
Abstract
mRNA, either with unmodified or modified nucleosides, has become an important modality in medicines and vaccines. Despite the inclusion of N1-methyl-pseudouridine (N1mѰ)-modified mRNA in approved coronavirus 2019 (COVID-19) vaccines and the various ongoing clinical studies with both N1mѰ-modified and unmodified mRNA therapeutics, there are limited nonclinical data directly comparing unmodified and N1mѰ-modified mRNA safety and tolerability. Here we present data from a repeat-dose toxicity study in Wistar Han rats, which were intramuscularly administered mRNA (either with unmodified or N1mѰ-modified nucleosides) encapsulated into lipid nanoparticles (LNPs). The study evaluated systemic toxicity, local tolerance, and immune responses after once-weekly intramuscular injections for 3 weeks, followed by a final injection 2 weeks later. Additionally, a subgroup of animals was maintained for a 2-week dose-free period following the last dose to assess recovery from any findings. Repeat IM administration of unmodified and N1mѰ-modified mRNA rabies vaccine to male and female Wistar Han rats was well tolerated at doses of 10 μg and 40 μg, with similar non-adverse findings irrespective of the test article used, confirming that the N1mѰ nucleoside modification does not impact the nonclinical safety of mRNA when encapsulated into the same LNP. As the field of RNA medicines evolves and matures, the ability to use existing knowledge of unmodified or N1mѰ-modified mRNA to support mRNA therapeutic development in a nonclinical safety context will allow for a more streamlined and rapid process when developing or improving mRNA therapeutics.
Keywords
Introduction
Messenger ribonucleic acid (mRNA)-based agents have a wide range of potential applications, including prophylactic vaccines, therapeutics, and diagnostic tools, and they are currently under investigation for a variety of indications including infectious disease, oncology, and metabolic diseases.1-3 In recent years, the field of vaccine development has seen significant progress, particularly with the advance of mRNA prophylactic vaccine technology following the coronavirus 2019 (COVID-19) pandemic, for which 2 mRNA vaccines were approved.2,4 The possibility of rapidly developing mRNA vaccines using existing knowledge and/or platform-approach technology has paved the way to bring much-needed medicines to patients more rapidly, as reflected by the approval of an mRNA-based Respiratory Syncytial Virus vaccine in 2024. 5 Indeed, emerging guidelines on the use of vaccine platform technology and prior knowledge approaches from the United States Food and Drug Administration (US FDA), European Medicines Agency (EMA), Medicines and Healthcare products Regulatory Agency (MHRA), and World Health Organization (WHO)6-9 are laying the groundwork for a more streamlined development path of mRNA medicines and vaccines. The highly modular nature of mRNA and the ability for rapid improvement and development underscore the role of mRNA in shaping the future of modern medicine.
Although the immunostimulatory nature of mRNA is considered a key strength for vaccine development, it also presents a challenge. 10 Exogenously delivered mRNA synthesized by in vitro translation is recognized by innate immune receptors in the body, producing inflammatory cytokines and type I interferons that suppress cellular mRNA translation and increase mRNA degradation, resulting in decreased expression of the mRNA-encoded protein. 11 A number of strategies have been employed to optimize mRNA pharmacology, including chemical base modifications of mRNA to overcome immunogenicity challenges and increase mRNA translation efficiency. 10 The groundbreaking impact of introducing modified ribonucleotide base to the mRNA sequence has been recognized as a pivotal advancement in designing mRNA medicines and vaccines, as evidenced by the awarding of the 2023 Nobel Prize in medicine for this discovery. 12
A ribonucleoside modification used in marketed mRNA vaccines is N1-methylpseudouridine (N1mѰ). 11 Pseudouridine (Ѱ) and N1mѰ are naturally occurring post-transcriptional modifications of uridine found in cellular RNAs, such as tRNA and 18S ribosomal RNA. 13 Ѱ is formed in an enzymatic reaction converting the N1-C1’ glycosidic bond between the uracil base and the ribose to a C5-C1’ bond, a further enzymatic methylation at the N1 position of the uracil base of Ѱ leads to the formation of N1mѰ. 13 These modifications have been successfully adopted for synthetic mRNA applications due to their favorable immunological and translational properties.14-16
Replacing uridine with Ѱ or N1mѰ reduces mRNA immunogenicity by limiting the activation of pattern-recognition receptors including toll-like receptors (TLR) 3, TLR7 and TLR8, the retinoic acid–inducible gene I protein (RIG-I) pathway protein kinase R, and 2’-5’- oligoadenylate synthetase.14,17-19 Ѱ-modification also reduces RNase L-mediated degradation and improves mRNA stability, leading to increased translation capacity.16-18 N1mѰ-modified mRNA was shown to further enhance synthetic mRNA properties by further evading the innate immune response and thus improving cellular viability and increase protein expression in vitro and in vivo, respectively, compared with Ѱ-modified mRNA. 14 The utilization of N1mѰ-modified mRNA in approved COVID-19 vaccines has been demonstrated to be effective, safe, and tolerable in humans enabling the use of higher vaccine doses whilst retaining improved reactogenicity profiles when compared with unmodified mRNA vaccine.2,20-22
However, not all mRNA medicines utilize modified nucleosides as there are various cancer, protein, and cellular immunotherapies in the clinic evaluating unmodified mRNA.2,23 It is possible that during the development of these immunotherapies, the mRNA may be optimized by changing the nucleosides from unmodified to modified or vice versa.
Therefore, understanding the safety profile of modified and unmodified mRNA vaccines in a nonclinical setting is an important step in the development of safe and effective mRNA-based therapeutics and vaccines. Despite promising results from nonclinical studies of unmodified and N1mѰ-modified mRNA vaccines against SARS-CoV-2,20,24-26 there is currently a lack of systematic characterization of the safety and local tolerability of unmodified mRNA vs N1mѰ-modified mRNA vaccines in a single nonclinical toxicity study.
To address this knowledge gap, a repeat-dose toxicity study in rats was conducted using an mRNA construct with or without N1mѰ-modified nucleosides, encapsulated into identical lipid nanoparticles (LNPs). The aim of the study was to evaluate the systemic toxicity, local tolerance, and immune response to N1mѰ-modified or unmodified investigational mRNA encoding rabies virus envelope glycoprotein (RABV-G) following repeated intramuscular (IM) administration over a 30-day dosing period. In addition, the potential for delayed onset and/or reversibility of findings was evaluated in a 2-week recovery period.
Materials and Methods
Compliance and Ethical Considerations
This repeat-dose toxicity study was conducted in accordance with the US Department of Health and Human Services, FDA, US Code of Federal Regulations (CFR), Title 21, Part 58: Good Laboratory Practice (GLP) for Nonclinical Laboratory Studies, 27 and as accepted by regulatory authorities throughout the European Union (Organisation for Economic Co-operation and Development (OECD) Principles of Good Laboratory Practice), Japan (Ministry of Health, Labour, and Welfare), and other countries that are signatories to the OECD Mutual Acceptance of Data Agreement. 28 All animal housing conditions and experiments were conducted in strict accordance with the National Research Council Guide for the Care and Use of Laboratory Animals (Eighth Edition, 2011) 29 and all applicable sections of the Office of Laboratory Animal Welfare Public Health Service Policy on Humane Care and Use of Laboratory Animals (NIH Publication No. 15-8013, revised 2015). 30 The study protocol was reviewed and approved by the Testing Facility (Charles River Laboratories; Ashland, Ohio) Institutional Animal Care and Use Committee.
