phase begins with the supporting limb positioned in front. During flight, with anterior displacement, the lower limbs are abducted in opposite directions, with the supporting limb moving forward while the working limb extends backward. Landing is performed with the knee of the supporting limb flexed and the hip of the working limb fully extended. The working limb refers to the limb performing the main motion after takeoff, while the supporting limb refers to the limb bearing the body weight during landing. The kinematics of the Sissone Ouvert Devant (Figure 1, middle panel) are similar to those of the Sissone en Avant, but the displacement occurs in the frontal plane, with the body moving laterally. During flight, the hip of the working limb abducts and moves away from the supporting limb, with both hips and knees extended. Landing occurs on the supporting limb with a flexed knee, while the working limb remains extended with the hip still abducted. In both Sissone jumps, pelvic and lower-limb joint alignment in the frontal plane should be maintained throughout the jump, without rotational compensations.

Upper panel: Sissone Ouvert En Avant; middle panel: Sissone Ouvert Devant; lower panel: Assemblé Dessus. For all jumps: (A) preparation phase, (B) flight phase, and (C) landing phase.
In the Assemblé Dessus (Figure 1, lower panel), the movement begins with the working limb positioned behind the supporting limb, which remains in contact with the ground. Jump preparation starts when the working limb moves sideways in abduction, while the supporting limb performs knee extension. The flight phase begins when both limbs leave the ground and come together in the air, maintaining the same position as during preparation; subsequently, the working limb crosses in front of the supporting limb. During landing, both limbs touch the ground simultaneously, and the technique requires that the pelvis remain in a neutral and aligned position, without tilting or rotating to the sides.
Data Acquisition and Analysis
We used eight infrared cameras (Vicon VERO; Oxford Metrics, Oxford, UK) to track the trajectory of sixteen 9.5 mm spherical reflective markers attached to the pelvis and lower limbs of each participant (Figure 2) and a force plate (AMTI OR-6-1000, Watertown, MA, USA) to record vertical ground reaction force data, both sampled at 100 Hz. We used four markers, positioned on the right and left anterior superior iliac spine and posterior superior iliac spine, to establish and track the pelvis anatomical reference frame (ARF). Each thigh ARF was defined using the lateral and medial femoral epicondyles and the estimated locations of the hip joint center, based on the regression equations by Davis et al. 23 To address previously reported limitations in the assessment of axial rotations using skin-mounted markers,10,13,17 we employed a rigid cluster-based tracking approach to improve measurement reliability. We positioned a wedge-shaped cluster, consisting of a 5-cm-high, lightweight triangular 3D-printed plastic prism fitted with four markers and presenting a concave face to allow greater contact with the skin, on the distal portion of the iliotibial band, 2 cm above the lateral epicondyle (Figure 2), 19 to track thigh movement during dynamic trials. We defined shank ARFs from the respective medial and lateral femoral epicondyles and the medial and lateral malleoli, and tracked with a rectangular 10 × 7.5 cm rigid cluster with four markers, positioned on the respective mid-lateral portion. Tracking the kinematics of lower limb joints via a conventional rigid cluster of markers on the shank and a wedge-shaped cluster on the thigh has been shown to enhance the accuracy of hip and thigh axial rotation measurements in comparison to skin-mounted markers, especially for dynamic tasks involving sustained large hip axial rotations18,19 like those occurring in ballet jumps. Thus, this markers’ configuration was selected to minimize the effect of soft tissue artefacts and to improve the robustness of kinematic measurements. Finally, we established and tracked each foot ARF from markers positioned on the medial and lateral malleoli, heel, and the base of the second metatarsal (Figure 2). To improve the consistency of markers’ placement, all participants were instrumented by the same operator.
