
Abstract
Background:
The unexpected observation of calretinin immunoreactivity in smooth muscle cells in the muscularis propria of the cecum led to a more detailed examination of calretinin expression and its possible relationship to propulsive contractile activity around the vermiform appendix.
Methods:
Immunohistochemistry and RNA in situ hybridization were performed to analyze calretinin expression in intestinal samples from 33 patients at ages ranging from mid-gestation fetuses to adults, as well as in some potentially relevant animal models. Dual immunolabeling was done to compare calretinin localization with markers of smooth muscle and interstitial cells of Cajal.
Results:
Calretinin expression was observed consistently in the innermost smooth muscle layers of the muscularis interna in the human cecum, appendiceal base, and proximal ascending colon, but not elsewhere in the intestinal tract. Calretinin-positive smooth muscle cells did not co-express markers located in adjacent interstitial cells of Cajal. Muscular calretinin immunoreactivity was not detected in the ceca of mice or macaques, species which lack appendices, nor in the rabbit cecum or appendix.
Conclusions:
Localized expression of calretinin in cecal smooth muscle cells may reduce the likelihood of retrograde, calcium-mediated propulsive contractions from the proximal colon and suppress pro-inflammatory fecal stasis in the appendix.
Introduction
Calretinin, also known as calbindin 2 (formerly “29 kd calbindin”), is an intracellular calcium-buffering protein that was discovered in 1987 after its cDNA was cloned from a chick retinal cDNA library. 1 Calretinin is a member of the EF-hand superfamily of calcium binding proteins. 2 The superfamily includes more than 200 proteins, which share structural and functional properties including multiple EF-hand domains, 29–30 amino acid-long regions that bind calcium. While not solely expressed in neurons, calretinin and the EF-hand-domain calcium-binding proteins, calbindin and parvalbumin, are present in the cytoplasm of discrete subtypes of neurons, where they appear to play a major role as intracellular calcium buffers. These buffers are important modulators of Ca++-mediated events in Leydig cells, 3 some neoplastic cell lines,4,5 and neurons, including long term potentiation in the hippocampus and granular cell excitability in the cerebellum.6,7
In the gastrointestinal tract, calretinin is expressed by mast cells, serosal mesothelial cells, and a subset of enteric neurons. Calretinin-specific antibodies label the cell soma of ~80% of submucosal ganglion cells and ~10% of myenteric ganglion cells in the human colon. 8 Immunoreactivity is also present in the neurites of these cells, including calretinin-immunoreactive mucosal innervation, which is absent in aganglionic bowel. 9 Calretinin immunohistochemistry has become a routine ancillary method in the histopathological diagnosis of Hirschsprung disease. 10 Expression of calretinin in a subset of enteric neurons is a conserved feature in other animals (e.g., mice, rats, sheep, and guinea pig), 11 but its function is unknown and mice that lack calretinin have no obvious gastrointestinal phenotype. 12 To date, calretinin expression has not been reported in skeletal, cardiac, or smooth muscle cells.
Methods
Specimens
A computerized search of surgical and autopsy reports was conducted to identify archived formalin-fixed paraffin-embedded (FFPE) sections of cecum and other gastrointestinal sites. Specimens were selected to represent a wide range of developmental stages (22 weeks gestation–18 years) and varied clinical settings (Table 1). Hematoxylin-and-eosin (H&E)-stained sections were reviewed to exclude tissues with moderate or severe histological autolysis. In some cases, frozen tissue sections of formalin-fixed samples were used for some of the immunofluorescence studies, as indicated in Table 1. Non-human tissue blocks were provided by investigators at Seattle Children’s Research Institute (mouse) or the University of Washington (macaque and rabbit). The rabbit blocks sampled the proximal, mid, and distal parts of their distal ileum, elongate cecum and non-vermiform appendices; mouse and macaque blocks included full-thickness samples of their distal ileum and cecum. Efforts to obtain cecal tissue blocks from non-human primates with true vermiform appendices (e.g., chimpanzees) were unsuccessful, in part because of restrictions on experimental research with these species.
Summary of Specimens and Calretinin Immunolabeling Results.
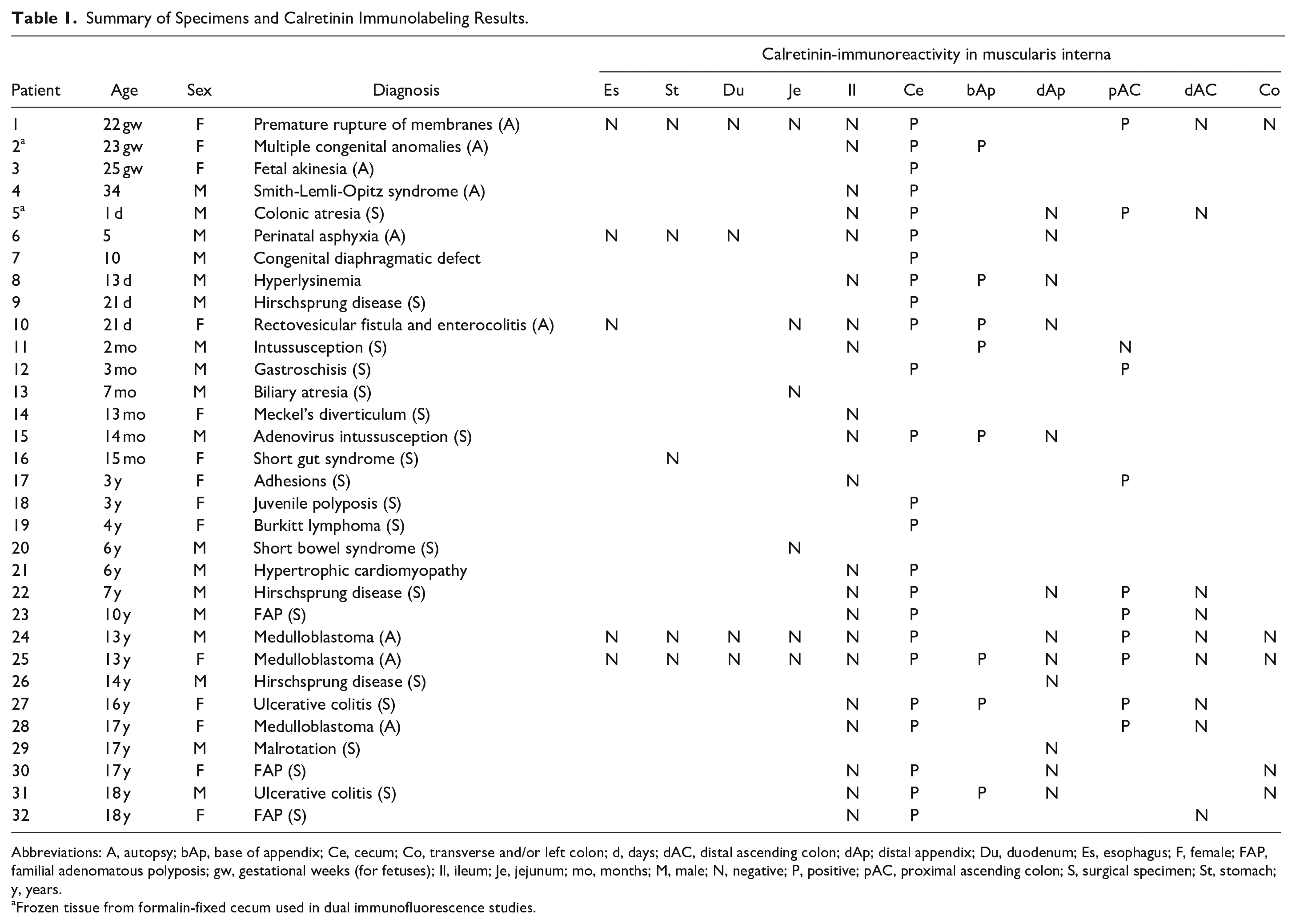
Abbreviations: A, autopsy; bAp, base of appendix; Ce, cecum; Co, transverse and/or left colon; d, days; dAC, distal ascending colon; dAp; distal appendix; Du, duodenum; Es, esophagus; F, female; FAP, familial adenomatous polyposis; gw, gestational weeks (for fetuses); Il, ileum; Je, jejunum; mo, months; M, male; N, negative; P, positive; pAC, proximal ascending colon; S, surgical specimen; St, stomach; y, years.