Animals and Husbandry
Male and female Wistar Han rats (obtained from Charles River Laboratories, Raleigh, North Carolina) were group housed (2-4 animals of the same sex and dosing group) in environmentally controlled rooms with a mean temperature between 20°C and 26°C, relative humidity of 30%-70%, and 12-hour light and dark cycles (except during designated procedures). The rats were allowed to acclimatize to study housing conditions for ≥7 days before the study began. At dose initiation, rats were 8-9 weeks old and weighed 169-263 g (males) or 121-186 g (females). All rats received a complete diet (LLC LabDiet Certified CR Rodent Diet 5CR4, PMI Nutrition International) and had access to treated tap water suitable for drinking; both were provided ad libitum. Only clinically healthy animals, as determined during the acclimation period, were included in the study.
Test and Control Articles
The test articles consisted of unmodified (F12311) and N1mѰ-modified (F12312) mRNA encoding the rabies virus glycoprotein G (RABV-G), hereon referred to as unmodified and N1mѰ-modified mRNA, respectively. Both test articles had the same non-coding mRNA elements, were manufactured in the same facility, using the same manufacturing process, and were encapsulated into the same 4 lipids (ALC-0315, 1,2-distearoyl-sn-glycero-3-phosphorylcholine, cholesterol, and ALC-0159, Acuitas Therapeutics, Canada) at the same ratios to form LNPs.
Dose formulations were prepared by diluting test article stock solutions at 0.96 (unmodified) and 0.97 (N1mѰ-modified) g/L mRNA with sterile saline solution (0.9% sodium chloride for injection, US Pharmacopeia (USP)) to final concentrations of 10 and 40 µg/dose with no adjustments for purity. Dose formulations were prepared fresh on the day of administration under RNase-free conditions and were used within 2 hours of preparation. Sterile saline solution was used as the vehicle control. Dose formulation analyses were conducted and confirmed that all animals received at least the respective protocol-specified dose.
Study Design
Repeat-Dose Toxicity Study in Rats: Experimental Design
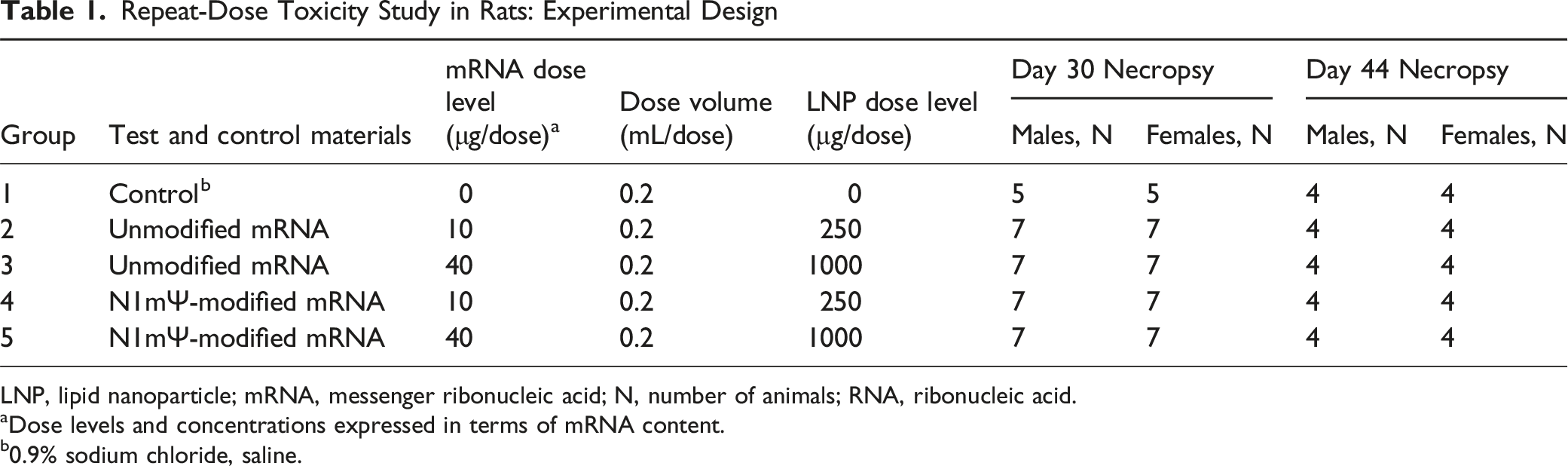
LNP, lipid nanoparticle; mRNA, messenger ribonucleic acid; N, number of animals; RNA, ribonucleic acid.
aDose levels and concentrations expressed in terms of mRNA content.
b0.9% sodium chloride, saline.
All animals received 4 intramuscular (IM) injections of either vehicle control (0.9% sodium chloride) or test article at a dose volume of 0.2 mL. Vehicle control of 0.9% sodium chloride was used as the comparator to the test articles, as LNPs can behave differently depending on whether they contain cargo due to their physicochemical properties including biodistribution, cellular uptake, or immune profile. 31 IM injections were administered once weekly for 3 weeks (Days 1, 8 and 15), followed by a final injection 2 weeks later (Day 29). Injections were administered into the right or left posterior lateral thigh (bicep femoris), using an appropriately sized syringe and needle. Injection sites were rotated each respective week of dosing. To ensure accurate dosing of animals, dose administration was conducted in group order, beginning with the control group and proceeding from low to high doses. Dose formulations for each test article and dose levels were uniquely barcoded. For each administration, the animal identification (ID) chip was scanned and electronically recorded against the corresponding formulation barcode to ensure correct dose-to-animal assignment. Twenty-four hours after the last injection (Day 30), 7 animals/sex/group (5 animals/sex for the control group) were euthanized, while the remaining 4 animals/sex/group were followed for a 2-week recovery period prior to being euthanized on Day 44 (Table 1). Euthanasia was performed via carbon dioxide inhalation.
In-Life Observations
Daily health observations were carried out 1-3 hours post-dose and at least once daily on non-dosing days. Detailed clinical observations were recorded within 4 days of arrival to the test facility, on Day −2, on Day 1 prior to dosing, at least once weekly during the study period, and on the day of necropsy. Measurements were generally performed in group order starting with Group 1. Individual body weights and the calculated body weight gains were recorded at least once on each day of dosing (unfasted, prior to dose administration on Days 1, 8, 15, and 29), daily for 3 consecutive days following each dosing day, on Days 22, 25, 36, 39, and 43, and finally on the day of scheduled necropsy (Day 30 and Day 44, fasted weight). Food consumption was measured twice weekly. Body temperature was collected using a Bio Medic Data Systems (BMDS) transponder implanted subcutaneously in the dorso-scapular region on each day of dosing (prior to dose administration) and at approximately 2-, 4-, 6-, 12-, 24-, and 48°h post-dose. Body temperature measurements continued daily if an animal had a temperature ≥39°C and continued until the value was below 39°C.
Local tolerance at the injection sites was assessed using the Draize scoring method 32 on each day of dosing, prior to the dose, and approximately 6-, 24- and 48 h post-dose. If a score of ≥1 was noted for either erythema or edema, scoring continued daily until the findings were resolved.
Hematology, Coagulation, Clinical Chemistry, and IgG
Hematology, coagulation, and clinical chemistry were evaluated on Day 30 (end of the dosing period) and Day 44 (end of the recovery period). Hematology (0.5 mL) and clinical chemistry (1 mL) blood samples were collected from the jugular vein by venipuncture. Hematology blood samples were collected in K2EDTA tubes and analyzed using ADVIA 2120i blood hematology analyzer. Clinical chemistry samples were collected without anticoagulant, processed in serum separating tubes, and the resultant serum analyzed using ADVIA 1800 clinical chemistry analyzer. Coagulation blood samples (1.8 mL) were collected from the vena cava after euthanasia into sodium citrate tubes and analyzed using STAGO STA Compact Max analyzer. Finally, blood samples (0.5 mL) were collected once during acclimation and on Day 30 to be processed to serum for the detection of anti-rabies antibodies.