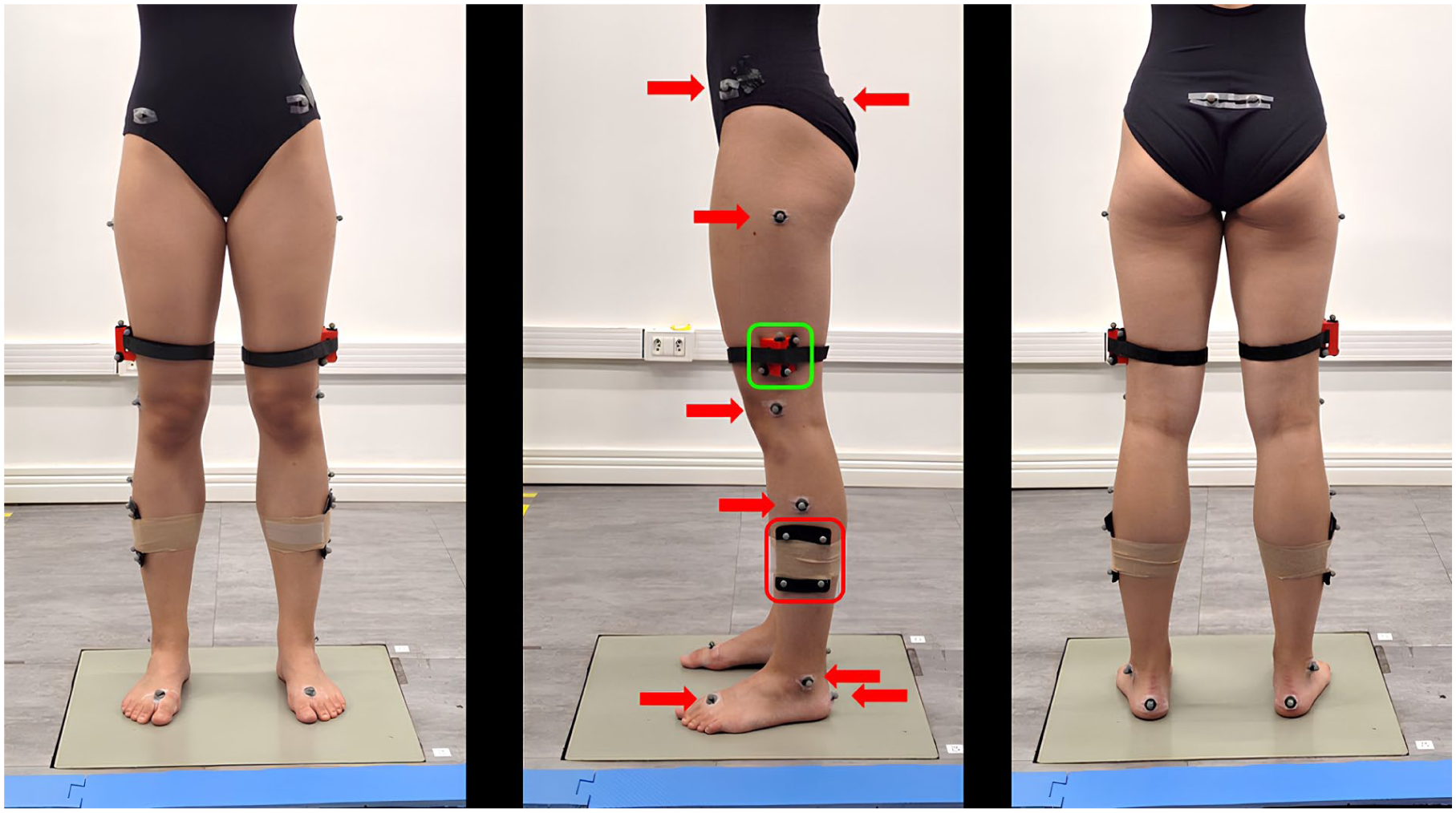
Ballerina with wedge-shaped cluster (green box, proximal) and conventional rigid cluster (red box, distal); plug-in Gait protocol markers (arrows).
As a pilot repeatability analysis, intra-rater reliability was tested using the intraclass correlation coefficient (ICC) [ICC(1,k)]10,24,25 based on repeated measures of skin marker placement conducted by the same evaluator on a subset of two participants, across three distinct sessions. Repeated measurements were taken for peak external rotation of hip and ankle joints during the Assemblé jump on the dominant limb of each participant, the same side that was assessed in the main experimental data acquisition. In each session, five trials were collected per participant, resulting in a total of 15 trials per subject. Excellent intra-operator reliability was observed for peak external rotations of hip and ankle joints (ICC hip = 0.92; ICC ankle = 0.99). However, the use of only two participants may limit the precision and stability of these estimates and should therefore be considered a limitation of this pilot reliability analysis.
To ensure proper jump execution, the dancers were instructed to initiate the jump outside the force plate and to land on it. Prior to data acquisition, they were given approximately 10 minutes to warm up and become familiar with the jump tasks. For each participant, data acquisition included an anatomical calibration trial (upright double-leg static) and a minimum of five valid trials per jump. When necessary, up to eight attempts per jump were collected to replace trials excluded due to incomplete marker tracking or technical issues. The different jump modalities were performed in a randomized order to minimize potential order effects, such as learning, fatigue, or performance adaptation, thereby reducing possible bias related to task sequence.