Frozen tissue from formalin-fixed cecum used in dual immunofluorescence studies.
Immunohistochemistry
Calretinin immunoperoxidase labeling was performed on 4 µm-thick FFPE sections with an automated immun-ostainer (Ventana BenchMark Ultra; Ventana Medical Systems, Tuscon, AZ). Rabbit anti-calretinin (SP65; catalog #790-4467; 1:2500; Zymed Laboratories, South San Francisco, CA) was used with “mild CC1-buffer” cell conditioning (a proprietary Tris-based buffer with a slightly basic pH; reference number 950-224, Ventana Medical Systems, Tuscon, AZ), 36 and 32 minutes incubation at 37°C and an A/B block. Sections were counterstained with hematoxylin.
Immunofluorescence studies were performed on frozen sections prepared from formalin-fixed tissue. For frozen sections, the fixed tissue was equilibrated 4–16 hours in 30% sucrose, embedded in OCT, and cut at a thickness of 8 µm. Antigen retrieval was performed in 10 mM citrate buffer, pH 6.0 for 10 minutes and then sections were incubated in primary antibodies in phosphate-buffered saline, pH 7.4 (PBS) + 0.3% Triton-X + 5% donkey serum (PBST) for 2 hours. One of 3 combinations of primary antibodies were used: rabbit anti-calretinin (SP13, catalog #232R; Cell Marque, Rocklin, CA; 1:100)/mouse anti-smooth muscle actin (catalog #MO85101, Dako, Santa Clara, CA; 1:100), goat anti-calretinin (catalog #AF5065; R&D Systems, Minneapolis, MN; 2 µg/ml)/mouse anti-smooth muscle actin (catalog #MO85101, Dako, Santa Clara, CA; 1:100), or mouse anti-calretinin (catalog #ab204990, Abcam, Waltham, MA; 1:100)/rabbit anti-cKIT (catalog #A4502, Dako, Santa Clara, CA; 1:100). Sections were washed with PBS and incubated in combination of 2 secondary antibodies: CY3-conjugated goat anti-rabbit (catalog #111-165-103, RRID AB_2340414; Jackson Labs, Bar Harbor, ME; 1:500) or 549-conjugated donkey anti-goat (catalog #750-506-147, RRID AB_2632576; Jackson Labs, Bar Harbor, ME; 1:500) and 488-conjugated donkey anti-mouse(catalog #715-545-150, RRID AB_2341099; Jackson Labs, Bar Harbor, ME; 1:500) in PBST for 1 hour. Finally, 4′,6-diamidino-2-phenylindole (DAPI; 5 µl per 50 ml PBS) was applied for 5 minutes, the sections were washed twice in PBS, and coverslips were applied with Fluoroscope mounting medi-um (SouthernBiotech, Birmingham, AL). Negative controls were performed in parallel by replacing the primary antibodies in the above protocol with PBST alone.
For calretinin/DOG1 dual immunohistochemistry, 5 µm FFPE sections were deparaffinized and conditioned with the Ventana Benchmark II automated immunostainer (CC1 × 64 minutes). After conditioning, the slides were removed from the instrument and processed by hand. After a brief rinse with PBS, slides were incubated overnight at 4°C in a combination of goat-anti-calretinin (catalog #AF5065; R&D Systems, Minneapolis, MN; 2 µg/ml) and rabbit-anti-DOG1 (catalog #SP31, RRID AB_1516844; Cell Marque, Rocklin, CA; 1:100) in PBST. Following three 5-minute washes with PBS, alkaline phosphatase-conjugated donkey anti-rabbit secondary antibody (catalog #711-055-152, RRID AB_2340591; Jackson Labs, Bar Harbor, ME; 1:500 × 30 minutes at room temperature) was applied, the slides were washed again and incubated with BCIP/NBT alkaline phosphatase substrate (catalog #SK-5400, Vector Laboratories, Newark, CA) until optimal staining was visualized. Then, sections were washed with PBS and incubated with peroxidase-conjugated horse anti-goat secondary antibody (catalog #PI-9500-1, RRID AB_2336526; Vector Laboratories, Newark, CA; 1:500 × 30 minutes at room temperature) in PBST, washed with PBS, and treated with TMB-DAB substrate (SK-4400, Vector Laboratories, Newark, CA) until reaction product was easily visible; the reaction product turned aqua after dehydration and application of Micromount coverslip medium (Leica Biosystems, Deer Park, IL).
RNA In Situ Hybridization
RNA in situ hybridization was performed on 5 µm FFPE sections using a human CALB2 probe (catalog #422171) and the RNAscope 2.5 HD Detection Kit - RED (catalog #322350) from Advanced Cell Diagnostics (Newark, CA) according to the manufacturer’s instructions. Final development reagents were applied 30–60 minutes until optimal labeling was obtained.
Results
Anti-Calretinin Immunoreactivity in Cecal Smooth Muscle
Calretinin immunohistochemistry was performed on paraffin sections of human gastrointestinal tract at various stages of development, which range between 22 weeks gestation and 18 years (Table 1). In every intestinal tissue section, intense immunoreactivity was observed in the cell soma of most submucosal neurons and a small minority of myenteric neurons, as well as neurites in the periganglionic plexuses, muscularis propria, and mucosa (Figures 1 and 2). Cytoplasmic immunoreactivity was also present in mast cells and mesothelium. No muscular immunoreactivity was observed in esophagus, stomach, or small or large intestine, except in the cecum, appendiceal base, and most proximal ascending colon.