Anti-rabies virus protein antibodies were measured using a validated enzyme-linked immunosorbent assay (ELISA). The positive control consisted of a mouse anti-rabies virus glycoprotein monoclonal antibody (Clone 1112-1, Millipore Sigma) spiked into pooled rat serum (BioIVT), and the negative control consisted of pooled rat serum to represent the background signal of the assay. Nunc Maxisorp 96-well clear plates (Thermo Fisher Scientific) were coated overnight with inactivated rabies virus (RABV) protein (Biorbyt), and control wells were coated with rat gamma globulin (Ig, Jackson ImmunoResearch) and then blocked with EZ Block (ScyTek Laboratories). On the day of the assay, controls and immunogenicity samples were diluted at a minimum required dilution of 50-fold in EZ Block. EZ Block was added to the control wells coated with rat Ig. Goat Anti-Rat IgG, Fcγ fragment-specific conjugated to peroxidase and Goat Anti-Mouse IgG, and Fcγ fragment-specific conjugated to peroxidase (both from Jackson ImmunoResearch) were added simultaneously to bind to anti-rabies virus protein antibodies captured on the plate by RABV protein. Absorbance signal (OD) was detected using a SpectraMax M5 or M2 Plate Reader. Positive samples were determined using a plate-specific cut point (PSCP) and were titered.
Necropsy
Necropsies were conducted by rotating through dose groups so similar numbers of animals from each group, including controls, were necropsied throughout the day. Blood for clinical pathology was collected in the same order on days of necropsy. All animals were fasted overnight prior to necropsy. Animals underwent complete necropsy examination, which included evaluation of the carcass, all external surfaces and orifices, cranial cavity, external surfaces of the brain, and thoracic, abdominal, and pelvic cavities with their associated organs and tissues. A standard organ panel (OECD Test no. 407) 33 was collected, weighed, and evaluated. Paired organs were weighed together. Organ to body weight and organ to brain weight ratios were also calculated.
Representative samples of tissues (artery (aorta), bone marrow (sternum), bone (femur and sternum), brain, cervix, epididymis, esophagus, eye, gland (adrenal, Harderian, mammary, parathyroid, pituitary, prostate, submandibular salivary, seminal vesicle, thyroid), gut-associated lymphoid tissue, heart, femorotibial joint, kidney, cecum, colon, rectum, liver, lung, lymph node (inguinal draining the administration site, mandibular, and mesenteric), quadriceps muscle, nerves (optic and sciatic), ovary, pancreas, intramuscular injection site, skin, duodenum, ileum, jejunum, spinal cord (cervical, lumbar and thoracic), spleen, stomach, testis, thymus, tongue, trachea, ureter, urinary bladder, uterus, vagina) were collected and preserved in 10% neutral buffered formalin, except for eyes and optic nerves which were preserved in Davidson’s fixative, and testes and epididymides which were preserved in modified Davidson’s fixative. All collected tissues were further processed and stained with hematoxylin-eosin for histopathology evaluations by a board-certified veterinary pathologist.
Statistical Analysis
An a priori sample size calculation was not conducted. Sample size was selected based on GLP standard toxicology guidelines. 9 Body weight, body weight gain, food consumption, body temperature, hematology, coagulation and clinical chemistry parameters, IgG titer, absolute, and relative organ weights were analyzed using Levene’s test to assess the homogeneity of group variances. Homogenous data was consequently analyzed using a one-way ANOVA F-test, and non-homogenous data was analyzed using Kruskal-Wallis test. Pairwise comparisons were performed for homogenous and non-homogenous data using a two-sided t-test or Wilcoxon Rank Sum Test, respectively, with adjustments for multiplicity based on the square root of the number of pairwise comparisons. All statistical tests were conducted at the 5% significance level. All pairwise comparisons were conducted at 1% and 5% significance levels. Groups with less than 3 observations were excluded from analysis. Means, standard deviations (or % coefficient of variation or standard error, when deemed appropriate), ratio, percentages, numbers, and/or incidences are reported as appropriate by dataset. SAS® software (v9) was used to perform statistical analyses.
Results
In-Life Observations
Overall, the administration of both unmodified and N1mѰ-modified mRNA vaccines was well tolerated at 10 and 40 µg doses with no test article-related clinical observations that were considered adverse. A swollen hindlimb leading to limited usage of that leg and abnormal gait was observed in 1 male administered 40 µg unmodified mRNA 1 day after the second administration, which was resolved by the next day. No similar findings were observed for any other animal or day. Skin scabs on the hindlimbs were noted in a small number of animals with similar frequency in the control and treated groups and were attributed to the IM injection procedure. All animals survived until their scheduled euthanasia.
Body Weight Gain and Food Consumption