For each jump, we analyzed a single limb according to its functional and biomechanical importance in the execution of the specific jump. All movements were performed using the dominant side only, defined as the limb preferentially used by the dancers to initiate movements and perform unipodal landings during ballet practice. Most dancers used the right dominant limb (right). In the bipodal Assemblé Dessus jump, we analyzed the working limb, as it initiates and executes the main movement, while the supporting limb primarily contributes to stabilization and body-weight acceptance. In the unipodal Sissone Ouvert jumps, we analyzed the supporting limb because it experiences greater biomechanical demand during landing and is responsible for maintaining body alignment in the final stage of the jump. This approach was adopted to reduce inter-individual variability and to focus on the functional demands of each jump.
We tracked marker trajectories in Nexus 2.6 and processed in Visual3D software (Visual3D, HAS-Motion, Ontario, Canada) to establish a six-degree-of-freedom model of both lower limbs, including the feet, shanks, thighs, and pelvis. Joint centers and reference frames of each segment were defined according to marker positions acquired in the calibration anatomical trial. Marker trajectories and joint angles were filtered using a zero-lag, fourth-order Butterworth filter with a cut-off frequency of 6 Hz.
Jump phases were segmented as follows: the preparation phase began when knee flexion velocity reached 10% of its peak; the flight phase started with the loss of foot contact with the ground, the flight phase started when both feet were off the ground (take-off); landing began at the first contact with the force platform, identified as the instant when the vertical ground reaction force reached 3N, and ended when the vertical velocity of the heel(s) reached zero. For the Assemblé Desus, take-off was defined as the instant when the vertical position of the contralateral second metatarsal head marker exceeded its static pre-jump value by 15 mm. For the Sissone Ouvert En Avant and the Sissone Ouvert Devant take-off was defined when the ipsilateral second metatarsal head marker reached the same threshold. The thresholds for preparation, flight, and landing phase onsets were empirically determined through preliminary analyses to optimize sensitivity in detecting jump events, while minimizing false positives.
The following discrete variables were extracted from the hip, knee, and ankle joint angle time series in the transverse plane for each jump and phase: (1) the external rotation peak of each joint (i.e., the minimum axial rotation angle) which together quantifies the turnout (the rotation of the lower limb as a whole); and (2) the timing of the external rotation peak, expressed as a percentage of jump duration. According to the adopted convention for angular motion, external rotation was defined as negative and internal rotation as positive; thus, external rotation peak corresponds to the minimum recorded rotation at each phase.
To address the primary research questions of the study: (1) which joint contributes most to turnout in each phase of the jump, and (2) how the contribution of each joint changes across phases, we compared these two variables across phases and joints using separate repeated-measures ANOVAs for each jump, followed by post hoc Newman-Keuls tests (P < .05) (Jamovi v. 2.5.3). This approach was chosen to allow more direct interpretation of joint-specific and phase-specific effects without conflating their meanings, and is consistent with previous three-dimensional kinematic studies in ballet that examined joint and phase effects as separate within-subject factors. 26
Results
All statistical results are shown in Table 1, including ANOVAs P values between phases and between joints and the post hoc P values are presented in the text for better readability. Figures 3 to 5 present the mean temporal profiles of hip, knee, and ankle internal/external rotations, as well as the mean values for the peak rotation angles of each joint across the jump phases. To facilitate visualization and interpretation, the bar plots display the absolute values of the peak rotation angles.
Mean, Standard Deviation, Effect Size (η2 P ), and P Values for the Comparisons of the External Rotation Peak and Rotation Peak Timing of the Hip, Knee, and Ankle During the Preparation, Flight, and Landing Phases of the Sissone Ouvert Devant, En Avant, and Assemblé Dessus Jumps.