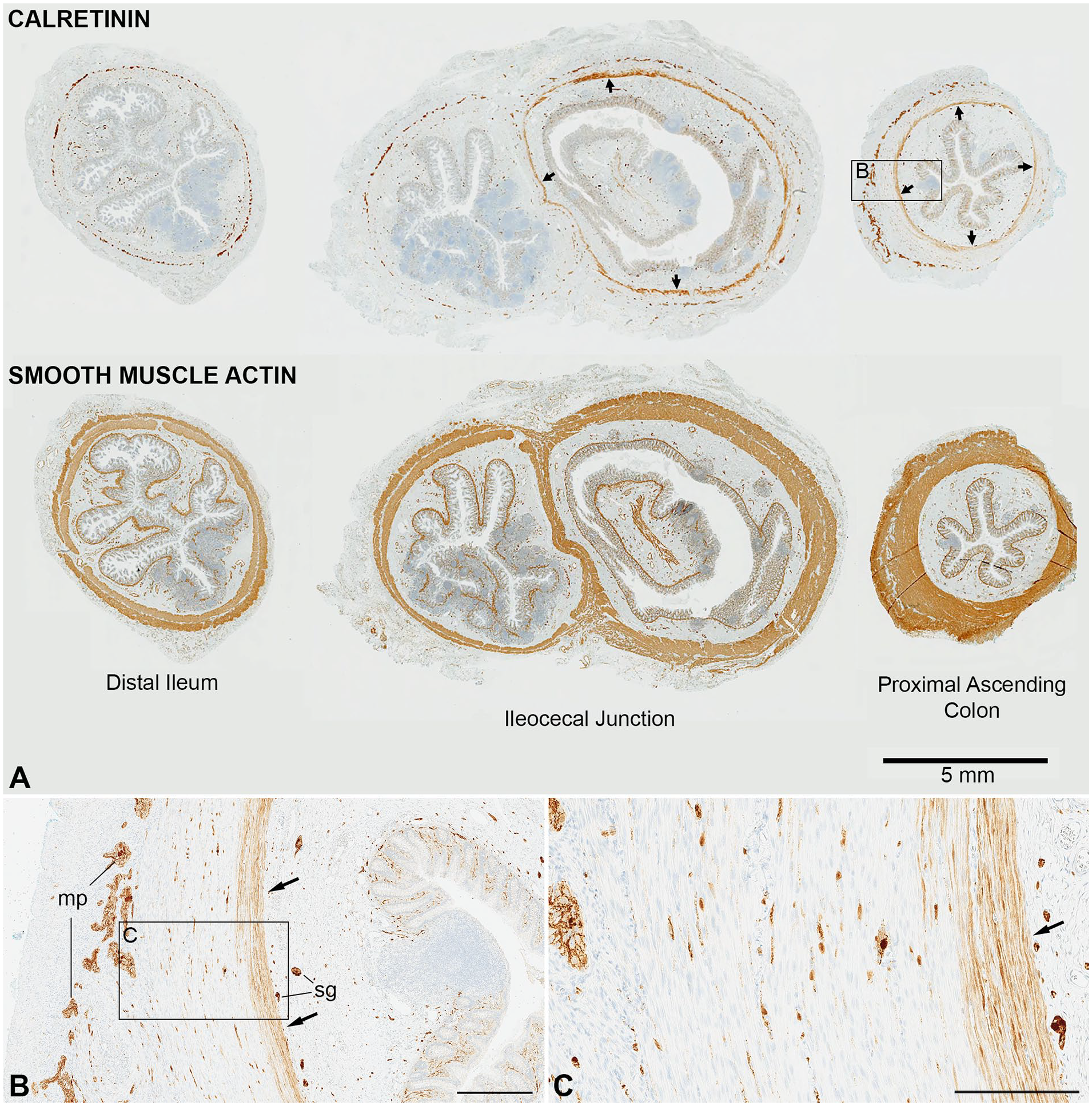
Calretinin and smooth muscle actin immunolabeling in the distal ileum, ileocecal junction, and proximal ascending colon. (A) Transverse sections of an ileocecal resection (3-month-old male, patient 13) show punctate calretinin immunolabeling of myenteric and submucosal ganglion cells and nerves in all areas. In the cecum and adjacent proximal ascending colon, but not the ileum, labeling in the inner part of the muscularis interna (arrows) is also present. Smooth muscle actin immunoreactivity highlights both layers of the muscularis propria with an intervening gap that corresponds to the myenteric plexus. (B) Magnified field from calretinin-labeled proximal ascending colon (rectangle in A) showing immunoreactive smooth muscle cells along the submucosal border (arrows) of the muscularis interna. (C) Magnified field from calretinin-labeled proximal ascending colon (rectangle in B). Abbreviations: mp, myenteric plexus; sg, submucosal ganglia. Scale bars: B, 400 µm; C, 200 µm.
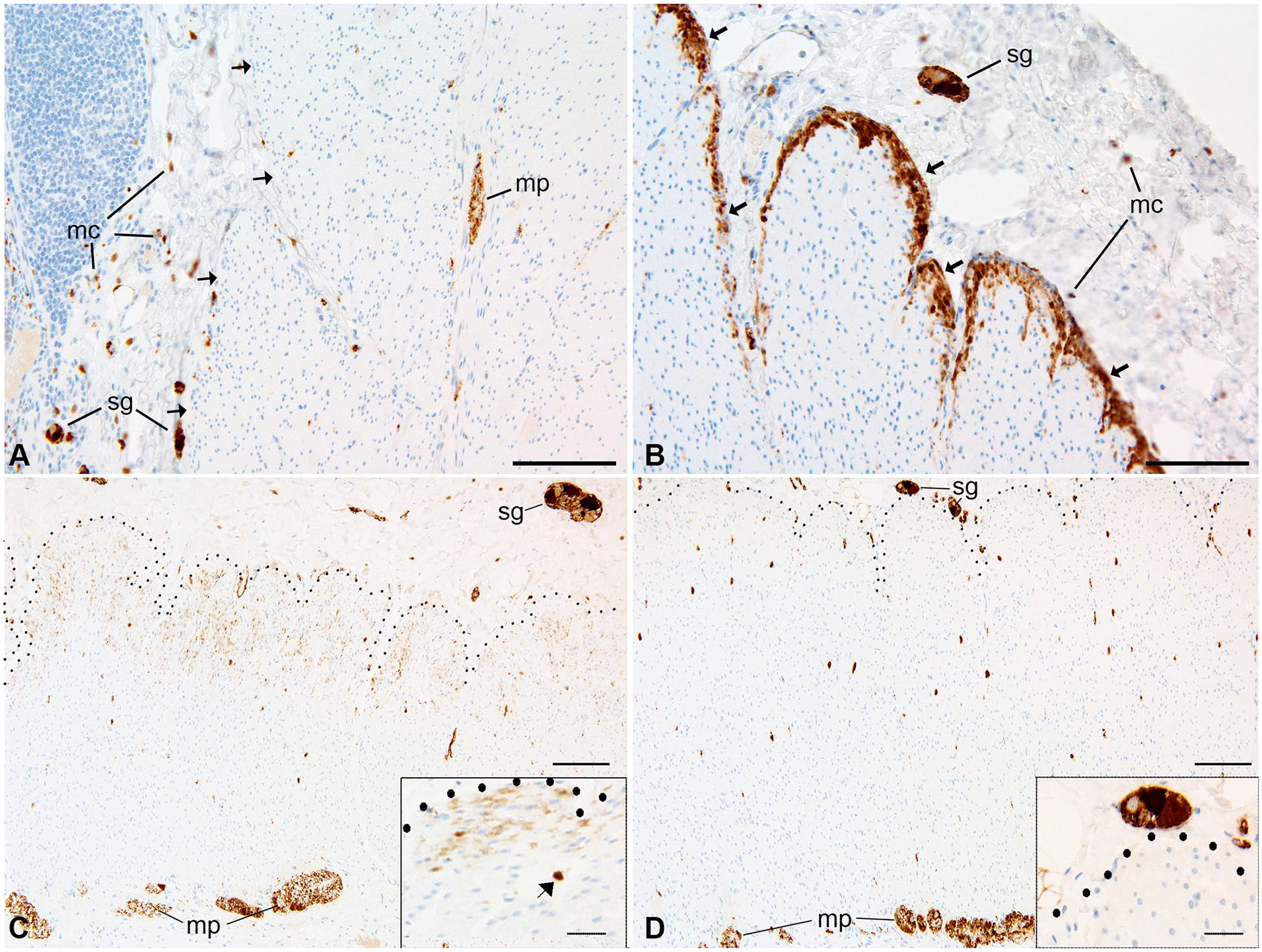
Restricted smooth muscle expression of calretinin in the cecal region. (A) In the small intestine (3-year-old, patient 18), calretinin labels submucosal ganglion cells (sg), nerves in the myenteric plexus (mp), and mast cells (mc) with negative staining in the muscularis propria. (B) In contrast, the innermost layers of the muscularis propria are intensely immunoreactive in the cecum of the same patient; submucosal ganglia (sg) and mast cells (mc) are also labeled. Arrows in (A) and (B) indicated submucosal border of muscularis propria. (C) Calretinin immunoreactivity (10-year-old, patient 21) is present in the innermost muscularis interna of the cecum (dotted lines show interface between muscularis and submucosa). The inset illustrates immunoreactive population at higher magnification of cecum (arrow indicates a mast cell). (D) This section from the mid-ascending colon of the same specimen demonstrates absence of muscular calretinin immunoreactivity, characteristic of the colon apart from the appendiceal-cecal region. Abbreviations: sg, submucosal ganglion; mp, myenteric plexus. Scale bars: A-D, 100 µm; insets, 25 µm.