A summary of mean body weight gain comparing unmodified vs N1mѰ-modified mRNA at the same dose along with the control group is provided in Figure 1. Generally, on the day following each vaccination, body weight gain was transiently reduced in both sexes in all groups, including vehicle control animals. However, overall, males receiving the unmodified mRNA exhibited significantly (P ≤ 0.05) dose-related lower mean body weight gain after each administration (Days 2, 9, 16, and 30) compared with the control or the corresponding N1mѰ-modified mRNA groups. However, these differences in body weight gain reductions in males at 40 µg were not statistically significant on Day 30 between the 2 test articles. Furthermore, during the intervals from Day 1 to Day 29 or Day 29 to Day 43, there were no marked differences compared to control or between test articles in body weight gain with the exception of males administered 40 µg unmodified mRNA, which showed a significant (P ≤ 0.05) reduction in body weight gain during the Day 1 to Day 29 period compared with the control group. Following the last dose, in males administered 40 µg unmodified or N1mѰ-modified mRNA, the reduced body weight gain after each administration throughout the study resulted in lower body weights (unfasted) on Day 29 (6.2% or 3.4%, respectively). These differences in final body weights were not statistically significant among the 2 test articles or vs the concurrent control. Mean body weight gain of male and female animals The mean unfasted body weight gain of animals is plotted following each administration of (A) 10 µg of unmodified or modified mRNA vaccine and the control group in males; (B) 40 µg of unmodified or modified mRNA vaccine and the control group in males; (C) both dose levels of unmodified and modified mRNA vaccine and the control group in males; (D) 10 µg of unmodified or modified mRNA vaccine and the control group in females; (E) 40 µg of unmodified or modified mRNA vaccine and the control group in females; and (F) both dose levels of unmodified and modified mRNA vaccine and the control group. The syringe reflects each day of administration, where body weight measurements were obtained prior to dose administration. Fasted body weight gains were excluded from these plots on study Days 30 and 44, designated by * on the X-axis. Error bars represent standard deviation, N = 9 (control group, Day −2 to Day 29), N = 11 (10 µg and 40 µg unmodified and modified mRNA administered groups, Day −2 to Day 29), and N = 4 (all groups Day 31 to Day 43)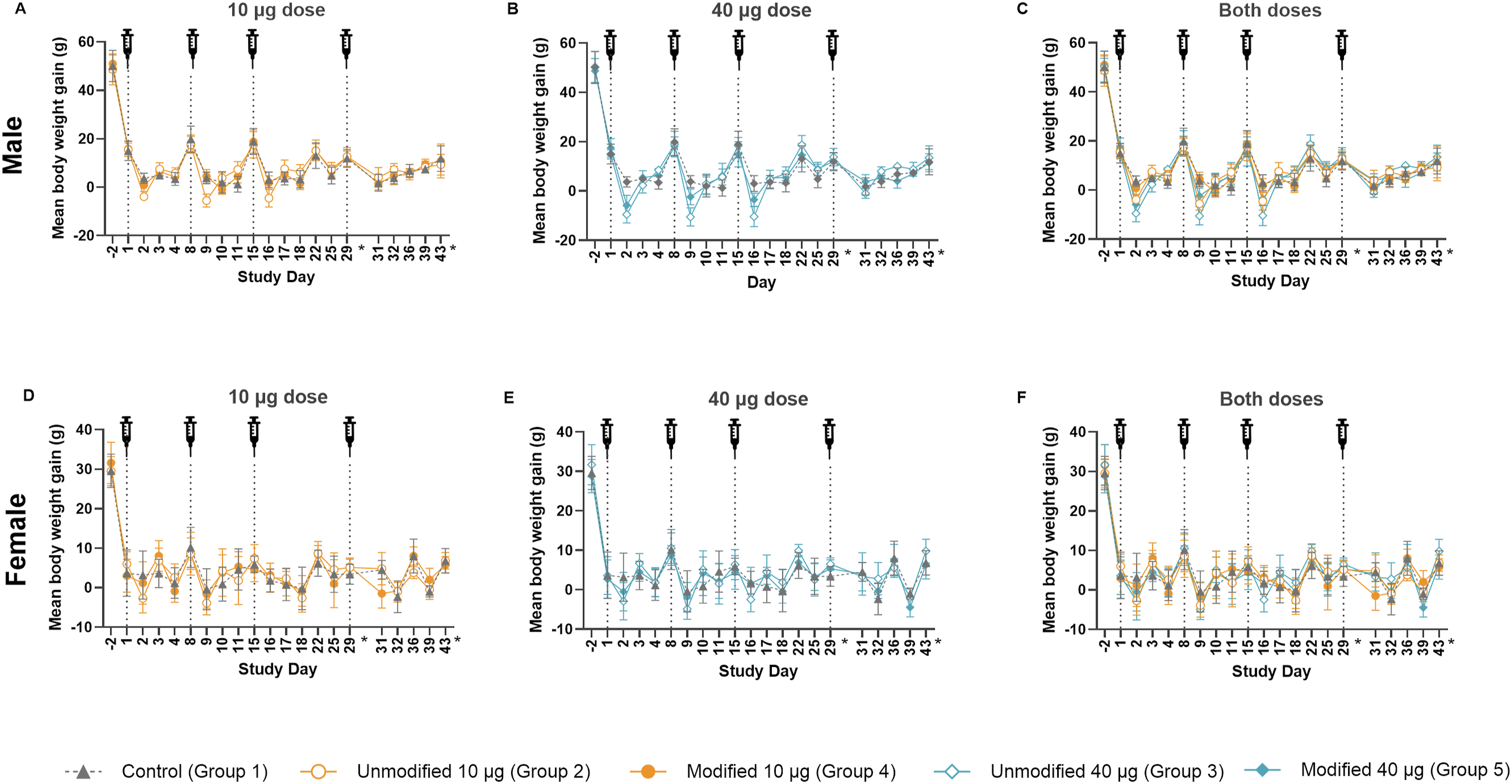
Body weight gain in females receiving unmodified or N1mѰ-modified mRNA appeared to be less affected regardless of the dose. Any changes noted in body weight gain in female groups generally recovered by the next day following dosing.
At the end of the recovery period (Day 43), body weight gain (Day 29 to 43) was generally similar to controls across both sexes administered unmodified or N1mѰ-modified mRNA, irrespective of the dose.
The observed reductions in body weight gain appeared to be dose-dependent, test-article, and/or injection procedure-related and were more pronounced in males receiving unmodified mRNA, indicating a slightly higher impact of the unmodified compared with the N1mѰ-modified mRNA, with effects largely resolving by the end of the recovery period.
Food consumption immediately following each vaccination was generally lower in all mRNA groups compared with the control group, although not statistically significant, which generally corresponded with decreased body weight gain in respective groups. Among the mRNA administered groups, mean food consumption following the second injection of the unmodified mRNA was significantly lower compared with N1mѰ-modified mRNA administration in males at both 10 and 40 µg (P ≤ 0.05) and in females at 10 µg (P ≤ 0.05). Exceptions were males administered 10 µg N1mѰ-modified mRNA, females administered 10 µg N1mѰ-modified mRNA after the first 2 injections, and males administered 40 µg N1mѰ-modified mRNA after the second and fourth administration, which had higher food consumption compared with control and in line with no observed decrease in body weight gain.
Overall, the transient reductions in food consumption appeared to be procedural, likely related to the test article, and were slightly more pronounced following unmodified compared with N1mѰ-modified mRNA.
Body Temperature
To evaluate the reactogenicity following the administration of unmodified or N1mѰ-modified mRNA at 10 or 40 µg, the body temperature was measured prior to dose administration and was followed for 48 hours post-administration (Figure 2). Mean body temperature increased in males administered 10 (Figure 2A) or 40 µg (Figure 2B) with either unmodified or N1mѰ-modified mRNA following each injection compared to controls, starting as early as 4 hours post-administration. Males which received 10 µg of either test article exhibited increased mean body temperature similarly with controls and resolved by 24 h. Statistically higher mean body temperature compared to the control animals was measured in males administered 40 µg unmodified mRNA following each administration (P ≤ 0.01) starting as early as 6 h post-administration and remaining elevated through 24 h post-dose and resolving by 48 h. In comparison, males administered 40 µg N1mѰ-modified mRNA vaccine exhibited shorter periods of increased mean body temperature compared to control animals, usually between 12 and 24 h (P ≤ 0.01) and resolving by 48 h following each administration. Mean body temperature in the first 48 h following each administration The mean body temperature is plotted prior (pre) and following each administration of (A) 10 µg of unmodified or modified mRNA vaccine and the control group in males; (B) 40 µg of unmodified or modified mRNA vaccine and the control group in males; (C) both dose levels of unmodified and modified mRNA vaccine and the control group in males; (D) 10 µg of unmodified or modified mRNA vaccine and the control group in females; (E) 40 µg of unmodified or modified mRNA vaccine and the control group in females; and (F) both dose levels of unmodified and modified mRNA vaccine and the control group in males. The syringe reflects each day of administration. The horizontal dotted lines designate the minimum and maximum mean body temperature for the male (35.91°C and 37.77°C) and female (35.83°C and 37.93°C) control animals. Error bars represent standard deviation, N = 9 (control group), N = 11 (10 µg and 40 µg unmodified and modified mRNA administered groups), and N = 4 (all groups at 48 h after fourth administration)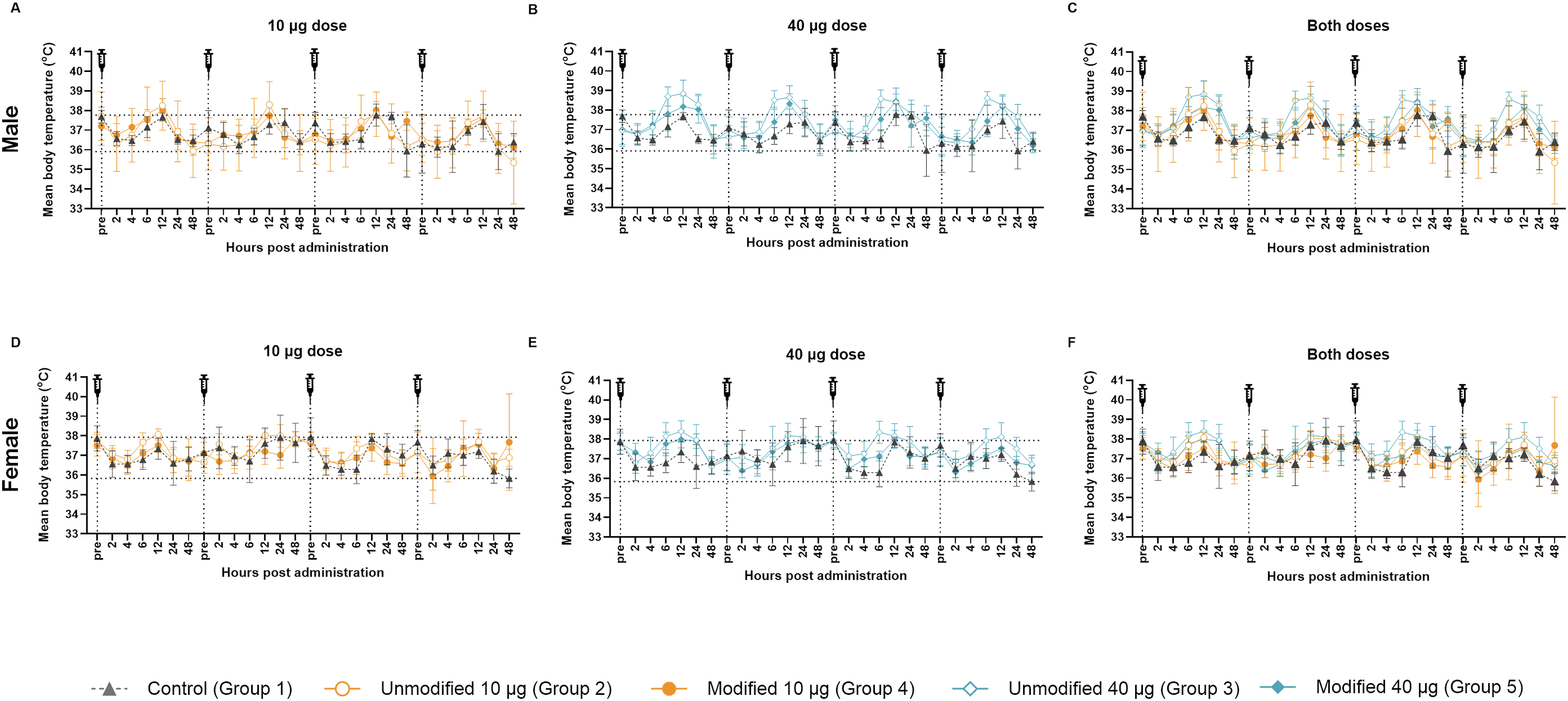