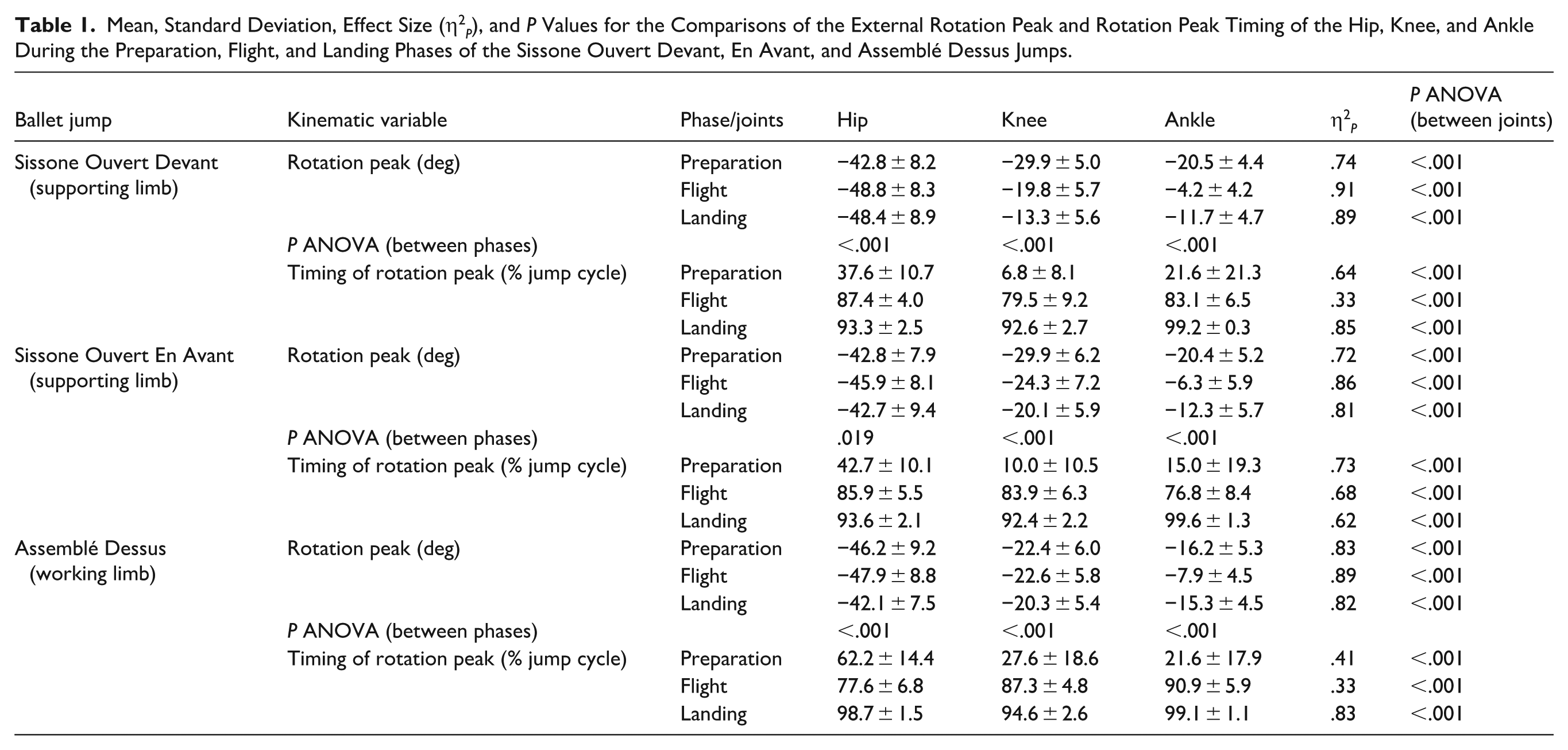
Upper panel: mean time series and standard error of the joint angles in the transverse plane for hip A), knee (B), and ankle (C) during Sissone Ouvert Devant jump; negative values indicate external rotation. Lower panel: mean and standard error of the absolute value for external rotation peak for hip (A), knee (B), and ankle (C) during preparation, flight, and landing phases of the Sissone Ouvert Devant.
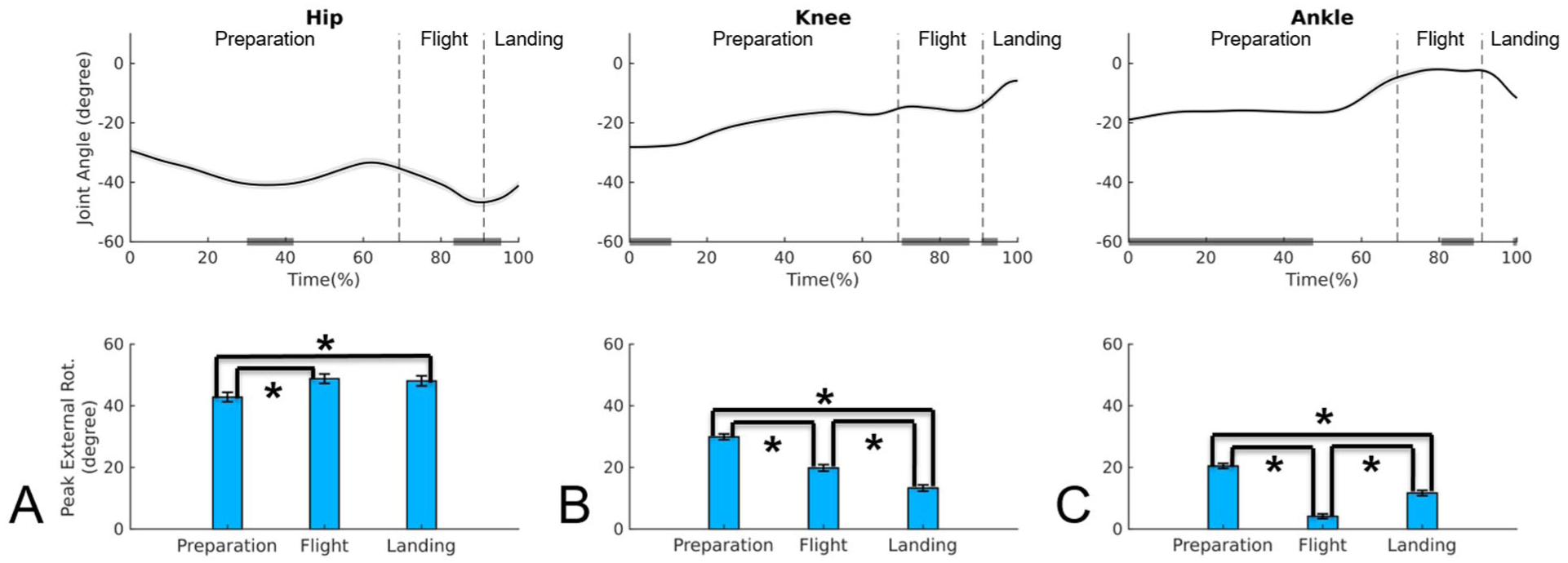
Upper panel: mean time series and standard error of the joint angles in the transverse plane for hip (A), knee (B), and ankle (C) during Sissone Ouvert En Avant jump; negative values indicate external rotation. Lower panel: mean and standard error of the absolute value for external rotation peak for hip (A), knee (B), and ankle (C) during preparation, flight, and landing phases of the Sissone Ouvert En Avant.
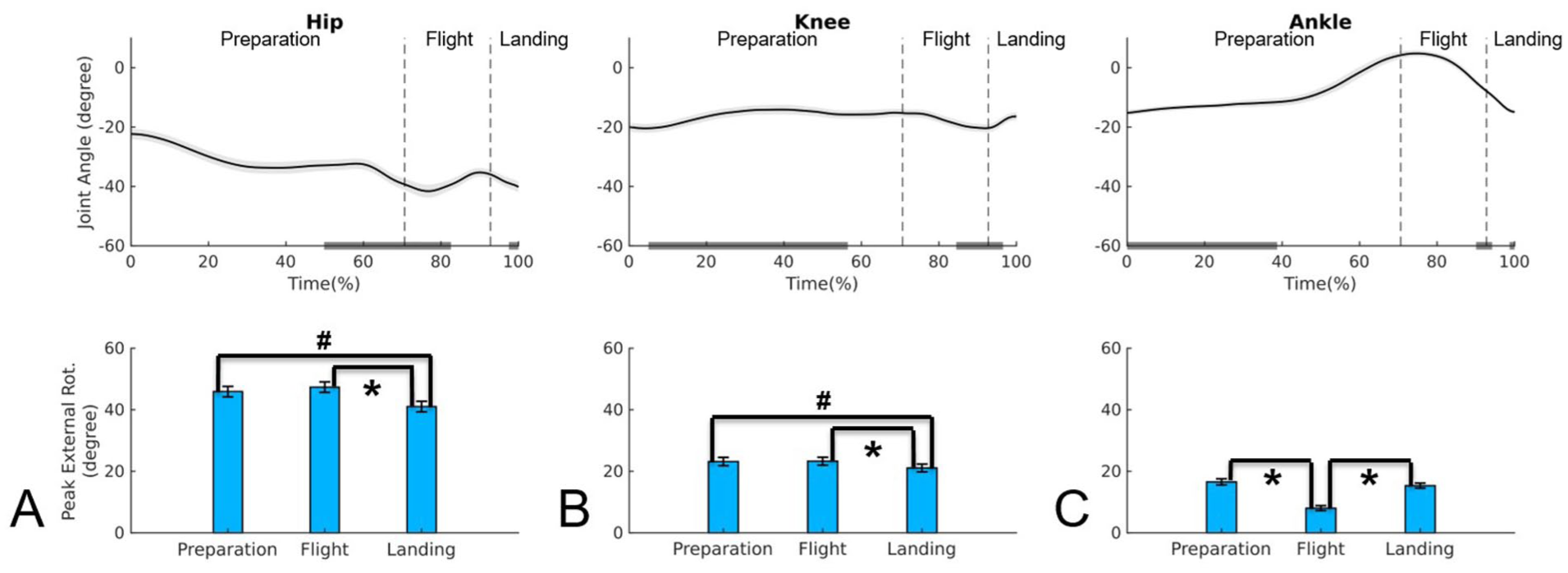
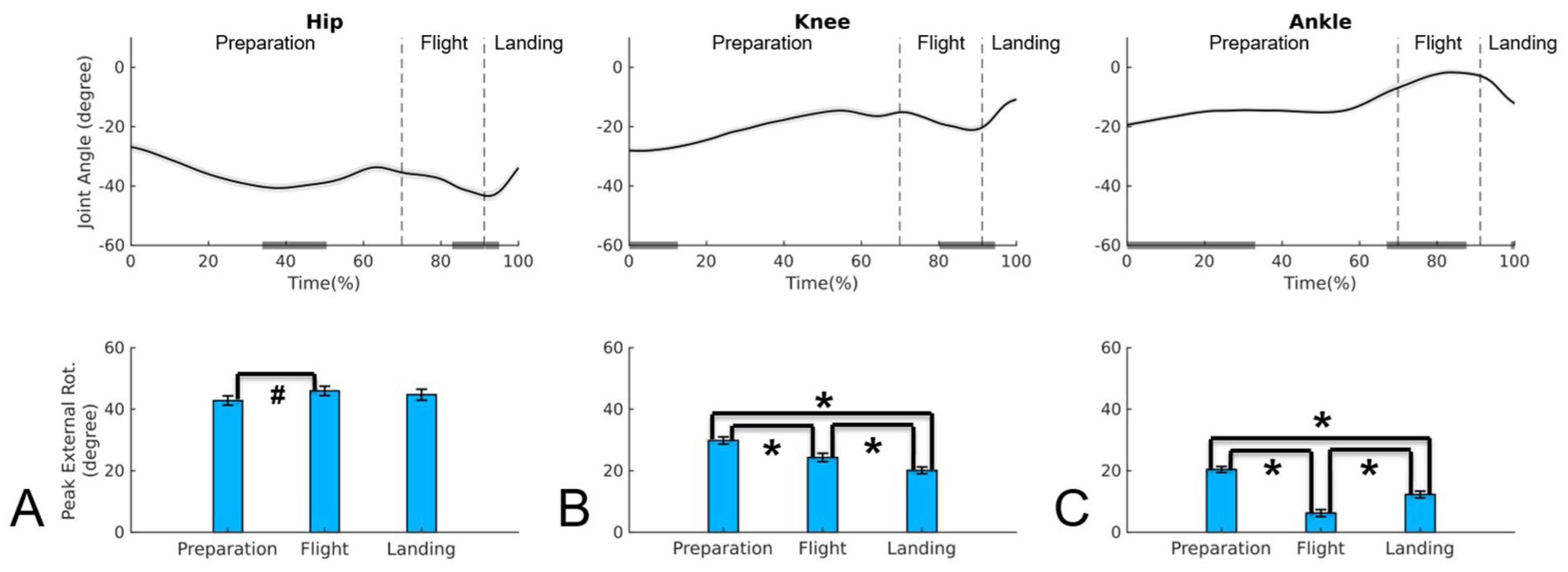
Upper panel: mean time series and standard error of the joint angles in the transverse plane for hip (A), knee (B), and ankle (C) during Assemblé Dessus jump; negative values