In the cecum, appendix, and ascending colon, antibody labeling was observed in the cytoplasm of smooth muscle cells, which comprised the innermost portion of the muscularis interna (circular portion of the muscularis propria) (Figure 1). The intensity of immunolabeling was generally less than in neurons in the same tissue section, but unequivocally above background and sharply in contrast to immediately adjacent non-reactive smooth muscle cells. The immunoreactive myocytes were restricted to the inner third of the muscularis interna, sparing the muscular tunic closest to the myenteric plexus; no immunostaining was ever detected the muscularis externa (longitudinal muscle layer). In the cecum, between the appendiceal base and the ileocecal valve, calretinin immunoreactive (CR+) smooth muscle cells formed a confluent population that circumscribed the inner muscularis interna. This “shell” of CR+ smooth muscle cells ended abruptly at the interface between small and large intestine along the ileocecal valve (Figure 1(A)). The CR+ population extended into the base of the appendix and proximal ascending colon, where it became discontinuous and interrupted by patches of unlabeled cells. Distal to first-third of the ascending colon all smooth muscle immunoreactivity disappeared (Figure 2(B)), with a transition between patchy positivity to complete negativity never farther that one-third of the way between the ileocecal valve and hepatic flexure. The extent of appendiceal immunoreactivity was limited in most cases to the very base with no smooth muscle labeling in the mid or distal appendix; occasionally smooth muscle calretinin immunoreactivity extended from the base into the proximal third of the appendix. The same distribution with respect to these anatomical landmarks was maintained from mid-gestation (earliest stage examined) until adulthood.
Calretinin RNA In Situ Hybridization
RNA in situ hybridization was performed as an alternative means to confirm that calretinin immunoreactivity in the muscularis propria correlates specifically with sites of CALB2 (abbreviation for the calretinin gene) expression. As expected, robust hybridization was identified in a subset of submucosal and myenteric (not shown) neurons, in which labeling was confined to the perikaryon, where RNA transcripts normally reside (Figure 3). Labeling in smooth muscle was restricted to the innermost layers of the muscularis propria in the cecum, but not more distal colon, concordant with the immunohistochemistry results. The density of hybridization signals in the inner muscularis interna of the cecum, while unequivocally positive in comparison with smooth muscle elsewhere, was significantly less than in the cell bodies of ganglion cells, suggesting that calretinin transcripts are less abundant in the smooth muscle cells. This finding correlates with weaker immunolabeling in the smooth muscle cell relative to ganglion cell soma, although the difference observed with RNA hybridization was more dramatic.

CALB2 (calretinin) RNA in situ hybridization. (A and B) Red punctate labeling is observed in the inner muscularis interna of the cecum (patients 5 and 24). The density of labeling is much less than in neighboring submucosal ganglion cell bodies (arrow). (C) In contrast to a submucosal ganglion cell (arrow), no hybridization signal is detected in the muscularis interna from the right colon (same patient as A). Scale bars: A-C, 50 µm; insets, 10 µm.
Dual Immunolabeling
Dual-label immunofluorescence or immunohistochemistry was performed on tissue sections for a subset of cases. Anti-smooth muscle actin highlighted all smooth muscle cells, including both layers of the muscularis propria. In the inner muscularis interna of the cecum, calretinin colocalized with smooth muscle actin, which confirmed the CR+ cells are myocytes (Figure 4(A) and (B)). Identical results were obtained with 2 different calretinin-specific antibodies (see Methods) and no labeling was observed in control sections when primary antibody was excluded from the protocol (Figure 4(C)). Anti-cKIT (CD117) and anti-DOG1 labeled interstitial cells of Cajal (ICC). ICC were located around the myenteric plexus and more sparsely within and along the submucosal border of the muscularis interna. CR+ smooth muscle cells closely approximated the latter ICC population but did not colocalize with cKIT or DOG1 in ICC (Figure 5(A)-(C)). Calretinin and cKIT colocalization was observed in mast cells, as expected (not shown).
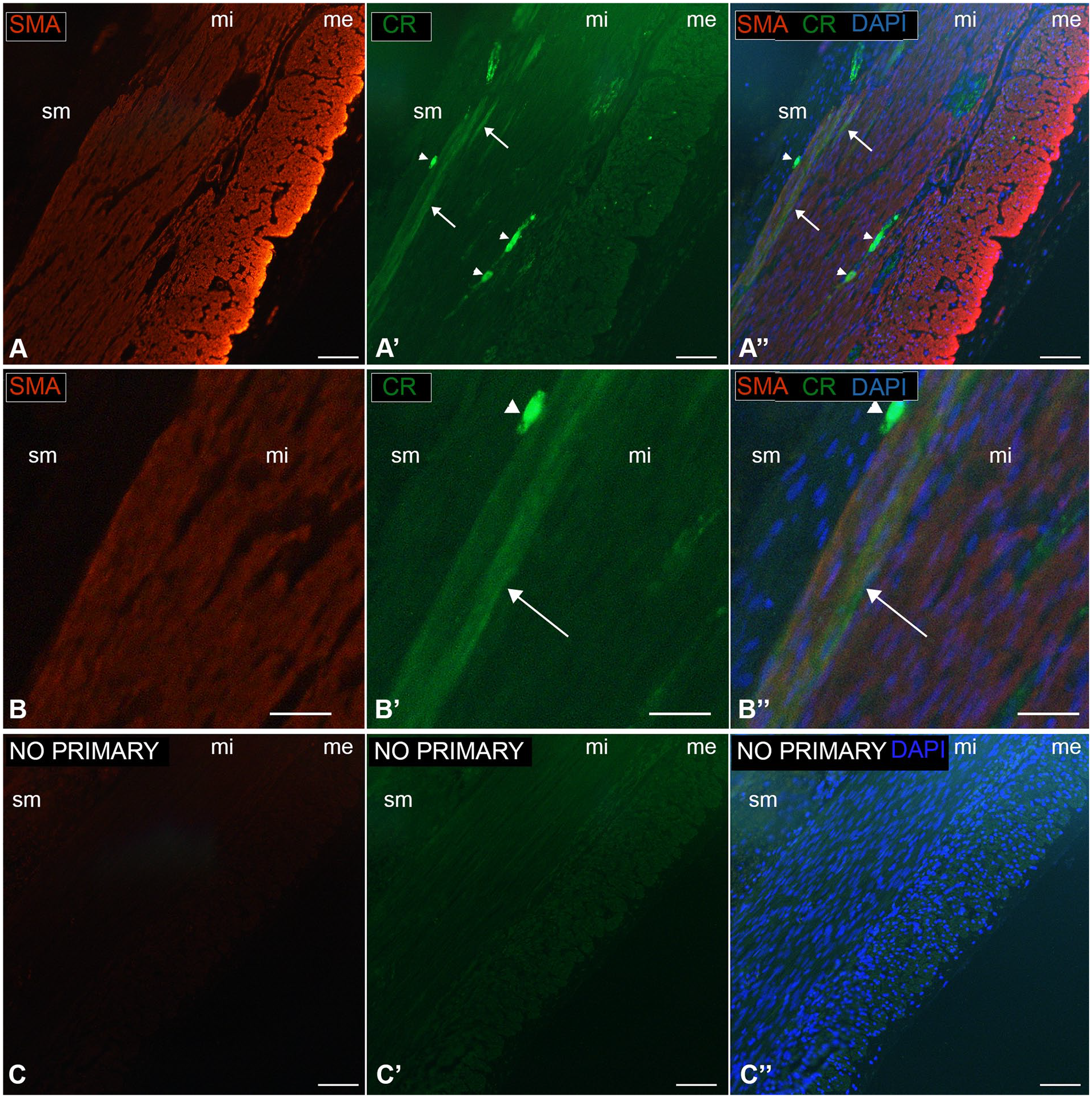
Mouse anti-smooth muscle actin and goat anti-calretinin dual immunolabeling (cecum, 10-day-old, patient 7). (A and B) Dual-immunofluorescence labeling shows that SMA is distributed throughout the muscularis interna (mi) and externa (me). (A′ B′) In the muscularis propria, CR immunoreactivity is only in the inner layers of the muscularis interna (arrows); arrowheads, immunoreactive submucosal and myenteric ganglia. (A″ B″) Overlay of SMA and CR with DAPI-stained nuclei (blue); arrows highlight areas of SMA and CR colocalization in smooth muscle cells. (C, C′, C″) Negative control images from sections of the same tissue block run in parallel with A, but without addition of primary antibody. sm, submucosa. Scale bars: 100 µm.