Comparable test article–related effects were observed in females administered 10 (Figure 2D) or 40 µg (Figure 2E) with unmodified or N1mѰ-modified mRNA, although the magnitude of body temperature changes in females was generally lower compared to males. In females at 10 µg of either mRNA, elevated temperatures were in the range of the control animals and typically resolved by 24 h post-dose. Whereas in the 40 µg groups, temperatures were higher compared to the control mainly following the first administration (P ≤ 0.01 both unmodified and N1mѰ-modified mRNA), and to a lesser extent following the third (P ≤ 0.01, unmodified and P ≤ 0.05, N1mѰ-modified mRNA) and fourth (P ≤ 0.01, unmodified only) injections, and generally remained elevated at 24 h and resolved by 48 h. Following the second administration of 40 µg unmodified or N1mѰ-modified mRNA to females, body temperatures were comparable to control animals indicating that the elevated temperature observed was likely procedure-related and not due to the test article.
The mean body temperatures were consistently higher (P ≤ 0.05) in the unmodified groups compared to their respective dose-matched N1mѰ-modified group particularly in males at 40 µg measured 6 hours following each administration. Females administered 40 µg unmodified mRNA were characterized with a higher (P ≤ 0.05) body temperature compared to N1mѰ-modified only at 2 h after the second administration and 6 h after the third administration. Following the administration of 10 µg mRNA, males did not show any difference in body temperature increase, while females exhibited a higher (P ≤ 0.05) body temperature in the unmodified groups compared to N1mѰ-modified 12 h following the first administration and 12 and 24 h following the second administration. It should also be noted that a lower (P ≤ 0.05) body temperature in the unmodified groups compared to their dose-matched N1mѰ-modified groups was observed 48 h after the third administration in males administered unmodified mRNA at 10 µg and at 24 h after the fourth administration in females at 40 µg. The mean body temperature changes between unmodified and N1mѰ-modified were comparable for all other timepoints.
In summary, the mean body temperatures measured in both males and females were highly variable including in controls. In animals administered 40 µg with either test article, statistically higher average body temperatures were noted compared to control animals, demonstrating a dose-related effect. Overall, females exhibited a less pronounced increase in body temperature. Animals administered the unmodified mRNA vaccine had an earlier elevation in body temperature than those receiving the N1mѰ-modified mRNA, which was demonstrated at 6 h after each administration with unmodified mRNA compared to N1mѰ-modified mRNA in males and resulted in a longer duration of elevated temperature. Nevertheless, the increased body temperature returned to baseline by 48 h for both the unmodified and N1mѰ-modified mRNA groups.
Local Tolerance
Injection sites were evaluated for dermal changes as part of local tolerance, which can act as indicators of reactogenicity in IM-injected vaccines. Dermal observations included edema and erythema noted at the injection sites in groups administered unmodified and N1mѰ-modified mRNA when compared to control animals and exhibited a dose-dependency in occurrence, duration, and severity (Figure 3). However, there were no consistent differences in dermal observations between the unmodified and N1mѰ-modified mRNA groups. Erythema was observed more frequently in females than males. Dermal observation scores of edema and erythema plotted following each administration and by day post-administration. Edema and erythema at the injection sites were monitored for 5 days after each administration. Prior to administration, the injection site was clear of dermal events. The aggregated severity scores are plotted: (A) Number of edema observations following each vaccine administration (Days 1, 8, 15, and 29); (B) number of edema observations by day post-administration for the first three administrations (Days 1, 8, and 15); (C) number of erythema observations following each vaccine administration (Days 1, 8, 15, and 29); (D) number of erythema observations by day post-administration for the first three administrations (Days 1, 8, and 15)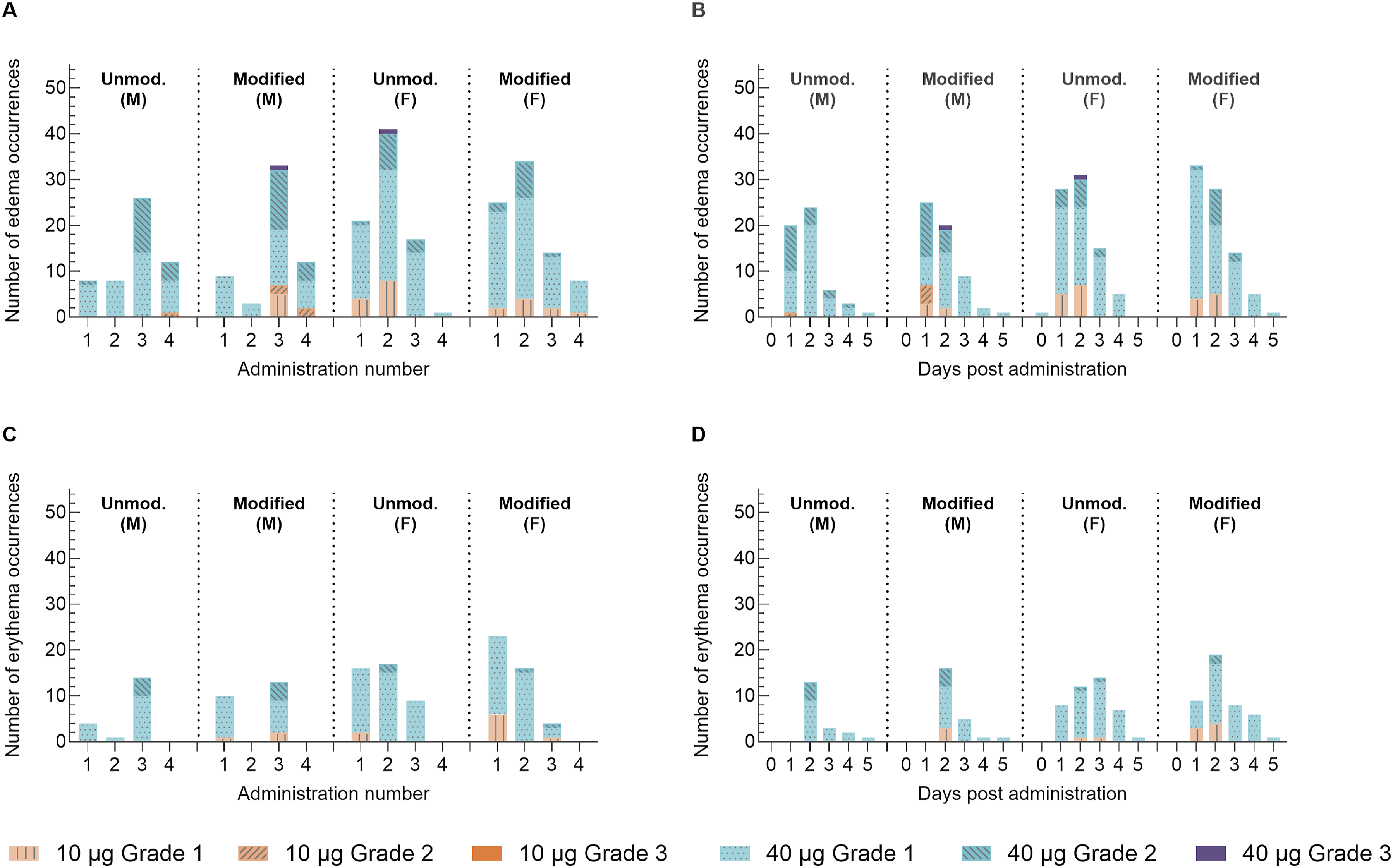
Edema and erythema severity scores at injection sites were generally Grade 1 or 2. The two Grade 3 edema incidences reported were in 1 female 2 days after the second administration of unmodified mRNA at 40 μg and in 1 male 2 days after the third administration of N1mѰ-modified mRNA at 40 μg (Figure 3A, B). No incidences of erythema Grade 3 or above were reported.