indicate external rotation. Lower panel: mean and standard error of the absolute value for external rotation peak for hip (A), knee (B), and ankle (C) during preparation, flight, and landing phases of the Assemblé Dessus.
Sissone Ouvert Devant
In the Sissone Ouvert Devant (Table 1, Figure 3), the hip external rotation peak was greater during flight compared to preparation (P < .001) and remained higher during landing (P = .380). In the knee, the external rotation peak progressively decreased from the preparation to the landing phase (P < .001). In the ankle, the peak was higher during preparation compared to other phases (P < .001) and lowest during flight (P < .001). When comparing joints, the hip showed larger peaks than both the knee and ankle across all phases (P < .001). Compared to the ankle, the knee exhibited higher peaks during preparation and flight (P < .001), while no difference was observed between them during landing (P = .377).
Regarding the timing of the external rotation peak (Table 1), the hip and ankle reached their peaks in the middle of the preparation phase. For some subjects, the ankle reached its peak at the beginning of the phase, while for others it occurred at the middle, explaining the extent of the gray bar. The knee peaked at the beginning of the preparation phase. During flight, the hip and ankle reached their peaks at the end of the phase, whereas the knee peaked in the middle. In the landing phase, both the hip and knee reached their peaks at the beginning, while the ankle reached its peak at the end.
Sissone Ouvert En Avant
In the Sissone Ouvert En Avant (Table 1, Figure 4), the peaks occurred in a pattern similar to that observed during Sissone Ouvert Devant. The hip rotation peak was greater during flight compared to preparation (P < .05), and this higher peak was maintained from the flight to the landing phase (P = .309). In the knee, a progressive decrease in peak rotation was observed from preparation to landing (P < .001). At the ankle, the peak was higher during preparation compared to other phases (P < .001), with the lowest value recorded during flight (P < .001). When comparing joints, the hip exhibited the highest peaks across all phases (P < .001), followed by the knee, which also presented higher peaks than the ankle in all phases (P < .001).