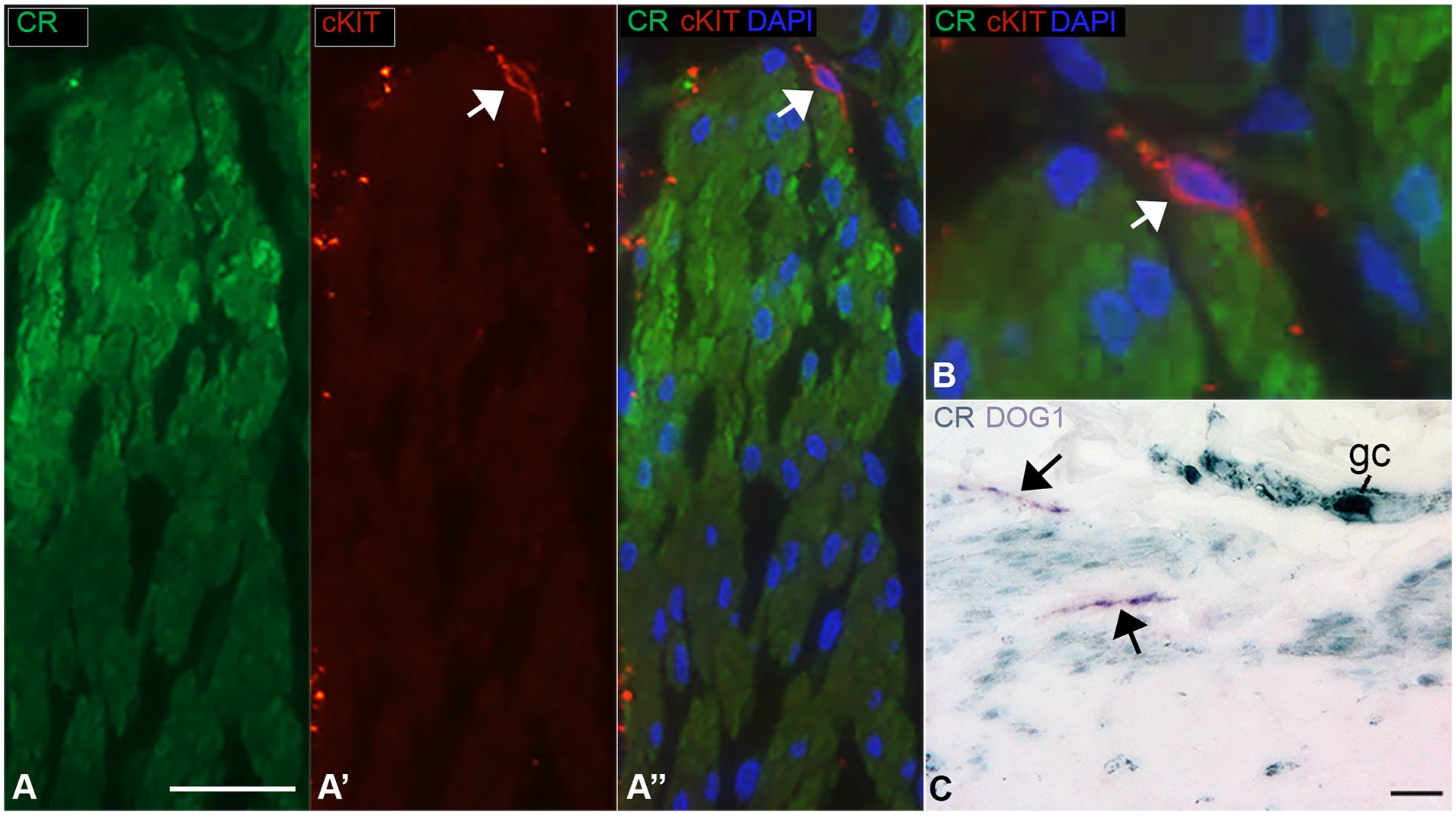
cKIT and calretinin dual immunolabeling (cecum, 1-day-old, patient 5) (A) Dual-immunofluorescence labeling shows CR expression in smooth muscle cells of the muscularis interna (mi) and an adjacent cKIT-positive, CR-negative interstitial cell of Cajal (arrow). (B) High magnification overlay of CR and cKIT labeling show that the 2 proteins do not colocalize in an interstitial cell of Cajal (arrow), the processes of which appear to contact CR-positive smooth muscle cells. (C) CR-peroxidase (aqua) and DOG1-alkaline phosphatase (purple) immunolabeling of smooth muscle cells and interstitial cells of Cajal (arrows), respectively. Scale bars: 25 µm.
Two different calretinin-specific antibodies were used for dual-labeling, and each yielded an identical pattern of smooth muscle positivity to the paraffin-based immunohistochemistry described above. In each case, negative control sections (no primary antibody) showed no significant background (not shown). The combination of identical results with different antibodies and appropriately negative controls suggests that the immunohistochemistry results are specific for calretinin and reflect previously unrecognized expression of the intracellular calcium buffering protein by a localized subpopulation of smooth muscle cells.
Calretinin Immunoreactivity in Other Species
Cecal appendices are thought to have evolved independently at least 29 times throughout mammalian evolution. 13 Unlike rabbits, humans and primates closely related to humans, neither mice nor macaques have appendi-ces. Calretinin immunohistochemistry was performed on sections of mouse (n = 3), rabbit (n = 2), and macaque (n = 3) ceca (Figure 6). In the ceca of each of these species, intense labeling of a subset of enteric ganglion cells and nerves served as an internal control and indicated the antibody recognized their calretinin protein. In none of the animal ceca nor the rabbit appendix was calretinin immunolabeling detected in smooth muscle cells.