Edema and erythema events generally resolved within 3 days post-administration of 10 μg mRNA and within 6 days post-administration of 40 μg mRNA, in line with a dose-related effect. Injection sites appeared normal prior to each dose administration.
Hematology, Coagulation, Clinical Chemistry, and IgG
The data described herein are those that were considered test article-related changes in hematology, coagulation, clinical chemistry, and IgG.
Hematology
Test article-related hematology changes compared to controls were noted in neutrophil counts, lymphocyte counts, monocyte counts, eosinophil counts, white blood cell (WBC) counts, gated red blood cell distribution width (RDWG), and platelet counts (Table S1).
On Day 30, neutrophil counts were significantly (P ≤ 0.05) increased in all mRNA groups vs the control group. Increases were significantly (P ≤ 0.05) higher in animals administered 10 µg unmodified mRNA (7x in males and females) vs 10 µg N1mѰ-modified mRNA (3x and 4.4x in males and females, respectively). At 40 µg doses, the increase (∼6x) in neutrophil counts was comparable for unmodified and N1mѰ-modified mRNA. Conversely, lymphocyte counts were statistically significantly (P ≤ 0.05) decreased compared with controls in females administered 10 µg unmodified mRNA (43%), both sexes administered 40 µg unmodified mRNA (43%-48% males and females, respectively), and females administered 40 µg N1mѰ-modified mRNA (30%). Although minimal, monocyte counts were significantly elevated in the 10 µg N1mѰ-modified mRNA groups (1.6x and 1.7x in males and females, respectively) compared with controls (P ≤ 0.05). Finally, eosinophil counts were significantly (P ≤ 0.05) increased in the female unmodified mRNA groups (10 and 40 µg; 2.3x and 2.8x, respectively) compared with the control group.
The combination of these changes resulted in overall higher total WBC counts on Day 30 in all vaccinated groups vs control. Nevertheless, statistical significance (P ≤ 0.05) compared with the controls could only be demonstrated for males receiving unmodified mRNA (10 and 40 µg) and 40 µg N1mѰ-modified mRNA. Moreover, the increase in WBC count was significantly (P ≤ 0.05) higher in males receiving 10 µg unmodified (1.8x) vs males receiving 10 µg N1mѰ-modified mRNA (1.2x).
On Day 30, RDWG was also increased for all groups vs control (P ≤ 0.05), except for animals administered 10 µg N1mѰ-modified mRNA. RDWG increases were significantly (P ≤ 0.05) higher in the female unmodified mRNA groups (10 and 40 µg; 1.1x and 1.3x, respectively) and the male unmodified mRNA 10 µg group (1.1x) compared with their respective N1mѰ-modified groups. Platelet counts at Day 30 were decreased in animals receiving unmodified mRNA at 40 µg (18%) relative to the control group, which was statistically significant for males (P ≤ 0.05). Taken together, the changes in hematology parameters were minimal and thus considered non-adverse based on their magnitude and lack of any corresponding findings.
After the 2-week recovery period, all WBC and platelet count changes were completely resolved, except for neutrophil counts in females administered N1mѰ-modified mRNA at 10 µg (1.8x) and eosinophil counts in females administered unmodified mRNA at 10 and 40 µg (2.2x and 2.1x, respectively), which showed evidence of recovery. The increase in RDWG was only partially resolved and still significantly higher compared with the control group on Day 44 (P ≤ 0.05).
Coagulation
There were no significant changes in prothrombin time; however, the activated partial thromboplastin time (APTT) was statistically significantly elevated in the 10 and 40 µg unmodified males on Day 30 vs control (1.2x, P ≤ 0.05; Table S2). The fibrinogen concentration was significantly (P ≤ 0.05) increased for all groups on Day 30 vs control, with a higher increase for the unmodified mRNA groups vs the N1mѰ-modified mRNA groups (significant P ≤ 0.05 for the 10 µg males and 40 µg males and females). At the end of the recovery period, all coagulation changes were fully resolved.
Clinical Chemistry
Clinical chemistry changes compared with controls were noted in total protein, albumin, globulin, albumin/globulin ratio (A/G), glucose, triglyceride, calcium, and chloride concentrations (Table S3). On Day 30, a small non-statistically significant decrease in total protein was observed in females administered N1mѰ-modified mRNA at 40 µg (4%), which resolved completely following the recovery period. Minimal changes were also observed in albumin and globulin levels, with only a statistically significantly lower albumin reported in females receiving 40 µg N1mѰ-modified mRNA (9%, P ≤ 0.05) on Day 30. The changes in albumin and globulin led to a significantly decreased A/G for both N1mѰ-modified and unmodified mRNA groups at 40 µg (range of 9%-13%, P ≤ 0.05). The decreased A/G generally resolved after a 2-week recovery period in males, but not in females administered 40 µg unmodified or N1mѰ-modified mRNA (10% and 15%, respectively). The persisting lower A/G was due to the significantly higher globulin levels on Day 44 for these groups compared with controls (1.07x and 1.15x, respectively, P ≤ 0.05). Glucose was slightly decreased (not statistically significant) across males and females in both unmodified and N1mѰ-modified mRNA 40 µg dose groups (11%-15%), which was fully resolved following the recovery period. Triglyceride was also decreased in males receiving 40 µg of unmodified mRNA (45%), with this difference being non-significant. Statistically significantly decreased calcium was noted on Day 30 in males and females receiving unmodified mRNA at 40 µg (6% and 4%, respectively) that did not resolve for females (P ≤ 0.05). Decreased chloride (P ≤ 0.05) was observed in females administered 40 µg unmodified mRNA (3%), which resolved completely following the recovery period.
IgG
Serum collected prior to dose initiation was negative for anti-rabies antibodies for all animals. On Day 30, all animals administered unmodified and N1mѰ-modified mRNA at 10 and 40 µg had positive titers for anti-rabies antibodies. Mean titers are given in Table S4.