Regarding the timing of external rotation peaks (Table 1), joints generally reached their peaks at similar points of the movement cycle as observed in the Sissone Ouvert Devant. Exceptions were observed for the knee and ankle during flight, with the knee peaking at the end of the phase and the ankle at the beginning.
Assemblé Dessus
In the Assemblé Dessus (Table 1, Figure 5), the joints exhibited a different kinematic behavior compared to the other two jumps. The highest hip external rotation peak occurred during preparation and flight, with no significant difference between them (P = .361). A significant decrease in hip rotation peak was observed from flight to landing (P < .001), with the landing peak being lower than that during preparation (P < .019). The knee displayed a pattern similar to the hip, with higher peaks during preparation and flight, followed by a reduction at landing (post hoc: P < .001), where the peak was lower than during preparation (P < .036). The ankle exhibited its highest peaks during preparation and landing, which did not differ significantly (P = .278); in addition, its value decreased from preparation to flight (P < .001) and increased from flight to landing (P < .001). When comparing joints across phases, the hip maintained the highest peaks in all phases (P < .001). The knee reached its highest peak during flight, whereas the ankle peaked during preparation, decreased during flight (P < .001), and recovered during landing (P < .001), with no significant difference between preparation and landing, a pattern consistent with the other two jumps.
Regarding the timing of the rotation peaks (Table 1), during preparation, the hip peak occurred at the end, while the knee and ankle peaks occurred at the beginning. During flight, the hip peaked at the beginning, the knee from the middle to the end, and the ankle at the end. In the landing phase, the hip and ankle reached their peaks at the end, while the knee peaked at the beginning.
Discussion
This study aimed to analyze the kinematic contributions of the hip, knee, and ankle joints to lower limb external rotation (turnout) during three classical ballet jumps, using a rigid cluster model with a wedge configuration to overcome previously described methodological limitations in measuring axial rotation. 18 Overall, the results confirmed our first hypothesis that the hip is the joint most responsible for turnout across all jumps and phases. The knee showed a greater contribution compared to the ankle, but this contribution was smaller than that of the hip, particularly during preparation, partially refuting our second hypothesis. The knee and ankle joints appear to act in a complementary manner, with the knee reducing its contribution and the ankle increasing its contribution from flight to landing. Confirming the third hypothesis, the ankle played a more prominent role during landing of the unipodal jumps, reinforcing its function not only as a complementary joint to the turnout but also as a key structure in maintaining alignment when postural control demands are higher. In the unipodal jumps (Sissone Ouvert En Avant and Sissone Ouvert Devant), the hip maintained its external rotation from flight to landing, whereas in the bipodal jump (Assemblé Dessus), a reduction in hip rotation was observed from preparation to landing. These findings highlight how different body aerial displacements, spatial orientations of the body and lower limbs, and landing support (uni- or bipodal) influence the kinematic contributions to turnout, providing new insights into joint alignment during complex technical ballet tasks. In addition, the magnitude of external rotation during all jumps were consistent with the expected clinical range of motion under static conditions using electrogoniometers (range 44 to 75°, mean 61.1 ± 8.5°).
In both Sissone jumps, the hip maintained higher peaks during flight and landing, suggesting that well-trained dancers can sustain turnout even under challenging postural control conditions, consistent with classical ballet principles. However, joint alignment throughout the lower limb is expected to be maintained during the entire jump, 27 which was not observed in all three jumps, as the three joints displayed different behaviors across phases (Figures 3-5). In the Assemblé Dessus, a reduction in hip rotation was observed from preparation to landing phase, which may reflect the characteristics of bipodal jumps, where simultaneous foot contact with the ground while landing tends to increase the rigidity of the kinematic chain, limiting fine rotational adjustments at the hip and knee. To compensate for this reduction in hip rotation during bipodal landing phase, the ankle played a complementary role, exceeding the range of motion of the knee, which could potentially increase joint overload while landing, as discussed in previous studies addressing joint compensations and their potential injury implications.9,28,29
The knee, unlike the hip, progressively reduced its external rotation peaks from preparation to landing in all three jumps, which may reflect a protective strategy to avoid rotational overload during high joint-demand moments, i.e., at landing. This has been documented in systematic reviews on may increase mechanical demands associated with injury risk, factors in ballet, which identify knee rotational overload as a factor linked to ligament and patellofemoral injuries.28,30 From a technical standpoint, the knee is expected to follow the external rotation of the hip without causing transverse-plane torsion, maintaining alignment between the femur, tibia, and foot throughout all jump phases. However, this was not consistently observed, as the knee did not follow the biomechanical pattern of the hip. This is particularly noticeable during landing of the Assemblé Dessus (see upper panel of Figure 5) and during preparation of both Sissone jumps (see upper panels of Figures 3 and 4).