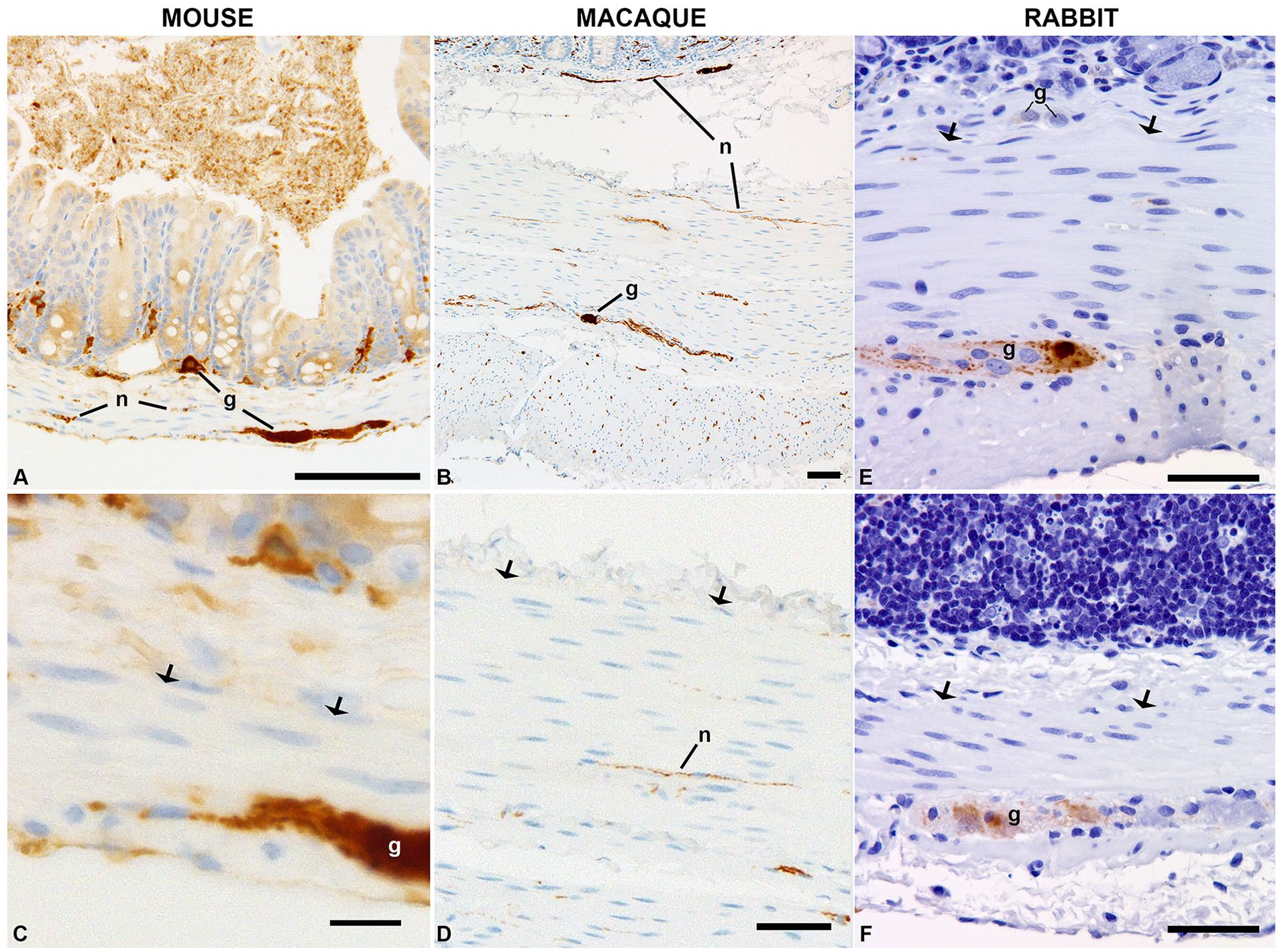
Calretinin immunolabeling of ceca from other species. Immunoreactivity was present in ganglion cells (g) and nerves (n) in the ceca of mice (A and C) and macaques (B and D) and in the rabbit small intestine (E) and cecum (F), but no smooth muscle labeling was identified in any of these species. Arrows indicate the interface between submucosa and muscularis propria, where calretinin-positive smooth muscle is observed in human ceca. Scale bars: A and B, 100 µm; C, 10 µm; D, 70 µm; E and F, 50 µm.
Discussion
Calretinin Expression in Human Cecal Smooth Muscle
Calretinin immunohistochemistry is performed routinely as part of the evaluation of rectal biopsies or other colonic specimens for possible Hirschsprung disease. 10 In this clinical context, attention is focused on expression by a subset of intrinsic enteric ganglion cells and their mucosal nerve processes. However, in this study, calretinin immunoreactivity and mRNA expression are described in a subset of enteric smooth muscle cells, which are confined to the inner layers of the muscularis propria in the cecum, base of the appendix, and proximal ascending colon. This pattern of expression is consistently observed in human colons, but not in the ceca of mice or macaques, species which lack appendices, or the appendices of rabbits. The findings allow for speculation about what purpose calretinin expression has in this particular population of smooth muscle cells. In order to develop a hypothesis, it is necessary to first review in detail some aspects of cecal/appendiceal comparative anatomy and colonic contractile physiology, as well as the established importance of calretinin as a calcium buffer in neurons.
Distinct Morphological Features of Inner Portion of the Muscularis Interna in the Human Cecum
The anatomy of the mammalian colon exhibits considerable species variation related in part to dietary differences. 14 Distinctive features of the human colon include a proportionately small cecum, haustra, consolidation of the muscularis externa into 3 taenia coli (longitudinal bands that extend from the cecum to the sigmoid colon), and a cecal appendix. Anatomical variations of the cecum are particularly notable. While most mammals have recognizable ceca, only a minority have cecal appendices. The presence or absence of a cecal appendix in a given species depends in part on how the structure is defined. Smith et al. 13 and Smith 16 have studied appendiceal morphotype and their evolution.13,15 They define the cecum as “any cecal appendage that has a distinct decrease in diameter compared to the main body of the cecum.” This definition excludes mere thickening of the wall at the cecal apex and/or concentration of lymphoid tissue in the cecum. Smith et al describe 2 morphotypes of cecal appendices: (a) vermiform—an elongate, cylindrical, worm-like morphology found in humans, and a few other primates; (b) tapering—a voluminous cecum that “converges distally into an appendix, as exemplified by rabbits and other lagomorphs. She also classifies short narrow projections from the intestines in the absence of a well-formed cecum as “appendix-like structures” which are found in wombats and other marsupials. Rigorous analysis based on these definitions suggests that some variation of a cecal appendix has evolved independently at least 29 times and been lost as many as 12 times. Among extant animals, cecal vermiform appendices are particularly prevalent in primates (10 of 13 taxa), in which cecal and appendiceal anatomy is very similar among hominoids (humans and apes), but appendices are lacking in most New and Old World monkeys.13,15,17,18 Spontaneous appendicitis is relatively common in humans and has been documented in a chimpanzee 19 and rabbits 20 and suspected in gorillas,19,21 but has not been reported in other appendix-bearing species. 16 Despite the frequency of appendicitis in humans, the pathogenesis is poorly understood. Fecal stasis within the appendiceal lumen is a proposed risk factor. 22
The microscopic anatomy of the human cecum and ascending colon differ from other parts of the colon in a manner that may correlate with some of the physiological properties describe below. Arguably, the most detailed anatomic studies of this subject have been reported by Faussone-Pellegrini et al.,23 -25 who conducted light and microscopic analyses on human colons. Some, but not all, of their observations were confirmed by others. 26 It is generally agreed that the innermost 2–15 rows of myocytes along the submucosal border of the muscularis interna (inner circular muscle cells, ICMC) have different staining and ultrastructural features from the outer circular muscle cells, OCMC). These myocytes have smaller diameters, more perinuclear organelles, and higher glycogen content than the rest of the muscularis interna. ICC-SM and nerves run along the submucosal surface of the ICMC, where slow waves originate. 27 Faussone-Pellegrini emphasized that these cytological features were confined to ICMC in the cecum and ascending colon and gradually transitioned along the transverse colon to a “meshlike” arrangement, which exists in the left colon. In contrast, Rumessen et al 26 explicitly stated that the cytological features of the ICMC were similar in all parts of the colon and the OCMC and ICMC were closely apposed throughout. 26 Irrespective of their differences, these studies concur that the ICMC differs from the OCMC and that the distinctive features of the ICMC may relate to their special role in the initiation and propagation of colonic contractility.