Among animals administered unmodified mRNA, titers ranged from 5000 to 50,000 ng/mL and were similar between males and females at the same dose level, and between the 10 and 40 µg dose groups, with the exception of 1 male in the 40 µg dose group with a reported titer of 500 000 ng/mL.
Among animals administered N1mѰ-modified mRNA, titers also generally ranged between 5000 and 50,000 ng/mL. However, titers were generally greater for males compared with females and for the 40 µg vs 10 µg dose groups. Outliers included single instances of titers up to 500 000 ng/mL in a 40 µg male and female, a titer of 50,000 000 ng/mL in a 40 µg male, and a titer 500 ng/mL in a 10 µg female.
Necropsy
Organ Weights and Gross Pathology
There were no gross findings associated with the administration of either unmodified or N1mѰ-modified mRNA at necropsy. Organ weight changes relative to control animals at terminal euthanasia on Day 30 were noted in the spleen, liver, and kidney (Table S5)
There were no differences in mean absolute kidney weights or mean kidney weight relative to brain weight compared with control in any group. Nevertheless, mean kidney weight relative to terminal body weight was significantly (P ≤ 0.05) higher in males and females administered 40 μg unmodified mRNA, compared to control animals. Additionally, compared to its N1mѰ-modified mRNA counterpart, male kidney weight relative to terminal body weight was significantly (P ≤ 0.05) higher in males administered 40 μg unmodified mRNA at the same dose. There were no correlating meaningful changes in kidney function parameters measured as blood urea nitrogen (BUN) and creatinine in animals administered unmodified mRNA at any dose level. Both BUN and creatinine were within normal range of control values in animals that were administered unmodified mRNA. These markers of kidney function were also within normal range of control values in animals administered N1mѰ-modified mRNA.
There were no microscopic correlates to the increased mean organ weights, and organ weight differences were resolved after a 2-week recovery period. There were no other test article-related organ weight changes.
Histopathology
Microscopic findings following terminal euthanasia on Day 30 were limited to mixed cell inflammation and/or mononuclear cell infiltration in the IM injection sites, sciatic nerves, inguinal lymph node, and quadriceps muscle in test article-administered animals. There were no consistent differences in incidence or severity between both test articles. Across affected tissues, findings primarily consisted of mixed cell inflammation characterized by the presence of neutrophils (often degenerated) and mononuclear cells with or without eosinophils, frequently accompanied by eosinophilic extracellular material consistent with fibrin and expanded space within the tissue spaces indicative of edema. Mononuclear cell infiltration was characterized by the presence of lymphocytes and histiocytes without accompanying features of active inflammation.
Findings in the injection site muscle were similar in incidence and severity (ranging from minimal to moderate) across the 2 test articles with no apparent dose-response relationship. However, females appeared to have higher incidences compared with males, irrespective of the dose or the test article.
In the sciatic nerve, mixed cell inflammation (minimal to moderate) along with minimal mononuclear cell infiltration was perineural in distribution and was not associated with visible changes within the nerve. These findings occurred at a higher incidence in females in a dose- and test-article independent manner.
Minimal to mild mixed cell infiltration in the inguinal lymph node, noted in only 2 animals (1 male and 1 female) administered N1mѰ-modified mRNA at 40 µg, was characterized by the presence of neutrophils, eosinophils, and mononuclear cells without degenerate neutrophils and fibrin.
Inflammation around the sciatic nerve, in the quadriceps muscle, and in the lymph node was consistent with extension from the local inflammatory changes at the injection site. These findings did not result in tissue degeneration or necrosis, were generally expected for local reactions to vaccination, and were not considered adverse.
Following a 2-week recovery period, similar findings were noted in the same tissues, albeit at a reduced incidence and/or severity. The findings were often less acute, trending towards inflammatory cell infiltration, rather than inflammation. The inguinal lymph nodes in males and females administered 40 µg N1mѰ-modified or unmodified mRNA had a new finding of histiocytic infiltration, which was characterized by the presence of clusters of plump macrophages within the lymph nodes. This finding was considered to be due to progression of the immune response following the administration of a vaccine and thus was not considered adverse. No clear differences in incidence and severity could be identified between the modified and unmodified groups.
Discussion
Throughout mRNA drug discovery and development, the decision to use modified or unmodified nucleosides in the design of the mRNA may change, particularly depending on the indication. Generally, during the discovery stage, sequence and nucleoside modifications are considered in relation to the immunogenicity elicited before a candidate is selected for further nonclinical safety development. While toxicity endpoints might be included in pharmacology studies, the doses used in these studies are generally not high enough and therefore, direct and complete comparison between unmodified and modified mRNA with regards to nonclinical safety is not readily available. 34 Hence, the aim of the current study was to directly compare the nonclinical safety profiles of unmodified and N1mѰ-modified mRNA encoding RABV-G protein encapsulated into LNP.
To evaluate the systemic toxicity and local tolerance of unmodified and N1mѰ-modified mRNA, Wistar Han rats received 3 once weekly IM injections followed by 1 final IM injection 2 weeks later. Rats were used as they have been shown to be pharmacologically relevant in terms of rabies IgG production 35 and have been widely used in mRNA development. An observation period of 2 weeks after the last administration was included to evaluate delayed onset and/or reversibility of any findings. Overall, both the unmodified and N1mѰ-modified mRNA vaccines (10 or 40 µg doses) were well tolerated following 4 IM administrations. Moreover, the observed findings generally remained consistent across the 2 test articles, albeit slightly more pronounced in animals administered unmodified mRNA, and were in line with expected immune responses following the administration of exogenous mRNA encapsulated into lipids to form LNPs.36-38
In this study, a dose-related decrease in body weight gain was observed following each vaccination, particularly among male rats, with the effect being greater in unmodified vs the N1mѰ-modified mRNA groups. The decreased body weight gain coincided with decreased food consumption immediately following each dose. However, body weights recovered prior to each subsequent dose injection. There was an absence of any clinical or health observations associated with decreased body weight gain or food consumption. In addition, body weight gain rapidly rebounded within 2 days post-dose; therefore, the changes in body weight gain were considered non-adverse. No biologically relevant differences in body weight gain were observed during the recovery period, confirming that the decrease in body weight gain after each dose was related to the stress of the dosing procedure. Transient body weight decreases following mRNA-LNP is a well-documented and expected finding associated with systemic reactogenicity (inflammation, cytokine induction, and reduced appetite). These findings have been reported in rats, rabbits, and non-human primates following mRNA-LNP administration.36,37,39
Following each IM administration, a transient dose-related increase was noted in mean body temperature in both sexes with both unmodified and N1mѰ-modified mRNA. This was observed to peak at 6 or 12 hours post-dose and generally resolved by 24 or 48 hours post-dose for the 10 and 40 µg doses, respectively, indicating that the duration and magnitude of the body temperature increase were dose-related. This dose-related increase in mean body temperature was slightly more pronounced in the unmodified mRNA groups relative to the N1mѰ-modified mRNA when compared with the concurrent control group as well as the historical control database. However, based on the high variability (especially in females at the high dose) and the low magnitude of body temperature increase, these changes were not considered adverse.
Edema and erythema at the injection site were also reported following each dose with either unmodified and N1mѰ-modified mRNA, and similarly were dose-related, transient in nature, and resolved completely prior to the next IM administration. Edema was noted more frequently than erythema with both test articles. Erythema was noted with greater frequency in females when compared with males, likely due to the lower body weight in females and therefore smaller muscle mass than males. 40 The difference in muscle mass results in a greater local tissue burden per injected volume, therefore increasing the likelihood of local irritation and erythema. The observed local injection site reactions were reflected in the microscopic findings of inflammation and/or mononuclear cell infiltration in the intramuscular injection sites and quadriceps muscle with both test articles.