The ankle, in contrast, displayed an inverse pattern relative to the knee across all three jumps: higher peaks during preparation, a decrease during flight, and a return to preparation angle values during landing. This suggests a distal compensatory function, particularly at ground contact, when the ankle assists with fine base-of-support adjustments and foot realignment to sustain turnout under impact demands, as described in biomechanical analyses identifying the ankle as a fundamental stabilizer during landing tasks.20,29 In classical ballet, landing requires the foot to remain externally rotated, sustaining the fifth position, without compromising joint stability, which demands active ankle control and precise eccentric strength. 31 While such compensations may serve a short-term functional purpose, they can compromise both aesthetics and joint health if perpetuated in habitual technique. 32
This shift in joint contribution across jump phases illustrates that turnout is not a static position but a complex, coordinated joint movement that adapts to the biomechanical requirements of each phase.26,28 The timing of external rotation peaks further reinforces this dynamic view. Rotational control is not simultaneous across joints; rather, it is distributed over time, with each joint reaching its rotation peak at a distinct moment. This requires specific coordination strategies and neuromuscular activation during each jump phase. Such patterns have been previously demonstrated in 3-dimensional kinematic analyses of dancers, highlighting the importance of joint timing for technical efficiency and injury prevention.27,30
This study has a few limitations. First, the analysis was limited to three specific jump types, which may not capture the full spectrum of technical demands in classical ballet. Other jumps, particularly those of greater complexity, may require different strategies for maintaining turnout. Furthermore, the sample consisted exclusively of pre-professional dancers. Professional dancers, with longer training histories and broader exposure to diverse repertoires, may develop distinct motor patterns or technical habits that could influence their turnout strategies.33-35 Finally, the cross-sectional study design restricts the ability to establish causal relationships between the observed variables and may increase mechanical demands associated with injury risk. Future longitudinal studies incorporating a broader range of movements and biomechanical variables are warranted to further advance the understanding of dynamic turnout in dancers.
Practical and Clinical Applications and Implications
From a methodological perspective, the use of wedge-shaped rigid clusters represented a significant improvement compared to a previous study by the same authors, 10 which used only the Plug-in-Gait protocol. In the present analysis, the external rotation angles differed from those previously obtained using isolated markers. The adoption of the wedge-shaped cluster 19 appears to be a promising solution for detecting variations in axial rotations in ballet, where pelvis control and hip external rotation must be maintained during unipodal support with aerial body displacement.
The results provide valuable insights for more precise training and clinical interventions. Based on these findings, technical training should focus on developing a more functional turnout, that is, biomechanically adequate and centered on the hip, to avoid harmful compensations at the knee and ankle that may increase mechanical demands associated with injury risk, and compromise movement aesthetics. Maintaining rotational alignment during landing, particularly the stabilizing role of the ankle, should be carefully addressed to ensure safety and efficiency in impact absorption.
Conclusion
In summary, in unipodal and bipodal jumps in classical ballet, the hip consistently exhibited the highest external rotation peaks across all phases, while the knee and ankle played specific roles, particularly during preparation and landing, respectively. The use of wedge-shaped rigid clusters appears to be a promising solution for detecting variations in axial rotations in ballet turnout. By demonstrating how turnout is organized and articulated, this study contributes to a deeper understanding of the kinematics and dynamics of ballet jumps, providing meaningful insights to support more effective teaching, injury prevention, and the consideration of dancers’ individual structural characteristics.
Footnotes
Ethical Considerations
This observational cross-sectional study was approved by the Research Ethics Committee of the School of Medicine of the University of São Paulo (CAAE 70365223.5.0000.0068). All participants (or their legal guardian, if younger than 18 years) signed an informed consent form.
Author Contributions
LCM and ICN participated in the design of the study, contributed to data collection, data reduction/analysis and interpretation of results; MOM and BMP participated in the data collection, data reduction/analysis and interpretation of results; MIVO participated in the design of the study, contributed to data analysis and interpretation of the results; PC contributed to data analysis and interpretation of results. All authors contributed to the manuscript writing. All authors have read and approved the final version of the manuscript, and agree with the order of presentation of the authors.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The National Council for Scientific and Technological Development (Conselho Nacional de Desenvolvimento e Pesquisa, CNPq/Brazil) funded I.C.N. Sacco (Process: 302558/2022-5) and Bianca Meneses Portela (process: #124672/2024-8). The Coordination for the Improvement of Higher Education Personnel (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, CAPES) funded Luciana Manfrim (Financial code 001).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The datasets generated and/or analyzed during the current study are accessible as anonymized data at 10.6084/m9.figshare.28851110.