Inner Circular Muscle Cells and Colonic Contractile Activity
Contractile events in the colon and other parts of the gastrointestinal tract are regulated by intrinsic myoelectric properties of the muscularis propria and superimposed modulation of these properties by the enteric nervous system. Fundamental to normal motility are slow waves—rhythmic changes in smooth muscle cell membrane potential, which intermittently depolarize the resting membrane potential of myocytes and bring these cells close to their threshold for contraction.28,29 Slow waves or pacemaker potentials are initiated in interstitial cells of Cajal (ICC) and propagate to adjacent smooth muscle cells via gap junctions. In conventional hematoxylin-and-eosin-stained paraffin sections, ICC are inconspicuous. They were originally visualized with vital dyes (e.g. methylene blue and zinc iodide/osmic acid) or in Golgi stained preparations, and later characterized by electron microscopy (reviewed by Thuneberg 30 ). Today, ICC are most often resolved with immunohistochemical markers including c-Kit (CD117) and DOG1 (ANO1). 31 ICC are stellate cells with relatively small cell bodies, round nuclei, and multiple thin long processes. 30 They reside along boundaries between the smooth muscle and connective tissue, where enteric nerves are found that modify their activity. In the colon, subpopulations of ICC are located along the submucosal surface of the muscularis interna (ICC-SM), the myenteric surfaces of the muscularis interna and muscularis externa (ICC-MY), sub-serosal surface of the muscularis externa (ICC-SS), and between muscle fascicles in the muscularis propria (ICC-IM). A second class of “fibroblast-like” interstitial cells is also present in many of the same locations as ICC, but express PDGFRα, as opposed to c-Kit.
Sanders et al. 32 introduced the Smooth muscle/ICC/PDGFRα cell (SIP) syncytium as a way to conceptualize the intimate spatial and functional relationships that exist between these cells muscularis propria. Slow waves and other electrical properties of cells in the SIP directly influence each other because they are connected by gap junctions. Therefore, neural, pharmacologic, toxic, or pathological alterations that affect any of the cell types can disrupt function of the entire network.
In the colon, slow wave pacemaker activity initiates at the submucosal border of the muscularis propria in the ICC-SM.33,34 Rhythmic depolarization of ICC-SM cells is transmitted rapidly to overlying smooth muscle cells and somewhat more slowly through the ICC network to upstream or downstream smooth muscle. Initiation and propagation of slow waves is dependent on calcium fluxes both in ICC and adjacent ICMC. As slow wave depolarization passes from ICC to neighboring ICMC, the membrane potential changes in the latter parallel those in the ICC, but are mediated by different channels, including voltage-sensitive T- and L-type Ca++ channels. 35 The net result is coordinated rhythmic changes in both membrane potential and intracellular Ca++, which are maximal in the innermost muscularis interna and diminish toward the outer muscularis interna.
Superimposed on the slow wave are membrane potential oscillations (MPOs). MPO’s begin in ICC-MY (and possibly ICC-IM), lack the regularity of slow waves, and propagate through the muscularis propria from the external surface toward the inner border. When a cresting slow wave and MPO collide in smooth muscle cells, phase summation takes place and the threshold for Ca++-mediated action potentials may be reached. 36 Neural inputs that lower or raise the action potential threshold will influence whether or not contraction is realized and neural networks allow for coordinated contraction and relaxation necessary for propulsive colonic motility. 37 Repolarization of smooth muscle cells in the SIP is mediated in large part by calcium-dependent potassium channels (BK and SK channels).35,38 Potassium conductance through these channels works to restore the resting membrane potential and relax the muscle cell.
Ultimately whether or not smooth muscle contraction takes place depends on the intracellular concentration of calcium in myocytes, which is intimately linked to membrane potential and production and frequency of action potentials. 39 Calcium entry during these electrical events initiates a chain of molecular phosphorylation events that culminates in contractile interaction between actin and myosin II. In brief, the canonical sequence of molecular events is a follows: (a) calcium binds to the calcium-sensor protein, calmodulin, and causes a conformational change to enable calmodulin to interact with other proteins, (b) interaction with myosin light chain kinase activates the kinase, (c) myosin light chain kinase phosphorylates the regulatory light chain of myosin II, (d) phosphorylated myosin II establishes cyclic force bridges with actin, and (e) contraction ensues. Physiological, pharmacological, or patholo-gical processes that reduce intracellular calcium inhibit contraction. Thus, intracellular calcium concentration plays an important role in the formation and propagation of electrical slow waves in the SIP and excitation-contraction coupling in smooth muscle cells.
Non-Propulsive and Propulsive Colonic Motor Patterns
Colonic motor patterns can be subclassified into non-propulsive and propulsive types. 40 Non-propulsive or segmental contractions are thought to be important for mixing of luminal contents, in contrast with propulsive contractions, which cause mass movements of materials through the length of the colon. In the human colon, manometric studies have demonstrated that most propulsive movements of luminal contents correlate with pressure waves that originate at or proximal to the mid-ascending colon and extend up to or through the mid-transverse colon.41,42 We lack detailed understanding of the cellular and electrophysiological events that underlie these colonic motor patterns. However, the combination of data collected from animal models and more limited studies of human tissue suggest that ICC-SM and contractile “ripples” produced by calcium-mediated activity in adjacent smooth muscle cells play an important role in the initiation or propagation of some of these contractile phenomena. Therefore, a factor like calretinin, which buffers against calcium fluxes in ICMC, is likely to inhibit the initiation and/or propagation of some types of non-propulsive and non-propulsive colonic motor patterns.
Calretinin’s Effect on Cellular Excitability
Expression of calretinin in smooth muscle cells has not been previously described. It seems logical to suggest that the calcium-buffering effect of calretinin might reduce ICMC excitability, but no direct experimental data exists regarding the physiological significance of calretinin in these cells. In contrast, a lot is known about the functions of calretinin and other calcium-buffering proteins in neurons, which may provide some useful insights. Experiments with calretinin-null (−/−) mice suggest that calretinin serves as a calcium buffer in cerebellar granule cells to modulate the electrophysiological properties of these cells.2,43 In the cerebellar cortex, excitatory input from mossy fibers stimulates granule cell neurons, which in turn excite Purkinje cells. Cerebellar granule cells each have 4 dendrites and each dendrite receives mossy fiber input from a single mossy fiber. 44 At these synapses, mossy fibers release glutamate, which activates AMPA and NMDA receptors. NMDA receptors allow for calcium influx, as do other types of calcium channels (e.g., L-type) expressed by granule cells. Mitochondria and endoplasmic reticulum may also be a source of calcium in response to granule cell stimulation. The net effect of excitatory glutaminergic stimulation is depolarization of the granule cell and an action potential. Restoration of the normal resting membrane potential is mediated largely by Ca++-activated potassium channels (BKCa channels), which are sensitive to intracellular calcium levels. 45 In calretinin −/− animals, the excitatory properties of granule layer neurons appear to be enhanced so that the neurons are more readily excited by presynaptic input, as expected if intracellular Ca++ levels are increased. Electrophysiologically, this has been observed in vitro by direct recordings from granule cell neurons 46 and indirectly in vivo by recording the granule cell-mediated activity of Purkinje cells.47,48 In alert mice, calretinin −/− mice have more “single spikes,” shorter “complex spikes,” and a prolonged delay of the single-spike pause after complex spikes. 47 The alterations in granule cell neuron excitability are rescued by intracellular injection of BAPTA (fast-acting Ca++ chelator), which supports the concept that they are due to the Ca++ buffering function of calretinin. 49
Conceptually, these observations in the cerebellum may be relevant to excitability of cecal smooth muscle cells. Like cerebellar granule cells, myocytes in the colonic muscularis interna receive excitatory (depolarizing) input from ICC-SM and ICC-MY, as well as excitatory and inhibitory input from neurons. The summative effect on myocyte membrane potential and ultimately intracellular Ca++ concentration will dictate whether contraction occurs. If calretinin in ICMC functions in a similar manner to cerebellar granule cells, it will reduce excitability and oppose the propagation of muscular contractions.