Changes noted in hematology with both unmodified and N1mѰ-modified mRNA were primarily related to an acute phase inflammatory response, supported by the microscopic findings at the intramuscular injection sites, sciatic nerve, lymph nodes, and/or quadriceps muscle. The increased RDWG in all unmodified groups and the high-dose N1mѰ-modified group indicated a greater variation in red blood cell size without concurrent changes in red blood cell mass, red blood cell indices, or reticulocytes. A dose-dependent, statistically significant increase in fibrinogen was observed particularly in the unmodified groups vs the N1mѰ-modified mRNA groups (P ≤ 0.05), indicating increased inflammation following administration with unmodified mRNA. Moreover, a dose-dependent increase in neutrophil counts in animals administered unmodified or N1mѰ-modified mRNA together with the decreased A/G ratio in the 40 µg unmodified and N1mѰ-modified groups further confirmed an acute phase inflammatory response. The positive titers for anti-rabies antibodies on Day 30 also corroborated the immune activation against the RABV-G protein with both test articles.
On Day 30, increased spleen and liver weights were noted for both males and females in the unmodified and N1mѰ-modified groups, with no microscopic correlates. Both the liver and spleen play crucial roles in the body’s immune response. The spleen filters blood and helps fight infections, while the liver produces acute-phase proteins, together forming the liver-spleen axis, which refers to the functional and anatomical relationship between the 2 organs, particularly in the context of immunity, infections, and metabolism. 41 Therefore, the observed organ weight changes were likely due to a stimulated immune response and thus were considered a non-adverse pharmacological response. Mean kidney weights relative to terminal body weight were significantly increased in both males and females administered 40 µg unmodified mRNA. In addition, kidney weight relative to terminal body weight was significantly (P ≤ 0.05) higher in males receiving 40 µg unmodified mRNA compared with N1mѰ-modified mRNA at the same dose. There were no microscopic correlates to the increased kidney weights, and no effects were observed on markers of kidney function (BUN and creatinine); therefore, these findings were not considered adverse. No other absolute organ weight changes were noted with either test article, and all observed changes resolved after the 2-week recovery period.
An acute inflammatory response to vaccination, including injection-site pain, swelling, and fever, is well described in humans. 42 The collective findings in body temperature, local injection site reactions, microscopic findings, body/organ weight, and clinical pathology were therefore considered consistent with, and expected, following vaccine administration. Taken together with the reversible or partially reversible microscopic findings, the no-observed-adverse-effect level following 4 IM administrations was determined to be 40 µg for both unmodified and N1mѰ-modified mRNA.
To the authors’ knowledge, this is the first study to directly compare the nonclinical safety profile of unmodified and N1mѰ-modified nucleosides of the same vaccine encapsulated into the LNP using the same lipids and administered at the same dose. Nucleoside modifications in mRNA vaccines primarily aim to enhance stability, reduce immunogenicity, and improve translation efficiency. 43 While the primary focus of these modifications is on the efficacy and safety of the mRNA therapeutic, there are indirect effects on lipid metabolism due to the LNP delivery system used in these vaccines. 43 Indeed, there is evidence that N1mѰ-modified mRNA has a better overall safety profile compared with unmodified mRNA based on the recent clinical experience with COVID-19 vaccines. 44 Nevertheless, there are many factors that contribute to the overall safety profile of mRNA therapies and the benefit-risk ratio differs between therapeutic areas. 45
It has been established that the LNP used to deliver the mRNA therapeutic can induce an immune response on its own and also impacts mRNA efficacy.43,46,47 Kauffman et al 48 demonstrated that pseudouridine modification in mRNA had no significant effect on the physical properties of LNP, in vivo protein expression, or mRNA immunogenicity compared with unmodified mRNA when delivered systemically with liver-targeting LNPs. Moreover, the transient extracellular innate immune response has been found to be similar between the pseudouridine modified and unmodified mRNA. Bernard et al 43 demonstrated similar findings in macaques and observed that the impact of nucleoside modification on mRNA vaccine efficacy varied with LNP composition, and not the nucleoside modification.
In nonclinical species, reactogenicity is measured through a multimodal assessment combining clinical observations, body weight, food consumption, body temperature, injection site reactions, and dermal scoring along with clinical pathology and histopathology. mRNA LNP constructs have been shown to produce transient inflammatory responses, aligning with known innate immune activation patterns.38,43 In line with the findings of Bernard et al 43 and Kaufmann et al, 48 the current study demonstrated that while the reactogenicity of the unmodified mRNA appears to be slightly higher compared with N1mѰ-modified mRNA, the overall safety profile is very similar. This is not surprising as modifications such as N1mѰ in mRNA vaccines help reduce the intrinsic immunostimulatory activity of mRNA and increase its translation efficiency.10,14 However, it is worth noting that this nonclinical study only evaluated 1 type of nucleoside modification and was limited to only 2 dose levels. In clinical settings, for example, in oncology, a greater number and frequency of doses may be required to elicit an immune response. It is important to note that in humans, the differences in lipid metabolism 49 and immune response50,51 between males and females may also influence reactogenicity. Therefore, while the nonclinical to clinical translation of reactogenicity is difficult, it may be prudent to evaluate different nucleoside modifications at possibly higher doses administered over a longer duration to further compare the nonclinical safety of unmodified vs modified mRNA in a single study.
Conclusion
In conclusion, repeated IM administration of an investigational mRNA rabies vaccine with unmodified or N1mѰ-modified nucleosides to male and female Wistar Han rats was well tolerated with no adverse findings at doses of 10 μg or 40 μg. While the findings were similar across the 2 test articles overall, effects in male and female rats administered unmodified mRNA were slightly more pronounced, consistent with higher reactogenicity. These data support the platform approach,6,9,52,53 where previously generated data from unmodified mRNA can be used to inform the nonclinical and clinical safety of N1mѰ-modified mRNA medicines, irrespective of the open reading frame, and vice versa, provided that the mRNA have the same noncoding elements and are encapsulated into the same LNPs.
Supplemental Material
Supplemental Material - Nonclinical Safety Profiles of mRNA Therapeutics Comprising Unmodified or N1-Methyl-Pseudouridine-Modified Nucleosides Are Similar Following Repeated Administration
Supplemental Material for Nonclinical Safety Profiles of mRNA Therapeutics Comprising Unmodified or N1-Methyl-Pseudouridine-Modified Nucleosides Are Similar Following Repeated Administration by Albena Draycheva, Joske Millecam, and Ebru Caba in International Journal of Toxicology.
Footnotes
Acknowledgments
Ethical Considerations
All animal housing and experiments were conducted in strict accordance with the National Research Council Guide for the Care and Use of Laboratory Animals (Eighth Edition, 2011) and all applicable sections of the Office of Laboratory Animal Welfare Public Health Service Policy on Humane Care and Use of Laboratory Animals (NIH Publication No. 15-8013, revised 2015). The study protocol was reviewed and approved by the Testing Facility (Charles River Laboratories; Ashland, Ohio) Institutional Animal Care and Use Committee.
Author Contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by CureVac SE. The Sponsor was involved in study conception and design, analysis and interpretation of the data, and the decision to submit the article for publication. The sponsor was also given the opportunity to review the manuscript for medical and scientific accuracy as well as intellectual property considerations.
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Albena Draycheva is an employee of CureVac Netherlands B.V. and may hold shares and/or stakes in the company. Joske Millecam is an employee of CureVac Belgium S.A. and may hold shares and/or stakes in the company. Ebru Caba is an employee of CureVac Inc. and may hold shares and/or stakes in the company.
Data Availability Statement
The datasets generated during the current study are not publicly available due to commercial reasons but are available from the corresponding author on reasonable request.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.