Why Is Calretinin Selectively Expressed in Human Cecal and Appendiceal ICMC?
I hypothesize that expression of calretinin to in the submucosal border of the muscularis propria in the cecum and proximal appendix evolved to reduce the likelihood that smooth muscle contractions will propagate toward the appendix. As described above, significant propulsive contractions are initiated in the proximal colon and propagate aborally. These contractions provide the force that moves content through the colon lumen toward the rectum. Were similar contractions to propagate in the opposite direction, luminal contents would move toward the base of the cecum and appendix, potentially increasing appendiceal stasis and the likelihood of appendicitis.
Contemporary models suggest that the initiation of colonic contraction depends on ICC-driven changes in the calcium levels in ICMC. By buffering the free calcium levels in the ICMC of the cecum and appendix, calretinin may increase the action potential threshold of these cells relative to their downstream ICMC neighbors and thereby promote the initiation and propagation of contractions away from the cecum (Figure 7). This hypothesis is admittedly speculative, but consistent with current understanding of colonic muscular physiology, the function of calretinin in some other cell types, and the restricted pattern of calretinin expression observed in human colon and appendix. The model allows for secondary hypotheses which could be tested by targeted expression of calretinin in smooth muscle cells in tissue culture and animal models. If calretinin expression in ICMC does play some type of protective role against appendiceal stasis, it is possible that genetic or environmental factors that alter the putative calcium buffering effect of calretinin are involved in some manner in the poorly understood and relatively frequent incidence of appendicitis in humans.
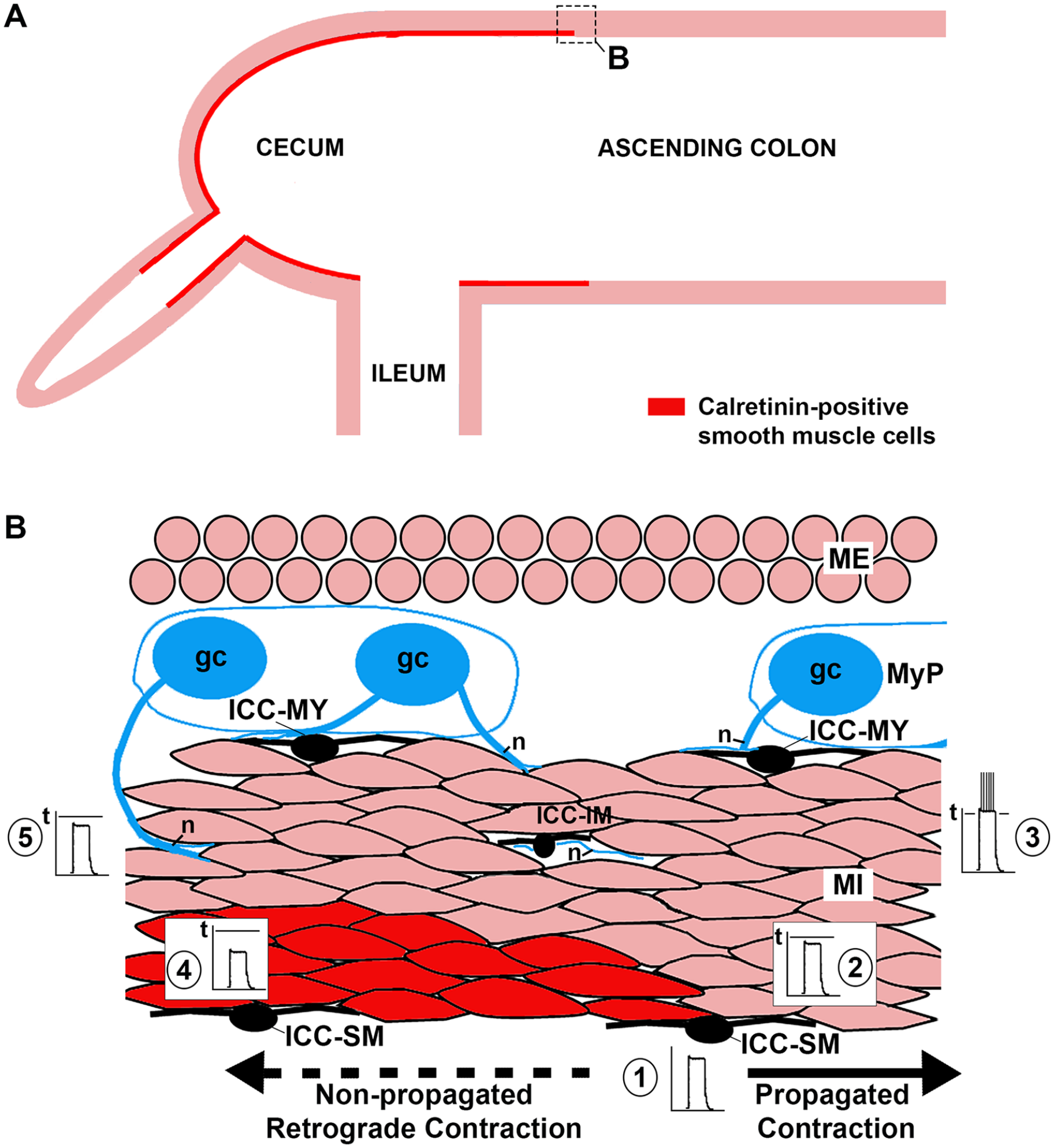
Hypothetical suppression of retrograde contractile propagation by calretinin calcium buffering in the cecal muscularis interna. (A) Observed distribution of calretinin-immunoreactive smooth muscle cells in the inner muscularis interna (rectangle corresponds with area illustrated in B). (B) Rhythmic slow wave potentials (1) are generated by interstitial cells of Cajal (ICC) at the submucosal border (ICC-SM) of the muscularis interna (MI) and passively conducted to adjacent smooth muscle cells. (2) In the aboral direction, the membrane potential of smooth muscle cells is brought close to the threshold (t) for action potentials and (3) driven past this potential by summation and neural excitation with consequent propagated propulsive contraction. (4) In the oral (retrograde) direction, the intracellular calcium-buffering effect of calretinin reduces the membrane potential of smooth muscle cells such that they are (5) less likely to reach the threshold for action potential formation and contraction.
Footnotes
Acknowledgements
The author thanks Dr. Kristina Adams-Waldorf and the University of Washington Department of Comparative Medicine for formalin-fixed intestinal tissue samples from non-human species.
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author received no financial support for the research, authorship, and/or publication of this article.